一种猪圆环病毒3型病毒样颗粒亚单位疫苗及其制备方法与应用
文献发布时间:2023-06-19 19:30:30
技术领域
本发明属于生物技术检测领域,具体涉及一种猪圆环病毒3型病毒样颗粒亚单位疫苗及其制备方法与应用。
背景技术
猪圆环病毒(PCV)是单股环状DNA病毒,基因长度约为1.7kb,是最小的动物DNA病毒之一,主要引起仔猪多系统衰竭综合征、肺炎、皮炎、肾病综合征、繁殖障碍,给生猪的养殖带来重大损失。PCV3是一种新型圆环病毒,国内尚无针对PCV3的商品疫苗。PCV3和PCV2Cap蛋白基因的同源性很低,仅有30%的一致性,这也暗示已有的PCV2疫苗无法保护生猪免受PCV3的侵害。因此PCV3相应疫苗的研发至关重要。
最近的研究表明PCV3 Cap蛋白可在多种系统中表达且获得亚单位或者嵌合表位疫苗,Cap蛋白可以自发组装形成模拟天然病毒形态的病毒样颗粒(VLPs),无病毒核酸,没有自主复制的能力,VLPs可以诱发宿主细胞产生免疫应答,因此,Cap VLPs可以作为研发猪圆环病毒的优选靶标。目前已有研究表明,PCV3 cap蛋白在大肠杆菌、昆虫杆状病毒系统、毕赤酵母中成功表达且能组装成病毒样颗粒,并且能获得较好的免疫效果。但是在大肠杆菌表达系统(CN108611359B)中获得的PCV3 Cap重组菌,主要形成包涵体,蛋白可溶性低,且有内毒素的污染,菌液需超声破碎收集上清,非常不利于下游生产,而后期去除内毒素工艺复杂且代价昂贵;大肠杆菌表达系统有着成本低,表达量大的优势。因此,大肠杆菌表达系统成为大批量生产的首选。但同时,大肠杆菌表达系统用于VLP制备也存在着许多技术难点。首先,因为密码子使用频率的不同,动物病毒的基因常常难于在大肠杆菌表达系统中表达。其次,大肠杆菌分子伴侣较少,常常造成重组蛋白无法正确的折叠,从而影响重组衣壳蛋白组装成VLP。另外,相对于真核表达系统,大肠杆菌缺乏很多关键的蛋白酶,无法去除阻碍衣壳蛋白组装的多余氨基酸序列。针对不同的动物病毒都需要摸索出特定的表达与包装条件,才能使用大肠杆菌表达系统制备出相应的VLP。
PCV3作为猪源病毒,其Cap蛋白在大肠杆菌中的表达量受着密码子使用频率偏好性的影响,Cap蛋白本身含有大量的疏水氨基酸,其在大量表达时容易形成不可溶的包涵体,从而无法形成VLP结构,另外,Cap蛋白自组装成VLP需要在特定的条件下进行,因此,本研究存在一定的难度。
SUMO是一种小分子泛素样修饰蛋白,广泛存在于各种真核细胞中,SUMO近年来被发现可用作分子伴侣来增加外源蛋白的稳定性和可溶性,其作用机理可能是SUMO蛋白作为一种高度疏水的核心为目的的蛋白的折叠提供成核位点,促进蛋白间的相互作用并使其正确折叠,最终增强融合蛋白的可溶性。
为使猪圆环病毒3型Cap蛋白能够在上清液中大量表达,王帅勇等人设计针对去核定位信号(34个氨基酸)的ORF2特异性引物,将截短后的PCV3 Cap序列连接到pCold TF载体上,构建了重组的原核表达质粒pCold TF-PCV3-dCap。
发明内容
为了提高大肠杆菌表达可溶性PCV3 Cap蛋白的表达量和稳定性,有利于工业化生产,本发明的技术方案是:
一种猪圆环病毒3型病毒样颗粒的制备方法,包括以下步骤:
S01.将pET28a(+)的T7启动子替换为如SEQ ID No.2所示的tac弱启动子的核苷酸序列,将如SEQ ID No.3所示GST的核苷酸序列插入空载质粒pET28a(+)的NcoI、BamHI之间,得到pETM载体;
S02.将如SEQ ID No.6所示的optPCV3 Cap基因连接至pETM载体,连接产物转化到E.coli DH5α感受态细胞,挑菌鉴定后抽提质粒,重组质粒命名为pETM-optPCV3 cap;
S03.将pETM-optPCV3 cap质粒转入大肠杆菌表达菌株BL21(DE3),命名为ETM-optPCV3 cap/BL21;
S04.ETM-optPCV3 cap/BL21菌液接种到含卡那霉素的液体LB培养基中,37℃,200rpm,培养至OD
S05.无菌1×PBS洗涤并重悬菌体,破碎重悬的菌体,离心收集上清液,过柱纯化,透析形成VLP,得到猪圆环病毒3型病毒样颗粒;
所述optPCV3 cap基因的核苷酸序列为SEQ ID No.6所示。
进一步的,所述步骤S01包括:J01.以如SEQ ID No.8所示的正向引物和如SEQ IDNo.9所示的反向引物为上下游引物,以空载质粒pET28a(+)为模板,采用PrimeSTAR MaxDNA Polymerase进行扩增,扩增产物加入DpnI消化模板,然后转化E.coli DH5α感受态细胞,涂布卡那霉素抗性的LB平板,37℃静置过夜,挑选单克隆转接至卡那霉素抗性的LB液体培养基,37℃振荡培养,挑取平板上的单菌落,PCR鉴定,抽提质粒,再进行NcoI和BamHI双酶切,并回收酶切产物;
J02.以正向引物:5’CATG
J03.将步骤J01的酶切产物和步骤J02的纯化产物在16℃连接过夜,之后将连接产物转化到E.coli DH5α感受态细胞,涂布含卡那霉素的LB平板,37℃培养12-16h,挑取平板上的单菌落,PCR鉴定,抽提质粒,重组质粒命名为pETM载体。
进一步的,所述步骤S02包括以下步骤:
(1)以f:5'CCG
(2)扩增产物经EcoRI和XhoI双酶切后回收纯化,pETM载体经EcoRI和XhoI双酶切后回收纯化,处理好的pETM载体和optPCV3 Cap在16℃连接过夜,之后将连接产物转化到E.coli DH5α感受态细胞,涂布含卡那霉素的LB平板,37℃培养12-16h,挑取平板上的单菌落,PCR鉴定,抽提质粒,重组质粒命名为pETM-optPCV3 cap。
本发明的另一个目的是提供上述方法制备的猪圆环病毒3型病毒样颗粒。
本发明的另一个目的是提供上述猪圆环病毒3型病毒样颗粒在制备猪圆环病毒3型抗体检测试剂中的应用。
本发明的另一个目的是提供上述猪圆环病毒3型病毒样颗粒制备的猪圆环病毒3型亚单位疫苗。
进一步的,所述疫苗包含按照权利要求4所述方法制备的病毒样颗粒和SummitPoly Solution佐剂。
进一步的,所述疫苗中病毒样颗粒和Summit Poly Solution佐剂的体积比为4:1,病毒样颗粒的浓度为120μg/ml。
与现有技术相比,本发明具有以下优点:
采用大肠杆菌表达系统,对空载质粒pET28a进行改造,将强启动子T7替换为弱启动子tac、引入促溶标签GST,得到优化的空载质粒pETM,降低了转录效率,有助于保护重组蛋白免受胞外蛋白酶的降解并提高其稳定性,然后对PCV3 ORF2序列依据大肠杆菌密码子偏好进行了优化,将合成的优化序列与空载质粒pETM连接,重组质粒转染大肠杆菌,在低温下诱导表达,使目标蛋白PCV3 Cap全部表达在上清,解决了包涵体的困扰,有利于下游生产放大。
纯化和透析后的PCV3 Cap能够形成病毒样颗粒(VLPs),可以和PCV3阳性血清进行特异性结合,纯化的PCV3 Cap制备的亚单位疫苗对猪圆环病毒3型具有良好的防控效果。
本发明首次采用猪圆环病毒3型PCV3-HK/2019株进行免疫攻毒保护试验。通过小鼠攻毒保护试验,发现本发明中的PCV3 Cap免疫组对应的病毒拷贝数显著低于SUMO-cap181免疫组和TF-PCV3-dCap免疫组,表明本发明制备的猪圆环病毒3型亚单位疫苗对PCV3攻毒具有很好的保护效果;另外,该亚单位疫苗免疫仔猪后也能获得较好的免疫保护力。
附图说明
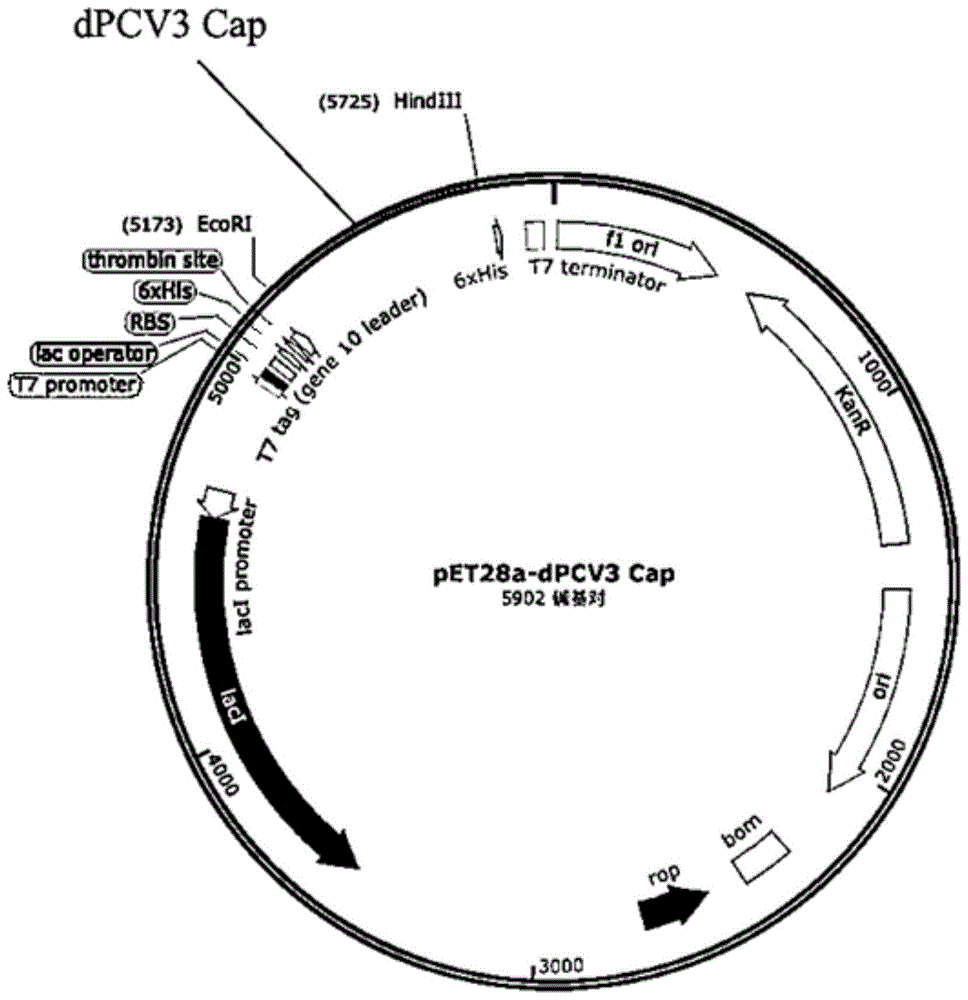
图1为表达载体pET28a-dPCV3 Cap的质粒图谱;
图2为不同实施例的重组质粒表达的PCV3 Cap蛋白的可溶性比较图;
M:蛋白质分子质量标准;1:实施例1超声沉淀;2:实施例1超声上清液;3;实施例2超声沉淀;4:实施例2超声上清液;5:实施例3超声沉淀;6:实施例3超声上清液;7:实施例4超声沉淀;8:实施例4超声上清液;9:实施例5超声沉淀;10:实施例5超声上清液;11:实施例6超声沉淀;12:实施例6超声上清液;箭头指向为目的条带(下同);
图3为空载质粒pETM的质粒图谱;
图4为dPCV3 Cap基因插入空载质粒pETM获得的重组质粒的质粒图谱;
图5为dPCV3 Cap的N端引入信号肽PhoA基因再插入空载质粒pETM获得的重组质粒的质粒图谱;
图6为优化密码子的PCV3 Cap基因插入空载质粒pETM获得的重组质粒的质粒图谱;
图7为优化密码子的PCV3 Cap去NLS后插入空载质粒pETM获得的重组质粒的质粒图谱;
图8为优化密码子的PCV3 Cap基因插入空载质粒pET-30a获得的重组质粒的质粒图谱;
图9为实施例3、实施例4和实施例5表达的PCV3 Cap上清Ni-NTA纯化结果;
M:蛋白质分子质量标准;1-3:依次为实施例3纯化后产物、实施例4纯化后产物、实施例5纯化后产物;
图10为重组大肠杆菌ETM-optPCV3 cap/BL21蛋白表达Western Blotting鉴定结果;
图11为HisTrap HP FPLC纯化的PCV3 Cap的SDS-PAGE验证结果;
图12为Western Blotting验证PCV3 Cap免疫原性结果;
图13为PCV3 Cap形成VLP颗粒的透射电子显微镜图;
图14为病毒拷贝数;
图15为免疫后猪圆环病毒3型ELISA抗体OD
***:表示P<0.001,差异极显著;
图16为二免攻毒后分毒情况。
具体实施方式
为使本发明更加容易理解,下面结合具体实施例,进一步阐述本发明。应理解为,以下实施例仅用于说明本发明而不用于限制本发明的范围。以下实施例中所使用的生物材料、化学试剂以及未提及的具体实验方法,均为本领域材料、试剂及按照常规实验方法进行。
(一)筛选重组质粒表达PCV3 Cap
实施例1
(1)含去NLS核酸序列的猪圆环病毒3型ORF2蛋白基因的合成
ORF2编码猪圆环病毒3型的唯一结构蛋白Cap,构成病毒的核衣壳,并且编码的该蛋白是病毒的主要免疫原性蛋白,Cap蛋白的N端有一段由34个氨基酸组成的核定位信号肽(NLS),其作用是帮助Cap蛋白进入细胞核,该NLS核酸序列含有丰富的Arg残基和大肠埃希菌稀有密码子,可导致新生多肽链的提前终止并阻止Cap蛋白在大肠埃希菌中的表达,因此Cap蛋白在大肠埃希菌的原核表达系统里很难得到大量表达,Cap蛋白在原核表达系统中的表达情况多以包涵体形式表达,对蛋白的纯化带来不便,其反应原性和免疫原性亦不如可溶性蛋白好,截掉该区域有利于大肠埃希菌表达系统表达PCV3 Cap。
本实施例以猪圆环病毒3型PCV3-HK/2019株基因组为模板,参照PCV3 ORF2序列(NCBI GenBank,登录号MW767462.1),设计引物,截去PCV3 Cap N端32个氨基酸(即96个核苷酸)以去掉核定位序列,得到如SEQ ID No.1所示的片段(546bp):dPCV3 Cap。
(2)构建重组表达质粒表达PCV3 Cap蛋白
将其插入到空载pET28a(+)的EcoRI/HindIII之间,形成表达载体pET28a-dPCV3Cap(如图1所示),转化大肠埃希菌感受态细胞BL21(DE3),获得重组菌株dPCV3 Cap/BL21,诱导条件:1%接种量转接dPCV3 Cap/BL21至液体LB培养基,37℃,180rpm振荡培养至OD
实施例2
(1)重组空载质粒
形成包涵体的原因通常认为是由于目标蛋白的表达速率快于所折叠出天然结构的速率,发生了错误折叠或者部分折叠,暴露在外的疏水区域就会与其它结构相似的蛋白质发生相互作用从而形成聚集物,这些聚集物会形成不溶于水的致密包涵体。实施例1的重组质粒中T7启动子是一种强启动子,可提高外源蛋白的表达量,转录效率较强,导致蛋白不能正确折叠,形成包涵体,为此本实施例将空载质粒pET28a(+)的T7启动子替换为tac弱启动子,降低转录效率,并配合低温的诱导条件,解决错误折叠的问题。同时将GST(谷胱甘肽巯基转移酶)插入空载质粒pET28a(+)的NcoI、BamHI之间,构建成新的空载质粒pETM(如图3所示),GST标签有助于保护重组蛋白免受胞外蛋白酶的降解并提高其稳定性,可直接利用还原型谷胱甘肽琼脂糖凝胶亲和树脂进行纯化。
本实施例以空载质粒pET28a(+)为模板,设计两条互补引物,其中包括如SEQ IDNo.8所示的正向引物:5'ATCTCGATCCCGCGAAATTTGACAATTAATCATCGGCTCGTATAATGGGAATTGTGAGCGGATAA3'和如SEQ ID No.9所示的反向引物:5'TTATCCGCTCACAATTCCCATTATACGAGCCGATGATTAATTGTCAAATTTCGCGGGATCGAGAT3',采用TaKaRa的PrimeSTAR Max DNA Polymerase进行扩增,反应体系为(50μL):PrimeSTAR Max Premix(2×)25μL,正向引物(10μM)1μL,反向引物(10μM)1μL,pET28a(+)(118ng/μL)2μL,ddH
其中tac启动子的核苷酸序列如SEQ ID No.2所示为(29bp),GST的核苷酸序列如SEQ ID No.3所示为(654bp)。
(2)含去NLS核酸序列的猪圆环病毒3型ORF2蛋白基因的合成
以猪圆环病毒3型PCV3-HK/2019株基因组为模板,参照PCV3 ORF2序列(NCBIGenBank,登录号MW767462.1),设计引物,截去PCV3 Cap N端32个氨基酸(即96个核苷酸)以去掉核定位序列,得到如SEQ ID No.1所示的片段(546bp):dPCV3 Cap。
(3)构建重组表达质粒表达PCV3 Cap蛋白
将SEQ ID No.1所示的dPCV3 Cap插入空载质粒pETM的EcoRI、XhoI之间(5’-3’),与促溶标签GST进行融合表达,形成表达载体pETM-dPCV3 Cap(如图4所示),再转化大肠埃希菌感受态细胞BL21(DE3),获得重组菌株为ETM-dPCV3 Cap/BL21。诱导条件:1%接种量转接ETM-dPCV3 Cap/BL21至液体LB培养基,37℃,180rpm振荡培养至OD
实施例3
(1)含去NLS核酸序列的猪圆环病毒3型ORF2蛋白基因的优化和合成
大肠埃希菌蛋白转运途径主要有Sec途径和Tat途径,大肠埃希菌在表达外源性蛋白时,信号肽能引导蛋白跨膜,转运至周质,并锚定在细胞膜上,能避免形成无活性的包涵体,并可保护蛋白,防止蛋白被胞内蛋白酶水解。碱性磷酸酶是一种周质蛋白,属于信号肽,被PhoA基因编码,PhoA基因的核苷酸序列如SEQ ID No.4所示为(63bp),PhoA基因编码的氨基酸序列如SEQ ID No.5所示为MKQSTIALALLPLLFTPVTKA。
为此本实施例在如SEQ ID No.1所示dPCV3 Cap片段的N端引入信号肽PhoA的核苷酸序列(SEQ ID No.4),得到重组基因片段PhoA-dPCV3 Cap,以促进表达的PCV3 Cap蛋白分泌性表达。
(2)构建重组表达质粒表达PCV3 Cap蛋白
将重组基因片段PhoA-dPCV3 Cap插入空载质粒pETM(如图3所示)的EcoRI、XhoI之间(5’-3’),形成表达载体pETM-PhoA-dPCV3 Cap(如图5所示),再转化大肠埃希菌感受态细胞BL21(DE3),获得重组菌株为ETM-PhoA-dPCV3 Cap/BL21,诱导条件:1%接种量转接ETM-PhoA-dPCV3 Cap/BL21至液体LB培养基,37℃,180rpm振荡培养至OD
实施例4
(1)优化猪圆环病毒3型ORF2蛋白基因
以猪圆环病毒3型PCV3-HK/2019株基因组为模板,参照PCV3 ORF2序列(NCBIGenBank,登录号MW767462.1)进行大肠埃希菌密码子优化,得到片段optPCV3 Cap,优化后的核苷酸序列如SEQ ID No.6所示为(663bp,核苷酸序列的第643~660位为6×His tag)。
(2)构建重组表达质粒表达PCV3 Cap蛋白
利用PCR方法,以f:5'CCG
原因可能是原核表达蛋白的效率与tRNA的含量有关,Cap蛋白NLS中高含量的精氨酸会减少大肠埃希菌中tRNA的浓度,导致蛋白质的合成速率下降,抑制外源蛋白的表达,未去掉核定位信号肽序列NLS时,降低了转录效率,在结合替换弱启动子后的空载质粒配合低温诱导表达条件,使目标蛋白的表达速率不快于所折叠出天然结构的速率,减少了错误折叠或者部分折叠,GST标签有助于保护重组蛋白免受胞外蛋白酶的降解并提高其稳定性。
实施例5
(1)优化猪圆环病毒3型ORF2蛋白基因
以猪圆环病毒3型PCV3-HK/2019株基因组为模板,参照PCV3 ORF2序列(NCBIGenBank,登录号MW767462.1)进行大肠杆菌密码子优化,得到片段optPCV3 Cap,优化后的核苷酸序列如SEQ ID No.6所示为(663bp,核苷酸序列的第643~660位为6×His tag)。设计引物,截去optPCV3 cap N端31个氨基酸(即93个核苷酸)以去掉核定位序列,得到如SEQID No.7所示的片段(570bp):optdPCV3 Cap。
(2)构建重组表达质粒表达PCV3 Cap蛋白
再将如SEQ ID No.7所示的片段optdPCV3 Cap插入空载质粒pETM的EcoRI、XhoI之间(5’-3’),形成表达载体pETM-optdPCV3 cap(如图7所示),再转化大肠埃希菌感受态细胞BL21(DE3),获得重组菌株为ETM-optdPCV3 cap/BL21,诱导条件:1%接种量转接ETM-optdPCV3 cap/BL21至液体LB培养基,37℃,180rpm振荡培养至OD
实施例6
(1)优化猪圆环病毒3型ORF2蛋白基因
以猪圆环病毒3型PCV3-HK/2019株基因组为模板,参照PCV3 ORF2序列(NCBIGenBank,登录号MW767462.1)进行大肠埃希菌密码子优化,得到片段optPCV3 Cap,优化后的核苷酸序列如SEQ ID No.6所示为(663bp,核苷酸序列的第643~660位为6×His tag)。
(2)构建重组表达质粒表达PCV3 Cap蛋白
利用PCR方法,以f:5'CCG
复筛
对比发现,只有实施例4中获得的PCV3 Cap全部表达在上清液,而实施例3和实施例5获得的PCV3 Cap部分在上清,因此,对实施例3、实施例4和实施例5进行复筛。
(1)按1%接种量,将重组菌株ETM-PhoA-dPCV3 Cap/BL21、ETM-optPCV3 cap/BL21和ETM-optdPCV3 cap/BL21对应的种子液分别转接至装有400mL液体LB培养基的锥形瓶中,37℃180rpm振荡培养至OD
(2)8,500rpm,4℃离心10min,收集菌体,并加入40ml无菌1×PBS进行菌体洗涤,然后再加入40ml无菌1×PBS重悬菌体,最后进行4℃水浴低温超高压连续流细胞破碎,压强设置为1,000bar,反复破碎2-3次,直到破碎液澄清;
(3)8,500rpm,4℃离心10min,收集上清液。
(4)上清进行Ni-NTA纯化,最后采用15ml洗脱液(26mM Tris,0.5M氯化钠,pH 7.4,500mM咪唑)洗脱,进行SDS-PAGE验证(如图9所示)。经过灰度法测定,实施例3洗脱液浓度为0.080mg/ml,实施例4洗脱液浓度为0.217mg/ml,实施例5洗脱液浓度为0.054mg/ml。
最终选取实施例4为最优方案。
(二)实施例4的重组菌株ETM-optPCV3 cap/BL21蛋白表达Western Blotting鉴定
取诱导前、后菌液,分别对其全菌、上清和沉淀进行Western Blotting鉴定PCV3Cap的表达,其中使用鼠源His-tag单克隆抗体和鼠源GST-tag单克隆抗体(稀释比例均为1:2000)分别作为一抗,二抗为HRP标记的羊抗鼠IgG(稀释比例为1:2000),结果如图10所示,在50kDa处出现特异性条带(箭头处)。
(三)实施例4的重组菌株ETM-optPCV3 cap/BL21高密度发酵及蛋白纯化
(1)将1L ETM-optPCV3 cap/BL21种子液转接至装有20L液体LB培养基(含200g甘油、20g消泡剂)的30L发酵罐,37℃,200rpm,培养至OD
(2)菌体处理:菌体用无菌1×PBS重悬,进行高压细胞破碎,然后10,000rpm,4℃,离心10min,收集上清。采用AKTA pure25蛋白质层析纯化系统、GE公司的HisTrap HP预装柱进行FPLC纯化。先通过以下步骤平衡预装柱:水洗:5CV(1CV=50mL,下同),流速5mL/min;Binding Buffer平衡(500mM NaCl,20mM Tris-base,5mM咪唑,pH7.5需过滤、需灭菌):2CV,流速7mL/min pH值、电导值稳定;然后上样:流速10mL/min;从waste管路中接流穿样品;将入口转移至Washing Buffer(500mM NaCl,27mM Tris-base,60mM咪唑,pH7.5需过滤、需灭菌)中,洗柱,2CV,流速10mL/min,收集洗杂样品;然后用Elution Buffer(500mM NaCl,27mMTris-base,500mM咪唑,pH7.5需过滤、需灭菌)进行洗脱,流速10mL/min,当Uv值达到期望值(400),等待2mL滞留体积开始收样,当Uv值降至期望值(100or 200),等待2mL滞留体积停止收样;
(3)对收集的流穿、洗杂和洗脱,进行SDS-PAGE验证(如图11所示),经过灰度法测定,浓度为0.573mg/ml。
(四)体外验证其免疫原性;
重组菌株ETM-optPCV3 cap/BL21高密度发酵后进行蛋白纯化获得的PCV3 Cap进行Western Blot验证,其中感染猪圆环病毒3型的猪阳性血清作为一抗(稀释比例为1:500),HRP标记的兔抗猪IgG作为二抗(稀释比例为1:2000)(见图12)。
(五)PCV3 cap蛋白VLP电镜图片;
将步骤(三)纯化后的PCV3 Cap用1×PBS 4℃透析过夜除盐,然后将透析后的组分用1×PBS稀释100倍,用镊子夹取200目的铜网放在样品上,室温吸附5min,用滤纸吸去多余的样品,用3%的磷酸钨染色3min,用滤纸吸去多余的样品,用透射电子显微镜观察样品,分析VLP的组装情况(见图13)。成功自组装了17~20nm的猪圆环病毒3型病毒样颗粒,而现有技术CN 201810420766.3使用含促溶标签SUMO的质粒连接截短的PCV3-cap181基因序列,获得重组质粒,虽然通过重组菌成功表达了可溶PCV3 Cap蛋白,但该目的蛋白需在特殊组分的缓冲溶液A中透析才能组装成病毒样颗粒。实施例4表达的PCV3 Cap蛋白没有截短,形成的VLPs更接近天然构象且组装效率更高。目前通过优化PCV3Cap蛋白完整全长基因并能表达可溶目标蛋白,且能自组装病毒样颗粒的仅有涂亚斌等人公布的申请号为201910863646.5的发明专利,未提及洗脱液中PCV3 Cap蛋白的浓度,该发明使用的载体为pET-30a,具有T7强启动子,其参照GenBank登录号MN 310686中的野生型优化后的PCV3 Cap蛋白完整全长基因如下,与实施例4中的序列的同源性为85.4%。
根据在“猪圆环病毒3型Cap蛋白的原核表达及其免疫原性分析”的报道涂亚斌等人在“Self-assembly into virus-like particles of the recombinant capsidprotein of porcine circovirus type 3and its application on antibodiesdetection”中公布了利用原核表达系统也无法表达完整的Cap蛋白或蛋白表达量过低。本发明在实施例6中采用该现有技术的方案,用优化后的optPCV3 cap给予连接到pET-30a上表达PCV3 Cap蛋白,发现无明显条带。
(六)含不同促溶标签的PCV3 Cap制苗后的攻毒保护效果对比
(1)将实施例4的PCV3 Cap 120μg/ml、SUMO-cap181 120μg/ml和TF-PCV3-dCap120μg/ml分别与Summit Poly Solution佐剂按4:1(V/V)混合,300~500rpm乳化30min,制备成对应的单苗;
(2)将40只18-22g的BALB/C小鼠饲喂适应一周,随即分成4组,即PCV3 Cap单苗组、SUMO-cap181单苗组、TF-PCV3-dCap单苗组和对照组。每只小鼠进行肌肉注射0.2ml,对照组注射同等剂量的PBS溶液,7天后,按照同等方法进行二次免疫,剂量为0.2ml。免疫后14d,进行猪圆环病毒3型PCV3-HK/2019株(NCBI GenBank,登录号MW767462.1)攻毒(病毒含量为10
(3)测定病毒拷贝数,需先进行荧光定量PCR,获得Ct值。荧光定量反应体系包括:2×AceQ qPCR SYBR Green Master Mix 10μL、上、下游引物各0.4μL、50×ROX ReferenceDye 1 0.4μL,病毒核酸2μL、ddH2O补至20μL。其中,上游引物序列为PCV3-F:5'-TGTGGTGCTACGAATGTCC-3',下游引物序列为PCV3-R:5'-GGTATCTCGCTATACTGCCTC-3'。荧光定量反应程序包括:95℃5min;95℃10s,60℃30s,40个循环,溶解曲线分析从60℃到95℃。其中,Ct值与拷贝数之间的关系为:Ct=-3.585×Log10(拷贝数)+42.591(R
(4)结果表明,PCV3 Cap单苗组对应的病毒拷贝数显著低于SUMO-cap181单苗组和TF-PCV3-dCap单苗组,表明本发明制备的猪圆环病毒3型亚单位疫苗对PCV3攻毒具有很好的保护效果,结果如图14所示。
(七)PCV3 Cap单苗免疫仔猪后的攻毒保护效果评估
(1)将PCV3 Cap 120μg/ml与Summit Poly Solution佐剂按4:1(V/V)混合,300~500rpm乳化30min,制备成猪圆环病毒3型亚单位疫苗;
(2)用14~21日龄健康易感仔猪15头,随机分成3组,每组5头,即PCV3 Cap单苗免疫组、非免疫攻毒组和空白对照组,免疫组每头仔猪肌肉注射2ml,非免疫攻毒组注射同等剂量的PBS溶液。首免后21天,按照同等方法进行二次免疫,剂量为2ml;
(3)二免后14日,对所有仔猪进行前腔静脉采血,然后将离心管中的血置于37℃恒温箱1小时,再置于4℃过夜,待血液凝固血块收缩后,4,000rpm离心10分钟,取上清于洁净的离心管中,即为待测血清,保存于-20℃;
(4)采用间接Elisa法检测血清效价,以PCV3 Cap包板,每孔包被量为50ng,对待测血清分别进行40倍稀释,然后加入酶标二抗(羊抗鼠),最后加入TMB显色,测定450nm处的吸光值。
(5)在二免后14日,对所有仔猪进行称重,将免疫组、非免疫攻毒组分别进行攻毒,每头猪滴鼻接种猪圆环病毒3型PCV3-HK/2019株病毒液3.0ml和颈部肌肉注射2.0ml(病毒含量为10
(6)攻毒后28日,剖杀所有仔猪并取肠系膜淋巴结和腹股沟淋巴结分别进行病毒分离,肠系膜淋巴结和腹股沟淋巴结两者任一组织分离到病毒即可判为病毒分离阳性。
(7)攻毒试验组(免疫组、非免疫攻毒组)仔猪与空白对照组(非免疫、非攻毒组)仔猪相比,若相对增长率≥5.0%,则判该仔猪生长发育不良,有猪圆环病毒3型引起的临床症状。
(a)空白对照组平均日增重=(攻毒后28日空白对照组仔猪体重之和-攻毒当日空白对照组仔猪体重之和)/(28日×5头);
(b)攻毒试验组仔猪平均日增重=(攻毒后28日攻毒试验组仔猪体重之和-攻毒当日攻毒试验组仔猪体重之和)/(28日×5头);
(c)攻毒试验仔猪平均日增重=(攻毒后28日该仔猪体重-攻毒当日该仔猪体重)/28日;(d)同时满足相对增长率<5%,且肠系膜淋巴结和腹股沟淋巴结组织均分离不到病毒即可判为保护。
(8)免疫后抗体见表1,PCV3 Cap单苗免疫组二免后14日的抗体效价显著高于非免疫攻毒组和空白对照组(如图15所示),二免攻毒后分毒情况如图16,且从表2结果可以看出,PCV3 Cap制苗免疫后,攻毒保护为4/5;非免疫攻毒组5/5发病,表明本发明所获得的猪圆环病毒3型病毒样颗粒亚单位疫苗对仔猪具有较好的攻毒保护效果。
表1免疫后猪圆环病毒3型ELISA抗体OD
表2免疫后猪圆环病毒3型PCV3-HK/2019株攻毒保护性试验结果
注:“+”表示病毒分离阳性;“-”表示病毒分离阴性。