番茄植物对ToBRFV的抗性
文献发布时间:2023-06-19 19:07:35
技术领域
本发明涉及番茄(Solanum lycopersicum)植物,也称为西红柿(Lycopersicumesculentum),对烟草花叶病毒番茄褐色皱果病毒(ToBRFV,以前缩写为TBRFV)的抗性。更具体地,本发明涉及包含导致对番茄褐色皱果病毒抗性的遗传决定因子的番茄植物和果实。根据本发明,抗性由DNA序列或QTL提供,从醋栗番茄(S.pimpinellifolium)基因渗入到番茄(S.lycopersicum)植物的9号或11号染色体上的基因组中。9号染色体上的基因渗入的QTL可以纯合或杂合地存在于番茄植物的基因组中。11号染色体上的基因渗入的QTL优选纯合地存在于番茄植物的基因组中。本发明还涉及与所述DNA序列关联的标记物,以及此标记物识别或选择DNA序列或QTL,并识别或选择携带此种抗性的植物的用途。本发明还涉及此类植物的种子和子代以及用于获得此类植物的繁殖材料,以及这些植物的不同用途。
背景技术
所有栽培和商业形式的番茄都属于最常被称为番茄(Lycopersicon esculentumMiller)的物种。番茄属(Lycopersicon)是极其庞大和多样化的茄科(Solanaceae)中的一个相对较小的属,茄科被认为由大约90个属组成,包括辣椒、烟草和茄子。番茄属已分为两个亚属,番茄(esculentum)复合物,其包含那些可以很容易地与商业番茄杂交的物种,以及秘鲁番茄(peruvianum)复合物,其包含那些很难杂交的物种(Stevens,M.和Rick,C.M.1986)。由于其作为作物的价值,番茄(L.esculentum)已在全世界广泛传播。即使栽培番茄的确切起源尚不清楚,但它似乎来自美洲,原产于厄瓜多尔、秘鲁和加拉帕戈斯群岛,最早于公元700年由阿兹特克人(Aztecs)和印加人(Incas)栽培。墨西哥似乎是最早引进的驯化地和来源地。据推测樱桃番茄(L.esculentum var.cerasiforme)是现代栽培形式的直系祖先。
番茄因其果实而种植,广泛用于新鲜市场或加工产品。作为作物,只要环境条件允许产生经济上可行的产量,番茄就可以进行商业化种植。大多数新鲜市场的番茄都是在藤蔓成熟和成熟的绿色成熟阶段手工采摘的。新鲜市场的番茄全年供应。加工番茄大多采用机械收获,并以多种形式使用,如番茄罐头、番茄汁、番茄酱、果泥、番茄膏或甚至番茄沙司。
番茄通常是简单的二倍体物种,具有十二对分化的染色体。然而,多倍体番茄也是本发明的一部分。栽培的番茄是自花授粉的,几乎完全自花授粉。番茄花是雌雄同体。商业栽培品种最初是开放授粉的。由于已在番茄中识别出杂种优势,由于具有更高产量和更均匀的植物特性而在农民中越来越受欢迎,杂种正在取代开放授粉品种。由于其传播广且价值高,番茄得到了集约化培育。这就解释了为什么现在存在如此广泛的番茄种类。形状从小到大,并且有樱桃型、李子型、梨型、块状、圆形和牛排型。番茄可以根据植物到供收获的成熟果实所需的时间进行分组,一般来说,栽培品种被认为是早熟、中熟或晚熟的。番茄也可以根据植物的生长习性进行分组;分组为确定的、半确定的或不确定的。确定的植物倾向于先长出枝叶,然后开花,如果授粉成功花就会成熟为果实。所有的果实倾向于在大约同一时间在植物上成熟。不确定的番茄由生长一些枝叶开始,然后在整个生长季节继续产生枝叶和花朵。在任何给定时间,这些植物往往会具有处于不同成熟阶段的番茄果实。半确定的番茄具有的表型在确定和不确定之间,它们是典型的确定型,除了长得比确定品种大以外。番茄育种的最新发展导致了更广泛的果实颜色种类。除了标准的红色成熟颜色外,番茄还可以是乳白色、柠檬绿、粉红色、黄色、金色、橙色或紫色。
可以通过人工授粉生产杂交商业番茄种子。收获雄性亲本的花粉,并人工施用于雌性近交系的柱头表面。在人工授粉之前和之后,花朵被覆盖,这样昆虫就不会带来外来花粉并创造混合物或杂质。花朵被标记以识别授粉的果实,从授粉的果实中收获种子。
多种病原体影响番茄植物的生产力,包括病毒、真菌、细菌、线虫和昆虫。番茄尤其易受许多病毒的影响,因此病毒抗性在农业上具有重要意义。
烟草花叶病毒是在农业中造成严重损害的最重要的植物病毒之一,尤其是对全世界的蔬菜和观赏作物。烟草花叶病毒很容易通过机械方式传播,以及通过种子传播。烟草花叶病毒的一般特征是约300nm的杆状颗粒包裹着编码四种蛋白质的单链阳性RNA基因组。在番茄中,烟草花叶病毒(TMV)和番茄花叶病毒(ToMV)受到全世界种植者的担忧,因为它们会严重损害作物生产,例如通过不规则的成熟(果实表面有黄色斑点,表面下方有褐色斑点)。然而,多年来植物育种者已经识别了多个基因,并且现在存在TMV和/或ToMV的抗性番茄品种。
在过去的几十年里,所有现代不确定番茄品种和许多确定番茄品种确实含有Tm-2基因或优选地该基因的Tm-2
2014至2015年期间,严重的病毒爆发影响了中东例如约旦和以色列的番茄产区。大多数受影响的番茄品种被认为具有TMV和/或ToMV抗性,但仍然受到严重影响并表现出典型的TMV/ToMV样症状:虽然枝叶的症状与TMV/ToMV症状非常相似,但果实症状比具有果实病变和变形的此类病毒的常见症状更为频繁和严重。果实质量很差,相当不易销售。Salem等人(Arch.Virol.161(2),503-506.2015)从约旦感染的有症状植物的果实和叶片中提取RNA,并进行了各种测试,最终识别出一种新的烟草花叶病毒物种,其序列对应于GenBank保藏号KT383474(SEQ ID NO:112);Salem等人提议将这种约旦病毒命名为:番茄褐色皱果病毒(以前是TBRFV,现在为ToBRFV)。与其他烟草花叶病毒序列的比较,表明番茄褐色皱果病毒确实是一种烟草花叶病毒,但不是TMV或ToMV。对TMV和/或ToMV的抗性并不赋予对这种新病毒ToBRFV的抗性。
Luria等人(PLoS One.2017;12(1):e0170429)同时分离并测序了在以色列感染番茄的以色列烟草花叶病毒的完整基因组,对应于GenBank登录号KX619418(SEQ ID NO:113)。因此,他们在以色列病毒和约旦病毒之间显示出非常高的序列同一性(超过99%的序列同一性),并得出了是两种不同的番茄褐色皱果病毒的分离株的结论。
最近,该病毒在欧洲被发现,特别是在西西里岛、德国、荷兰和法国以及墨西哥,因此现在它被认为是对番茄作物的主要全球威胁。识别的毒株似乎基本上是以色列毒株,而不是约旦毒株。
近期在WO2018/219941中描述了对番茄褐色皱果病毒表现出抗性的番茄植物的识别以及遗传决定因子的定位和识别,下文也称为QTL(数量性状位点)导致对番茄褐色皱果病毒的抗性,,尽管在此出版物中提到了耐受性。两个QTL,即QTL1和QTL2,分别发现于6号和9号染色体上,并且当纯合出现在番茄背景时独立地或组合地赋予感染或可能感染ToBRFV的番茄植物果实的改善的耐受性或抗性。第三个QTL,QTL3在11号染色体上发现,当纯合出现时,它赋予感染或可能感染ToBRFV的番茄植物叶的改善的耐受性或抗性。
尽管这些单独或组合的QTL,提供对ToBRFV的耐受性或抗性,但发明人现已确定,在大多数情况下,它们不能赋予番茄植物足够高水平的抗性,使得很大一部分果实受到影响并无法销售。此外,这些QTL被描述为当纯合出现时提供抗性。只要9号染色体上的QTL2存在于与Tm-2
WO2019/110130和WO2019/110821公开了对6、11和12号染色体上的3个不同QTL的识别,该QTL从醋栗番茄基因渗入并据称赋予对ToBRFV的抗性或耐受性。然而,11号染色体上的QTL被描述为位于2个标记物之间,这2个标记物定义了55Mb的区域,该区域对应于11号染色体的几乎整个序列。没有任何更明确的描述,11号染色体上的此QTL不能用于基因基因渗入渗入。
WO2020/018783公开了番茄11号染色体上的遗传区域,该区域包含来自醋栗番茄的匍柄霉属(Stemphylium)抗性等位基因,并且据称还包含相关的ToBRFV抗性等位基因。由于根据此文献的公开内容,两个等位基因都与相同的标记物相关联,因此不可以仅对ToBRFV抗性进行基因渗入,尤其是在已经对匍柄霉属具有抗性但对ToBRFV易感的植物中。
由于烟草花叶病毒不容易控制,而是通过在抗性基因繁殖中识别和使用进行遗传改良,并且由于目前可用于控制TMV和/或ToMV的抗性基因对新的番茄褐色皱果病毒的损害无用,并且已经确定的耐受性或抗性QTL并不总是足够有效,没有充分表征并且不能与Tm-2
发明内容
本发明人已经在野生型醋栗番茄(S.pimpinellifolium)植物中识别出对ToBRFV的抗性,并且已经能够将该抗性基因基因渗入渗入到番茄(S.lycopersicum)植物中,从而获得对ToBRFV具有抗性的番茄(S.lycopersicum)植物。本发明的抗性由新发现的序列赋予,该序列与加性数量性状基因座(QTL)连锁,可转移到不同的番茄遗传背景。
新发现的QTL基本上在被病毒感染的番茄植物的果实水平上,对于9号染色体上的QTL在被病毒感染的番茄植物的叶水平上,以及对于11号染色体上的QTL基本上是在受感染植物的叶水平上赋予对番茄褐色皱果病毒(ToBRFV)的抗性。
因此,本发明提供了这些基因渗入的序列,此处也称为QTL,它们在被ToBRFV感染的番茄植株的番茄叶和/或果实水平上赋予ToBRFV抗性表型。
本发明提供了对ToBRFV表现出抗性的番茄植物,包括商业植物、品系和杂交种,以及生产或识别对ToBRFV表现出抗性的番茄植物或种群(种质)的方法。本发明还公开了分子遗传标记物,尤其是单核苷酸多态性(SNP),其与本发明的负责对ToBRFV抗性的QTL连锁。还提供了通过这些分子标记物的方法和用途获得的植物。
此外,所述抗性可容易地转移到不同的遗传背景,即转移到多种番茄中,并且本发明还扩展到允许QTL赋予表型的转移或基因渗入的不同方法。
本发明还提供与赋予ToBRFV抗性的QTL相关的这些SNP相关的信息的多种方法和用途,尤其是用于识别抗ToBRFV植物的方法和用于识别与该抗性相关的进一步分子标记物的方法,以及在受ToBRFV侵染的环境中改善番茄产量的方法以及保护番茄田地免受ToBRFV侵染的方法。
术语“抗性”由ISF(国际种子联合会)蔬菜和观赏作物科定义,用于描述植物对害虫或病原体的反应,以及蔬菜种子行业的非生物胁迫。具体而言,抗性是指与在类似环境条件和害虫或病原体压力下的易感植物品种相比,植物品种限制特定害虫或病原体的生长和发育和/或它们造成的损害的能力。抗性品种在严重的害虫或病原体压力下可能表现出一些疾病症状或损害。定义了两个级别的抗性:
高抗性(HR):与易感植物相比,高度限制特定害虫的生长和/或发育和/或其在正常害虫压力下造成的损害的植物。然而,这些植物在严重的害虫压力下可能表现出一些症状或损害。
中等抗性(IR):高度限制特定害虫的生长和/或发育和/或其造成的损害,但与高抗性植物相比可能表现出更大范围的症状或损害的植物。当在类似的环境条件和/或害虫压力下生长时,中等抗性植物仍将表现出比易感植物更轻的症状或损害。
术语“耐受性”通常用于描述植物承受非生物压力而不会对生长、外观和产量造成严重后果的能力。
然而,在文献和专利中,该术语也用于表示植物的表型,其中在所述植物暴露于感染剂量的病毒时至少一些疾病症状仍然不存在,由此存在全身或局部感染、病毒繁殖,至少在某些培养条件下,至少可以确定所述植物细胞中病毒基因组序列的存在和/或其基因组整合。因此,耐受性植物对症状表现是抗性的,但对病毒携带者是无症状的。有时,病毒序列可以存在或甚至在植物中繁殖而不引起疾病症状。应当理解,耐受性植物虽然被病毒感染,但通常能够至少适度地限制病毒的生长和发育。
出于这个原因,根据这个定义的耐受植物最佳被表征为中等抗性植物。
在ToBRFV的情况下,叶抗性或枝叶抗性是指植物的表型,其中当所述植物暴露于感染剂量的ToBRFV时,叶片上的疾病症状仍然不存在或不太显著。然而,果实上的疾病症状可能出现在受感染的植物上。
在ToBRFV的情况下,果实抗性是指植物的表型,其中当所述植物暴露于感染剂量的ToBRFV时,果实上的疾病症状仍然不存在或不太显著。然而,叶片上的疾病症状可能出现在受感染的植物上。
ToBRFV感染的叶片上的症状通常包括花叶、小叶变形,在许多情况下还包括带状症状。ToBRFV感染的果实上的症状通常包括典型的黄色病变(变色)和果实变形。在许多情况下,果实上也有“巧克力斑点”。
易感性:植物无能力限制特定害虫或病原体的生长和发育;易感植物表现出与病毒感染相关的有害症状,即在ToBRFV感染的情况下枝叶损害和果实损害。
对番茄褐色皱果病毒易感的番茄植物,如在2015年Salem等人的出版物中提到的市售品种Candela。
到目前为止,即在本发明之前,在ToBRFV感染地区种植的所有市售番茄品种都对ToBRFV易感,或者对于那些携带耐受性QTL的植物没有足够的抗性,例如PCT申请WO2018/219941中提到的HAZTBRFVRES1的保藏种子。
因此,相对于品种Candela,更普遍地相对于在ToBRFV感染地区种植的任何商业番茄品种,包括耐受性植物,并且相对于HAZTBRFVRES1,根据本发明的植物至少改善了对ToBRFV的抗性或耐受性,。相对于对应于HAZTBRFVRES1的植物的改善的抗性在实验部分的实施例5中得到证明。
如本文所用,术语“后代”或“子代”是指从一种或多种亲本植物或其后代的无性或有性繁殖产生的作为子代的任何植物。例如,后代植物可以通过亲本植物的克隆或自交,或通过杂交两个亲本植物获得,并且包括自交以及F1或F2或更进一步的世代。F1是由亲本生产的第一代后代,其中至少一个亲本是首次用作性状的供体,而第二代(F2)或后续世代(F3、F4等)的后代是F1’、F2’等从自交生产的样本。因此,F1可以是(并且通常是)由两个真实遗传的亲本杂交产生的杂种(真实遗传对于性状是纯合的),而F2可以是(并且通常是)由所述F1杂种的自花授粉产生的后代。
如本文所用,术语“杂交”、“杂交的”、“异花授粉”或“杂交育种”是指将一株植物上的一朵花的花粉(人工或自然地)施加到另一株植物上的花的胚珠(柱头)上的过程。
如本文所用,术语“遗传决定因子(genetic determinant)”和/或“QTL”是指与生物学功能相关的任何DNA片段。因此,QTL和/或遗传决定因子包括但不限于基因、编码序列和/或它们表达所需的调控序列。QTL和/或遗传决定因子还可以包括非表达的DNA片段,例如,形成其他蛋白质的识别序列。
如本文所用,术语“基因型”是指单个细胞、细胞培养物、组织、生物体(例如植物)或生物体群的遗传构成。
如本文所用,术语“嫁接”是根状茎与接穗嫁接的操作。嫁接的主要动机是在疾病管理的遗传或化学方法不可用时,避免土壤传播的害虫和病原体造成的损害。将易感接穗嫁接到抗性根状茎上,可以提供抗性栽培品种,而无需将抗性培育到栽培品种中。此外,嫁接可以增强对非生物应激的耐受性,增加产量并导致更有效的水和养分使用。
如本文所用,术语“杂合子”是指二倍体或多倍体个体细胞或植物,其具有至少存在于一个基因座的不同等位基因(给定基因、遗传决定因子或序列的形式)。
如本文所用,术语“杂合的”是指在特定基因座处存在不同的等位基因(给定基因、遗传决定因子或序列的形式)。
如本文所用,“同源染色体”或“同系物”(或同源物)指在减数分裂期间彼此配对的一组一个母本染色体和一个父本染色体。这些拷贝在相同的基因座和相同的着丝粒位置具有相同类型的基因,但它们的序列或等位基因可能不同。
如本文所用,术语“纯合子”是指在所有同源染色体上的一个或多个基因座具有相同等位基因的个体细胞或植物。
如本文所用,术语“纯合的”是指在同源染色体区段中的一个或多个基因座处存在相同的等位基因。
如本文所用,术语“杂种”是指由在一个或多个基因上不同的亲本之间的杂交产生的任何个体细胞、组织或植物。
如本文所用,术语“基因座(locus)”(复数:“基因座(loci)”)是指任何在基因上被定义的位点,这可以是单个位置(核苷酸)或染色体区域。基因座可以是基因、遗传决定因子、基因的一部分或DNA序列,并且可以被不同的序列占据。位点也可以由SNP(单核苷酸多态性)、多个SNP或两个侧翼SNP来定义。
如本文所用,术语“根状茎”是能够在嫁接过程中接受接穗的植物的较低部分。
如本文所用,术语“接穗”是能够在嫁接过程中嫁接到根状茎上的植物的较高部分。
本发明包括不同倍性水平的植物,主要是二倍体植物,但也包括三倍体植物、四倍体植物等。
在本发明的上下文中,根据Illumina开发的TOP/BOT指定方法DNA链和等位基因被设计为TOP:
(https://www.illumina.com/documents/products/technotes/technote_topbot.p df)。
具体实施方式
本发明人已识别QTL,当其单独或组合存在于番茄植物中时,在感染或可能感染番茄褐色皱果病毒(ToBRFV)的番茄植物的果实和/或叶片中提供改善的抗性。
本发明人已经识别了9号染色体上的一个主要QTL,以下称为QTL9,当存在于番茄背景时,QTL9赋予对ToBRFV感染的抗性,尤其是果实抗性;以及11号染色体上的另一个加性QTL,在下文中称为QTL11,QTL11也赋予对ToBRFV的抗性,尤其是叶片抗性,并且可以与上文定义的QTL9组合。
根据本发明的种子和植物已经从醋栗番茄的野生植物(基因渗入的配偶体显示目的表型但在另一个物种中)与番茄植物(轮回易感亲本)之间的初始杂交获得,以便将抗性转移到番茄遗传背景中。
源自该初始杂交的抗性番茄植物的种子纯合地包含QTL9和QTL11,并称为LVSTBRFVRES2,已由发明人于2020年4月1日保藏在NCIMB,保藏号为NCIMB 43591。从这些保藏的种子长出的植物是番茄(S.lycopersicum),对ToBRFV具有抗性,即相对于任何已知的番茄植物,尤其是任何商业番茄植物,至少显示出对该病毒的改善的果实抗性。
如实施例中所示,根据本发明的植物的表型最好表征为对ToBRFV的抗性而非耐受性,即果实抗性或枝叶和果实抗性,耐受性仅适用于非生物应激。就耐受性也广泛用于表征抗性或中等抗性,然而本发明的植物也可以表征为耐受性植物。在下文中,参考对ToBRFV的抗性;然而,这种表型包括耐受性表型,如某些出版物中定义,以及中间抗性。
此外,还如实施例中所证实的,尤其是实施例4、5和8中所证实的,根据本发明的ToBRFV抗性不同于现有技术中公开的耐受性/抗性。事实上,实施例5证实了根据本发明的抗性水平高于WO2018/219941中描述的抗性水平。此外,实施例4证实了,根据本发明和根据WO2018/219941,负责ToBRFV抗性的序列是不同的。实施例8证实了,根据本发明的ToBRFV抗性与匍柄霉属抗性无关,与WO2020/018783中公开的遗传决定因子相反,因此不同于该抗性。
因此,根据第一个方面,本发明涉及番茄植物,在其基因组中包含第9染色体上的QTL,下文称为QTL9,和/或第11号染色体上的QTL,下文称为QTL11,在感染的情况下赋予对ToBRFV的改善的抗性,特别是对于QTL9在果实水平和对于QTL11在枝叶水平;,以及在其它们基因组中包含QTL9和/或QTL11的此种番茄植物的种子和细胞。
赋予抗性的所述QTL是最初从野生醋栗番茄基因渗入的,因此在下文中称为抗性QTL、或QTL9或QTL11、或本发明的基因渗入序列。本发明还涉及此类植物或种子的细胞,其包含这些赋予抗性的基因渗入序列。
耐受性/抗性表型可以如实验部分中所述进行测试和评分,尤其是在实施例1中,通过自然感染或通过人工接种,在第一叶片水平或果实水平。
赋予对ToBRFV的改善的抗性的QTL优选位于9号染色体上的由SNP TO-0201220(SEQ ID NO.1)和具有SEQ ID NO.101的SNP限定的染色体区间或区域内。
本发明人在实施例部分确实证明了该区域内与QTL9相对应的基因渗入序列与目的的表型一起被遗传。
根据本发明并赋予对ToBRFV改善的抗性的QTL9选自LVSTBRFVRES2种子基因组中存在的QTL9。因此,QTL9存在于这些保藏种子的基因组中。这些番茄种子的样本已由HM.Clause S.A.(rue Louis Saillant,26800Portes-les-Valence,法国)保藏,根据并满足《关于国际承认用于专利程序的微生物保藏的布达佩斯条约》(“布达佩斯条约”)的要求,于2020年4月1日保藏在国家工业、食品和海洋微生物保藏中心(NCIMB)(NCIMB,Ltd,Ferguson Building,Craibstone Estate,Bucksburn,Aberdeen AB21 9YA,英国)保藏号为43591。此番茄种子的保藏由HM.Clause S.A.(rue Louis Saillant,26800Portes-les-Valence,法国)保存。
根据本发明的赋予对ToBRFV改善的抗性的其他QTL优选位于11号染色体上,由SNPTO-0201237(SEQ ID NO:102)和SL2.50ch11_9924232(SEQ ID NO:115)限定的染色体间隔或区域内,优选由SNP TO-0201237(SEQ ID NO:102)和SNP TO-0201241(SEQ ID NO:106)限定的染色体间隔或区域内。根据本发明并赋予对ToBRFV改善的抗性的QTL11,可与QTL9组合,选自LVSTBRFVRES2、NCIMB 43591的种子的基因组中存在的那些。QTL11确实存在于LVSTBRFVRES2 NCIMB,保藏号43591的种子的基因组中。
与本说明书中提到的SNP(单核苷酸多态性)或标记物对应的特定多态性,以及这些SNP或标记物在番茄基因组中的侧翼序列,在实验部分(尤其参见针对QTL9的表G和H,和针对QTL11的表K)和随附的序列表中给出。它们的位置相对于9号和11号染色体上的番茄基因组2.50版,及其侧翼序列也进行了说明。
在这方面要注意,根据定义,SNP是指基因组中的单个核苷酸,它根据存在的等位基因而变化,而侧翼核苷酸是相同的。为了便于清楚地识别不同SNP的位置,它们的位置在表G、H和K中给出,通过参考其版本2.50中的番茄基因组序列和通过参考它们的侧翼序列,通过SEQ ID号识别。在本申请中与特定SNP相关的序列中,例如SNP TO-0201220的SEQ IDNO:1,序列中只有一个核苷酸实际对应于多态性,即SEQ ID NO:1的第201个核苷酸对应SNPTO-0201220的多态性位置,其可以是表G中所示的A或G。侧翼序列用于在基因组中定位SNP,但不是多态性本身的一部分。因此,检测SNP标记物或该SNP的等位基因是指检测该标记物的多态性核苷酸,并且不需要所有侧翼序列都相同。
类似地,说明书和表K中提到的其他标记物,严格来说不是SNP,是INDEL标记物,使单个核苷酸插入。例如,对于根据表K在SL2.50的9684449位置处的标记物SL2.50ch11_9684449(SEQ ID NO:112),这意味着位置9684449是等位基因,即是C或CT,对应于T的插入。在SL2.50中“C”的位置是表K中提到的位置9684449。只要等位基因仅涉及这些INDEL标记物的一个位置,为了简单起见,在下文中也可以通过语言扩展将它们称为SNP标记物。
由侧翼序列识别的基因组或染色体区域,例如SNP或INDEL标记物(下文同化为SNP)因此被明确且不含糊地定义。
由两个SNP X和Y限定的基因组区域是指基因组的部分,更具体地是染色体的部分,位于这两个SNP的位置之间,优选包括所述SNP,因此该染色体区域的核苷酸序列以对应于SNP X的核苷酸开始,以对应于SNP Y的核苷酸结束,即根据本发明,SNP包含在它们限定的区域内。
在给定的基因组区域所谓“来自醋栗番茄的基因渗入序列”,可以理解为,在该区域中发现的基因组序列与在醋栗番茄供体中发现的相应基因组序列具有相同的序列,即在基因渗入配偶体中,与在相同基因座的LVSTBRFVRES2(NCIMB 43591)中发现的相应基因组序列位于相同的基因座和具有相同的序列。所谓具有“相同序列”,是指要比较的两个序列是相同的,除了在基因组区域向子代传递过程中可能发生的潜在点突变外,即优选在1kb的长度上至少99%相同。
可以得出结论,在本发明的意义上,给定的基因组区域与在相同基因座的醋栗番茄供体中发现的相应基因组区域具有相同的序列,如果所述基因组区域也能够赋予对ToBRFV的抗性并且是源于醋栗番茄的。
例如,可以通过GISH(遗传原位杂交)显示番茄植物、种子或细胞的基因组中存在基因渗入的序列。GISH确实是强大的技术,用于检测染色质物质从一个物种或亚种基因渗入到另一个物种。GISH的优点是,基因渗入过程是通过“基因渗入的基因组图片”方式可视化的。使用这种技术,还可以确定基因组的特定区域是纯合子还是杂合子的,归功于使用共显性的分子细胞遗传学标记物。通过这种技术,还可以确定在哪条染色体中存在目的的基因渗入的基因。
本发明人已经识别并定位了赋予本发明的ToBRFV抗性的QTL,主要是通过识别代表在沿上述9号和11号染色体区域的不同基因座上的基因渗入的QTL的序列的存在,即由具有QTL9的SEQ ID NO.1至101的101个SNP定义的101个不同基因座,以及在由具有QTL11的SEQ ID NO:102至115的14个标记物定义的14个不同基因座。这些SNP在下文中称为本发明的SNP,或本发明对于QTL9的101个SNP。其中优选的SNP是具有SEQ ID NO:1至14的14个SNP;特别是具有SEQ ID NO:1、2、10、12和14的SNP。
因此,在本发明的植物、种子或细胞的基因组中,可以基于这些SNP标记物来识别赋予抗性表型的基因渗入的序列或QTL的存在。
优选地,对于QTL9,通过具有SEQ ID NO:1-101的101个SNP之一,并且优选通过选自包含SNP TO-0201220(SEQ ID NO:1)、TO-0201221(SEQ ID NO:2)、TO-0201222(SEQ IDNO:3)、TO-0201223(SEQ ID NO:4)、TO-0201224(SEQ ID NO:5)、TO-0201225(SEQ ID NO:6)、TO-0201226(SEQ ID NO:7)、TO-0201227(SEQ ID NO:8)、TO-0201228(SEQ ID NO:9)、TO-0201229(SEQ ID NO:10)、TO-0201230(SEQ ID NO:11)、TO-0201231(SEQ ID NO:12)、TO-0201232(SEQ ID NO:13)和TO-0201233(SEQ ID NO:14)的14个SNP列表中的SNP,更优选通过TO-0201220、TO-0201221、TO-0201229、TO-0201231和TO-0201233的5种SNP中的一个在番茄植株中识别或表征该基因渗入的序列或QTL的存在。本发明的QTL9可以例如由SNP TO-0201220或SNP TO-0201229识别或表征。
另一个合适的SNP是具有SEQ ID NO:15、16、17、18、19、20、21、22、23、24、25、26;27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100和101的那些。
根据优选的实施方式,对于QTL9,本发明的番茄植株、细胞或种子中基因渗入的序列的存在是通过至少2个,优选至少3个,或至少5个所述101个SNP标记物,或所述14个SNP标记物,优选其中至少一个是SNP TO-0201220或SNP TO-0201229可识别。例如,赋予ToBRFV抗性的基因渗入的序列的存在通过由至少2个SNP构成的单倍体的存在来检测,例如一个SNP是TO-0201229,另一个SNP是从具有SEQ ID NO:1-101(除SEQ ID NO:10外)的SNP中选择的不同SNP。
对于本发明的101个SNP,在表H的最后一列中报告了代表赋予本发明抗性的QTL或基因渗入序列的这些分子标记物的等位基因。对于上述14个SNP标记物,代表基因渗入QTL的等位基因是SNP TO-0201220的等位基因G、TO-0201221的等位基因G、TO-0201222的等位基因A、TO-0201223的等位基因A、TO-0201224的等位基因A、TO-0201225的等位基因A、TO-0201226的等位基因A、TO-0201227的等位基因C、TO-0201228的等位基因C、TO-0201229的等位基因A、TO-0201230的等位基因C、TO-0201231的等位基因C、TO-020132的等位基因G和TO-020133的等位基因G。
因此,根据本发明的番茄植物、细胞或种子的基因组中存在QTL可以通过检测代表在所述基因座的QTL的序列来检测或揭示,更优选地通过检测具有SEQ ID NO:1-101的SNP的一个或多个抗性等位基因来检测或揭示,如表H的最后一列报告的,例如,通过检测SNPTO-0201220的等位基因G、TO-0201221的等位基因G、TO-0201222的等位基因A、TO-0201223的等位基因A、TO-0201224的等位基因A、TO-0201225的等位基因A、TO-0201226的等位基因A、TO-0201227的等位基因C、TO-0201228的等位基因C、TO-0201229的等位基因A、TO-0201230的等位基因C、TO-0201231的等位基因C、TO-0201232的等位基因G和TO-0201233的等位基因G中的至少一个,更优选通过检测SNP TO-0201220的等位基因G、SEQ ID TO-0201221的等位基因G、TO-0201229的等位基因A、TO-0201231的等位基因C或TO-0201233的等位基因G,并且甚至更优选通过检测SNP TO-0201220的等位基因G和/或TO-0201229的等位基因A来检测或揭示。
根据优选的实施方式,在番茄植物、细胞或种子的基因组中通过检测至少2个,优选3个,优选至少5个具有SEQ ID NO:1至101的SNP的抗性等位基因来检测本发明的在9号染色体的QTL,即QTL9,优选至少一个检测到的抗性等位基因是SNP TO-0201220的等位基因G和/或TO-0201229的等位基因A。
QTL9位于9号染色体的染色体间隔上,该染色体间隔的一侧由侧翼SNP TO-0201220限定或侧翼,另一侧由具有SEQ ID NO.101的侧翼SNP标记物限定或侧翼。发现QTL9的更优选的9号染色体的染色体间隔是由TO-0201220和TO-0201233限定的间隔。更优选的间隔是由具有SEQ ID NO:20和TO-0201233的SNP限定的间隔,或具有SEQ ID NO:22和TO-0201233的SNP之间的间隔,或具有SEQ ID NO:26和TO-0201233的SNP之间的间隔,或具有SEQ ID NO:30和TO-0201233的SNP之间的间隔,或者具有SEQ ID NO:34和TO-0201233的SNP之间的间隔,或具有SEQ ID NO:38和TO-0201233的SNP之间的间隔,或者优选SNP TO-0201221和TO-0201233之间的间隔。
根据本发明的QTL11优选由具有SEQ ID NO:102至115的标记物之一检测,优选由具有SEQ ID NO:102至111的SNP标记物之一检测,优选由SNP TO-0201237(SEQ ID NO:102)、TO-0201238(SEQ ID NO:103)、TO-0201239(SEQ ID NO:104)、TO-0201240(SEQ IDNO:105)和TO-0201241(SEQ ID NO:106)中的至少一个检测,和/或由标记物SL2.50ch11_9684449(SEQ ID NO:112)、SL2.50ch11_9779896(SEQ ID NO:113)、SL2.50ch11_9823405(SEQ ID NO:114)和SL2.50ch11_9924232(SEQ ID NO:115)中的至少一个检测。优选地,该QTL11的存在可以通过检测具有SEQ ID NO:102至115的标记物中的抗性等位基因之一来表征,优选通过具有SEQ ID NO:102至111的SNP的抗性等位基因之一来表征和/或具有SEQ IDNO:112至115的标记物之一来表征,如表K最后一列所公开的。根据优选的实施方式,QTL11的存在通过检测SL2.50ch11_9684449的等位基因CT、SL2.50ch11_9779896的等位基因AT、SL2.50ch11_9823405的等位基因C、SL2.50ch11_9924232的等位基因GT、TO-0201237的等位基因G、TO-0201238的等位基因A、TO-0201239的等位基因A、TO-0201240的等位基因A和TO-0201241的等位基因A中的至少一个表征,优选由TO-0201237的等位基因G、TO-0201238的等位基因A、TO-0201239的等位基因A、TO-0201240的等位基因A和TO-0201241的等位基因A中的至少一个表征,和/或SL2.50ch11_9684449的等位基因CT、SL2.50ch11_9779896的等位基因AT、SL2.50ch11_9823405的等位基因C和SL2.50ch11_9924232的等位基因GT中的至少一个表征。因此,QTLH的优选标记物是具有SEQ ID NO:102至115的那些(表K),或者具有SEQID 102-111的那些,或者标记物TO-0201237、TO-0201238、TO-0201239、TO-0201240、TO-0201241、SL2.50ch11_9684449、SL2.50ch11_9779896、SL2.50ch11_9823405和SL2.50ch11_9924232的列表,或者标记物TO-0201237、TO-0201238、TO-0201239、TO-0201240和TO-0201241的列表,或者标记物SL2.50ch11_9684449、SL2.50ch11_9779896、SL2.50ch11_9823405和SL2.50ch11_9924232的列表。优选的抗性等位基因是对应于这些不同列表并且在表K中给出的那些。这些标记物和/或抗性等位基因的列表适用于本发明的所有不同方面。
本发明的番茄植株、细胞或种子对于QTL9、QTL11或赋予ToBRFV抗性的本发明的基因渗入序列可以是纯合的。然而本发明不限于这种纯合的植株、细胞或种子。事实上,发明人还证明了由这种QTL9赋予的抗性是加性的,这种杂合地具有本发明的QTL9的植物也对ToBRFV具有抗性(见实验部分),抗性水平低于纯合地包含QTL的植物的抗性水平,但高于易感植物的水平。因此本发明还包括本发明如上文定义的在9号染色体上的其基因组中杂合地具有QTL9或基因渗入序列的番茄(S.lycopersicum)植株、细胞或种子。
QTL11也可以纯合地或杂合地存在于本发明的番茄植物、细胞或种子中,但仅在纯合阶段赋予ToBRFV抗性,因为抗性等位基因是隐性的(参见这方面的实施例9)。QTL9和QTL11的杂合的或纯合的存在可以独立定义。
根据优选的实施方式,本发明的植物、种子或细胞杂合地或纯合地包含QTL9,以及如上文所定义的纯合或杂合地包含QTL11。优选的组合是QTL9和QTL11都纯合地存在,QTL9杂合与QTL11纯合地,以及两个QTL都杂合地存在。
优选地,根据本发明的番茄植物是商业植物或品系。这种商业植物或品系优选地也表现出对ToMV(番茄花叶病毒)的抗性,例如由于存在Tm-2基因(等位基因Tm-2或Tm-2
根据本发明还设想了其它抗性或耐受性。
根据优选的实施方式,本发明的植物对凤果花叶病毒(Pepino Mosaic Virus,PepMV)没有抗性。根据另一个实施方式,本发明的番茄植物还具有PepMV抗性。
此外,本发明的商业植物在合适的条件下结出果实,其至少10克,优选地在完全成熟时25克,优选地完全成熟时至少100克,和/或甚至更优选地完全成熟时至少150克或至少200克。此外,每株植物的果实数量基本上不受本发明的QTL9的存在的影响,即根据本发明的植物的生产力对于具有相同基因型但缺失所述QTL9的植物不逊于超过20%。因此本发明的植物,例如从保藏的种子生长的植物,通常结出每簇至少3个番茄,优选地约4个,并且这些果实具有优选地150克至180克的重量。
仍然根据另一个实施方式,本发明的植物是确定的、不确定的或半不确定的植物或其种子或细胞,即对应于确定的、不确定的或半不确定的生长习性。
对于确定的,其是指倾向于先长出枝叶,然后开花,如果授粉成功则花就会成熟为果实的番茄植物。所有的果实往往几乎同时在植物上成熟。不确定的番茄由生长一些枝叶开始,然后在整个生长季节继续生产枝叶和花朵。在任何时间,这些植物往往会具有处于不同成熟阶段的番茄果实。半确定的番茄具有的表型介于确定和不确定之间,它们是典型的确定型,除了长得比确定品种大以外。
本发明还涉及番茄的杂交植物,可通过将纯合地携带本发明的QTL9的植物与另一种番茄杂交获得。由于本发明的QTL9是加性的,由上述杂交生产的番茄的杂交植物将具有对ToBRFV的抗性。优选地,其它番茄的杂交配偶体是没有本发明所述的QTL9,但可以包含WO2018/219941中描述的QTL之一,优选9号染色体上的QTL2或11号染色体上的QTL3。或者,其他番茄的杂交配偶体可以包含WO2020/018783、WO2019/110130和WO2019/110821中描述的11号染色体上的赋予ToBRFV抗性的QTL。
因此本发明还涉及番茄植物、种子或细胞,其纯合或杂合地包括所公开的QTL9,以及:
-9号染色体上的QTL2,如WO2018/219941中公开的;杂合地;
-11号染色体上的QTL3,如WO2018/219941中公开的,优选纯合地;
-11号染色体上的一个或多个QTL,如W02020/018783、WO2019/110130和WO2019/110821中公开的,纯合地或杂合地。
此外,就本发明的发明人已经识别出的11号染色体上的QTL,在下文中称为QTL11,其赋予ToBRFV抗性,该QTL11可以与本发明的QTL9组合存在。在11号染色体上的该QTL,也对应于从提供QTL9的醋栗番茄的基因渗入配偶体基因渗入的序列。与QTL11相对应的基因渗入的序列发现于11号染色体上的由SNP TO-0201237(SEQ ID NO:102)和SNP TO-0201241(SEQ ID NO:106)限定的区域内。如在本申请实验部分所证实的,该QTL11当存在于番茄植物的基因组中时,特别是与QTL9组合存在于番茄植物的基因组中时;相对于缺失所述QTL11的相同植物,提供增加的对ToBRFV的抗性。因此,每个QTL独立地赋予ToBRFV抗性,但是相对于仅由一个QTL提供的抗性,QTL的组合提供增加的抗性,即至少是累积的,特别是因为QTL11的抗性基本上在叶片水平,QTL9的抗性在果实水平。因此本发明还涉及其基因组中纯合地或杂合地包含此QTL11的番茄植物、细胞和种子,但优选纯合地以便赋予ToBRFV抗性,优选叶片抗性。优选地,发现此QTL11与本发明的QTL9组合。
因此,本发明还涉及包含QTL9和QTL11的番茄植物,从而相对于缺失所述QTL11的相应植物,增加了植物对ToBRFV的抗性水平;本发明还包括其细胞和种子。
此外,如上所公开的,提供ToMV和TMV抗性的抗性基因的存在是有利的,特别是对于不确定的商业品种。根据本发明的一些优选实施方式,赋予ToMV和TMV抗性的Tm-2
根据实施方式,本发明因此涉及植物、细胞或种子,其杂合地包含根据本发明的QTL9以及提供ToMV和TMV抗性的Tm-2
根据优选的实施方式,本发明的番茄植物、细胞或种子,因此杂合地包含QTL9和Tm-2
如同一申请人名下的申请PCT/IB2019/00674中公开的,Tm-1基因的存在也可以改善ToBRFV的抗性。因此本发明的植物、细胞或种子有利地也包含Tm-1基因。Tm-1基因尤其在Ishibashi等人,2007年的出版物(病毒RNA复制的抑制剂由植物抗性基因编码(Aninhibitor of viral RNA replication is encoded by a plant resistance gene)。PNAS August 21,2007 104(34)13833-13838)中定义;在文献中报道的优选‘Tm-1基因’是指编码具有Tm-1活性的蛋白质的基因序列,即抑制Tm-1易感的野生型ToMV菌株的病毒复制的能力,例如该文献中公开的ToMV-L菌株。根据优选的实施方式,根据本发明的Tm-1基因是Ishibashi等人中报道的编码具有754个氨基酸序列的蛋白质的基因,对应于SEQ ID No:115(NCBI BAF75724)或编码与该序列具有至少75%、优选至少80%的序列同一性并表现出Tm-1活性(Ishibashi等人,2007中报告的),即抑制病毒RNA复制的野生型Tm-1易感ToMV菌株的能力的蛋白质的基因。
因此,除了本发明的QTL9之外,本发明还包括纯合地或杂合地包含Tm-1基因的番茄植物、细胞或种子,还有潜在的Tm-2
仍根据另一种实施方式,本发明的植物在嫁接过程中用作接穗或根状茎。嫁接是一种在作物如葫芦科等中使用多年的处理,但最近才用于番茄。嫁接可用于对大地上的病原体(telluric pathogens),如疫霉属(Phytophthora)或某些线虫提供一定水平的抗性。因此,嫁接倾向于防止待栽培的植物或品种与受感染的土壤接触。有兴趣用作嫁接物或接穗的品种,可选为F1杂交种,嫁接到用作根状茎的抗性植物上。抗性根状茎保持健康,并从土壤中为从疾病中分离出的嫁接物提供正常供应。
如上所述,本发明涉及表现出改善的ToBRFV抗性的番茄植物,以及产生这些植物的种子,和这些植物或种子的细胞,或其他植物部分,包括从醋栗番茄基因渗入的其基因组中的抗性QTL9和/或QTL11,和涉及包含所述QTL的本发明植物的子代。
子代包括与根据本发明的植物杂交的第一、第二和所有进一步的后代,其中杂交包括与自身的杂交或与另一植物的杂交。根据本发明的植物或种子可以是从保藏的种子LVSTBRFVRES2生长的植物的子代或后代,保藏在NCIMB,保藏号NCIMB 43591。对于赋予改善表型的本发明的QTL9以及QTL11,从保藏的种子生长的植物确实是纯合的;因此,它们的基因组中携带9号和11号染色体的每个同源物上的目的QTL。它们可用于通过杂交和自交和/或回交将这些序列转移到另一个背景中。
本发明还涉及LVSTBRFVRES2(NCIMB 43591)的保藏的种子以及从这些种子之一生长的植物,纯合地包含有赋予目的表型的QTL9,以及QTL11。值得注意的是,这些种子不对应于植物品种,除了本发明的QTL之外,它们对于大多数基因不是纯合的;因此,在繁殖过程中它们的表型不是固定的,除了本发明的ToBRFV抗性/耐受性外;在繁殖过程中它们的大多数表型性状分离,除了本发明的ToBRFV抗性外。
本发明还涉及如上所定义的植物或种子,即含有纯合的或杂合的状态的目的QTL9和/或QTL11,所述序列赋予改善的表型,可能组合,其植物或种子通过从番茄植物转移QTL进入另一个番茄遗传背景可获得,番茄植物的代表性种子在NCIMB保藏号NCIMB-43591保藏,例如通过将所述植物与第二个番茄植物亲本杂交并选择带有控制目的表型的QTL9和/或QTL11的植物。在这种杂交中,可以转移QTL9以及如果适用QTL11。
需要注意的是,本发明的种子或植物可以通过不同的处理获得,并且不排他地通过本质上的生物处理获得。
根据此方面,本发明涉及番茄植物或种子,优选非天然存在的番茄植物或种子,在其基因组中可包含一个或多个突变,其为植物提供对番茄褐色皱果病毒的果实和/或枝叶抗性,突变存在于例如,在植物的基因组中,其代表性样本保藏于NCIMB保藏号NCIMB 43591下。
在另一种实施方式中,本发明涉及获得在其基因组中携带一个或多个突变的番茄植物或种子的方法,该方法为该植物提供对番茄褐色皱果病毒的果实和/或枝叶抗性。这种方法在实施例7中示出,并且可以包括:
a)用诱变剂处理待修饰的番茄植物的M0种子,以获得M1种子;
b)从如此获得的M1种子中生长植物以获得M1植物;
c)通过M1植物的自花授粉生产M2种子;和
d)可选地重复步骤b)和c)n次以获得M1+n种子。
M1+n种子生长成植物并接受ToBRFV感染。幸存的植物,或那些ToBRFV感染症状较轻的植物,再繁殖一代或多代,同时继续选择其对ToBRFV的果实和/或枝叶抗性。
在该方法中,步骤a)的M1种子可以通过化学诱变(例如EMS诱变)获得。其他化学诱变剂包括但不限于,硫酸二乙酯(des)、乙烯亚胺(ei)、丙磺酸内酯、N-甲基-N-亚硝基氨基甲酸酯(mnu)、N-亚硝基-N-甲基脲(NMU)、N-乙基-N-亚硝基脲(enu)和叠氮化钠。
或者,通过辐射诱导突变,辐射例如选自X射线、快中子、紫外线辐射。
在本发明的另一个实施方式中,突变是通过基因工程诱导的。这种突变还包括赋予ToBRFV果实和/或枝叶抗性的序列整合,以及用赋予ToBRFV果实和/或枝叶抗性或耐受性的替代序列替换驻留序列。优选地,突变是如上所述的QTL9和/或QTL11的整合,以替换番茄植物的同源序列。更优选地,突变是包含在番茄基因组的9号染色体上的SNP TO-0201220(SEQ ID NO:1)和SNP TO-0201233(SEQ ID NO:14)中的序列或其片段替换为存在于植物基因组中的9号染色体上的同源序列,该植物的代表性样品以保藏号NCIMB43591保藏于NCIMB,其中序列或其片段赋予对ToBRFV的抗性。根据另一种实施方式,突变是将番茄基因组的11号染色体上的SNP TO-0201237(SEQ ID NO:102)和SL2.50ch11_9924232(SEQ IDNO:115)中包含的序列或其片段(例如包含在SNP TO-0201237和SNP TO-0201241中的序列)替换为植物基因组中存在的11号染色体上的同源序列,其中代表性样本以保藏号NCIMB43591保藏于NCIMB,其中序列或其片段纯合地存在时赋予对ToBRFV的抗性。
可以使用的基因工程手段包括使用所有称为新育种技术(New BreedingTechniques)的技术,该技术是开发和/或用于通过遗传变异在植物中创造新特征的各种新技术,目的是靶向诱变,靶向引入新基因或基因沉默(RdDM)。这种新育种技术的例子是通过使用以下技术促进靶向序列变化:锌指核酸酶(ZFN)技术(ZFN-1,ZFN-2和ZFN-3,参见美国专利号9,145,565)、寡核苷酸定向诱变(ODM),同源转基因(Cisgenesis)和内源转基因(intragenesis)、嫁接(在GM根状茎)、反向育种、农杆菌渗入法(Agro-infiltration)(农杆菌渗入法“严格意义上”,农杆菌接种,浸花法),转录激活因子样效应物核酸酶(TALENs,参见美国专利号8,586,363和9,181,535)、CRISPR/Cas系统(参见美国专利号8,697,359;8,771,945;8,795,965;8,865,406;8,871,445;8,889,356;8,895,308;8,906,616;8,932,814;8,945,839;8,993,233;和8,999,641)、工程化大范围核酸酶(meganuclease)重新改造的归巢核酸内切酶、DNA引导基因组编辑(Gao等人,Nature Biotechnology(2016))和合成基因组学。靶向基因组编辑的主要部分,即新育种技术的另一个名称,是在基因组中预期修饰的选定位置诱导DNA双链断裂(DSB)的应用。此类应用可用于产生突变(例如,靶向突变或精确天然基因编辑)以及基因的精确插入(例如,同源基因、内源基因内或异源基因)。导致突变的应用通常被确定为定点核酸酶(SDN)技术,例如SDN1、SDN2和SDN3。对于SDN1,结果是靶向的,非特异性的基因缺失突变:DNA DSB的位置被精确选择,但宿主细胞的DNA修复是随机的,并导致小的核苷酸缺失、添加或替换。对于SDN2,SDN用于生成靶向DSB,并使用DNA修复模板(与目的DSB DNA序列相同的短DNA序列,除了一个或几个核苷酸变化外)来修复DSB:这导致所需的目的基因中的靶向和预定点突变。对于SDN3,SDN与包含新DNA序列(例如基因)的DNA修复模板一起使用。该技术的结果是将该DNA序列整合到植物基因组中。说明使用SDN3的最可能的应用是在选定的基因组位置插入顺基因、基因内的或转基因表达盒。每种技术的完整描述可以在欧盟委员会联合研究中心(JRC)前瞻性技术研究所2011年发布的题为“新植物育种技术-商业发展的现有技术和前景”的报告中找到。
根据本发明,对ToBRFV的抗性或中间抗性对应于本发明的抗性植物的非销售果实的显著减少;特别地,本文公开的抗性植物结出番茄,使得至少70%的果实在成熟时可销售,即没有变色斑点,没有褐变花萼,没有尺寸过小且表面粗糙,没有棕色坏死斑点。优选地,即使在双重感染的情况下,至少80%的果实在成熟时仍可销售,优选至少90%的果实在成熟时仍可销售。
如上所定义的,本发明植物的特征在于,在9号染色体上在由SNP TO-0201220和具有SEQ ID NO:101的SNP限定的该染色体区域存在基因渗入序列,和/或在11号染色体上在由SNP TO-0201237和SL2.50ch11_9924232,优选由SNP TO-0201237和SNP TO-0201241限定的该染色体区域存在基因渗入序列。然而,来自醋栗番茄的基因渗入序列可以在这些边界或侧翼序列之外发现。类似地,在上述区域内可以找到基因渗入序列,但整个区域不一定由基因渗入序列组成。鉴于发明人识别和使用的QTL9的标记物,优选在本发明的植物、种子或细胞的基因组中在以下的基因座发现赋予ToBRFV抗性的基因渗入序列:包含具有表H中提到的SEQ ID NO:1-101的101个SNP的101个基因座中的至少一个或多个,更优选在9号染色体上的以下14个基因座中的至少一个:包含TO-0201220的基因座、包含TO-0201221的基因座、包含TO-0201222的基因座、包含TO-0201223的基因座、包含TO-0201224的基因座、包含TO-0201225的基因座、包含TO-0201226的基因座、包括TO-0201227的基因座、包含TO-0201228的基因座、包含TO-0201229的基因座、包含TO-0201230的基因座、包含TO-0201231的基因座、包含TO-0201232的基因座和包含TO-0201233的基因座。
所谓“包含SNP标记物的基因座”,是指围绕SNP多态性的序列,优选从SNP上游约2兆碱基延伸到SNP下游约2兆碱基的序列,优选在SNP上游和下游1兆碱基,优选甚至0.5兆碱基的序列。
这些基因座的基因渗入序列是在对应于NICMB43591的种子LVSTBRFVRES2中的相应基因座上发现的序列。
对于QTL11,优选在本发明的植物、种子或细胞的基因组中在以下的基因座发现当纯合地存在时,赋予ToBRFV抗性的基因渗入序列,所述基因座为包含在表K中提到的具有SEQ ID NO:102-115的14个标记物的14个基因座中的至少一个或多个。这些基因座的基因渗入序列也是在对应于NICMB43591的种子LVSTBRFVRES2中的相应基因座上发现的序列。
本发明在另一方面还涉及可能从如上所述的本发明的种子或植物中获得的任何植物,以及这种植物的植物部分,以及最优选外植体、接穗、插条、种子、果实、根、根状茎、花粉、胚珠、胚、原生质体、叶、花药、茎、叶柄、子叶、花、根尖、下胚轴和任何其他植物部分,其中所述植物、外植体、接穗、插条、种子、果实、根、根状茎、花粉、胚珠、胚、原生质体、叶、花药、茎、叶柄、子叶、花、根尖、下胚轴和/或植物部分从根据本发明的第一方面的种子或植物中可获得,即在其基因组中纯合地或杂合地携带目的的QTL9和/或QTL11。这些植物部分,特别是外植体、接穗、插条、种子、果实、根、根状茎、花粉、胚珠、胚、原生质体、叶、花药、茎、叶柄、子叶、花、根尖或下胚轴,在其基因组中包括赋予目的表型的QTL9和/或QTL11,即对ToBRFV的抗性,特别是对QTL9的果实抗性和对QTL11的枝叶抗性。根据优选的实施方式,本发明涉及如上所述的种子发育成根据本发明第一方面的植物,从而由于抗性QTL9或QTL11或基因渗入序列的存在,而对ToBRFV感染具有抗性,如上文所定义。
本发明的这个方面中提到的QTL9和QTL11是上面在本发明植物的上下文中定义的那些。
植物部分可以有利地包含如上所定义的QTL11,以补充或代替QTL9。
与本发明第一方面相关定义的QTL的不同特征经必要修正后适用于本发明的这一方面。因此,QTL9优选选自与保藏材料LVSTBRFVRES2(NCIMB保藏号43591)相对应的植物基因组中存在的那些。其有利的特征在于:存在表H的SNP的至少一个抗性等位基因;优选存在SNP TO-0201220的等位基因G、TO-0201221的等位基因G、TO-0201222的等位基因A、TO-0201223的等位基因A、TO-0201224的等位基因A、TO-0201225的等位基因A、TO-0201226的等位基因A、TO-0201227的等位基因C、TO-0201228的等位基因C、TO-0201229的等位基因A、TO-0201230的等位基因C、TO-0201231的等位基因C、TO-0201232的等位基因G和/或TO-020133的等位基因G,更优选存在SNP TO-0201220的等位基因G、SEQ ID TO-0201221的等位基因G、TO-0201229的等位基因A、TO-0201231的等位基因C或TO-0201233的等位基因G,甚至更优选存在SNP TO-0201220的等位基因G和/或TO-0201229的等位基因A。
QTL11优选地选自保藏材料LVSTBRFVRES2(NCIMB保藏号43591)相对应的植物基因组中存在的那些。其有利的特征在于:存在表K的标记物的至少一个抗性等位基因;优选存在TO-0201237的等位基因G、TO-0201238的等位基因A、TO-0201239的等位基因A、TO-0201240的等位基因A、TO-0201241的等位基因A、SL2.50ch11_9684449的等位基因CT、SL2.50ch11_9779896的等位基因AT、SL2.50ch11_9823405的等位基因C和SL2.50ch11_9924232的等位基因GT;例如,存在TO-0201237的等位基因G、TO-0201238的等位基因A、TO-0201239的等位基因A、TO-0201240的等位基因A和TO-0201241的等位基因A中的至少一个。
本发明还涉及番茄植物的细胞,使得这些细胞在其基因组中包含独立地赋予番茄植物目的表型的本发明的QTL9或QTL11,以及潜在的QTL9和QTL11。QTL9以及QTL11是已经在本发明框架中定义的一种,其特征在于:已经公开的关于根据本发明前述方面的植物和种子的相同特征和优选实施方式。这种QTL的存在,即QTL9或QTL11可以通过上述公开的技术揭示,并且为本领域技术人员所熟知。除其他外,可以确定QTL是否纯合或杂合地存在于本发明的此细胞的基因组中。QTL9的有利特征在于:存在表H的SNP的至少一个抗性等位基因;优选存在SNP TO-0201220的等位基因G、TO-0201221的等位基因G、TO-0201222的等位基因A、TO-0201223的等位基因A、TO-0201224的等位基因A、TO-0201225的等位基因A、TO-0201226的等位基因A、TO-0201227的等位基因C、TO-0201228的等位基因C、TO-0201229的等位基因A、TO-0201230的等位基因C、TO-0201231的等位基因C、TO-0201232的等位基因G和/或TO-020133的等位基因G,更优选存在SNP TO-0201220的等位基因G、SEQ ID TO-0201221的等位基因G、TO-0201229的等位基因A、TO-0201231的等位基因C或TO-0201233的等位基因G,甚至更优选存在SNP TO-0201220的等位基因G和/或TO-0201229的等位基因A。QTL11的有利特征在于:存在表K的标记物的至少一个抗性等位基因,优选存在TO-0201237的等位基因G、TO-0201238的等位基因A、TO-0201239的等位基因A、TO-0201240的等位基因A、TO-0201241的等位基因A、SL2.50ch11_9684449的等位基因CT、SL2.50ch11_9779896的等位基因AT、SL2.50ch11_9823405的等位基因C和SL2.50ch11_9924232的等位基因GT;例如,存在TO-0201237的等位基因G、TO-0201238的等位基因A、TO-0201239的等位基因A、TO-0201240的等位基因A和TO-0201241的等位基因A中的至少一个。
根据本发明的细胞可以是携带有目的QTL9和/或QTL11的任何类型的番茄细胞,特别是分离的细胞和/或能够再生整个番茄植物的细胞。
本发明还涉及根据本发明上述定义的植物的非再生或可再生细胞的组织培养物;优选地,可再生细胞来源于本发明的胚、原生质体、分生组织细胞、愈伤组织、花粉、叶、花药、茎、叶柄、根、根尖、果实、种子、花、子叶和/或下胚轴,并且细胞在其基因组中包含独立地赋予改善的表型的QTL9和/或QTL11,即QTL9的对ToBRFV的果实抗性和QTL11的对ToBRFV的枝叶抗性。
优选地,该细胞纯合地或杂合地包含QTL9并且还包括QTL11,如本发明上下文中定义的。
该细胞还有利地包含任何额外的抗性或耐受性基因,如在本发明的第一方面上下文中所公开的,也适用于此。
组织培养物优选能够再生具有前述番茄植物的生理和形态特征的植物,以及再生具有与前述番茄植物基本相同基因型的植物。本发明还提供了从本发明的组织培养物再生的番茄植物。
本发明还提供了上述定义的植物或者来自上述定义的组织培养物的原生质体,所述原生质体包含赋予本发明的改善的表型的QTL9和/或QTL11。
本发明还涉及本发明植物的组织;该组织可以是未分化的组织,或是分化的组织。这样的组织包含一个或多个包含本发明QTL的细胞。
本发明还涉及繁殖材料,其能够生产根据本发明的抗性番茄植物,包含如上定义的基因渗入序列或QTL。
根据另一方面,本发明还涉及本发明的番茄植物优选纯合地包含本发明的QTL9,作为育种方案中的育种配偶体用于获得具有本发明的改善表型的用途。事实上,这样的育种配偶体在其基因组中纯合地包含赋予目的表型的QTL9。通过将该植物与番茄植物,尤其是品系杂交,因此可以将赋予所需表型的本发明的QTL9转移给后代。因此,根据本发明的植物可以用作育种配偶体,用于将赋予所需表型的QTL9基因渗入到番茄植物或种质,即ToBRFV抗性。尽管杂合地携带目的QTL9的植物或种子也可以用作如上所述的育种配偶体,但表型的分离可能会使育种程序更加复杂。
本发明的改善的表型是对ToBRFV的抗性,特别是果实抗性或枝叶抗性,或果实和枝叶抗性。
育种配偶体还可以包含本发明中定义的QTL11,优选纯合地包含本发明中定义的QTL11。
基因渗入的QTL9将有利地被引入包含其他所需遗传性状的品种中,例如抗病性、果实早熟、耐旱性、果实形状等。优选地,基因渗入的QTL9将有利地引入包含Tm-2
根据另一种实施方式,本发明还涉及与本发明的番茄植物的相同用途,但纯合地包含本发明的QTL11作为育种配偶体。
本发明还涉及与纯合地包含QTL9和/或QTL11的在保藏号NCIMB 43591下保藏在NCIMB的LVSTBRFVRES2的植物或种子,以及源自其的植物相同的用途。所述植物也适合作为育种程序的基因渗入配偶体,旨在将所需的表型赋予番茄植物或种质。
在这样的育种程序中,选择显示所需表型或携带与所需表型相连的QTL9的子代可以有利地基于SNP标记物的等位基因进行,特别是基于具有SEQ ID NO:1-101的本发明的SNP标记物的等位基因进行。
对于QTL9,优选地在SNP TO-0201220的等位基因G、TO-0201221的等位基因G、TO-0201222的等位基因A、TO-0201223的等位基因A、TO-0201224的等位基因A、TO-0201225的等位基因A、TO-0201226的等位基因A、TO-0201227的等位基因C、TO-0201228的等位基因C、TO-0201229的等位基因A、TO-0201230的等位基因C、TO-0201231的等位基因C、TO-0201232的等位基因G和/或TO-020133的等位基因G的存在下选择植物的子代,更优选在SNP TO-0201220的等位基因G、SEQ ID TO-0201221的等位基因G、TO-0201229的等位基因A、TO-0201231的等位基因C或TO-0201233的等位基因G的存在下选择植物的子代,甚至更优选在SNP TO-0201220的等位基因G和/或TO-0201229的等位基因A的存在下选择植物的子代。
或者,可以基于与改善的表型相关的本发明的101个SNP的任何一种抗性等位基因或这些等位基因的组合的存在来进行选择。
这种选择将根据待选择植物的遗传物质样本中目的等位基因的存在进行。这个或这些等位基因的存在确实证实了本发明的QTL9或基因渗入序列在所述SNP定义的基因座的存在。然而,在点突变或重组事件之后,可以想象这些等位基因中的至少有1个或2个丢失,剩余的携带目的QTL9的染色体片段仍然赋予目的表型。
携带QTL11的子代的选择可以有利地基于具有SEQ ID NO:102-115的标记物的等位基因进行,优选具有SEQ ID NO:102-111的SNP标记物的等位基因进行,优选基于TO-0201237的等位基因G、TO-0201238的等位基因A、TO-0201239的等位基因A、TO-0201240的等位基因A、TO-0201241的等位基因A、SL2.50ch11_9684449的等位基因CT、SL2.50ch11_9779896的等位基因AT、SL2.50ch11_9823405的等位基因C和SL2.50ch11_9924232的等位基因GT的存在进行;例如,通过TO-0201237的等位基因G、TO-0201238的等位基因A、TO-0201239的等位基因A、TO-0201240的等位基因A和TO-0201241的等位基因A中的至少一个的存在进行。
因此,根据本发明的植物或从以保藏号NCIMB 43591保藏的种子生长的植物,在用于获得具有本发明的改善表型的商业番茄品系和品种的标记物辅助选择中特别有价值。
本发明还涉及所述植物在旨在识别、测序和/或克隆赋予所需表型的基因序列的程序中的用途。
本发明的前文方面描述的任何具体实施方式也适用于本发明的这个方面,特别是关于赋予目的表型的QTL9和QTL11的特征。
根据又一方面,本发明还涉及具有所需表型的番茄植物,特别是商业植物和近交系的生产或育种的方法或处理。本发明确实还涉及将赋予ToBRFV抗性的本发明的QTL或基因渗入序列转移到其它番茄植物,特别是其他番茄品种,或其他物种或近交系,并且可用于生产番茄的新类型和品种。
在这方面,本发明还包括育种具有ToBRFV抗性的番茄植物的方法,包括以下步骤:使从携带赋予ToBRFV抗性的本发明的QTL9的保藏的种子LVSTBRFVRES2 NCIMB 43591或其子代长出的植物与优选缺乏所述QTL的初始番茄植物杂交。QTL如上定义,即从醋栗番茄基因渗入,优选存在于LVSTBRFVRES2、NCIMB保藏号43591的种子的基因组中。该QTL可由具有SEQ ID NO:1-101的SNP标记物的至少一个抗性等位基因识别。
本发明还涉及一种对番茄植物赋予ToBRFV抗性的方法,包括对所述植物进行基因修饰以引入本发明的QTL9抗性。所述QTL9如上所定义,并且优选存在于LVSTBRFVRES2、NCIMB保藏号43591的种子的基因组中。基因修饰可以通过本领域技术人员熟知的任何方法或手段进行。
本发明还涉及用于培育具有与QTL11而非QTL9相关的ToBRFV抗性的番茄植物和赋予番茄植物与QTL11而非QTL9相关的ToBRFV抗性的相同方法。在这种情况下,QTL可以通过具有SEQ ID NO:102-115的标记物的至少一个抗性等位基因来识别。
因此,本发明涉及一种具有ToBRFV抗性的番茄植物的育种方法,包括以下步骤:使从携带在9号染色体上的从醋栗番茄基因渗入并赋予ToBRFV抗性的QTL9的保藏的种子NCIMB 43591或其子代长出的植物与缺乏所述QTL9的初始番茄植物杂交,其中,所述9号染色体上的QTL9存在于植物种子LVSTBRFVRES2、NCIMB保藏号43591的基因组中,可通过SNPTO-0201220的等位基因G、SEQ ID TO-0201221的等位基因G、TO-0201229的等位基因A、TO-0201231的等位基因C或TO-0201233的等位基因G识别。
本发明还涉及一种具有对ToBRFV抗性的番茄植物的育种方法,包括以下步骤:使从携带在11号染色体上的从醋栗番茄基因渗入并赋予ToBRFV抗性的QTL11的保藏的种子NCIMB 43591或其子代长出的植物与缺乏所述QTL11的初始番茄植物杂交,其中11号染色体上的所述QTL11存在于植物种子LVSTBRFVRES2、NCIMB保藏号43591的基因组中,可通过TO-0201237的等位基因G、TO-0201238的等位基因A、TO-0201239的等位基因A、TO-0201240的等位基因A、TO-020124的等位基因A、SL2.50ch11_9684449的等位基因CT、SL2.50ch11_9779896的等位基因AT、SL2.50ch11_9823405的等位基因C或SL2.50ch11_9924232的等位基因GT识别。
具体地,本发明还涉及用于生产具有ToBRFV抗性的植物的方法或过程,包括以下步骤:
a)将从包括赋予ToBRFV抗性的QTL9的保藏的种子NCIMB 43591或其子代生长的植物,和优选缺失所述QTL的初始番茄植物杂交,
b)在这样获得的子代中选择包含本发明的QTL9的一种植物;
c)任选地,步骤b)中获得的植物自花授粉一次或多次,并在子代中选择由此获得的对ToBRFV具有抗性的植物,无论是果实抗性、枝叶抗性还是两者都有。
可选地,该方法或过程可以包括代替步骤a)的以下步骤:
a1)使对应于包括赋予ToBRFV抗性的QTL9的保藏的种子(NCIMB 43591)或其子代生长的植物,和优选缺失所述QTL的初始番茄植物杂交,
a2)通过自交的方式增加F1杂种,以创造F2种群。
在上述方法或过程中,优选SNP标记物用于步骤b)和/或c)中,用于选择携带赋予目的抗性表型的序列的植物。
所述SNP标记物优选为具有SEQ ID NO:1至101的本发明的101个SNP标记物中的一个或多个,包括本申请别处提及的它们的所有组合,并且优选具有SEQ ID NO:1-14的SNP。
通过根据一个或多个SNP的等位基因选择植物,可以理解,当SNP的等位基因是对应于该SNP的LVSTBRFVRES2亲本的等位基因的等位基因而不是初始番茄植物的等位基因时,该植物被选择为相对于初始植物具有ToBRFV抗性,无论是果实耐受性/抗性,枝叶耐受性/抗性还是两者兼有。例如,当检测到SNP TO-0201220的等位基因G、TO-0201221的等位基因G、TO-0201222的等位基因A、TO-0201223的等位基因A、TO-0201224的等位基因A、TO-0201225的等位基因A、TO-0201226的等位基因A、TO-0201227的等位基因C、TO-0201228的等位基因C、TO-0201229的等位基因A、TO-0201230的等位基因C、TO-0201231的等位基因C、TO-0201232的等位基因G和/或TO-020133的等位基因G时,更优选当检测到SNP TO-0201220的等位基因G、SEQ ID TO-0201221的等位基因G、TO-0201229的等位基因A、TO-0201231的等位基因C、TO-0201232的等位基因G或TO-0201233的等位基因G时,甚至更优选当检测到SNPTO-0201220的等位基因G和/或TO-0201229的等位基因A时,可以选择具有本发明的改善表型的植物。
优选地,步骤a)的番茄植物是优良品系,用于获得具有商业所需性状或所需园艺性状的植物。有利地,由于Tm-2
如上所定义的方法或过程可以有利地包括回交步骤,优选在步骤c)之后,以获得具有番茄植物的所有表征特征的植物。因此,生产具有这些特征的植物的方法或过程还可以包括以下附加步骤:
d)将步骤b)或c)中选择的抗性植物与番茄植物回交;
e)选择携带本发明的QTL9或基因渗入序列的植物。
步骤a)中使用的植物,即对应于保藏的种子的植物可以是从保藏的种子生长的植物;其也可以是根据本发明第一方面的、携带QTL9或基因渗入序列、赋予表型,优选纯合地携带这些序列的任何植物。
优选这样的植物还包括所定义的QTL11,优选纯合地包括所定义的QTL11。
在步骤e)中,SNP标记物可用于选择相对于初始植物具有ToBRFV抗性的植物。SNP标记物是本发明的标记物,如前面各部分所述。根据优选的实施方式,本发明的方法或过程的进行使得,对于至少一个选择步骤,即b)、c)和/或e),选择是基于具有SEQ ID NO:1-101的SNP的至少一个抗性等位基因的检测进行的。优选的等位基因和组合已经公开并且适用于本发明的此实施方式。
需要注意的是,当选择具有改善的表型并纯合地携带赋予该表型的QTL的植物时,选择是基于本发明的一个或多个SNP,在代表QTL的等位基因,即等位基因LVSTBRFVRES2亲本的存在下,结合不存在代表复发易感番茄亲本的等位基因进行的。
也可以基于与基因渗入序列相关的任何其他标记物进行选择,并通过与易感亲本的常驻序列(resident sequences)相反来代表这些基因渗入序列的存在。定义替代标记物的方法也在本发明的范围内并在另一部分中公开。
在步骤e)中选择的植物优选为商业植物,特别是在正常培养条件下完全成熟时,果实重量至少为10g但优选25g,至少100g、至少150g或至少200g的植物。
优选地,步骤d)和e)重复至少两次,优选三次,不一定用相同的番茄植物。所述番茄植物优选为育种系。
在上述公开的每个过程的选择步骤中,可以另外选择对线虫性状具有抗性或对ToMV具有抗性。
自花授粉和回交步骤可以按任何顺序进行,并且可以插入,例如可以在一次或多次自花授粉之前和之后进行回交,并且可以设想在一次或多次回交之前和之后进行自花授粉。
也可以通过特别是在实施例中公开的方案,基于来自番茄亲本的ToBRFV抗性的比较选择具有所需改善表型的子代;测试的抗性/耐受性可以是果实抗性/耐受性或枝叶抗性/耐受性,或两者兼有。
用于等位基因检测的方法可以基于允许区分特定染色体上的SNP的两个不同等位基因的任何技术。
本发明还涉及相同的方法,其中在步骤a)中,使用从包含赋予ToBRFV抗性的QTL11的保藏种子NCIMB 43591或其子代生长的植物。然后针对QTL11进行所有检测/选择步骤,特别是具有SEQ ID NO:102-115的标记物,更优选基于TO-0201237的等位基因G、TO-0201238的等位基因A、TO-0201239的等位基因A、TO-0201240的等位基因A、TO-0201241的等位基因A、SL2.50ch11_9684449的等位基因CT、SL2.50ch11_9779896的等位基因AT、SL2.50ch11_9823405的等位基因C和/或SL2.50ch11_9924232的等位基因GT的存在;例如,通过TO-0201237的等位基因G、TO-0201238的等位基因A、TO-0201239的等位基因A、TO-0201240的等位基因A和TO-0201241的等位基因A中的至少一个的存在。标记物和抗性等位基因的替代优选列表已在上文中公开。
因此,本发明还涉及一种赋予番茄植物对ToBRFV的抗性的方法,包括以下步骤:
a)使从保藏种子NCIMB 43591或其子代生长的植物,与优选缺失所述QTL的初始番茄植物杂交,保藏种子NCIMB 43591或其子代携带从醋栗番茄基因渗入并在NCIMB 43591中独立赋予ToBRFV抗性的、在9号染色体上的QTL9和/或在11号染色体上的QTL11;
b)在由此获得的子代中选择携带QTL9和/或QTL11的植物;
c)任选地,在步骤b)中获得的植物自花授粉一次或多次,并在子代中选择由此获得的对ToBRFV具有抗性的植物。
根据另一种实施方式,本发明涉及一种赋予番茄植物对ToBRFV的抗性的方法,包括以下步骤:
a1)将从保藏的种子NCIMB 43591或其子代生长的植物与优选缺乏所述QTL的初始番茄植物杂交,从而产生F1种群,该保藏的种子NCIMB 43591或其子代携带从醋栗番茄基因渗入并独立赋予在NCIMB 43591中的ToBRFV抗性的在9号染色体上的QTL9和/或在11号染色体上的QTL11,,
a2)F1杂交种自花授粉,以产生F2种群,
b)选择由此获得的对ToBRFV具有抗性的子代个体。
SNP标记在步骤b)和/或c)中有利地用于选择携带独立赋予ToBRFV抗性的QTL9和/或QTL11的植物。
本发明还涉及一种获得商业番茄植物或其近交亲本系的方法,该商业番茄植物或其近交亲本系相对于初始商业番茄植物,具有对应于对番茄褐色皱果病毒具有果实和/或枝叶耐受性和/或抗性的所需的改善表型,所述方法包括以下步骤:
a)将通过保藏的种子LVSTBRFVRES2 NCIMB保藏号43591发芽或其后代获得的植物与商业番茄植物进行回交,该保藏的种子LVSTBRFVRES2NCIMB保藏号43591火气后代携带赋予ToBRFV抗性的QTL9,
b)选择携带本发明的QTL9的植物。
优选地,所述筛选是基于本发明的101个SNP中的一个或多个进行的,如本发明其它方法所详述的。
或者,步骤a)的子代是携带QTL11的子代,并且步骤b)的选择是基于QTL11,优选基于具有SEQ ID NO:102-115的本发明的14个标记物中的一个或多个,如本发明的其它方法所详述的。
根据本发明的所有方法和过程中,初始番茄植物是确定的、不确定的或半确定的。
如已经公开的,根据本发明的番茄植株优选还对番茄花叶病毒、线虫、TYLCV以及镰刀菌属和轮枝孢属具有抗性。为了在本发明的过程和方法中获得该植物,育种方案中使用的番茄亲本优选携带赋予对番茄花叶病毒、线虫、TYLCV以及镰刀菌属和轮枝孢属的抗性的序列;并且进行选择步骤以选择具有这些抗性序列,以及赋予本发明的改善表型的QTL的植物。
本发明还涉及通过以上公开的任何方法和过程获得或可获得的番茄植物和种子。这种植物确实是具有根据本发明第一方面的改善的表型的番茄植物。这种番茄的种子优选用单独或组合的活性物种如植物营养素、增强微生物或用于对种子和植物的环境进行消毒的产品进行包衣或造粒。这些物种和化学物质可以是促进植物生长的产品,例如激素,或增加其对环境应激的抗性,例如防御刺激剂,或稳定基质及其周围环境的pH值,或者其他营养素。
它们也可以是用于防止不利于幼苗生长的试剂的产品,包含本文中的病毒和病原微生物,例如通过接触、摄入或气体扩散起作用的杀真菌剂、杀细菌剂、杀线虫剂(hematicidal)、杀昆虫剂或除草剂产品;例如,它是任何合适的精油,例如百里香提取物。所有这些产品都加强了植物的抗性反应,和/或消毒或调节所述植物的环境。它们也可以是活的生物材料,例如非致病性微生物,例如至少一种真菌或细菌或病毒,必要时具有确保其生存能力的培养基;而这种微生物,例如假单胞菌属(pseudomonas)、芽孢杆菌属(bacillus)、木霉菌属(trichoderma)、螺旋聚孢霉属(clonostachys)、镰孢菌属(fusarium)、丝核菌属(rhizoctonia)等类型刺激植物的生长,或保护其免受病原体的侵害。
在所有前文的方法和过程中,携带负责ToBRFV抗性的QTL或基因渗入序列的植物的识别可以通过检测与抗性QTL9相关的SNP的至少一个等位基因来完成,可以与本发明的SNP的其他等位基因形式的缺失相结合,以便在需要时确认QTL的纯合状态。因此,识别纯合地携带本发明的QTL或基因渗入序列的植物将基于识别针对QTL9的至少一个具有SEQ IDNO:1-101的SNP的抗性等位基因,以及所述SNP的易感等位基因的缺失。例如,纯合地携带本发明的QTL9的植物的识别将基于SNP TO-0201220的等位基因G、TO-0201221的等位基因G、TO-0201222的等位基因A、TO-0201223的等位基因A、TO-0201224的等位基因A、TO-0201225的等位基因A、TO-0201226的等位基因A、TO-0201227的等位基因C、TO-0201228的等位基因C、TO-0201229的等位基因A,TO-0201230的等位基因C、TO-0201231的等位基因C、TO-0201232的等位基因G和/或TO-020133的等位基因G的识别,以及相应易感等位基因的缺少,即SNP TO-0201220的等位基因A、TO-0201221的等位基因A、TO-0201222的等位基因G、TO-0201223的等位基因G、TO-0201224的等位基因C、TO-0201225的等位基因G、TO-0201226的等位基因G、TO-0201227的等位基因A、TO-0201228的等位基因G、TO-0201229的等位基因G、TO-0201230的等位基因A、TO-0201231的等位基因A、TO-0201232的等位基因A和/或TO-0201233的等位基因A。
类似地,对纯合地携带本发明的QTL或基因渗入序列的植物的识别将基于针对QTL11的至少一个具有SEQ ID NO:102-115的标记物的抗性等位基因的识别,以及所述SNP的易感等位基因的缺失。例如,纯合地携带本发明的QTL11的植物的识别将基于TO-0201237的等位基因G、TO-0201238的等位基因A、TO-0201239的等位基因A、TO-0201240的等位基因A、TO-0201241的等位基因A、SL2.50ch11_9684449的等位基因CT、SL2.50ch11_9779896的等位基因AT、SL2.50ch11_9823405的等位基因C和/或SL2.50ch11_9924232的等位基因GT的识别,以及相应的易感等位基因的缺失,即TO-0201237的等位基因A、TO-0201238的等位基因T、TO-0201239的等位基因C、TO-0201240的等位基因C,TO-0201241的等位基因C、SL2.50ch11_9684449的等位基因C、SL2.50ch11_9779896的等位基因A、SL2.50ch11_9823405的等位基因T和SL2.50ch11_9924232的等位基因G。
本发明还涉及由本发明人在此提供的信息的用途,即存在于保藏种子LVSTBRFVRES2中的QTL9和QTL11的存在,并赋予番茄植物改善的表型,并公开与这些QTL或基因渗入序列相关的分子标记物。除其他外,这些知识尤其可用于精确定位QTL、定义它们的序列、识别包含赋予改良表型的QTL的番茄植物以及识别与这些QTL相关的进一步或替代标记物。这种进一步的标记物的特征在于它们的位置,即接近本发明公开的101个标记物,并且优选来自针对QTL9的具有SEQ ID NO:1-14的14个SNP,并通过它们与本发明揭示的ToBRFV抗性的关联。对于QTL11,适用的标记物是那些具有SEQ ID NO:102至115的标记物。
在这方面,本发明还涉及一种用于识别、检测和/或选择具有本发明的QTL9的番茄植物的方法,如在LVSTBRFVRES2(NCIMB保藏号43591)的种子基因组中发现的QTL9,所述QTL相对于缺失所述序列的相应植物赋予对ToBRFV的改善抗性,该方法包括检测表H的SNP标记物()的至少一个抗性等位基因,尤其是SNP TO-0201220的等位基因G、TO-0201221的等位基因G、TO-0201222的等位基因A、TO-0201223的等位基因A、TO-0201224的等位基因A、TO-0201225的等位基因A、TO-0201226的等位基因A、TO-0201227的等位基因C、TO-0201228的等位基因C、TO-0201229的等位基因A、TO-0201230的等位基因C、TO-0201231的等位基因C、TO-0201232的等位基因G和TO-020133的等位基因G之一,更优选SNP TO-0201220的等位基因G、SEQ ID TO-0201221的等位基因G、TO-0201229的等位基因A、TO-0201231的等位基因C和TO-0201233的等位基因G之一,例如待识别和/或选择的植物的遗传物质样本中SNP TO-0201220的等位基因G和TO-0201229的等位基因A之一。优选地,具有SEQ ID NO:1-101的SNP的抗性等位基因中的至少2个或3个或5个被检测。
本发明还涉及一种检测或选择的番茄植物的方法,该植物具有赋予ToBRFV抗性的QTL9,并且具有至少一个具有SEQ ID NO:1-101的SNP的抗性等位基因,特别是具有SEQ IDNO:1-14的那些,其中检测或选择是在ToBRFV感染的条件下进行的,包括在待测试的植物上接种ToBRFV,人工感染或自然感染。目的表型的存在是本发明的QTL9或基因渗入序列的存在的信息,特别是在包含携带本发明的QTL9的亲本的育种方案中。
本发明还涉及与在种子LVSTBRFVRES2的基因组中发现的方法相同的,用于识别、检测和/或选择具有本发明的QTL11的番茄植物的方法,相对于缺失所述序列的相应植物所述QTL赋予对ToBRFV的改善抗性,该方法包括在待识别和/或筛选的植物遗传物质样本中检测表K的标记物的至少一个抗性等位基因,尤其是TO-0201237的等位基因G、TO-0201238的等位基因A、TO-0201239的等位基因A、TO-0201240的等位基因A、TO-0201241的等位基因A、SL2.50ch11_9684449的等位基因CT、SL2.50ch11_9779896的等位基因AT、SL2.50ch11_9823405的等位基因C和SL2.50ch11_9924232的等位基因GT之一。优选地,检测具有SEQ IDNO:102-115的SNP的抗性等位基因中的至少2个或3个或5个,或9个标记物TO-0201237至TO-0201241和SL2.50ch11_9684449、SL2.50ch11_9779896、SL2.50ch11_9823405和SL2.50ch11_9924232中的2个、3个或4个。根据另一种实施方式,标记物SL2.50ch11_9684449、SL2.50ch11_9779896、SL2.50ch11_9823405和SL2.50ch11_9924232的抗性等位基因中的至少1个、2个或3个被检测。标记物和抗性等位基因的替代优选列表已在上文中公开。可以在ToBRFV感染的条件下进行检测或选择。目的表型的存在是纯和的本发明的QTL11或基因渗入序列存在的信息。
本发明还涉及一种检测和/或筛选番茄植物、特别是商业番茄植物的方法,该植物具有本发明的QTL,该方法包含在待选择的植物的遗传物质样本中检测至少一种上述抗性等位基因,即
-对于9号染色体上的QTL,检测SNP TO-0201220的等位基因G、TO-0201221的等位基因G、TO-0201222的等位基因A、TO-0201223的等位基因A、TO-0201224的等位基因A、TO-0201225的等位基因A、TO-0201226的等位基因A、TO-0201227的等位基因C、TO-0201228的等位基因C、TO-0201229的等位基因A、TO-0201230的等位基因C、TO-0201231的等位基因C、TO-0201232的等位基因G和TO-0201233的等位基因G,优选SNP TO-0201220的等位基因G、SEQID TO-0201221的等位基因G、TO-0201229的等位基因A、TO-0201231的等位基因C、TO-0201232的等位基因G或TO-0201233的等位基因G中的至少一个,或
-对于在11号染色体上的QTL,检测TO-0201237的等位基因G、TO-0201238的等位基因A、TO-0201239的等位基因A、TO-0201240的等位基因A、TO-0201241的等位基因A、SL2.50ch11_9684449的等位基因CT、SL2.50ch11_9779896的等位基因AT、SL2.50ch11_9823405的等位基因C和SL2.50ch11_9924232的等位基因GT,例如TO-0201237的等位基因G、TO-0201238的等位基因A、TO-0201239的等位基因A、TO-0201240的等位基因A或TO-0201241的等位基因A中的至少一个。
抗性等位基因的替代优选的列表已在上文公开。
该方法特别适用于具有LVSTBRFVRES2(NCIMB保藏号43591)作为包含赋予ToBRFV抗性的本发明的QTL的初始亲本或其子代的育种程序。
本发明还涉及一种检测和/或选择具有本发明的赋予ToBRFV抗性的QTL9和/或QTL11的番茄植物的方法,该方法基于揭示所述QTL的存在的任何分子标记物的检测。事实上,目前本发明的QTL9和QTL11已经被本发明人识别出来,识别和之后分子标记物的使用,除了本发明的101个SNP(SEQ ID NO:1至101)或14个标记物(SEQ ID NO:102至115)之外,可以容易地被本领域技术人员实现。QTL9的特征可以是:存在本发明的101个SNP中的至少一个,但其也可以通过使用不同的、替代的标记物来识别;这同样适用于QTL11。因此,在本发明在还包括用于识别番茄基因组中本发明的QTL的任何此类分子标记物的方法和用途,其中相对于缺失所述QTL的相应植物,所述QTL赋予对ToBRFV的抗性,该QTL的特征在于:存在具有SEQ ID NO:1至101,优选1至14的SNP的至少一个抗性等位基因。
还包括用于识别番茄基因组中本发明的QTL9的任何此类替代分子标记物的方法和用途,其中所述QTL赋予ToBRFV抗性,其中所述QTL的特征在于:存在具有SEQ ID NO:1至101的SNP的至少一个抗性等位基因,优选SNP TO-0201220的等位基因G、TO-0201221的等位基因G、TO-0201222的等位基因A、TO-0201223的等位基因A、TO-0201224的等位基因A、TO-0201225的等位基因A、TO-0201226的等位基因A、TO-0201227的等位基因C、TO-0201228的等位基因C、TO-0201229的等位基因A、TO-0201230的等位基因C、TO-0201231的等位基因C、TO-0201232的等位基因G和TO-020133的等位基因G之一,更优选SNP TO-0201220的等位基因G、SEQ ID TO-0201221的等位基因G、TO-0201229的等位基因A、TO-0201231的等位基因C和TO-0201233的等位基因G之一,甚至更优选SNP TO-0201220的等位基因G和TO-0201229的等位基因A之一。
还包括用于识别番茄基因组中本发明的QTL11的任何此类替代分子标记物的方法和用途,其中当纯合地存在时所述QTL赋予ToBRFV抗性,其中所述QTL的特征在于:存在具有SEQ ID NO:102至115的标记物的至少一个抗性等位基因,优选具有SEQ ID NO:102至111的SNP的至少一个抗性等位基因,更优选TO-0201237的等位基因G、TO-0201238的等位基因A、TO-0201239的等位基因A、TO-0201240的等位基因A、TO-0201241的等位基因A、SL2.50ch11_9684449的等位基因CT、SL2.50ch11_9779896的等位基因AT、SL2.50ch11_9823405的等位基因C和SL2.50ch11_9924232的等位基因GT之一;例如TO-0201237的等位基因G、TO-0201238的等位基因A,TO-0201239的等位基因A、TO-0201240的等位基因A和TO-0201241的等位基因A之一。抗性等位基因的替代优选列表已在上文公开。
本发明还涉及一种检测和/或选择具有前文定义的赋予ToBRFV抗性的抗性QTL的番茄植物的方法,所述方法包括:
a)测定番茄植物是否存在至少一个与涉及ToBRFV抗性,尤其是在番茄植物中赋予所述抗性的QTL9或QTL11遗传有关或相关的遗传标记物,
b)选择包含遗传标记物和参与ToBRFV抗性的有关或相关的QTL9或QTL11的植物,
其中,对于QTL9,QTL和遗传标记物发现于由TO-0201220和具有SEQ ID NO:101的SNP限定的基因组区域中,优选在番茄基因组中由TO-0201220和TO-0201233限定的区域,和对于QTL11,在由具有SEQ ID NO:102和115的标记物限定的基因组区域,优选在由具有SEQID NO:102和111的标记物限定的区域。
通过关联或遗传关联,更具体地说是遗传连锁,应该理解遗传标记物(例如SNP标记物的特定等位基因)的多态性和目的表型同时发生,即是共同遗传,比偶然发生的预期更频繁,即等位基因和负责表型的基因序列之间存在非随机关联,由于它们的基因组靠近。
遗传标记物是上文公开的QTL9的101个标记物之一或替代标记物,并且优选在超过90%的减数分裂中,优选在超过95%、96%、98%或99%的减数分裂中与目的表型一起遗传。这同样适用于QTL11。
本发明的QTL或基因渗入序列的定义和优选特征如本说明书其他部分中所定义。赋予ToBRFV抗性的QTL如在种子LVSTBRFVRES2的基因组中发现的是有利的。
因此,本发明涉及一个或多个分子或遗传标记物用于精细定位或识别番茄基因组中赋予本发明的ToBRFV抗性的QTL的用途,其中所述一个或多个标记位于以下染色体区域之一:
-在由SNP TO-0201220(SEQ ID NO:1)和具有SEQ ID NO:101的SNP在9号染色体上限定的染色体区域,
-在距本发明的101个SNP标记物之一的基因座小于2兆碱基单位处,优选距具有SEQ ID NO:1至14的SNP之一的基因座,甚至更优选距TO-0201220、TO-0201221、TO-0201229、TO-0201231或TO-0201233之一的基因座小于2兆碱基单位处。
根据优选的实施方案,如上所公开的,所述一个或多个标记物在由TO-0201210和具有SEQ ID NO:101的SNP限定的染色体区域中,或由TO-0201210和TO-0201233,或由TO-0201221和TO-0201233限定的染色体区域中。
此外,所述一个或多个分子或遗传标记物优选地,与0.05或更小的p值,与具有QEDID NO:1至101的SNP的以下抗性等位基因中的至少一种相关,例如与SNP TO-0201220的等位基因G、TO-0201221的等位基因G、TO-0201222的等位基因A、TO-0201223的等位基因A、TO-0201224的等位基因A、TO-0201225的等位基因A、TO-0201226的等位基因A、TO-0201227的等位基因C、TO-0201228的等位基因C、TO-0201229的等位基因A、TO-0201230的等位基因C、TO-0201231的等位基因C、TO-0201232的等位基因G和/或TO-020133的等位基因G相关。
所述分子或遗传标记物优选为SNP标记物。更优选地在距离本发明的101个SNP中的至少一个的基因座小于1兆碱基,优选小于0.5兆碱基。
p值优选小于0.01。
此外,本发明涉及本发明的101个SNP标记物中的至少一个,与赋予ToBRFV抗性的9号染色体上的QTL相关联,用于识别与所述QTL关联的一个或多个替代分子或遗传标记物的用途,其中所述一个或多个替代分子或遗传标记物是:
-在由SNP TO-0201220(SEQ ID NO:1)和具有SEQ ID NO:101的SNP在9号染色体上限定的染色体区域,
-在距离本发明的101个SNP标记物之一的基因座小于2兆碱基单位,优选距离具有SEQ ID NO:1至14的SNP之一的基因座,甚至更优选距离TO-0201220、TO-0201221、TO-0201229、TO-0201231或TO-0201233之一的基因座小于2兆碱基单位。
根据优选的实施方式,所述替代标记物在上述优选的染色体区域中。遗传关联或连锁可以有利地通过跟踪替代标记物和由包含目的QTL的植物产生的子代中QTL的存在来检测。
所述替代分子标记物优选与所述QTL关联,p值为0.05或更小,优选小于0.01。QTL优选在保藏种子NCIMB 43591的基因组中发现。
分子或遗传标记物和抗性表型是在优选大于90%的减数分裂,优选大于95%的减数分裂中共同遗传。
根据本发明此方面的分子或遗传标记物优选为SNP。它们更优选距离本发明的101个SNP中的至少一个的基因座小于1兆碱基,优选小于0.5兆碱基。
本发明还涉及相同的方法和用途,其中标记物位于:
-在由具有SEQ ID NO:102的标记物和具有SEQ ID NO:115的标记物在11号染色体上限定的染色体区域,
-在距离表K的14个标记物之一的基因座小于2兆碱基单位,优选距离TO-0201237、TO-0201238、TO-0201239、TO-0201240、TO-0201241、SL2.50ch11_9684449、SL2.50ch11_9779896、SL2.50ch11_9823405或SL2.50ch11_9924232之一的基因座小于2兆碱基单位。
类似地,本发明还包括一种用于识别与赋予番茄植物对ToBRFV抗性的QTL相关的分子或遗传标记物的方法,如本申请中所述,所述方法包括以下步骤:
-在由TO-0201220和TO-0201233限定的基因组区间中或在距离本发明的101个SNP之一的基因座小于2兆碱基单位,优选小于0.5兆碱基单位识别分子或遗传标记物;和
-确定所述分子或遗传标记物的等位基因或状态是否与显示ToBRFV抗性的植物发出的分离种群中,例如在与保藏种子相对应的植物发出的分离种群中的ToBRFV抗性表型关联或相关。
根据又另一方面,本发明还涉及针对与ToBRFV感染的抗性或耐受性相关的至少一种遗传标记物的存在对植物,优选番茄植物或番茄种质进行基因分型的方法,其中该方法包括在被测试植物的基因组中测定或检测包含本发明的101个标记物中的至少一种或包含如上公开的替代分子标记物中的至少一种的核酸。优选地,该方法包括在待测试的植物样品中识别在包含本发明的SNP的至少一个抗性等位基因的核酸中与ToBRFV抗性相关的特定序列的步骤。
根据该方法的最优选实施方式,该方法包括在被测试植物中检测包含SNP TO-0201220的等位基因G或SNP TO-0201229的等位基因A的核酸的存在。
本发明还涉及与QTL11相关以及关于由具有SEQ ID NO:102的标记物和具有SEQID NO:115的标记物在11号染色体上限定的染色体区域的相同方法。该区域的相关标记物或SNP已经在本发明中公开,以及优选列表。
鉴于本发明的抗性植物限制由ToBRFV感染造成的损害的能力,它们有利地生长在被ToBRFV感染或可能被感染或被侵染的环境中;在这些条件下,本发明的抗性或耐受性植物比易感植物产生更适销的西红柿。因此,本发明还涉及一种在ToBRFV感染的环境中改善番茄植物产量的方法,包括生长其基因组中包含在9号染色体上的QTL9和/或在11号染色体上的QTL11的番茄植物,如根据本发明的前述方面定义的,并赋予所述植物对ToBRFV的抗性。
优选地,该方法包括第一步,选择或筛选包含所述QTL或目的基因渗入序列的番茄植物。该方法也可以定义为提高番茄田、隧道或温室生产力的方法,或定义为减少番茄生产中化学或杀真菌剂应用的强度或数量的方法。
本发明还涉及一种在ToBRFV侵染或感染条件下减少番茄生产损失的方法,包括生长如上所定义的番茄植物。
这些方法对于田间、隧道或温室中的番茄植物种群特别有价值。
或者,所述改善产量或减少番茄生产损失的方法可以包括:第一步,即识别对ToBRFV有抗性/耐受性的番茄植物,并在其基因组中包含赋予所述植物ToBRFV抗性的本发明的QTL9和/或QTL11;然后在受病毒感染或可能被病毒感染的环境中生长所述抗性植物。根据优选的实施方式,在第一步中待识别的植物包含TO-0201229的等位基因A,或具有SEQID NO:1至101的SNP的至少一个抗性等位基因。
本发明的抗性植物还能够限制ToBRFV的生长,从而限制其他植物的感染和病毒的传播。因此,本发明还涉及保护田地、隧道或温室或任何其他类型的种植园免受ToBRFV感染的方法,或至少限制所述田地、隧道或温室的被ToBRFV感染的水平,或限制ToBRFV在田间、隧道或温室中的传播,特别是在番茄田中的传播的方法。这种方法优选包括生长本发明的抗性或耐受性植物的步骤,即在其基因组中包含赋予所述植物ToBRFV抗性的9号染色体上的QTL9的植物。待使用的本发明的植物优选包含TO-0201229的等位基因A,或具有SEQ IDNO:1至101的SNP的至少一个抗性等位基因。根据另一种实施方式,待使用的植物包含具有SEQ ID NO:102至115的标记物的至少一个抗性等位基因。
本发明还涉及ToBRFV抗性植物用于控制田间、隧道或温室或其他种植园中ToBRFV感染或侵扰的用途;这种植物是本发明的植物,在其基因组中包含QTL9和/或QTL11,或如上所定义的9号或11号染色体上的来自醋栗番茄的基因渗入的序列。这种用途或方法也是一种通过减少病毒种群来对田地、隧道或温室进行消毒的方法。
QTL的所有优选特征如结合本发明的其他方面所定义,即优选存在于LVSTBRFVRES2(NCIMB保藏号43591)的种子中,并且可通过针对QTL9的具有SEQ ID NO:1至101的SNP标记物来识别;优选具有SEQ ID NO:1至14的SNP标记物来识别,优选通过SNP TO-0201220的等位基因G、TO-0201221的等位基因G、TO-0201222的等位基因A、TO-0201223的等位基因A、TO-0201224的等位基因A、TO-0201225的等位基因A、TO-0201226的等位基因A、TO-0201227的等位基因C、TO-0201228的等位基因C、TO-0201229的等位基因A、TO-0201230的等位基因C、TO-0201231的等位基因C、TO-0202132的等位基因G和/或TO-020133的等位基因G识别,并且可通过针对QTL11的具有SEQ ID NO:102至115的标记物来识别,优选针对QTL11的TO-0201237的等位基因G、TO-0201238的等位基因A、TO-0201239的等位基因A、TO-0201240的等位基因A、TO-0201241的等位基因A、SL2.50ch11_9684449的等位基因CT、SL2.50ch11_9779896的等位基因AT、SL2.50ch11_9823405的等位基因C和/或SL2.50ch11_9924232的等位基因GT来识别。
在进一步的方面,本发明还涉及一种生产番茄的方法,包括:
a)生长包含如前所定义的QTL9和/或QTL11的发明的番茄植物;
b)允许所述植物结果实;和
c)收获所述植物的果实,优选在成熟时和/或成熟前。
关于QTL9和QTL11的所有优选实施方式已在本发明的前述方面的上下文中公开。该方法可有利地包括将所述番茄加工成番茄加工食品的进一步步骤。
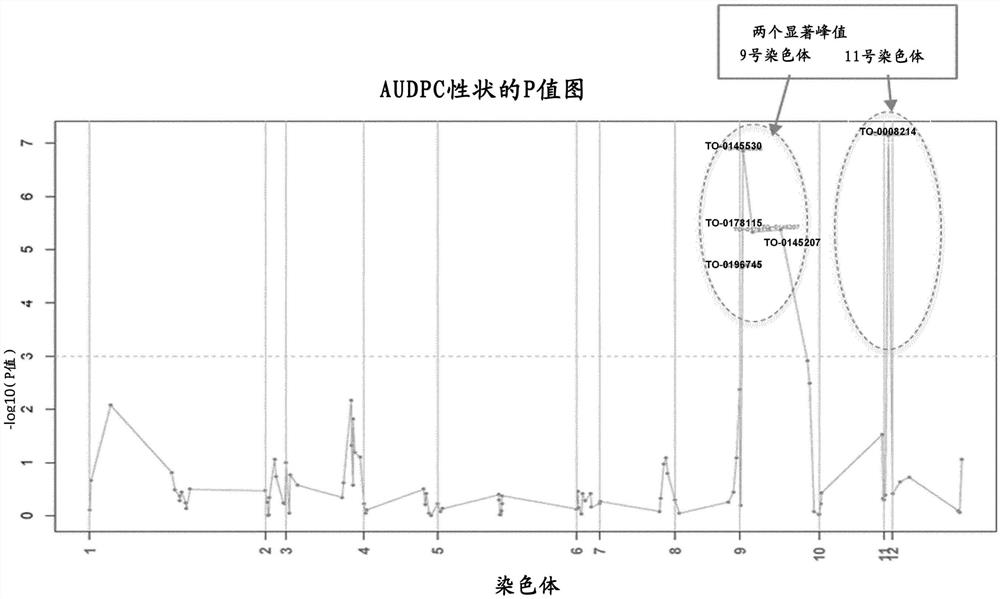
附图说明
图1:基于F2群体(基于来源D和HMC1),对应于AUDPC的性状的与ToBRFV抗性相关的QTL的p值图,。
纵轴(y轴)显示—log10(p值)和水平轴(x轴)表示通过沿物理图谱的染色体的SNP位置(以物理距离bp)的所有SNP。
图2A:果实抗性的调整值:Turkey检验图示
图2B:根据QTL9处抗性亲本等位基因的存在,果实抗性调整值和置信区间
图3:不同基因型的果实抗性调整值
图4:通过QTL9基因型的果实评分重新分配
图5:具有置信区间的叶片抗性调整值
图6:具有置信区间的不同基因型的枝叶抗性调整值
实施例
该测试进行3次重复,每个测试的种质或遗传背景大约15株植物。播种和在2叶期感染植物。然后在感染后7、14和28天通过对叶片的视觉评估进行评分。
对于此测试,标准如下:
9:无症状
5:中度花叶或/和坏死症状
1:强烈的花叶或/和坏死症状
在32天dpi时,对4个种质逐个植物进行Elisa测试,并对两个易感对照进行批量测试。
播种和在2叶期感染植物。然后在感染后7、14和28天通过对叶片的视觉评估进行评分。
对于此测试,标准如下:
9:无症状
7:轻度花叶症状
5:中度花叶症状
3:强烈的花叶症状和/或鼓泡
1:非常强烈的花叶症状和/或鼓泡和/或变形
对于QTL分析,发明人使用了不同的表型变量/性状,特别是14dpi的注释,21dpi的注释,28dpi的注释,AUDPC。
AUDPC(疾病进展曲线下面积)使用以下公式计算:
其中,“n”是症状评估的次数,“y”是症状强度(1至9),“t”是以dpi(接种后天数)计的时间。
根据制造商的程序,使用
接种阶段:种植后10天和17天。植物因此被感染两次。每次接种最嫩的2片叶片。最后,接种4片不同的叶片。
分离:ToBRFV约旦当地菌株2017。ToBRFV感染的嫩叶取自自然感染的植物,来自对TYLCV(Ty)和TMV具有抗性的植物,以避免接种物中存在多种病毒。
通过研磨制备4mL接种物需要1g具有ToBRFV症状的幼叶。
接种前,使用TMV免疫试纸(该免疫试纸不具有特异性,也可识别ToBRFV)检查接种物中ToBRFV的存在,并使用PepMV免疫试纸检查PepMV(Pepino)的缺失。只要受感染的植物具有TMV抗性,TMV免疫试纸就可以检测接种物中ToBRFV的存在。
待应用的接种物在TMV免疫试纸上呈阳性,在PepMV免疫试纸上呈阴性。
通过用浸泡在接种物中的粗糙海绵轻轻摩擦此叶片,将接种物应用到待测植物的两片嫩叶上。
当第一串果实是红色的,并且第二串果实变红时,进行第一次评估。
当至少第3串为红色时,进行第2次评估。
叶片症状的标准:
9:无症状,/7:在少数叶片弱症状,/5:在部分叶片上中等症状,/3:在所有叶片上中度症状,/1:在所有叶片上强烈症状。
果实症状的标准:
9:所有果实都没有任何症状
7:在一个或几个果实上轻微症状(变色)
5:在至少2-3个果实上轻度/中度果实变色
3:在超过30%的果实上中度/强烈的果实变色和/或小果实变形。
1:在超过50%的果实上非常强烈的变色和/或中度/强烈的果实变形和/或果实坏死斑点。
匍柄霉属是一种植物病原体真菌;它是番茄灰斑病的致病性病原体。来自醋栗番茄(Lycopersicum pimpinellifolium)的Sm基因提供了对匍柄霉属的遗传显性抗性。
匍柄霉属,西西里菌株,储存在-80℃。在培养皿上用V8培养基培养后,直接从冷冻保存的试管中制备接种物。分生孢子是通过刮擦培养基表面获得的,悬浮在含有1%葡萄糖的水中,然后在平纹细布上过滤。获得每毫升包含10
待测试的幼苗处于3片展开叶的阶段,对应于播种后17至24天。通过喷洒在所有叶片表面上应用接种物,直到形成液滴。
症状评估:
然后在感染后7至8天通过对叶片的视觉评估进行评分。
对于此测试,标准如下:
9:无症状
7:一些棕色坏死病变,数量少于易感植物
1:叶片两面有或小或大的棕色坏死病变。
发明人筛选了超过500种不同的野生种质以识别对ToBRFV抗性的潜在来源,从而潜在地将这些野生种质中赋予抗性的序列基因渗入到番茄背景中,特别是在商业植物中。
尽管发明人预期的抗性类型是果实耐受性/抗性,但这种测试不能应用于具有不同果实大小和形式的野生种质。因此,发明人决定在叶片症状测试中对来源进行排序,作为果实抗性的替代指标,因为无法筛选该参数。
在筛选的野生种质中,在多毛番茄(S.habrochaites)、智利番茄(S.chilense)和醋栗番茄物种中仅识别出4个潜在来源(0.9%),这表明与其他病毒相反,很难找到对ToBRFV的抗性来源。
所有这些来源都有很高比例的无症状植物,然而这些植物通过ELISA测试全部是阳性,表明没有发现总抗性或免疫力。
对于对照,使用了已知对ToBRFV易感的两种植物S1和S2;然而,由于Tm-2
评估植物叶片的潜在抗性/耐受性;事实上,鉴于这些野生种质的果实形状、颜色、大小不同,无法对果实水平上的抗性进行排名。
表A报告了获得的结果:
然而,在32dpi之后,所有植物通过ELISA检测均为阳性,表明这些植物携带的抗性并非全部(表A)。
鉴于获得的结果,发明人决定将进一步的工作集中在来源D上,它显示了在28dpi时叶片抗性的更好水平,尽管该研究的目的是识别果实抗性来源而不是叶片抗性来源。
F2定位-HMC1*来源D-人工测试
通过将抗性野生来源D与易感亲本HMC1杂交,开发了四个F2群体,每个群体有240个个体。HMC1是育种品系,生长习性不确定,红色圆形果实约100g,它包含Tm-2
视觉评分在不同的日期进行:7、14和28dpi(见上表B)。
表B显示了人工条件下F2筛选结果:植物以1至9的等级评分,其中得分为“1”或“3”的植物将被视为易感,得分为“5”的植物将被视为中等抗性,得分为“7”或“9”的植物在叶抗性方面具有高度抗性。
得分为1、3、5、7和9的植物数量报告在第4-8列。第9列(1=S)表示得分为1、3、5或7的植物数量。第10列(9=R)表示得分为9的植物数量。第11列和第12列报告得分为“S”(得分为1、3、5或7)和得分为“R”(得分为9)的植物的百分比。
表B:来源D及其子代的评分
QTL分析
如实施例1中详述提取DNA。对于QTL分析,发明人使用了不同的表型变量/性状:注释14dpi,注释21dpi,注释28dpi,AUDPC。
AUDPC(疾病进展曲线下面积)使用以下公式计算:
其中,“n”是症状评估的次数,“y”是症状强度(1至9),“t”是以dpi(接种后天数)计的时间。
F2群体的基因分型(基于来源D和HMC1)是使用一组169个SNP完成的。这些SNP是根据以下选择的:
·多态性/等位基因频率
·SNP物理图谱距离均匀分布
QTL分析是使用MAST-标记物辅助选择工具(A Marker Assisted SelectionTool,专有软件)中的双亲群体的QTL检测(ANOVA)模型进行的。
定位结果(见图1,说明对应于AUDPC的性状的P值图)揭示了与ToBRFV抗性相关的两个QTL候选物,它们位于9号染色体(QTL9)和11号染色体(QTL11)上,阳性等位基因来自来源D。
表C总结了与ToBRFV抗性相关的QTL9和QTL11显著相关的标记物及其在番茄基因组中的位置。
结果显示,负责ToBRFV抗性的QTL9位于番茄基因组的SL2.50版本的9号染色体上的位置3987296和位置40039587之间。9号染色体的这个区域是已知重组率低的区域。
负责ToBRFV抗性的QTL11位于11号染色体上的位置4524671,也是基于番茄基因组的SL2.50版本。
来源D和易感亲本HMC1之间的BC1F2种群已使用表C中所述的SNP开发。158株个体植物已经在约旦的田间接种条件下进行了表型分型,如实施例1中所述。
果实的症状以9到1的标准进行评估:
9:无症状-7:少数果实轻微症状-5:部分果实中等症状-3:所有果实中等症状-1:强烈症状。
表D显示了在田间接种条件下BC1F2筛选的结果:植物以1至9分的标准评分,其中1或3分的植物将被视为易感,5分的植物将被视为中等抗性,7或9分的植物将被视为高度抗性。由于果实大小,即由于醋栗番茄来源的小果实,没有对来源D进行果实评估。
表D:HMC1和BC1F2群体的果实评分
如实施例1所述从叶片中提取DNA。
BC1F2群体用在F2定位群体中与ToBRFV抗性显著关联的SNP亚群进行基因分型。标记物性状关联是通过MAST-标记物辅助选择工具(专有软件)中的交叉方差分析完成的。
定位结果显示,QTL9与对ToBRFV的果实抗性有关;即已经与叶片抗性相关的同一区域。
BC3F2 QTL定位-田间测试
在接种的田间条件下,对来源D和HMC1之间各约140个个体的3个BC3F2群体进行了评估。接种方案和症状评估与用于BC1F2和实施例1中描述的方案相同。
这些植物的评分详见表E;得分1到9的标准与BC1F2的详细情况相同。
表E:HMC1和BC3F2群体的果实评分
如实施例1所述从叶片中提取DNA。
BC3F2个体用在9号和11号染色体上的被识别为与F2定位群体中对ToBRFV抗性关联的多态性SNP的亚群进行基因分型。
使用JoinMap软件构建9号和11号染色体的遗传图谱,以确认标记物的位置和顺序。
QTL检测是使用MapQTL软件和对这些家族完成的遗传图谱进行的。
QTL定位结果证实了9号染色体上存在对ToBRFV的果实抗性的主要QTL。峰相关标记物在表F中描述。
表F:基于BC3F2分析的峰相关SNP
实施例4:品系重新测序和唯一SNP的识别。
对6个番茄品系(包括3个ToBRFV耐受性/抗性来源,包括来源D和WO2018/219941(HAZTBRFVRES1)中使用的品系)和在定位群体中用作复现的3个易感品系进行了重新测序。
播下种子,从新鲜叶片中提取DNA,并进行全基因组测序。
设定的测序深度最小为20X。使用lllumina NovaSeq 2x150nt技术进行测序。
将读数定位到SL2.40番茄参考基因组上,并用samtools进行变体调取分析(calling analysis)。为了尽可能唯一地识别来源D的SNP,过滤SNP,并将来源D中发现的等位基因与项目其他品系和360个番茄基因组中识别的SNP等位基因进行比较(“Genomicanalyses provide insights into the history of tomato breeding”;Lin etal.Nature Genetics,2014)。
在此基础上,在包含QTL9(从SL2.40ch09:10Mb到55Mb)的区间内识别来自来源D的310个唯一SNP。为了选择最好的SNP,应用了一些质量过滤器(番茄基因组上独特的Blast和侧翼序列中的%AT)。选择了101个SNP的列表(见表H)。
14个SNP(见表G)已经在大量背景下进行了测试,并且它们已经确认它们追踪QTL9的存在的能力。其中5个SNP(见表G)在对来源D的特异性方面显示出非常好的结果。
表G:QTL9间隔中来源D重新测序的14个特异性SNP的列表。
该表给出了14个SNP的名称,在SL2.50基因组中的位置,括号中具有多态性的序列以及易感和抗性等位基因。显示非常好的结果的5个SNP用星号表示。
多态性在括号内表示。“S”列报告易感等位基因,复现亲本HMC的存在,而“R”列表示抗性等位基因,如来源D所示。
表H:QTL9间隔中来源D重新测序的101个特异性SNP的列表。
表H给出了本发明人识别的101个SNP在SL2.50基因组中的位置、序列表中序列的SEQ ID号以及易感和抗性等位基因。上面提到的14个SNP和5个SNP显示非常好的结果,分别用一个或两个星号表示。
表H.QTL9间隔中来自LVSTBRFVRES2重新测序的101个特异性SNP的列表
因此,这些SNP允许区分具有本发明所述的QTL9的植物,和WO2018/219941中所述的源自HAZTBRFVRES1的植物,所述源自HAZTBRFVRES1的植物在9号染色体上具有提供对ToBRFV的耐受性的不同的QTL。因此,本发明中公开的QTL与WO2018/219941中公开的QTL在序列上明显不同。
实施例5:田间试验中新QTL9的表征。
在约旦进行了两项不同的试验,以确认除了本发明的QTL9和WO2018/219941中提到的QTL的序列差异外,这些不同的序列还赋予不同类型的ToBRFV抗性。
第一个试验T1(372株植物)于夏季在一个隧道中,使用不同的优良品系(检查)、实施例2-4中提到的对照S1和S2以及包含WO2018/219941中提到的9号和11号染色体上的QTL的植物进行。
第二次试验T2(1165株植物)在接下来的冬天,使用两条隧道,包括与先前试验T1相同的优良品系(检查)和对照,以及从来源D产生的BC3F2进行,如实施例4所述。
在此实施例中,在测试植物的6号和9号染色体上的QTL9和QTL11的基因座发现的序列的类型/起源定义如下:
→在QTL9和QTL11基因座的优良品系中发现,以及优良品系中这些基因座的扩展“等位基因”的序列,被编码为“Re”;
→在QTL9和QTL11基因座的易感品系S1和S2中发现的序列(易感品系中这些基因座的“等位基因”)编码为“S”;
→源自WO2018/219941中描述的HAZTBRFVRES1的耐受性/抗性植物中发现的位于QTL9和QTL11基因座(HAZTBRFVRES1中这些基因座的“等位基因”)的序列编码为“Rh”;
→在来自来源D的抗性植物BC3F2中发现的位于QTL9和QTL11基因座(本发明植物中这些基因座的“等位基因”)的序列被编码为“Rd”。
将植物接种两次,第一次在种植一周后,第二次在两周后,如实施例1所述。
然后根据实施例1.4中提到的标准,关于果实和叶片症状对植物进行评分。就叶片症状在成年阶段(植物结出红色果实)进行评估,然而,鉴于稀有嫩叶,叶片症状很难评估。
结果的统计分析。
使用混合模型分析数据,以确定由在植物的QTL9基因座发现的“等位基因”赋予的抗性水平,同时考虑到其他潜在影响,即在QTL11基因座发现的“等位基因”,植物的基因型和隧道的影响。混合模型如下:
分数=μ+(基因型)随机+隧道+QTL9+QTL11+ε
其中,分数(果实抗性或叶片抗性)是观测变量,μ是性状的平均值,ε是残差,其中(基因型)随机、隧道、QTL9和QTL11是效应。
由于所有隧道中都存在检查,因此估计隧道效应。
基因型效应被处理为随机效应,以捕捉群体的变异性,测试品种来自该群体。
对于QTL9基因座的每个基因型和每个性状(果实症状和叶片症状),发明人提取了调整后的值,以便独立于其他效应更好地估计每个基因型的潜力。
例如,对于与Rd/Rd对应的基因型,调整后的值为:
μ+QTL9[RdRd]+平均(基因型)+平均(隧道)+平均(QTL11)
其中μ是性状(果实症状或叶片症状)的估计平均值,QTL9[Rd/Rd]是对QTL9的基因型RdRd的估计效应,平均(基因型)、平均(隧道)和平均(QTL11)是相应的估计效应的平均值。
Tukey检验用于在调整值之间进行多重比较
结果:
QTL9对果实症状的影响:
对于与果实症状相对应的性状,使用上面详述的混合模型计算不同测试基因型关于QTL9基因座的调整值。结果在表I中报告,并在图2A和2B中示出。
表I:QTL9对果实症状的影响
N=相应植物的数量
组别:根据Tukey检验分配的组
对于QTL9基因座的序列,Rd、Rh、Re和S对应于植物的基因型。
从这些结果可以推断,本发明中定义的QTL9的存在(对应于此实施例中的Rd基因型)提供的抗性水平显著不同于且显著高于WO2018/219941中定义的QTL2提供的抗性水平(对应于此实施例中的Rh基因型)。
此外,对QTL9处的等位基因对果实抗性的影响的分析表明Rd等位基因具有显著的加性效应,如下(表I的摘录)和图3所示。
果实评分重新分配也已由QTL9基因型确定。结果如图4所示。
如这些结果所示,根据本发明的QTL9以纯合状态(此实施例中为基因型RdRd)时,给出超过80%的植物的果实评分为5、7或9(分别为208株中的48、61和59株)。大约60%的植物(208株中的120株)在两轮感染后得分为7或9分,即果实几乎没有症状。
这些结果也证实根据本发明的QTL9,以杂合状态(在此实施例中为基因型RdS)存在时,给出约50%的植物的果实评分为5、7或9(225株中分别为63、36和13株)。大约22%的植物(225株中的49株)在两轮感染后得分为7或9分,即果实几乎没有症状。
相比之下,对于RhRh基因型,也大约80%的植物的果实评分为5、7或9,但小于35%的植物的得分为7或9(89株中分别为30和1株),即在两轮感染后,果实几乎没有症状。
QTL9对叶片症状的影响:
对于与叶片症状相对应的性状,使用上文详述的混合模型计算不同测试基因型关于QTL9基因座的调整值。结果在表J中报告,并在图5中示出。
表J:QTL9对叶片症状的影响
N=相应植物的数量
组别:根据Tukey检验分配的组别
对于QTL9基因座的序列,Rd,Rh,Re和S对应于植物的基因型。从这些结果可以推断,本发明所定义的QTL9(对应于此实施例中的Rd基因型)的存在提供的叶片抗性水平显著不同于且显著高于WO2018/219941中定义的由QTL2提供的叶片抗性水平(对应于此实施例中的Rh基因型)。
此外,结果还表明,QTL9杂合地存在足以提供高水平的叶片抗性(基因型RdRd和RdS的调整值是统计学上相同的)。
实施例6:11号染色体上唯一SNP的品系重新测序和识别。
实施例4中对于9号染色体上的QTL公开的相同实验应用于发明人识别的11号染色体上的QTL。SNP分析是对相同植物群体进行的。
SNP是关于根据本发明的QTL11的存在的信息,该SNP在下面的表K中报告(SEQ IDNO:102-111),以及它们在SL2.50版本的基因组中的位置以及易感和抗性等位基因。
然后,发明人进行进一步的搜索,以识别对于赋予ToBRFV抗性的基因渗入序列的存在的其他标记物。这些标记物对来自赋予ToBRFV抗性的来源D的基因渗入序列具有非常的特异性。
表K:与QTL11对应的标记物
实施例7:通过甲烷磺酸乙酯(EMS)对番茄种子的基因进行修饰
番茄品种的种子用EMS通过在室温下,将每个品种约2000粒种子浸入0.5%(w/v)或0.7%EMS的充气溶液中,保持24小时进行处理。
每个品种每剂EMS大约1500粒处理过的种子发芽,所得植物优选在温室中生长,例如从5月到9月,以生产种子。
成熟后,每次处理将M2种子收获并成批地装在一个池中。所得的M2种子池用作起始材料,以识别单个M2种子和对番茄褐色皱纹果实病毒具有抗性的植物。
实施例8:对匍柄霉属的抗性
W02020/018783公开了番茄11号染色体上的遗传区域,该区域包含来自醋栗番茄的匍柄霉属抗性等位基因,据称还包含TBRFV抗性等位基因,它们两者之间的联系如此紧密,以至于它们被表征为非常相同的标记物,因此同时基因渗入。为了确认本发明人识别的11号染色体上的QTL与W02020/018783中公开的抗性等位基因不同,本发明人对根据本发明的植物的匍柄霉属抗性进行了测试。
匍柄霉属抗性的方案如实施例1.5中公开的。
结果:
测试了来自5种不同基因型的超过157种不同植物,每个基因型至少18个重复。
测试的基因型/栽培品种为:
-一种匍柄霉属抗性对照(R)
-一种匍柄霉属中等抗性对照(IR)
-一种匍柄霉属易感对照(S)
-来源D
-BC5F3 HMC1
匍柄霉属抗性的结果报告于下表L中。
表L:匍柄霉属抗性。
“平均值”表示根据1.5中详述的症状评估,来自相同科的所有植物的平均得分。释义表明该栽培品种是否被认为是抗性(R)或易感(S)。QTL11是根据本发明的QTL;它的存在是用前述实施例中公开的标记物测试的。NT表示未测试。
结论:根据本发明的来自来源D的提供ToBRFV抗性的QTL11,与匍柄霉属抗性无关,与根据W02020/018783的ToBRFV遗传抗性相反,因为所测试的植物在包含本发明的QTL11的同时,对匍柄霉属无抗性。
因此,可以得出结论,本发明的QTL11与W02020/018783中公开的在11号染色体上的抗性不同。
实施例9:QTL11提供的ToBRFV抗性的分析。
来源D和易感亲本HMC2之间的F2群体已经获得。HMC2是一种在叶片水平上对ToBRFV高度易感的品系。如实施例1.B中所述,在接种ToBRFV后对134株个体植物的叶片症状进行表型分型,并基于QTL11的SNP标记物进行基因分型。
对于与枝叶症状相对应的性状,使用类似于针对QTL9使用的模型的混合模型计算不同测试基因型关于QTL11基因座的调整值。结果在表M中报告并在图6中示出。
表M
QTL11处的等位基因对枝叶抗性的影响的分析显示抗性等位基因的显著隐性效应,如表M所示和如图6所示。
序列表
<110> 维尔莫林公司(VILMORIN & CIE)
<120> 番茄植物对ToBRFV的抗性
<130> B14263WO CS
<150> PCT/IB2020/000546
<151> 2020-06-05
<160> 115
<170> PatentIn version 3.3
<210> 1
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; G=抗性等位基因
<400> 1
gttgggaggc agaagggtat aagattggac actgaaattt ttgctactta ccgtaagctc 60
atcaacatcg actgttagaa acatcagaga aagatatttc tcagacagct cacagtagaa 120
tggagagatc attctacatg gaccacacca tgaagcactg aaatttgcaa taacctgcca 180
aaatttcact tgcgtaagaa rggtccaaaa tgccaaagcc agcaaaaatg taaaatttta 240
agcaagcaaa aggcgaggga tccatatggg gaaactgctg caccattctg ctattatcct 300
tctcttcgca tgtgtgcaaa gcattagagt taaagtgcct tgagtattgt tttttaacgt 360
cttttgaatt ttagtgcagc tgggctagag gattaagtta c 401
<210> 2
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; G=抗性等位基因
<400> 2
tactgtacca atattcagtg caaggtatag gtgtccattc atgagggctg tcacccatga 60
acaccgaaaa tgctgctcac agtacttgct gagagcttca gacgaaccat ccatttgcag 120
ataatatcca gtgctcaaga taggggtgtt tgttggagcc ggtgaatttt ttccagaaga 180
gagtatatgt ttgggatcac rgtctgtgac aatgacctcg ggatgtccat gtagcctaat 240
aatttcagaa gcaaatgaag aagccatagt ttgagctgtg aaggtagaag aaaaggcaat 300
gattgattgc ccatatttga gttaccagtc aaccaacgtc aagattgtta ccttcccttc 360
gaccccaaaa ggcaagtaat gcattggtac ctccttaaac a 401
<210> 3
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 3
tactatcgaa gggtaaaaat aaaaaacaag ttgttcaaat atccattaga atgaagaaaa 60
gaaaagcaag ttaggggctc ttggaccggg tggtatcaat gtctaagcaa tggttctcca 120
gaggcgtcac accattccaa gccttattca gcatggagaa aaatccctga aatctaatgg 180
ctcaagtcta gaaatagcct rgtcgataga cacacaagta aaaattacct ttaccgtctt 240
ctacatcatt agcattaggc gttctctgac cactcccttc ctcttctcct tttgcaaatg 300
acgacagaac atctcgactt cgagggaagt tttgggtggg attggagtaa aatcgtgtat 360
tcactcccca tactcagcct ccaagtccta tttaacctgt g 401
<210> 4
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 4
aagtaggaat acggtccatg gtctgtgttc atggattgag acataactta cccaatcact 60
taaatgaaca acagatgacc agcgcggagc attgttcaat ctattgtccg tatgtttgat 120
cataactgag gttagcagtt agttacggga aatttacgga ttggatcaac ttaaaattgt 180
cataaatttt agcacaaaat raattaggtt ctccataacc catgatatga tagataattt 240
aatattctct ccatatccac tgagtttgct aaaatttgac ctccgagtga aaagttatgc 300
ctattttagt gaaggcctat tgagtagacc ctaacgacag accgtctatt gaacaacggc 360
ccatcagtcc aggtcgtcga tttccgcgac aacaattagc t 401
<210> 5
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> C=易感等位基因 ; A=抗性等位基因
<400> 5
aacaacccaa tattcttagg aaatgatcct taaaatggtg catcttgctt agacaaccaa 60
tgtaagggct acttgactag agagagaagt ctctgggatg attgaagcta ccacaacatt 120
tggtttagct cccctccatg tgtctattga tgatttgacc gctagggtca caacttgtga 180
gagcaagaaa ggggagaaat mtgaggcttt gtcttttaaa gacaaggtag caaagcagaa 240
gaaggacata gactatccaa agtctactga cttcacttca ttactagagg ctgctaatga 300
tttagaaaca attgatactt tagagattac ttcgcctacc accagagagg tacataggga 360
ggatgctaga attgatgtat tagatgctga aactgatgag g 401
<210> 6
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 6
cattgcgaaa ttgcatttgt tctcttaatc aaagtcttgt aactttgggt gatttagtgt 60
gttctcggtg aaccgatcgg caactcaccg actacacatt tatgtcgccg acttgatttt 120
tccccctccc ctccgggcta gtatacttga actgtaggcg aattggggag ccactcggca 180
gttcaccaat tggttttggc rattaccagg atcttctttt tctccacttg ttcagctcgt 240
ttttctccct tttttctaga agtgtcctgc cttgcttatt aactcatata ccttaaaatt 300
aatgctttaa caatagtttg tagaataaaa taggcattta aggacactca aactaccaaa 360
aaatatccct aaatgagtaa aatctttgac tcattattta t 401
<210> 7
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 7
gtaatagggt cataaaatac ttggtgtgaa tttaccaaac accctaagca ttggcagaaa 60
acatatagtg tttgaacccc tcaaagacga gacctaacaa atagaccatc tgcgcatcaa 120
cggaccgttt gtggggtctc gtgggttgac acttagcttt acatggagcc tgactaacaa 180
accttgaaga tccaacagat rgagcaagaa gacaggccat cggttcaacc attggctgtc 240
gttgcctttg ttggtgaaca cctcaaggaa atttttaacc aagatttggg ctctttctct 300
agggcccctt gtgggtcaaa ggtggggtcg tactaaacaa ttaaattcct taactatgta 360
atataggttt aggaatatct catgcaggtt ccaataaaaa a 401
<210> 8
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; C=抗性等位基因
<400> 8
tgaaatgttt aaggtttaga ttggttggtt cgctcacata ggagggtaag tgtgggtgcc 60
agccccagcc cggttttggg tcgtgacaat tggacttcca tctgtggtga ttttaataca 120
ccgatatggg tttttcccat gctattatct tcactttagt ttcatgagtg caagcatccc 180
ctagttgaaa gagacctttc macattgaag gccatttgat ttaaggaaat gttacagcca 240
acattctgta tgtatccata ataataacat attttactct actcatgtcg gatgctcttc 300
cttaatatat gcttgcgcac ctattctttc acaagatcca acatattatg acatgtctgg 360
cttataagag tagtttgtgt gccccatttt ggtgttttgt t 401
<210> 9
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; C=抗性等位基因
<400> 9
gacttgactc ttaactttta gtcaattgaa ttatggattc aaagttaatt atctcatgtt 60
tatggatgat tttaagattt ttgagtgtac cttaaagtgt gggaataaac tagaaaacat 120
aggtatgttg ctagggtact agtacgaaat gaagtgggaa aagattgatg aattggtaac 180
cctggcgcat taagaggtgc sggccgcaag tgggtaacat cagatactag tgtagggttt 240
tttggctggt atggagcccc agattccaaa cttcagcgcc cctaatgagg cgctttggct 300
ggaacagcgc cttagtccct tgcccagttg aattccaact tttcttgctc atttttcaac 360
tctaaacccc taatttcaac ttgattcttt acccaaacac a 401
<210> 10
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 10
tatgagagag aattttttaa accatctaat ccatgaaaag gcttcacaga cattagatca 60
gtacaaggac cgtcaatggg gttcgtcaac ctaaaactac agacagtgtg atatttttat 120
ttttttccat tgttagctcg tcatcctaaa agtgttttga gtgtgccgtt aagatatatt 180
ggtactaaaa atttctccgc rggtactatg tctcccaaga ggcaagtggt ttatacaaaa 240
aggggcaaat tcaagttagt tggcccttct tcttggttaa ttgatgaaga cctagactag 300
aagagggacc cagcctacat tccttcggac ataaaaacac caccaacaaa accctaaact 360
actaagagca ccattctaat gaggaccgca cactgttcgg c 401
<210> 11
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; C=抗性等位基因
<400> 11
gaactatcta acttctttca tcctcctttt atgcgataga gttcatcttt ataaaagttt 60
ctttctaact cgtgcttgca cgtatctttt agatcatgcc tccatgaaga tttgtctgag 120
ctagtcgttc taggagcaac gttaaggatc aagaggtgcc caatgcacta gaagtgcgac 180
cctaaggaga ggtcacttct mggatcggat aatgagacag attgcgacca acaaagctat 240
gaaataaaga aagaattgac aggagctagt tgatacttct agcattcgtg agtacttaag 300
gatgaatttt tcaagcttca ataattcaag tgtgactgag gatccagaat attttataaa 360
atagttgtag aaagtgattg aggtcatgcg tgttgcttat g 401
<210> 12
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; C=抗性等位基因
<400> 12
aaaatttaac catcaaatct tcattaaaca tagacgtatg ggatctaatt cacctttgag 60
acctctttcc tcaaaggttt tcttcaaagt attttcaaaa gttgagtttt cttgtaggtg 120
cattcatccg aatttggacc tcccacagtg gttcaaagtt caaaaatttg atgtcattgg 180
aggaataggg aaccccttag mgcatctaag gtcctattgt tacaaactaa tgggagttag 240
gagtgataaa gctatattga tgcaactttt tggcagaaga ttgagcggag aagctttgga 300
actgtttaaa tctcacgaga ccagacaatg agattgctgg aatgcctcgg ctaatgaatt 360
cattgagcga gttgcttaca acgtagagat cgtccctgac t 401
<210> 13
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; G=抗性等位基因
<400> 13
agttagtgag ttctatagtc tttggcaaga gttttggagt ataatatgat ggtaaaagct 60
tgtaaaaatg gtttccgtac ttacaatagg agtttaaggg gtccgggtac gactccccaa 120
ataccaccaa ggggtccttg aggaggaccc taacactgac caaagagatt tcaaaacaac 180
ttggttgatg gttgcaattc rtgaccagta ggttggtcta caccccgtaa gtcaagtcat 240
gcaccactcc tacaaagctg atcctcgggt gcaaaccaca aaggtggttg acgtacagta 300
acttctccta cagtccgtag gtggaggggt gtaggtgttt ccagttttaa gtaatttatt 360
ttagttaggt gtttggtaat cgattagtta tacaattacg g 401
<210> 14
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; G=抗性等位基因
<400> 14
cattaagaga ctaaggtcgt ggggtaacaa tccatggacc aggaccacga cctgtgacct 60
ggcctaagat ccgtggatct ggcagtgggt cgacccttag aggattttct ggaccttttt 120
gggaagggtt gtaggttgaa atccacggac caccacttac agtctgtgcg tcaccctagt 180
ggtaactatg gattgcacat rtagcttctg gaattaacat gcaacttttg gatgttggtt 240
ggtcttttta ggataaaagg tgttacatta tctccccctt ggaaataatc ttcctcgagt 300
gaagactaaa ctagctgaat atgaaggaaa agagcttcca accctaccac taattactag 360
aaactgattt ctgactgaaa taagttccaa ggacatgcaa a 401
<210> 15
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> C=易感等位基因 ; A=抗性等位基因
<400> 15
gaactatgag cttctgtcaa catctaattt gtcaactcat cagcatttaa aacacataat 60
tgactttgat atcgaggaca ccatctcccc attgtgcaaa agcctcaatg aaattttcag 120
atgttgactt ctataagtca actcatacca gatcaagatc ttgcttaccc ttcacatctg 180
atacaaaaga tgattccaaa maattttgaa cctctacaca accttcatta gaatatacca 240
tctaaacatc caatctggaa aatctatgaa aatcatgaac agaatctttc ttatcatcct 300
caacaggaga tacactacct aactataacc gactaagaac atttgtaacc aaatttatat 360
ttttaggatg atatagggaa cttatgtcat aatccttcta c 401
<210> 16
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 16
gtcattatta taattagaat caataggaag tgataaaaca cttcagaaag acttaaacgg 60
acagaatagc gaaaagctgg taaaaatgat gatttgggtg catcttgggc gcaccccacc 120
agctccgaat tgattagggc tcaaaaatag cgcactgcta gcatgctact ctagtctgcg 180
tggcgctatt tgggactgtc rtgccatcct ttacttggga aaagattggc aaacttttga 240
tctcattttt taagttttaa atcgattaat ttcgattact tttctaaaat tcactttaat 300
gtaagatatc caaagcatat acacaaaatc taactcaaat aactcaagga aacaacactt 360
taatatataa aaaaaactct ttaatctcaa cctaactcaa g 401
<210> 17
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 17
ttagctagtg gatccaccta aaagatgaaa tatataggtt taccttagga aaataggaca 60
tatgtttttg tgtgggacat gacaccagat tctatacatc tcacattatt tatgtcggtt 120
aaggctaatt ccctacatga caagatcatg aacataaact atatttatga cttctcaagg 180
ggttctactt agtgtgagta rgggtatgag acttcactcc tgtattccac gagtagcttt 240
gaaaaggttt tatagtgtgg ttttcttaat ctatgaatat atatgaattt atgtatgtac 300
aatatggatt attgctataa gtggatttcg ttgttacaag ttgattaaca tatatgtttc 360
attgtttatg gatgaggtca aggtatgtta tgtatgatta g 401
<210> 18
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; G=抗性等位基因
<400> 18
gcagcatgtc tcattaggtc tatctctcag ttagtagcta gccttttaaa ataatacttc 60
aacctagaca gacaaactag ccaataatat ttatttatgt tagaaacaac caacaaactc 120
actatttttt taatttataa ttcttgaaaa ttaccttctt gtcgatttct ttatccaaaa 180
gaagaagaaa gaaaaaataa ratagtctca ttgtcactta cactcacatc attagtggaa 240
tatcttttgc ataaaaacca caaacacata taaagggtga ttccaattaa acttccctat 300
aattgacaat tacctattcg ttaacagaat ctagagatat tatgacaatg atattacaca 360
tacatacact gaatattaaa agtctatatg attaacagaa a 401
<210> 19
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> C=易感等位基因 ; A=抗性等位基因
<400> 19
tcatgaggct tctctcgggt agcggcataa ggaaatcata cagagggtgt agtatcatca 60
ttgtgactgg ccattttgag agagattaca aaaaataagg taagttagga tagtatagac 120
acgataggga tgcctagacg gaaagagaat ttttttggga aaattaagcc aatctactga 180
cctaagtcat cttgatgtag mggcaatcat cccgctacaa cgggatgatg caggccacta 240
caacgacctt aatctaggac gaacttcaaa cgcaaagagg gcgtattttt gtcccaaaat 300
ctgagtttta tcagtttttt agcatatcaa agtctataaa ctatcctaga acaagaacta 360
actaacatgt caaccaaata actcctaaat tgtgtctaaa t 401
<210> 20
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 20
ttgattacta acaacaagtg acttgattaa taagacaaca attgtttcat gatagaagat 60
tgaatcacta gtgaagtctt ctcgaagtat aacttcaatt tttaagaact ccagttatga 120
acttcatgat aattctgagt tcaagattcc tcatcatctt ttgtcttggg ggcaacattt 180
aaatcaaact tcagatgctc raccaaaata gattttgtgt ttcttttttt actttctcca 240
tccttaactt caatgtcaac ttattctcct tcatatgctt gtcttgaggt aattctgagc 300
aaaggtgcag aatctcatgc attagcactc tctttttttt tttccgttac atgtttgcaa 360
ttctttatag agatcttcca atgcctcaaa attatttgat t 401
<210> 21
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; C=抗性等位基因
<400> 21
gttttcttat tgtctattcc aatacattac ttctaacttt tctatttcca gatgaagata 60
cattcatatt aaagagattt caccatctca caaagagcta gcctagatgt aaaatgttaa 120
tgcactaatc ttatcgaaga tgaaaacata aatggaaatt aatgaatgta tgagcgtttt 180
ctttgattta tcacttttcc matattcact agttattaga aagttcaact aaatttatca 240
atttaggcaa tataaggcct cattaaagtc tgaggatgaa aaaagctgac ttaataacat 300
aagaaatcca taatctttca tttttaatct aaaacctcat atcgatggat aattgattac 360
taacaacaag tgacttgatt aataagacaa caattgtttc a 401
<210> 22
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 22
taaagtaaag ttaggcggca gaagataaca catggaaggg acaaggtctt tatctgaagg 60
ataagtaagt gactaggaga ctttaattga tgcgaaaaaa ctcaaatatg ccatcgaaat 120
tttagaaaag acttatttat atcatcttct agatatttgg ttaatttatg tcattgttgt 180
taaagaaaag acgcattcac rttttactcc aattttacaa aaccattttc tacacctcaa 240
ataaggtttg acgcttttca attggagttt ttggatcata tgcacacttt ttgcttcttt 300
ttctttaaga acaacaagaa cacatgaaga acataaacaa ctcccaaaat tttcaacatc 360
ttcaactctt cacgaaatca tctcaatttt cggctacatc a 401
<210> 23
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> T=易感等位基因 ; A=抗性等位基因
<400> 23
acatttgata aatgcaacat aaaaaataaa ttcaatccat tcacagaacc aacacataca 60
aagcaatcaa acttttatca gatgtgatat tcgatccaat caaaagataa cccatatcga 120
tgtggtattc gagataatcg atatgtatat caagggtggt atccgagcct atgaaaagat 180
atcatgtatc aattgtggta wttagaccaa ttgaatatac cacatcacac aatcagaatc 240
ataataaata caatcacatt tgtataaaca agtcaattgg aaaagtttat cgcatgagat 300
tgacgacaaa tatcatttca atttcatcca ttttctagtc cctaaccaca tattgtacaa 360
gtctactagc aaaataatta acttgaacat attacacaat g 401
<210> 24
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; G=抗性等位基因
<400> 24
tatacatgag aaaattgttc gagaagggtc gtcgaggtag gtgatggttg aattattttt 60
cattgtgtaa tatgttcaag ttaattattt tgctagtaga cttgtacaat atgtggttag 120
ggactagaaa atggatgaaa ttgaaatgat atttgtcgtc aatctcatgc gataaacttt 180
tccaattgac ttgtttatac raatgtgatt gtatttatta tgattctgat tgtgtgatgt 240
ggtatattca attggtctaa ataccacaat tgatacatga tatcttttca taggctcgga 300
taccaccctt gatatacata tcgattatct cgaataccac atcgatatgg gttatctttt 360
gattggatcg aatatcacat ctgataaaag tttgattgct t 401
<210> 25
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 25
atagagcctt aattttggtc atgccatctg tggatagtgg aactgcacat tttgaaggct 60
agagttaaga ttcaccgcaa tagtagaact tgttttagat gatgagttac atttacacca 120
tcatatgcac ttactagtag tagatctaac cgggggattc acaaatccca tgatgattgg 180
tttcagagac taacgaggta ragtgatgtg atgaataact aacctttgag ttttgattgg 240
ttaaaattag tgtatacttg agtgattcca taattatgcc atctcctaaa aacaacaaga 300
aggtacgcat taaaactatt ttagcattcg aggtcaagaa tcaaagatct cactttgatt 360
ataaatagaa ttaaagatct cgccttcact gtagcttgaa t 401
<210> 26
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 26
ttggtgagtc tattctagct gagattgtat cgtgattgta ccatttccgt caatcacaaa 60
agaaccatgg atgacttagt tgagttagat ataattttga attcattcta ggtatgaatg 120
gtttcattct tgttatgact caatcgattg aagaactcga gttgtgaagt tcaaaatccc 180
taatgagaca cttattgagt rgagaagtag ttttgtagtg cctaagggtc attttttttt 240
gtatcttaag gcgataaagt tagtttctaa gggttgttcc tatcactttg ttagagttaa 300
cgactctagt gttgagatgc ctcttattca acaagtcgca gttgtgagtg aagtttccag 360
atgatcttcc cgaagtccct catatgagaa atagacctcg a 401
<210> 27
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; G=抗性等位基因
<400> 27
ctttaaatat gtttttcgaa ctaaaaaagg ataagaaata ctccaactag cacaggtatt 60
ttgtacagaa aataaagaag agccaagtaa ctttttttaa aaaaaatcca aaagaaactc 120
tcatgtttga gcaacatcta atacttctag gacaacaaca aaaaatgctt aaaagatggt 180
aataacacta tgaatcatca rattaaacta attgcatggt aacataatga acagaacacg 240
tatacaccta aagacatgga tcatatgctt ggactaacga gaatacaata aaataaaata 300
aaacacattg tgtagtgaat ggggataatc ttaaataaag aaccaacaga ttgattaatt 360
ttgaggttct gcctcatact cttgttgctt gaatattggc a 401
<210> 28
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> C=易感等位基因 ; A=抗性等位基因
<400> 28
tttgtaaacg aggatatgtt ttcaacaata ttaacaagat caaatcacat tatatttact 60
ctcaatacct aacattgatg actacaagac ttattctcaa agattcactc cttactatca 120
ccattatact taaggttcag ggtattaaaa tcacaatttt cattagtaga tgattctagg 180
cataattcgc catagaggtt maatttcaat tttactattc atgatccata ttatgcacta 240
aaatctagtt caattcattt ctatatgttc ctaatcttga aaattatgaa agttgatatc 300
aaatttagaa gatcaccctt gaatcctcaa ttttacctct catggcataa acccaattaa 360
tatcaattct atcaactaga ctagggttaa tcatgaaaat c 401
<210> 29
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 29
ttaattctgc actagttttc acgtcttatg taatgaatga tagcgaaaag gctagtacaa 60
gaccttcgaa aggttaatta cacaattgtt ccgatccaag ggtgccttat tctacggtgt 120
actacgggtc atgaaaaatt ttacccatag agtccattca gaacttgatt tctaagcttg 180
cgaatttccc caaaaattag rgttcaatct aagtcaaaat ttctctttca agacctttct 240
aatatttaat ttatcaataa ttatgattga attgttgatt aaatattgat ttatgaatcc 300
ccatatttct aaattgtgaa tttatgattt gggttgatcg aacataccaa aatgtgaagt 360
tattatgtct atattgattg agttattaac taatgatatt a 401
<210> 30
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> C=易感等位基因 ; G=抗性等位基因
<400> 30
cgaggagttc ttggagatga gaagttatga ataaatgaat gttatacgca aaagcctaca 60
atttgggcat tttcaaactc tacaccacga tctgattact agcaattttg cagtttagct 120
tcaactgaat cccttttgta aaagtcatca atactctact tgtttaatct tttcttcata 180
aggacatatt tcaggtgaat stagagaaat cgtcgacaat cacgaagatg tatcttttcc 240
cacttctgct ttaaattctc atttgtccgc aaatatccat gtggagaagc tctaatggat 300
tgattgcggt gacacctttc tttgacctga aagatgatct tctttgattc cttcttacgc 360
aagcctcaca aattttattt tctagagaac atttttattc c 401
<210> 31
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> C=易感等位基因 ; A=抗性等位基因
<400> 31
ttaccattgc atgtactata atccatggat ttttggtgat acgtcaatcc aacttgtttt 60
ttgacgaata tttttatggg aattggttag gaccttatta attacgtgat tggtaagtag 120
gggcaaacaa acacttttta aagttcgaaa tagccccaat gtggaagtga ccgaaattac 180
tattttacgc ctgttatagt mcatgaattt ttggtgatat attaatccgt cttgtttttg 240
gtcgaaattt ttatggacgt ccgttaaaaa cttattaatg gcatgaattg gtcaaaaggg 300
acaaagtaac aaatgcattt tcaagttcaa aatagtccaa tgcagtaaga gcatccccaa 360
gagtttgttc tactaactaa aaaacatgtt gaggattcaa a 401
<210> 32
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; C=抗性等位基因
<400> 32
atgcacaaac aaatttggat tatttatgcc acctactaga aatatttcac tgacttaaag 60
acttgatcct acaataatgt gagcatatat ttcttgagtg acatccaaaa taggcaataa 120
tgcttgtaga tatttcattg ctagttgtct tctttctgct gtttgattgc atcaacagaa 180
ccattactga aacggctgtt scttcttttg tttctatgtc gatattggga ccataacttt 240
gctacactct gaatgaaatc aacttgtata cttctatcat aaacttgagt attatccatt 300
ttaaaatatg cttttgcgca ggactctttg tcctctaact cgatgaaagc ataacagtgt 360
ctatccccag tttcgtgatc tttgataatt ttagcaaata t 401
<210> 33
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; G=抗性等位基因
<400> 33
tcgtaccatt ttttgtctta aggcttgtaa gatgatctct aaagagtgtt catatcatat 60
tgtcagagtc aaatatttag actctgaaac tcctcccatg gagttagtct ccatagtgag 120
gaaatttcct aaggtctttc ctaatgatat tcccataact cctctcgaat gagaaattga 180
ttttgtatcg cattgctacc rgacacaaat cccatttcaa ttcctccttg tcggatggat 240
ctggccgaat taaaagattt gaagcttaaa ctcaagaatt tactaaagaa aggattcatt 300
aaaccttgca tttccccatg gggtgctctg tttctaattg ggaagaaaaa ggatggatct 360
cttaggatgt gtattgacta ccagcaactc aataaagtga c 401
<210> 34
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; G=抗性等位基因
<400> 34
aggaagtttc ttccgtaaaa gttcaatgga ggaatcattt agttgagggt gctactcagg 60
agccaagtcc gatagtatgt tgatatcctc atctatttcc ttctactcct actctagttt 120
gaggtaatga tttcctcttg gtttgtttta tgtgttgtga gtcatgtaca ttttatgtgt 180
ttccatgatc tcattttgtt rtgcatgttc ttgaaaaagc ctatgttttg aaagaaatga 240
cttatatgat ttattttcct catgatgtgt gcatttagta tgagttgcct atagacttca 300
taatatgaat agatgaacat gtataagata aactttttga gaatgaattg tttaaaggct 360
ccatgagatt gtgttgagcg tgttttagct tggagaagtt a 401
<210> 35
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 35
gaagtgagtc ttacatcttt ttgttcaact ctttttttct ttcttttctc tttctctttt 60
ttttccttgt aattgataaa ttttcgtgag tcacctctct tgattaattt ttctattttt 120
tcctcattac ttttttcatc ccgacttttt gggtcatcac tcataatagc caccctccac 180
ttatgaattt gccatgagtc ragctacata tttcccaagg ttaggtcaag gccaagacaa 240
ggccagtttc tgcattaacc tctccactta tgattttgac ctaagttgag ttgtacattt 300
tcaagtaggg accacggcca atacattaaa ctcatggtgg gtgacttggt aaaaaaagaa 360
aggtctatta taggttcaac aactttggat caaaaaatga t 401
<210> 36
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 36
ggatgtttgg actagaccca tgatatcctg aaaaacataa aagaagatgg aatccttcgg 60
aagcaagaac cctcaccata gttaacttga actccctatt gtatcaacag agctcctatt 120
gatgatcatg aataattggt tcttcattat aaaatgatgt agataaattg tcttagtatg 180
tgaaatatac gaatatttaa rgggaattca aaagcgtaaa cataagattg aactggtgca 240
atagaaacac acttaactct tctaaactaa gtcgtctcaa cgaaacatag ttcaattcta 300
ctcaattata tgatattcaa atcagtgaaa gaagagtcaa atcaacaata agatattcaa 360
cgctaggtac tcatcgatca ctagtttttc cacttaggat a 401
<210> 37
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; C=抗性等位基因
<400> 37
atgcacccat atatttccca ctcactttgg atgaaaatga cgttgaaaga tatttttaag 60
tcgcatgtat tcaatggtaa aagatgtgca ccccgtctat ttcatgttaa agtcaggaga 120
gtctgacata gaatgatatt tttatgttac ttttattcaa atgtggatga tgtgcacctt 180
atagaatacg tattgaggta sgaggagccc gatgttgagg gatattcata gtaaattata 240
ggaattagaa gtctttacat atatctaatc gatgaactat gatatgtcga attgatggac 300
atgatttatt tttcccaaag atactaccag ttgctcatct tattctttct acagatgtac 360
atacattcat atctgacgaa tggattatct tcattcatcc t 401
<210> 38
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; G=抗性等位基因
<400> 38
caactcatac cccaaacaac aaatttcatg aaagaagaaa aggaatgaaa caaattgcta 60
gaatgcttta tagcttcctg aagattagat gtggactcaa cacatcattc tggatgattc 120
tattgcacat tttctcttct actggagacc ggtaacctgc gctctcatac aaattttttt 180
acgacccaac ccactggacc rtgcgggaac ctactctaat atcttaaaag gagaacccta 240
ttttcctaat tgttataata tacagaagaa taagaaaatt aagatataga aataatttca 300
tatagattta tataaataag aatttaaatt tttcaaaatt aaggcttgta aagtaattaa 360
aaccctactg atctagtcta aataggtata ttatccagta a 401
<210> 39
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 39
ctaatttaag caaatattaa gacgttaatc acgagatgat tagtcgcaac aaattgaatg 60
aaaacagatt atgacccaat taatacaact aaattaaacg caaagatttt caagtatgac 120
atagtgatga caatctgggc aaaatttaat caaccccaat atgcaaataa tagagtaaaa 180
agagcaaata ttgtagggtc rtaaatgctt agaacaaggt tcataactat tattcaaaga 240
aatcacagaa ccatagatgg aatttaggaa aattttgtga gaaaagtaga tagtaattta 300
aacaaaagca aacccgaacg gatctgtcga gatccccttc agatgaaaac tctagaactt 360
catgtgaagc tgatgactga tgtggtggct catcgctgtt g 401
<210> 40
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; G=抗性等位基因
<400> 40
ccacttcccg tgtagcctct tcttttttct tgttcctcca aagtattttg accaaagcta 60
tgtctctact ccttaacttc atgacttgtc ggtccagtat agccactggg acttcctcaa 120
atgataggtc ttgtgtaggt ttagtgtctt cggttttgat gacatgggaa ggttgtacta 180
aaaatgcata acatagatac rtggaagatt gggtgcactg cagctaactc aggaggtaac 240
actaactaat acgctacctg gccagttaag cataaaattt tataaggccc aatatatcta 300
ggactcaaca tatctttcct tcaaaatagc ataacacctt tcataggtga caccgtaaaa 360
aatacaaaat catcaatcta aaaatctaaa tccctacgcc g 401
<210> 41
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; G=抗性等位基因
<400> 41
ccatactacc caggcccttg gagcacttag acttactccg atttggccga aatgggcatg 60
ttgatgtctt ttatccttaa gaatgacgtc aattgctcca taatagggga aaatatgaca 120
tttggcacaa tacggggtta ctcagacttc accctactag accaaacaaa tattcaacac 180
aatacaataa ttttgactca rtttgaacaa taaaagaaag ggaatcatga ttaaagatcg 240
ataaataatg atttcactta aagttaggca aaggaatagt cacacgatga ttacccagtg 300
atattcaaaa acaaaataca gatttcattc gagttcaaac acatatagag tttgagtttg 360
gaaaccaagc ttttttgttg aaatatattt gaaatgttag g 401
<210> 42
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 42
ttttttgttg ttggcgaaaa gttgttgcca attctatatt ttgttgtaat ggacatgcat 60
gcatacacga tttacttaat tttaggttac tcatgactac tttcatggat ggtataacct 120
ctccatattt tgtaatgatg gctaagcctc ctcactctta agggtgtgat tatgggattg 180
gtattgattt aacttagtgg rtttattgaa ttgctctatt ttatggtcat ttatcatgca 240
tttgaaaatt ttgcaatgtg tttaattcat aaattgaggt ggccaactga tttttgtatc 300
gatcttacat tgatgcttga aagtgaccaa tatgagtggg tgattagttc attgtttgat 360
aactagtagt aaatccaagg taatggattg gatttaaaat t 401
<210> 43
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 43
atgatgtaat gttgttgtta ttggtggtgg tgttggaagg ctctacttct acaaaggaat 60
ccattagttt ttttattttt ttttgagagt tttttagggt ttaataagag aaggacatgg 120
tgtggaggtt gaggttaatt gttgatttga ataatcaaaa gatttctata tttgtaagat 180
acaagataga gactagagaa rgattgaaaa ttggagaggc aaaaaaggaa gagtttgatg 240
aaaggattga tgggagagtg aatgaaagga ttcctttttc tctctctttg tttagctacg 300
ataaaaataa cttttaacga taatatatat atatatataa ataaaagtac taaatcttaa 360
cgatattagt caatagtcaa aggattcaat gttaatataa t 401
<210> 44
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; C=抗性等位基因
<400> 44
tgtgcaatca aaatacaaat ccaatgtacc atgttttatt gttatttttt gattctcatt 60
ctgcgtgcag agtccatatt gaagctccga tttaatatga atcaccacta taggcaattc 120
aggggtgact tattttctca caatccaaaa ctcaaatttg aaatattaat taagaataaa 180
acaatttcac taccgcaacg mtacctttat caaatataac atatttatag agtctaacaa 240
ctctaaactt agagtccaaa atccaaggcc ccacggctat gtaaaccgta caaacaaacc 300
cattgtctcg tccacatcaa aactaggcaa accacaaacg ggtcgaattg ctaacacact 360
cagatccgtg ggcctcattg ctcctaccac ttaagataat a 401
<210> 45
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; G=抗性等位基因
<400> 45
caaatccaat gtaccatgtt ttattgttat tttttgattc tcattctgcg tgcagagtcc 60
atattgaagc tccgatttaa tatgaatcac cactataggc aattcagggg tgacttattt 120
tctcacaatc caaaactcaa atttgaaata ttaattaaga ataaaacaat ttcactaccg 180
caacgatacc tttatcaaat rtaacatatt tatagagtct aacaactcta aacttagagt 240
ccaaaatcca aggccccacg gctatgtaaa ccgtacaaac aaacccattg tctcgtccac 300
atcaaaacta ggcaaaccac aaacgggtcg aattgctaac acactcagat ccgtgggcct 360
cattgctcct accacttaag ataataagat aataatagta t 401
<210> 46
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 46
attatatcat attagtaata taattaacta tattgtatcg tatatcgata gttaaatatt 60
aaaagttcaa tcgattaact attttaccgt tttttttata aattagaggt ggcaaagctg 120
aactcgagcg aatggtactt cttcagtttt cgagatcgaa aatacgctac tggttttcgt 180
acaaatcggg ccacaatttc rggttattgg aaagcgacag gcaaagatcg gacggttgta 240
gatccgaaaa ctcgttcgat cgtagggatg agaaaaactc tggttttcta catgaataga 300
gctcctaatg gaattaaaac tggttggatc atgcatgaat ttcgcctcga aaatccacat 360
atacccccta aggtaattct tccttcattt taatttattt a 401
<210> 47
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; G=抗性等位基因
<400> 47
tatttatgga gttcagatat ccctctccta ataagaagtc tgaggtgtaa caattctcct 60
ctaaagagag ctcaaagaac atagagcata aagctgagat tgcaaaagga agtattgatc 120
ttccttcgta tgtaggtcct tataccatgg ttgggtaata aacatctaaa tacacaaatc 180
tagatttttg cgattttaaa raggaaaagt atgagcactg cgataaggtt gtgaaggtat 240
tgtctgccaa tgataacctg gatgccattc ctagattcta tgcaatcatc aagaagattt 300
tgtctcctgc atttaaattg cacataacaa ggttagttct agagcctcta aataaaggtt 360
aaaccaaatg actatatgag ggatttctag cttcttgtac t 401
<210> 48
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 48
aaacatcaaa gatcgccaat tgttcacata gtttagtaaa tgcatgtttt ggttgaaatt 60
tgtggtattt ttaggtcaca tagtatctag caaatgtata gaggttgata caaagaaatt 120
ggactccgta gagaaatggc atagaccact attgcagatc gatattagga gattttttgg 180
tttaatagga cattattgga rgtttgttga agtattctca tatgttgctt ctcatttgat 240
gaccttaacc aaaataaata taagtttgag tggctaaaag cttgttagaa agtcttccat 300
gaattgaagg ataggaagct tgtgagaaag acttccatga atagaaggat aagcttatct 360
ccaccccagt gtttacttta ccggagggta ctgatggctt t 401
<210> 49
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; T=抗性等位基因
<400> 49
ttgttttggg ttttgcatta tgaaagttga atatatgaaa atgtgcatac cttctcttta 60
tgaaaatgat gtaaataaat ttgatgatgc tttcactttt aaaatatcat aagtaacatg 120
aataattaaa tgaatatatg atcgtgaatg tatgtttatg actcaagaaa gattatagat 180
gagagtgtgt atatgttcta wgaagctttg ccacatgaca tgaatcacac atctagactt 240
gccttcaacg ctttttctgc tgttttctta aattatgtct taattaggat attattcgta 300
tatgtcacca taattataag tcacgttttc aatattattt ccactagtag taaagcatct 360
ctaacatgaa gtgaatcaat ctgggcctta aaagctcttt t 401
<210> 50
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> C=易感等位基因 ; A=抗性等位基因
<400> 50
gacaaccatc agtatgagaa ttttgtttat acaacgttat acaaatattg ccagagtcgc 60
cctatacatt taaaaagtat aaaactgaac tgaattgatg gatcgactaa aacactttct 120
tgaggcaggt gaggagccgc ccgacttgcg gtttgaatct atctacgatt gctgcatact 180
ttatgattat gtaaaatact maatcgattt agataattac tcaaacttgt gtttttacaa 240
ggtaactcaa gtgtactcaa tctctatgaa aaccctttac atttcgggaa gtagtacact 300
cttttcaaag tgaaaaaaat atctttttaa gggtaaactg aacttctctt tgtctagact 360
gaaaagcttt gaaaaattgt ttattctctt ctagaaaata g 401
<210> 51
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 51
gacttttgta gagataccaa ttaaacgatt aaaagtccta aaactggaga aaagaaacta 60
gagtatatca aaaggaaaaa tacaagtata tatatatata gaactaaaat aagtaaaata 120
catcatgtgc tcacccccct gtccggatca aaatgctcca cactagatca atgttggcta 180
ctggtgctca gactttcatc rcaaaaatag atgcagagcg tagcatgaaa accaaacaac 240
aggtaaccag taagaatact aaaatgacta aacttgaaat tagaagtaaa acgaaaacaa 300
ggaatagcaa gaccataggt caagaacaga agtttaagta acaatgaaat gcacacaagt 360
gtagaaagaa ctacaaaagt aatgcataag ctatctacaa c 401
<210> 52
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 52
tcaggcactt tataatctct aggtacttgt caataacccc atctatagac acacattttt 60
catcaagaat catctttaac cactctgcct tttatctttt cattgacttt ggcggaggac 120
catgtagtgg atgtgccaac actataacct gcacaggtat tggaggagta tcagtggtag 180
ggtaagtctt tgtgatccta ratggatcat ttagcttctt agtttattgt tcattccttt 240
gagttattga tagatcatcc ttatcagaac aaacaccatc acattctgat ttgtgtgggt 300
catttttgta atagccatca ctgtcagtaa tagaatctga taactcaaag atctggtctt 360
agtctttgcc tttctcatca gttttagaaa gtttaatagc c 401
<210> 53
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 53
ttgctacata tgactattct atggcaagat gtaattcata tgttgataca taaaattgtt 60
atgttattac aaataaatta catgtgacaa cataagtaag tcaaatattg caacatatga 120
ctattttgtg atgaaatgga agtcatgtgt tgctacatat aactgttcga ttgtgacaaa 180
taaatcacat gtgtcaacat rtgtaggtta tatggtgtca catatggctt tctgttgtga 240
aatacaaata tgttattaca ttaatatata aatcacatgt tttgataaaa tgtataagtc 300
atatgttgca acatatgtct attctgtcgc gagtaaaata accactcagt tgcaacacat 360
ataaccatct gttggaaaca cgaatgtgaa atattgaaaa a 401
<210> 54
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 54
agaatcaatt ggagcaactt acgtcccttt catcgaccaa cagaacatcg attccaccat 60
acctcaacac ctagcaaaac cattggtgga agagtactta cggaagaaac aacgggccgt 120
acatggattg aaggggcatc gatgcctccg tattcccaaa cttagcctgg tggtgaaagt 180
ccccaatatt acaagtccct raccaagacg acggatgtgc agcacgaacg aggggcattc 240
cttagagtca ccaacggacc gtcgacgggg tctcgttagt gaggtcagtt aactttgagt 300
cacattctat ttgggtcatt taaattagtt aggagtttag ggggttaatt aagggattaa 360
gagaggtcta attaagttta ttagtcaccc aataaagtcc a 401
<210> 55
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 55
gggtattgct cctagagtaa acattgttga gagagatatt acatatttat tgagatattt 60
tgtgagaatg aatcctccta ttattcatgg ctctaaggtt tgagaggatc cctaagagta 120
cctagatggt atgtagaagg tgttgagtga catgggggtg acttctaggg agaacgctga 180
gttagatttg tacaaaatga rggaggttgc taaagtgtgg ttcactcaat ggaaatataa 240
taggccgcta gagttgatcc ctataaagtg ggaagaactt aaagaagatt ttataggaaa 300
gtactttcca catgagaaga gggaggtgaa ggtcgagggg tttataaatc tcaagatagg 360
aaatatgagt gttgactaat actttttaaa gttctctatg a 401
<210> 56
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> C=易感等位基因 ; A=抗性等位基因
<400> 56
tcttaggaaa tgacctatga tgctgcccaa tacaggtact cttgtttatt ttgtagaact 60
tgtcatgttt tattttggta tcatcttggg tatggattag ctttatgagt gttttgcttc 120
cacaggttgt agaacaaggt cagtgtagtt ccaatttcct aatgaaacta ttctagagtg 180
gaaggcatga aatttgatgc maaggggtca aatcatttct tctctgatgt ctcgtatgat 240
catagccaag gttttttata ccatgtggtg ctggtaagga cctagaatgt caaacttctt 300
ttattgagtc aggcctatgc ttagagaata cctagaagtt tttcctattg accttccaag 360
agttcctcca aaataagaaa ttctcttcgg ttggcattta t 401
<210> 57
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; T=抗性等位基因
<400> 57
ttttagttaa attaattaac tattcttccc ttaataccta aataatcaca gaaccactta 60
atttattata tgatttatta taaaaacatg caggaaattt aaccctcaca ggcgaaaccc 120
agttgacaat ccatttgtga gtcaacggtt ggtgcttgcc tccgtccaca acaatcgttg 180
tcttggtcag agactggtgt wggcaagggc ttcctccatt tttctaagtg tggttctacg 240
gtttgcaatt acaccccatc gatccataca caaatcgttg tctgtatctc gtcgatgcat 300
atttcctagg atgtgatgac atcaactttc tacgatcctc ctcgagggcc attgattggt 360
cattgtggag tcatatccgg acgttttgac catgaatcac c 401
<210> 58
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 58
atgacttaaa attctttgat tgaagattta atttacgatt ttgtaagagg tagttgaatt 60
ggctaaggtt gcattgtgta gtgtcggttg ggctgcatat aagtaaaaga aatatgattg 120
gagaagaggt gactgatagt caaccaggtc atcaggacaa tatagatgat aacaatagtg 180
cttataagtc atatcatgct rgcaaaacgg gtgctatttt cttacaaaaa gatcatggca 240
atgtaacatt tcaagtcact agcatgagtc tttatctcct taaaatgaag gaacattatc 300
gagggctaga ccatgaatac ccgcatcagc atgtaagaaa ttttatagag atatgcagcc 360
cgttttaatt caaggagtca acccggcttt ggttatttca a 401
<210> 59
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; G=抗性等位基因
<400> 59
aaatttctat ttgttggaga agagaaccat cataggcaaa tgattagttc cattatacag 60
ccactactaa aaggtattgt gatactctta ccactatcat aggcaaacat atgcgactcg 120
cgtagcagta cctacaatca accccacaac aagccaaaag gtttttccct gaacaacttc 180
aacctttcct tcaatattgc racttctagg tccatcaaca ttccaagttc aaagcagtaa 240
aacaacggga gaaaaattac acaactacaa tccaaaatca ctagagtcct ccttcaacac 300
aataaaacac taatacaaca aaaacaatgt atagacttaa ctttcaaaga ttcaagataa 360
tataataggt tcagaatttt tacctagaac accaactcaa g 401
<210> 60
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 60
atgcttgtga tattccttgt atatctaaca tatctgtatt gctatgctca tcatgggagg 60
caacaatatc taataactca aaatttgatt ttgtatcagt gttggggtta agtaaggctt 120
cttcgagctt tctttttaat cttttttagt cataatcctt ttgagatgat gaacttgtac 180
ttaacttatc atcgtagaca rctagagctt atcctttgat cttaatgatt ttgcttaaat 240
atttgatcct ctctaattca tgaatattga aggaccaact aataatataa gagatctttt 300
tggtgataaa gaattgcata atttaatata ttctgataag gtaaatatat ttctctctac 360
ttgaaacttc tcaaactgta tttgatgttg agttgacaat a 401
<210> 61
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 61
aaacactata ggaaaaaatc cttgtacttg atctgccagc aataacagct tttatgtcct 60
gattttgaaa ctgaaatttc tcattcttca atcttctcgg ggatagacct tgtttttgag 120
tcacacatta gatttcctct tccatgtcct gattatttta aggagtaaat cagtagtcta 180
atcatcatca tcaagtttct rtcaatattg attattcatg tatacttcat attcttaatt 240
cataacttca taatcatcaa tcaaagggac aatatttact tggcaattgc catgtacttt 300
atcaggaggt caactaggag gaggttgtta aaatattggt ctgctcagaa tgttacgcgt 360
acggtgttga ttatcctaat catcaagttc atgtctttta t 401
<210> 62
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; C=抗性等位基因
<400> 62
ggagggtgac gattctaaca aagaaactaa aacagatcaa taacctcaaa gaggaccttg 60
gctaaatagt aacaatcatg agtaagtgta ggaagatact agaaagcatg gattttctca 120
taggggtaga aaagaatcta cgaaaaataa aactgatagt tttaacagac aacatagtag 180
ggctaggagt agaggtgttt satcattaaa gccatatttt ttaatgctag gagtataaat 240
acagttggtg catttgaaag actataaaac cttaaaaaac ttcataattt gactatgatt 300
gatgtgttgg aaccttttgc aaatcattgg tatattaaat caattatgtg tcatttacaa 360
atggatcatg cttattggaa tgtcaataat aaaatttggt c 401
<210> 63
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; G=抗性等位基因
<400> 63
caactagact agcttccgga ggtctttgtt gttcttggac gtttcacttt caaaatcctc 60
taagtgactt gtaaagctaa tattcaacat ttttagacct tacaacctca tgaaacatat 120
attaggctct atttatacct aaatcatttt gggttccctt cagttatact ttatggaccc 180
aaattttgaa aaacttaagc rcatctccta ccctaaacaa gtatttcaac attcaatcat 240
gagaaattgt catactgagg cttccaagta cttaatttta aatttgaacc ttaagtcaaa 300
acttaaatac agatcatgaa gttctttttt acagtatgaa cagactatct aacttggtgt 360
tgctaattaa acaatcgcat atagttatgg acaatacaaa a 401
<210> 64
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; G=抗性等位基因
<400> 64
aatttgaacc ttaagtcaaa acttaaatac agatcatgaa gttctttttt acagtatgaa 60
cagactatct aacttggtgt tgctaattaa acaatcgcat atagttatgg acaatacaaa 120
accccatggt caagacccac tcctgaattc caaaaaattt aactatgaac aaatattcat 180
gttttctaaa ggaaattgtt rccttagacg agacaaggag tggtgtgaat gtgaagcgac 240
actaagtttc caattaagaa cgcacaactt cactgaaccc ggacaccgaa gtaatatggg 300
aatggaggtt aagaagaatt tgtgtgactt ggatgatata gggggttttg gggatgatat 360
gagttggagt agggattgat tggtgggaaa aaggagtgaa t 401
<210> 65
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 65
ttcctgatca ctgtcattgt catgttgctt acattgtctc cttttagccg catccttttt 60
ttgttttcta ggcttgtttt tactcttggg agtcctttcc agagtactag tatcaatcta 120
cttcccttta tctttatttt tcactttttc aagttgtttt tttttaaaaa aagaattatg 180
tttcttgtta tccctgttct rcacctcttg ttgactctgt tgttaatcta tttgttgatt 240
cctatttgta gaattatcaa tggatctaag gtttgaatta tggtcagtat cgaacctctc 300
atacaaaaaa agaggcaaac tccctctttg gtatcccctt cctgcatgtt agtggttttc 360
tcctatcacc ccccatcaat acctccaacc acttcatcag t 401
<210> 66
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 66
ctctacagcc tgatcattcg aaggacttaa ggctttaatt agagactcaa tagtatcatc 60
atcctcatac gtatcatcta gagaatggtt gtccctatcc aattcatctt cagaaatatt 120
cacagcatac tcatcaatca aaggtattgt attaacttga catatctgtt tcatcttatc 180
tggtggttct attgacggtg ragtcatata tctagtctgc aaaggtataa tatctaaacc 240
tttgttgtca ttggccatca caaatttttt tcatggttca tgatcctttc ctaagcttgt 300
tactattcac tgttctgctg tatactttct ctcctattag catcatccct tttttgtttg 360
ctttggctta ttttttctct tgggagtgct ttcaggagta c 401
<210> 67
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> C=易感等位基因 ; A=抗性等位基因
<400> 67
tgttataaaa aaaatgagat catctcagat aatattttca gtatataacc agtaatatca 60
tatccagtaa tagaattgtc ccacatgcaa gttaaaaagg ttcagtaaca catttacttt 120
atataccatg tgaaaatcat ccaaattata aaatatcaat tgtggtaaga ttattactca 180
cagtacttgt gttgatgcac maaacttgtc agagaaatca ataaggtacc aaactcgcag 240
agaaatctat aaggcttcct tcgataaatc acaatcacat gacatggatc tcgagataat 300
tatttgctaa gactaattca aaactaagat aaatattatt ttaaactctt ttggactcca 360
ttttccatca aatctctaaa ataaaattat tcaactcata a 401
<210> 68
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 68
aaaaatttta ggcatgggta gctacctgca ttgcggttct tgttatcacc tagatatact 60
cttccagaca ccttttcaat atttacctaa gccccgaaga aatgaccttt agcaggaaaa 120
atagagaaat aagttattat acaggaaaca agattgcaag acaatattca attagaagtt 180
ttaccttctc aaatctttct rcgtacactg tgtgaaatca aagagaacac acaaataaga 240
catcaaaaca taaaatgacc atactcacaa aaaagcagag aaaataaacc ctcaaaaacg 300
gtacacagga aaccaacatg tgctcctctt ctgtccaagg agtccctctt cgccatttaa 360
tatatgcttt catattttgg ttgccattgt tttgcaagtt t 401
<210> 69
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 69
catcatatgc ttgagagtct tacctttatc attgacaagt gatgttgcag tgtgattttc 60
taagcttgca tataactcaa ttcatacatt ggaacaattg catacggtgt tcatgtaaat 120
gtacttcccg ttgtctaaga agttgaaaca caaagacaaa cttaacaact tcatggaact 180
acctggagaa tctgtgagta rttctggtac aagttcattg cattcataag aagtgttttg 240
aatcaccgca ttgattatga gtcactcaaa gagtattttg gttaaggaca ggatgacaat 300
cgtaagaaaa tgcttgatac tatcatggag gatcatatgg tgaattcact tttgaggaga 360
tcggtaaaaa attagagaat atttcccaaa ataataaagt a 401
<210> 70
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; C=抗性等位基因
<400> 70
tgtggtacat caaaaccctc aactaagaat tgttggtgtc atcaagcaac atattgagaa 60
tctagacaac acttgtatag aagcaacaaa tttaaaacca agcaatcata tgtctctcca 120
tattgaatct tattttaagt aaagttttgt tctcttagac gcaaacagtc taattttttt 180
aacatccaaa caagacataa maaaccaata aataacacca aatataatgt ttctctatcc 240
ctagagtacc aattttcaga caacgtcacc attgccctta tttcaacaga attccccttt 300
ttcgctcaat gtatatgaag atttagtata gggattcctt caactctcat gctcagaagt 360
aattatcaag gacatgtaat tctcaatacc ataggattga c 401
<210> 71
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; C=抗性等位基因
<400> 71
caaagtcata tagtaacaca ccattccaca aaagactcat tgaaactata acaactccta 60
acatggttta accataacat gacatacatt tcccaacata aaataccact tcacatatat 120
ctcatatata cccacagggg aactatataa aaataagcca aagttaactt ttacacatac 180
tcatcaaacc ataataccaa sataagcata aaaacataca ttttctccct caggcataaa 240
gataccatct catccaagca aacacatcta caagaccaca ggacaagaaa atataagaga 300
gagatcatat agatttaact tctttgtcac gacatgagaa tgatgaaaaa gggaaaatct 360
atgttctata tcatcacgct catggattat cagtcaccac c 401
<210> 72
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 72
ttctctttga gcaagcgatg ttggcaaatg aaaggcagtg taaccctagt cttgcacgta 60
taacatatgc catgtgcgca ttggcagggc aacattagat cgaccaaggg caggcaacat 120
catccaagtc tattgtgctt caggaaagag acaacggtca atgtcaagta gtatcgtcaa 180
ggtgacctta gttatggaat rtcacattag ttgttgatat tgcacgtcgc tcaatcaagg 240
gtaggcatgg atcgctccat catctgtggt ctgtgcacac tactcagcag ttgttaggct 300
tggtctttct gcatcgccct tggcctgcat acaatcacta gacaacatca gatgtgggag 360
gcactcattt tttcatggcc tgcaaattgc tcagccgact a 401
<210> 73
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 73
ttctctttga gcaagcgatg ttggcaaatg aaaggcagtg taaccctagt cttgcacgta 60
taacatatgc catgtgcgca ttggcagggc aacattagat cgaccaaggg caggcaacat 120
catccaagtc tattgtgctt caggaaagag acaacggtca atgtcaagta gtatcgtcaa 180
ggtgacctta gttatggaat rtcacattag ttgttgatat tgcacgtcgc tcaatcaagg 240
gtaggcatgg atcgctccat catctgtggt ctgtgcacac tactcagcag ttgttaggct 300
tggtctttct gcatcgccct tggcctgcat acaatcacta gacaacatca gatgtgggag 360
gcactcattt tttcatggcc tgcaaattgc tcagccgact a 401
<210> 74
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> C=易感等位基因 ; A=抗性等位基因
<400> 74
cacctagacc atcaatatga ataataggcg cgggataaca gatacacttc ttaagaatgg 60
ataaacggaa aacatgatga atcgaatcca ataaactagg taacctcaac tcataaggaa 120
ccttacgaat atttttcaaa atttcatagg gacacacata actagtactc aacttacctt 180
tcatgccaaa tctcaccacc mctttcatag aaattttttt ttgatacacg ttatcatttt 240
cttcaaactc tatatctcct ctccaattat catcataaga attttttaaa ctatagtctg 300
tttgcaaccg gttcctgatg atatgaactt tctctaaggt cttttagata aaattgggac 360
caagaagcga agactcccca accctaaacc acacaacctg a 401
<210> 75
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; C=抗性等位基因
<400> 75
ttataactta gatgacagaa aagatgttcc gtacctctat gatctcttgt ttgggatttt 60
tttattgttt ggtttgtgaa ttggtggagg ttgtggattt tacataacaa aagtttaata 120
gaggagttga caaagcgata catttgtatg tgtgcaacca taagatttta aaggctgaaa 180
tggtgtgtgt ctgagtgaag maaaatcaag attcttaagt aaaaaaggag ggcgggtaaa 240
agaaacaaaa aaatgatgag aagtactccc tccatcactt tcacatatat tctttggaat 300
tcacacacct cttaaaaaaa tgatttaaat aagttcttac tattataccc aaaatgttcc 360
attaacctcg ttaatggggt tttcttatct tcactggatt a 401
<210> 76
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; C=抗性等位基因
<400> 76
acaaatgctt tgtcttaaca catactcata ggtcatcaag accttcacat tatgcacatg 60
cttaagttat aatacgataa taccctaatt aataaagtaa tgttcgatga gtccagatag 120
tttcaaaata tataaagtaa caccgacgat gccacattgt ttcacaagca tagcacataa 180
acctataata tatattatag mgaaagatag gtcataatac caccaataaa atctcacaac 240
tttactccac aataaatcaa taaacttaat atgccaaggc tcttacccat agccatgatt 300
actaattcaa tcaaataatt caaatctaca attaattgaa gatatgaatt tatcatttta 360
ggaaacctta cttacaattg tatcataatt tttcataatt c 401
<210> 77
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> C=易感等位基因 ; A=抗性等位基因
<400> 77
attttttaat aaggttcaca cacaatataa tatatcctta atcttatatt gaatccttgg 60
ataaatagtc ctaatctcgt ggtccctatt cttcttcttc tcatatggtg ttcgtgagag 120
ttccaatgag agagacatgg gttttgattt gaagtctaga ttctaagggg ttcttttatt 180
ttttaaatag tcataaaaca mctaaaaata ccttaagtaa ccacatggtt tgacttttcc 240
aattcactct tttcacttaa aagtgaattg tgagaaatgc aggctccata gactatttga 300
cattgatcga ccactggttt gaaaattgtc catcctgtat atacgtagtc ttggtaagag 360
acttctacaa ggttcccttt tctcaggaga gagtctcaat t 401
<210> 78
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; T=抗性等位基因
<400> 78
gcatatatgt ttgtatttaa taataatatg aatcgtacca aatgaaatga atgaatatga 60
ataattgtat cccatgaatc aacatttgat tgtttaatta tgagaattaa caatgtgatt 120
gtgattgtta catgtagaat acattaggta tttcgttcca acacaatagc tgggagcgga 180
tatcaggtat cgacatatat wttatactgg gcgtcaaaat ccatcatact aatagtttgg 240
gtgtaggttt aataagagtg acattaactt gtcatataag catattgtat ttaatgtgta 300
aatggacatc ttttcataaa gataattgga ttttatatta tcttcttata tgtgacttca 360
tagtgttgta agctaattgc ttatatctca taatgaatga c 401
<210> 79
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> T=易感等位基因 ; A=抗性等位基因
<400> 79
gacttacact attttttttg gagaggctac atgtgtgtca atattgtagc ttgaataaaa 60
agaaggtaat atgtagtttt taggtgtgta ataccctata taaaaatata taataagggc 120
atatatgtca tttgatacta ttttaaaaag aactaagttg aattgttttt atattgacct 180
aatggagttg tttaaaaaaa wtgggctata accctatagg aaaagttgta cctggcaaca 240
aataaaaaaa gataatgggg atcaagaggg gttcgctgcg tacatcacca agaagaaaaa 300
gaaaattttg aataagaaag agaggaagcg tgaaaaggag aggggacata tgtagaaaga 360
aaagcaaaga attcaactga agaacttaga ggggagactt t 401
<210> 80
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> T=易感等位基因 ; A=抗性等位基因
<400> 80
gacttacact attttttttg gagaggctac atgtgtgtca atattgtagc ttgaataaaa 60
agaaggtaat atgtagtttt taggtgtgta ataccctata taaaaatata taataagggc 120
atatatgtca tttgatacta ttttaaaaag aactaagttg aattgttttt atattgacct 180
aatggagttg tttaaaaaaa wtgggctata accctatagg aaaagttgta cctggcaaca 240
aataaaaaaa gataatgggg atcaagaggg gttcgctgcg tacatcacca agaagaaaaa 300
gaaaattttg aataagaaag agaggaagcg tgaaaaggag aggggacata tgtagaaaga 360
aaagcaaaga attcaactga agaacttaga ggggagactt t 401
<210> 81
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> C=易感等位基因 ; A=抗性等位基因
<400> 81
atgaaccccc aactctgtaa cagagcctta tggtataaat gtgtactttt ttaaaaaatg 60
tttgggatat tataaatgaa gacttgatgg gagtggaatt agattttttc agtgacaagg 120
agatccccat gtattttact cattctttta tagtgcttct actcaaagtg aatgatcctt 180
aaaatttgaa ggagttaagg mctattagtc tgagcaactt cattagtaaa atcatttcca 240
aactattatg cacaagattg agccctattc tgcctaaatt aatctatctc aacaagtatg 300
gttttgtaaa gggaagaagt gtaactgaaa atatcatgtc ggcacaggaa atcacccatc 360
aaattaaaaa tactatcata ggtagcaatg tcattatcaa a 401
<210> 82
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> C=易感等位基因 ; G=抗性等位基因
<400> 82
agcaaaatga ctgacttatt tctcatcctc ctatgaatca attacaagaa gtggtcttct 60
ctatgaaccc ccaactctgt aacagagcct tatggtataa atgtgtactt ttttaaaaaa 120
tgtttgggat attataaatg aagacttgat gggagtggaa ttagattttt tcagtgacaa 180
ggagatcccc atgtatttta stcattcttt tatagtgctt ctactcaaag tgaatgatcc 240
ttaaaatttg aaggagttaa ggcctattag tctgagcaac ttcattagta aaatcatttc 300
caaactatta tgcacaagat tgagccctat tctgcctaaa ttaatctatc tcaacaagta 360
tggttttgta aagggaagaa gtgtaactga aaatatcatg t 401
<210> 83
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> C=易感等位基因 ; A=抗性等位基因
<400> 83
ctggcagctg ttgatgagga gaaatctata ttgctagaat gtttgaaagt tggctagtag 60
gtgcctgccc tattagcttg cttcaactcc atttcccttc ctcctgaaat tcaaccaatt 120
ttgagttgtt aaatctgtta ctatttgtgg tgaattgttc taatggtcca attccaagcc 180
aattatctca ccaaaaggaa maattccaag cttgaagttt ccattggatg tattgctcta 240
cttgttgtag gttcatcagc atgtatttcc aagttaaaga atctccagta tcccaatttt 300
tgcttacagg gttggacctt tgttggtatt ttactctaaa aaattctccc cataaggttt 360
gtcttgttca aaaaatccac catcgtttga attggaaggc c 401
<210> 84
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; C=抗性等位基因
<400> 84
agtttatgtg atcaatatag gctatattga tgccttatat gtctatacac aatgttgaat 60
tttgaatctt cctaattgac tatatgaaca ctatgatgag ttattgtgtt tattttgtag 120
aagtgtgtag ggggacttgg agtattctaa agtttaacct acgtgatact tgaataatat 180
tgtacatgtg ataatatgat mactaagtgt gtcttgttta ggtaaaccga tatcccttac 240
ttgatacatg aaagaatgtg aataagaata tgcatccttg acttaataag aaaagcaccg 300
ttttgattaa tgtatatgta tgaataaata tgatctttaa tatagtaaag gtcatttatg 360
tggaatcatt tagaaagaag tttacgatat ctttgatatt g 401
<210> 85
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 85
atttttagtt cctttagtta ctccttttga cattatcaat ttatagattt gcacttttaa 60
cataagacaa gttcgtcatt tctagggttc caattatatt ctattattaa cctacttaag 120
tcacaacaag gagctctaag ttggaatatt tagacagaca gagacacgga ctttttagat 180
agacttaata gactacacaa rcgcattcta tcattccaag ggacatcact acactagttt 240
tagcatgcaa tttcaaataa tttagtccaa aggacaagac caccctctgt atttctgtaa 300
caaactatga actagaattc aatgttaagg gaaaacaaaa taccatcaat gcagagtggt 360
gttgttgtga gttgaactat taaggagtat cctacgcaca c 401
<210> 86
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; T=抗性等位基因
<400> 86
atcatcacta cttaatgata caaggaaatg ttttgccaaa ttgttcaaaa ctcaaatcac 60
taagtataga gcggaataat atatggataa ataagtaaca ttgggcatga cacataggca 120
aactagacca aaagggagca tgcaagttat acaagcattg cgcaatagaa ttacacggtg 180
tactagtgca tatgacctat wgggtagagt gtgagcgaaa gaactaaatc aaatgtagga 240
atggagtaga tgatcacatg aatacataat ttaagacatg tagactatat gacatgcaga 300
catataaaaa tgaactactg accagaaatc acattgagta agtgtattat atcgcataaa 360
taagtggaat aacatggcaa tgagtacaca tatgtaatat g 401
<210> 87
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; G=抗性等位基因
<400> 87
tcaaggaccc ttcgggcggt ccttagggat tcgtgactag tcgtttggac cctttatact 60
atatttgacc tatttaactt gtttttacaa gttttagact tcatatgaaa ctcccaaacc 120
tccaccacaa ttagggagtc tactacttcg atttagctat tttctagacg tcaatgtttt 180
tagttttcat tttggatcta rccttttcag taacatactt tattacatca aaataggctt 240
ggaaacatca ttttaaccat tatgtaaaag tcgacgcgca agtctaagtc atggaacttc 300
aagagtgtta cacttaagcc cccacacaac agtctatgac aataacaatg gaccagcagg 360
atggtccatt ggtaagttga tcttccttcg actgtatcgt c 401
<210> 88
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; C=抗性等位基因
<400> 88
cttggggttc tcattcactt tctacacaaa gaagatagga gggttcatcc tagtgaattc 60
cttcaagcgg gaagccgtgg tactagcatt ttgttaaaca cggagtccaa catccctatg 120
tacttgaacc tccatggctg cgacttgtga tttggcagtc atagcttgat ccaaagtaaa 180
aaatgtggac catatatctt maatcatgaa aggtggagga atgaccgaag gttgttcttg 240
aggaggcact tagttatatt gtggagggac ttggttttct tggggaggaa atccctcctt 300
agacaagtct tccccaactt ttctttcagc tgcacgtgta gtagtcatat cctatatcca 360
caaaaaaaat gattagaagg aaaaagacat gagaggattt t 401
<210> 89
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; G=抗性等位基因
<400> 89
actttgaatg ccttgtacgg accataggac cttgtatagt ctgtacttgc ccttgtttaa 60
acttgtatga gcacactctt ttgcactttt attttcaact gaacttttaa tttcggaggt 120
gctacaaaag gctcattcat ataattattt attcactcaa atgaaatgaa tgaacaattt 180
ttttaatcaa taccattagc rctatatatc agtcggttca gctaagtttt gaagtttatc 240
gatttgactt atcgataatc agtttgtaga tattctaaac cattgagata ttttcattag 300
tgcggttatc aatttatcaa ttaagtgtga cataaacaac aatctttgag aaattgcata 360
tgagttgaga aattgcactt gctaagacca tctccaacct c 401
<210> 90
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 90
ctagttatca gtgtgaagca tgccatatct ataatttagg agactacaac actgcacaaa 60
atttctagct tctttcacac tcatttcgtg cattagagtt tgacctccga aaagtttctt 120
cctaatacgt gcattcgtgt gtttttaagt taatcatgcc tccacaaaga gcacttaaag 180
gttgtcctcc taggataaat rttgaggatt aaggggtaac caatgcacct gaagtgcaac 240
cccttgtaaa agttaacaat gttgagttct ctaagtctat ctcgatgtat agtaaaggtg 300
ttactaacca tgtttggaaa aaatgagggg ctagaaagga tgtagcgcaa cttcggggat 360
acgagagttc tagaggataa atccttcaag tttcactcta a 401
<210> 91
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; T=抗性等位基因
<400> 91
aaagtttctc taaatttgta actacaaaga aaggaaactt tgggtaagga aactgacatc 60
ttttacatct ctgcttatta gttaaagagt tacaactata gattcacaca gacatttaat 120
ttgaagtgtt ccactttgtt taagaacata agtcatcaat aatcagtagt ggatatatct 180
acagaatgca ataatttgtt wactttgtga caggtctatt tgaaactcca agcacatgct 240
gcagcaagaa tgggaaataa tgtcaagaac tcatcagctt tcctaaagtc tccaacttct 300
acattacttc atacaattag tgaatctgag aaggcatcat atgttgccca tattaacaac 360
tatctcgtgg gagatgaatt tttgaaaaaa ttccttccta t 401
<210> 92
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 92
atttggttat cttatcttaa tgtatccttg ctaatgccat acattgacag aagaggtgct 60
caaacatgat tttcagcaat tattttgcta atcaagaact tggtttttgc tcaatatgca 120
aagagtcctt tacactttgt cctcacgtct ttgttacgat gtattaatac caatccattc 180
atgtgaacga tttctcaaat rtgtccacca tatctctctc tcaatgaatg acaacgaata 240
tgtattgaac atagtaggac ttagaagttt taaagggaat tagtttggca acctctcgcg 300
aactacaatt atcagggaac tacatgcacc aaggcaaaac agatccaaag aatgactgaa 360
tgttttttcg ctagtgagac cttagggtta tcaacaattt c 401
<210> 93
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 93
aagatagcct agtagtggta agttcctttg aaactatgtg tgacttccat ggtttaaatc 60
cccctggcta gaattttact ttttttagtt tttttttttt taagttccaa ttccgttttt 120
acagtttttc ttttaaaaat taaaacatgc taaaagtgtg gttttaaatc caaaagaaat 180
caagggtcac atttcttggg raaaaaattg ttggcttatt tacacgttta tatttttatt 240
ttttgaatcc ctgaatgaaa atcctgggtc tgccactgct aggtgtgtgt ttatcgacgc 300
aagtcctaaa aaaattacta ctgctttcaa aactaataag ccagaaaaaa ctgaaccaaa 360
tggacggtaa ttctttagtt tggtttgatt tggttttaga a 401
<210> 94
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 94
gttttttggt tcatctattg gtgtatttat agtgtatata ctggtagaat tgttaaattt 60
aggtcatata taaacaccaa ctagcatgat agttttttgg tttatctatt tgtgtattta 120
atgtataatc tggtgtagtt gatatattta aggtcataca tagtcaccaa ctaacatggt 180
agtttttatg ttcatcgatt rgtgtatttg aggcatctac attgattcag agaaaaaggg 240
cttgagttgc agcaacttct attcttaatg gacaacaggc tatcacagat gatagccttt 300
ttactaggaa ttcagatagt gtattttttg gtgtagttga tatatttagg tcgtatataa 360
acatcaattt tcatggtata tttttggtcc atccattggt g 401
<210> 95
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 95
ttcttgtcat atctatagtc tatagttata tgaacaaaag agttacagaa ctccacttct 60
ctacaatata ggatttacaa gatccctact taatgttttt gcttaaggtc ttttcttggt 120
cttcgacttg ctctgtgcgt catctgtgag attgatcttg actctctatt ggcggcttgt 180
ggatcattct caggtaactt ragttggcgg ttcaaactct aaaataaaca tcacaaaatg 240
acgaatctga atgggatgtg ttttttttag gaaaagaaag atacgagaat ccggaactaa 300
gacgtataat cctagattga aaaggagaaa taaggtgtat tgccttgaga gtttcaaatg 360
agacgtattg tcattgtggg ttgacctacg attatgttat c 401
<210> 96
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; C=抗性等位基因
<400> 96
agcaaccaca accgatgagc ttagacagca aagcatagtt gtcttaagca gacaagtctt 60
actttcatga cgccttgtta gttgtttagg agtgttagtc taagctatta gaacaacttg 120
atctgtgtca gtcatagcca tcaaaaaacg cttgatctct tttgttgaat cctatttgca 180
tgaggttaac tttgacacct scagctgaat tattgaagct cttctacatg tttaggctta 240
atgttgttga tatcttaatg gaccttatga aactgctttg tgctaggctg tggatgaagc 300
ggaagaaaat gtttttctgt tttctcgtgg tatcaaaaat ttgtgggtcc aaggaaaaaa 360
ataatgtgat gttctgcgtt cgattctctc tattttgagc a 401
<210> 97
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; G=抗性等位基因
<400> 97
ataccaagtt ttacacgccc tcaacattca ctttggatgc gactgataat caagaactat 60
tgttgaccac aagcgagccc ttggcgtggc ttaattctta gaaggagact tactcatgat 120
aagattattt taaaagtaac aataactcaa atgataactt agaattttta gaaataaaaa 180
ttcaactacc caaagtggta rctcaagtaa acataaaata aatgaaaagg agaagacttt 240
agaactagta tctatatatc tatctttgaa atctctaaat actatgagag atatagggac 300
aagactacca accaccctaa caaaactgaa taacaaaaca taataaatgg atcttccgaa 360
aatcaaggag gctcatcaaa tgactctgaa ctcgactgga t 401
<210> 98
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> C=易感等位基因 ; A=抗性等位基因
<400> 98
actagaggct ttatgaggag tcacccgaat caatggcacg atcctatcct acggtggttg 60
tgtagttatg gggtataatt tatctaaaca ctttcctaac tccgtattca atacaaaaac 120
tctcaaatca atgcaatgat tctcaataac tgtataaggt tccatgttag aaaaaaatgt 180
cccaaggaaa aattaagaat mtctatcttg aattacaact caatttaggt tcgactctct 240
gagtgttggg aagtgatctt ttgggctatt gtagtaatcg gggaacttcc acggatttat 300
ggattatgta ggtaaatggg gccaaagact acatttgagt tcctcaggat agtttgagcc 360
aatgtggtgt aataattttc ctcgattcta atgttgttag t 401
<210> 99
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; T=抗性等位基因
<400> 99
attatcatac cagtgttgtt gaaactgatt tttgaaatga taatttcaaa taggacaaat 60
attttttaat gacgatttag acttgtcttg taaaaatata atgttgattt ttgatgcatt 120
attttggaaa aggggcattg ataatatatg agaagtacca taattttgat actaaattag 180
tggatttttt gtggcactaa wggggtacaa tgatgtaaaa tgaatttcaa aagtcattga 240
tggatatgtg tctatcctta acagtgtcat aagatgaaaa tagctaagtt tttgtttagg 300
agtagactga gaacatttgg aaataattgt gcatctagcc acatatataa cattatcaca 360
ccaaattaag agtgtccacg atttgtttca caaacagatc c 401
<210> 100
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; G=抗性等位基因
<400> 100
actcattcaa caccatcgtc cttcatcagc ttctactttt gagcgtactc atacttttgc 60
gacccttcac tctaaatttc cttataattt cttttgcata cttcttctgt caaatgaata 120
tttcatcttg gatttgttta acctccattc caagaaagaa agtcatctct ccgagatccg 180
tcatttcaaa gactttcatc rtctcctttt tcaactcttc tatgagcaca acattactcc 240
tcttactaaa agatcatcaa catacaaaga aattataatg atgtcagtgt taaaaatgct 300
ttatgtaaag agttgaagca cttaggcttt tttcaaaacc taagtaagaa agatgatcat 360
cgattctact ataccaaacc cttggaactt gctttaattt g 401
<210> 101
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 101
ctaaggattc cgtacgactt tacagaattg aattcgggtg ggtataactt tcgaaattga 60
tcttagagtg ttttctgagt aacgttggtt agtggtgcat tgtgaaatat ggaattgatt 120
aaatatatca ctttttcgtg taagcctccc tagcactgga ttttatcaat ctggcgggac 180
cattgtggtg gatcagatgc ractttaagt tgtaaatgaa ccccaaatgc ctttattctg 240
tacttttcga ctttggaatc taaggaatgg ccctgcaact ttttgatagt ttcccaccat 300
tcttaagtct aaaggtaagt gttcacctcc caaatccatt tttctatttg taaaacctat 360
tctcttgggt tattatcaat gggggaggat tttttaattg g 401
<210> 102
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; G=抗性等位基因
<400> 102
gttactttaa tatttaattg tattataata aattctatat actaagggat gcagttattt 60
tataattttg agaaaataca tggcatattc gttcatctta cttgtgacct tttttttaat 120
agattatagt acattcctta ttaaatgggt taaagctatg atctggaaat ctccatgctt 180
aacgtctgta tgttttcaat rctatggtca aacaatttgc tcatataatt tgtgtacgtg 240
ttatgaattt gggatttttc atatgagtaa aatagttttt aaaaaaaatt ctgttatgtc 300
cttgtgggca taggttcaac cttgaagtaa atttagggat cataataatg ggtgttgtta 360
attttgaaat cttattatgg tcatttgttt gagtagattg c 401
<210> 103
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> T=易感等位基因 ; A=抗性等位基因
<400> 103
tgcaacttct attcttcctt ccgcaggtgg gagctgtgta ttttatttat cttgtctagt 60
tgtactctta ttagcttttg tacttgttta actagatcca tgagagttga taaattactt 120
tacaagtttt aagtaaatgt gtgtctaaag atattattta taaaatcata ttattgcaat 180
tacttctgtt tccttaaaaa wtttcacatg tttaattatt gggatacgag agttctctta 240
cttaggtgtt agagtggtgc ctggacggct cggtgggttg gatcgtgaca taatatatga 300
atttgtcaaa aaaatttaca ttatttttgt gatcgctatt tcaatataat atacagtaag 360
catgcatata tgtacaatgg atatattttt gtcatgctct a 401
<210> 104
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> C=易感等位基因 ; A=抗性等位基因
<400> 104
tgcgaggcac tcgccttgat ggttcttact acacttgggg caagtggggt aagtcttggt 60
gcttgaaaca cttccctaag acttagattc tggtgcccta accttctgat catacatgtt 120
cttagaggat ggaatactag ttatcatagt tctgctatgg ggttgaaaac ttttcctgac 180
tctacgagcg atttccacca mcaaatttct gctaagagta gtcatagttc ccagtcctaa 240
cctcttattc tccttggctt gtttcctaag cttatcaccc tcaacctgct gatcatgagt 300
cataagctta aggatgttca tatctcgtag taacatagaa cttctgcact cagttttcac 360
caaattttga ctccgtacaa gaaattattc atctgagccc t 401
<210> 105
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> C=易感等位基因 ; A=抗性等位基因
<400> 105
agtacaaatt gcaacttttt caatgttaac gtctgctttt ttagtgtata aaaaagaagg 60
attcatgaac tcctcagtca tattctattt taaaatactt aaacatcttt tttaattatg 120
tctcaagcat gtcaaacatc taatgcaaaa aagacttacg tttgtcgttg tggtaattta 180
gctattttga ggatttcaca maccgatacg aacccagatc gataattatt taattgtgca 240
attggtgcat tgatcttttt ttaagtggct tgattatgat tcatcaacga gcaacccatc 300
aaaattgagt ctcggattcg agacatcaaa ttttcaaagc gttatcaaat ttcaaatatt 360
ctttatctcc ttattttatc tcatgtacaa cttgattgac t 401
<210> 106
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> C=易感等位基因 ; A=抗性等位基因
<400> 106
tcatcgtagt cagtaagatt ttaccaagga attaaaattt attaagtatt aagctcatta 60
cattagaatt agcaccaaga tttccaaaaa gaactaaatt ggaattagca aaaatctgaa 120
atttgaagag ttaagttaag tatgagtttt gagtcaactt caaaggacca taaatattag 180
aacacgatga gttagatgtg mtacaagata ccatagggaa gatattcgaa taacctttgc 240
aaagcctttg agtttactaa gtttcaagtt catacaagtg agatatgacc ttttgaagtt 300
aggttctcca gttaagcaaa gttagccaaa aatagtgaag gtattttggt ccttccctta 360
cccaaacaga tttaattcgt tttagtcata ttttagggtt a 401
<210> 107
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> C=易感等位基因 ; G=抗性等位基因
<400> 107
aaaacaattt caattatgta gaatcatttt tcggccacat taaatcatat ttttatgtca 60
ttgaaccttg agaaaatact catccatatc aattattaaa agtttaattt atctatacgc 120
tatatttcaa ttaacaataa cgacatgcac gaattaaatt tttaacaaat gacataaatg 180
tgtcttgtct caaaattcga sgagatcttt cgttttcttt aatattcatg aaaaaacatt 240
gatccaaagt ggaaagttaa ggatgagttg gacatttttc acttaaaata atgtaacgtg 300
actccctcca ttttccccaa acgtaagtcc aagtcaactt gagacaccat attattataa 360
taaaaataga acaaacacaa atccataaat ttacaacttg a 401
<210> 108
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 108
ttccaatggg agaaacttga gaggaagtga aaatgtggtg gagataaaca tggttatgga 60
gagacatttg aggcaaaata tgagagaaat atttgagaga ttttagggga ttaaataagt 120
gagagagaat attaattatt aattattaat aaagaaatgc caaatttatt tacacaaaac 180
gtgtaaaatg gacattttgt rtcaatcttt taaaatctgg ctaatggatg ctataaattg 240
aagtctgagt tgacatgcta tgtcattttc ctcaatgggc tgttggagaa gagaattggg 300
ttcaatatgg gagacaaaac ttggctagaa ttgagttaat tactaagttt ggttaaaaat 360
taaataagat aaataaatta atgttactta attatgggaa a 401
<210> 109
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> C=易感等位基因 ; A=抗性等位基因
<400> 109
cgtggacggt actcgacgac cattgttggt ccctaagcga acccttatcc tggctgacta 60
caagcggaag actaacttaa gcataccaaa aatcttaaaa ctgaaatgaa caacttagaa 120
taaaataatc tatttaacta gtcattaaat aggcaacttg agtctttaaa acatttaata 180
aaataaatta ttaataaaaa mttctcaact agactatcta tggagcctct aactgaaaaa 240
gatggaaatc gggacaccac gacatcttga ttaaactgaa aagtaaataa ggtcctccgg 300
aaacaaggag gttcaacgac taactcgaac tctaggatgt attaataaag ctcttgttaa 360
tgatcccgaa gacctgtgtt tgcatcatca attatgcaac c 401
<210> 110
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> G=易感等位基因 ; A=抗性等位基因
<400> 110
agcttcctta cttatttaaa tgtgaaaaca tatcatttgg ggatacttag ttcccctata 60
cttttgaaga aaagaacttg aacttaactc tttactcttt acttagcttg aaacttgact 120
cttaaggaat atttagttcc cttatatttt ttgagaaatg cactcactct ttgctctttg 180
ctctatttga aacttaactc rtagggaata cttaattccc ttataaattt tgagttttaa 240
actcaaatca ttactctttt ctcaacttgt agtttaagcc ttaaaacaaa gtttaaaatg 300
ttgtttaaga ctctttaaaa tagtaagaac ttactttgac tttcttttta tttgtaagac 360
ttgattctta aattctttgg ctttgatatt aactttcctt g 401
<210> 111
<211> 401
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (201)..(201)
<223> A=易感等位基因 ; T=抗性等位基因
<400> 111
tatatcagaa tgatcaaata atcgcgtctt tagaatttta taggcatcat tgttgctgct 60
aagatatttt gacctaatga tcaaataatc gcgtctttag aattttatag gcatcattgt 120
tgctgctaag atattttgat aggtcaaatg atgtaatagg gtttttgtag aggccagatc 180
gtaaattggt gaaggcaaaa wcagaaactg tacaatatga ttttgataga gatcagttct 240
ttttcttgag atgttatgga ggattcatct attaggatct agtacaagga aaatttgtta 300
tgtattgaag tatttgttga gaacattttt gaattacctt gtgtaagttt aagattatga 360
agaattcttt tctttttctt tctactagat gggtaagcac g 401
<210> 112
<211> 301
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> allele
<222> (151)..(151)
<223> 易感等位基因 = C; 抗性等位基因 = CT
<400> 112
agtgatattg tcttcagcta tagaatatta tccagcccct ctagctttgg tcgatttttt 60
tttataacaa attttatcca agctatcatt gttttttttt ttttttcaat ttacagcaat 120
tgtttcccga ctagcctatc aatgttagag cttttttttg gtttgaacat cgttgtcttg 180
agaagcttaa tccatgttca ctgtaggcat tgttatttaa ttttcaattg agtttgttaa 240
tctgcctata gatattaatt tatttcctct tctttcgatg catatgatca atccagtcta 300
t 301
<210> 113
<211> 301
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> allele
<222> (151)..(151)
<223> 易感等位基因 = A; 抗性等位基因 = AT
<400> 113
acaatgacat gacatcaaaa ataatttaat ttatcatgtc aaatttgttt aaaaataaat 60
tgtaatcgag tgattttaaa gaataaagat gttaagttga gtgattttaa agacacaaac 120
taaaatttag tgactttaca agataatttc atatagttca ttgacctttt aagatattaa 180
ctcaaaaaat caataagtag tcacacacaa aaggtgattg caaagtaaat ttcaattttt 240
agcataagct tagtggggaa taccaatgac gaatctaaaa gtatcaatac aaaaaataga 300
a 301
<210> 114
<211> 301
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 多态性
<222> (151)..(151)
<223> T=易感等位基因 ; C=抗性等位基因
<400> 114
aaatgttaat ttttttattt ataataaata ttaatatgtc ttagacttga acttttaagt 60
tcattaaaga tagttaaata tcgtatttga tatacttaac gaaaaagcta aaacgtaaaa 120
gaaaatataa taagtttata tcttcttctt ytttttttaa atttggattt gtgatagtag 180
ggctttgaga gaaaattctt tttataattt tttttttcat attcaaggct cgaacctaag 240
atctatgatt aaggaagaag tagcctttcc actatacaat acacgttcat aatgagttaa 300
t 301
<210> 115
<211> 301
<212> DNA
<213> 番茄 (Lycopersicon esculentum)
<220>
<221> 等位基因
<222> (151)..(151)
<223> 易感等位基因 = G; 抗性等位基因 = GT
<400> 115
gcaaaaaaat aaaaaataaa aaaacgctag acagattgat gactatttat gctcatttca 60
aaacataact aatttgtatt taattttttt ttaccttttt tgattatttt ttttgctgta 120
tgattttctc ggttctatta ttagttattt gtttttttaa atattaatca taataacata 180
aatacataat taatccctta acaaaataaa ataatgacat ttttaataaa caataaaatt 240
gttgctaaga agtcccacta aaagcttaaa tattgttgtt ttgaaaaaaa tttccccaag 300
g 301
- 烟草花叶病毒组抗性番茄植物、烟草花叶病毒组抗性番茄植物的生产方法、番茄植物的烟草花叶病毒组抗性的赋予方法、烟草花叶病毒组抗性番茄植物的筛选方法和番茄植物的烟草花叶病毒组抗性的检测方法
- 植物体、植物体的制造方法、抗性赋予方法、番茄以及番茄的制造方法