通过全细胞生物转化产生甜菊醇糖苷
文献发布时间:2023-06-19 11:06:50
优先权
本申请要求2018年7月16日提交的美国临时申请号62/698,617的优先权和权益,所述临时申请特此以引用的方式整体并入。
背景技术
小分子的糖基转移酶由植物界中的大的多基因家族编码。这些酶将糖从核苷酸糖转移至广泛范围的次级代谢物,从而改变受体分子的理化性质。例如,甜菊醇糖苷是在甜叶菊(Stevia rebaudiana Bertoni)的叶中发现的一类化合物,甜叶菊是原产于南美洲的某些地区的一种菊科(Asteracae(Compositae))多年生灌木。它们的结构特征是单碱基甜菊醇,不同之处在于位置C13和C19处存在碳水化合物残基。它们积聚在甜叶菊叶中,占总干重的大约10%至20%。按干重计,在甜叶菊叶中发现的四种主要糖苷通常包括甜菊苷(9.1%)、莱鲍迪苷A(3.8%)、莱鲍迪苷C(0.6%-1.0%)和杜克苷A(0.3%)。其他甜菊醇糖苷以少量或痕量存在,包括莱鲍迪苷B、D、E、F、G、H、I、J、K、L、M和O、杜克苷B、甜菊双糖苷和甜茶苷。
较小的糖基化产物莱鲍迪苷M(RebM)的效力估计比蔗糖高约200-350倍,并且被描述为具有纯净的甜味,略带苦味或甘草余味。Prakash I.等人,
仍然需要经济的方法来产生作为天然植物提取物的次要产物的高价值糖苷。
发明内容
在各个方面和实施方案,本发明提供微生物细胞和用于由低级糖基化中间体产生高级糖基化产物的方法。所述微生物细胞表达一种或多种用于所述中间体的糖基化的UDP依赖性糖基转移酶。当将微生物菌株与包含糖苷中间体的植物提取物或其级分一起孵育时,这些中间体可用于通过微生物细胞进一步糖基化。在各种实施方案中,从培养基和/或微生物细胞回收所述高级糖基化产物。
糖苷中间体可以是任何糖基化的次级代谢物,如在植物提取物(包括糖基化萜类化合物、类黄酮、大麻素、聚酮化合物、茋类化合物和多酚等)中天然发现那些。在一些实施方案中,所述糖基化的中间体是糖基化的萜类化合物(如甜菊醇糖苷或罗汉果苷),并且可用作甜味剂。在一些实施方案中,所述产物的生物合成涉及微生物细胞中糖苷中间体的至少两种糖基化反应。
在一些实施方案中,所述糖苷中间体来自甜叶菊叶提取物。例如,RebM和其他高级糖基化产物可由甜菊醇糖苷中间体如甜菊苷、甜菊双糖苷、莱鲍迪苷A(RebA)和莱鲍迪苷C(RebC)等生物合成。所述微生物细胞在细胞质中表达一种或多种UDP依赖性糖基转移酶,以用于较低价值中间体的糖基化。当将微生物菌株与包含甜菊醇糖苷中间体的甜叶菊叶提取物或其级分一起孵育时,这些中间体可用于细胞进行进一步糖基化,并且可从培养基和/或微生物细胞中回收这些产物。因此,这种方法在甜叶菊叶提取物中使用高级中间体,即具有一至五种糖基化的甜菊醇糖苷(以及在一些实施方案中,糖苷配基核心,甜菊醇)。来自甜叶菊叶提取物的高级中间体可从甜菊醇糖苷的现有工业提取物中容易地获得。
在各种实施方案中,微生物细胞表达使糖苷中间体糖基化的至少一种或至少两种或至少三种或至少四种UGT酶,并且可选自表1中列出的那些和本文提供为SEQ ID NO:1-45的那些及其衍生物。在各种实施方案中,所述UGT酶独立地选自1-2'糖基化UGT酶、1-3'糖基化UGT酶和O-糖基化UGT酶。在各种实施方案中,这些酶在细胞内表达,因为它们不含膜易位或分泌信号。
在一些实施方案中,特别是关于甜叶菊叶提取物中的甜菊醇糖苷中间体的糖基化,所述微生物细胞表达四种UGT酶,如13-O UGT糖基化酶、19-O UGT糖基化酶、1-2'UGT糖基化酶和1-3'UGT糖基化酶。
在各种实施方案中,所述微生物细胞表达1-3'糖基化UGT酶。例如,所述1-3'糖基化UGT酶可选自SrUGT76G1、MbUGT1-3及其衍生物(例如,UGT76G1_L200A或MbUGT1-3_1、MbUGT1-3_1.5或MbUGT1-3_2)。
在这些和其他实施方案中,所述微生物细胞表达1-2'糖基化UGT酶。例如,所述1-2'糖基化UGT酶可选自SrUGT91D2、SrUGT91D1、SrUGT91D2e、OsUGT1-2、MbUGT1,2、MbUGT1,2.2及其衍生物。
在这些或其它实施方案中,所述微生物细胞表达C13 O-糖基化UGT酶。例如,所述C13 O-糖基化UGT酶可选自SrUGT85C2及其衍生物(例如,MbUGTC13)。
在这些或其它实施方案中,所述微生物细胞表达C19 O-糖基化UGT酶。例如,所述C19 O-糖基化酶可选自SrUGT74G1、MbUGTc19及其衍生物(例如,MbUGTc19-2)。
全细胞转化需要在细胞内表达的酶作用于外部进料的底物(例如,糖基化中间体),并且细胞提供UDP-葡萄糖辅因子再生。这与依赖于在细胞外纯化或分泌的酶的过程相反,所述过程需要外源性UDP-葡萄糖供应或UDP-葡萄糖前体或UDP-葡萄糖再生机制或UDP-葡萄糖再生酶系统。在本发明的实施方案中,用活的微生物细胞进行催化(糖基化)。UDP-葡萄糖辅因子再循环使用天然细胞代谢发生,而无需其他外部提供的酶或底物进料。
根据本公开,对微生物细胞的遗传修饰使得糖苷中间体可用于通过细胞内表达的酶进行进一步糖基化。在一些实施方案中,与从裂解细胞中提取相反,可从培养基中回收高级糖基化产物。在一些实施方案中,所述微生物细胞具有一种或多种遗传修饰,所述遗传修饰增加UDP-葡萄糖可用性。在一些实施方案中,不希望受理论束缚,这些修饰也可使细胞对葡萄糖可用性产生应力,从而导致内源性转运蛋白的表达增加,以将糖苷中间体输入细胞。在一些实施方案中,不希望受理论束缚,通过遗传修饰或培养基组分使细胞具有渗透性,从而允许产物和底物的被动扩散。
在各种实施方案中,所述微生物细胞具有一种或多种内源性转运蛋白的过表达(例如,与亲本微生物菌株相比),或在某些实施方案中,经修饰以表达重组和/或工程化的转运蛋白。在一些实施方案中,所述微生物细胞表达内源性转运蛋白或其衍生物的一个或多个其他拷贝。例如,转运蛋白的表达或活性可进行修饰以增加甜菊醇糖苷底物(例如,甜菊苷、甜菊双糖苷、RebA和/或RebC中的一种或多种)转运至细胞中,同时输出产物,如RebM和/或RebD以及其他高级糖基化产物。可过表达或工程化以改变微生物细胞中的活性或底物特异性的示例性转运蛋白包括大肠杆菌、acrAD、xylE、ascF、bglF、chbA、ptsG/crr、wzxE、rfbX以及其直向同源物或衍生物。其他转运蛋白包括选自转运所需糖苷中间体的细菌或内源性转运蛋白的那些。例如,转运蛋白可来自宿主物种,或另一种细菌或酵母物种,并且可被工程化以调节其对特定糖苷中间体或产物的亲和力。
在各种实施方案中,所述方法使得糖苷中间体至少40%转化、或至少50%转化或至少75%转化为所需产物(例如,甜菊苷、甜菊双糖苷和RebA转化为RebD、RebM和/或其他高度糖基化的莱鲍迪苷)。在RebM的产生中,与RebD相比,产品特征可能更偏向RebM。
所述方法可通过分批发酵、补料-分批发酵、连续发酵或半连续发酵进行。例如,在一些实施方案中,所述方法通过分批发酵进行,其中孵育时间小于约72小时,或者在一些实施方案中小于约48小时或小于约24小时。
在一些方面,本发明提供了用于制备包含高级甜菊醇糖苷如RebM的产品的方法。所述方法包括将目标甜菊醇糖苷掺入产品,如食品、饮料、口腔护理产品、甜味剂、调味剂或其他产品中。在一些方面,本发明提供了用于制备包含多种高甜度甜味剂的甜味剂组合物的方法,所述高甜度甜味剂如甜菊醇糖苷、罗汉果苷、三氯蔗糖、阿斯巴甜、纽甜、爱德万甜(advantame)、乙酰磺胺酸钾、糖精、环己基氨基磺酸盐、新橙皮苷二氢查尔酮、买麻藤素E和/或白皮杉醇4'-O-β-D-吡喃葡萄糖苷中的两种或更多种。
本发明的其他方面和实施方案将根据以下具体实施方式而是显而易见的。
附图说明
图1.RebM的结构。甜菊醇支架(二萜类化合物)显示为加框。RebM含有六种糖基化:(1)C13 O-糖基化,(2)C13 1-2'糖基化,(3)C131-3'糖基化,(4)C19 O-糖基化,(5)C19 1-2'糖基化,和(6)C19 1-3'糖基化。
图2.通过UGT酶对甜菊醇和甜菊醇糖苷中间体的作用产生的糖基化产物。
图3.通过UGT酶对含有鼠李糖的甜菊醇糖苷中间体的作用产生的糖基化产物。
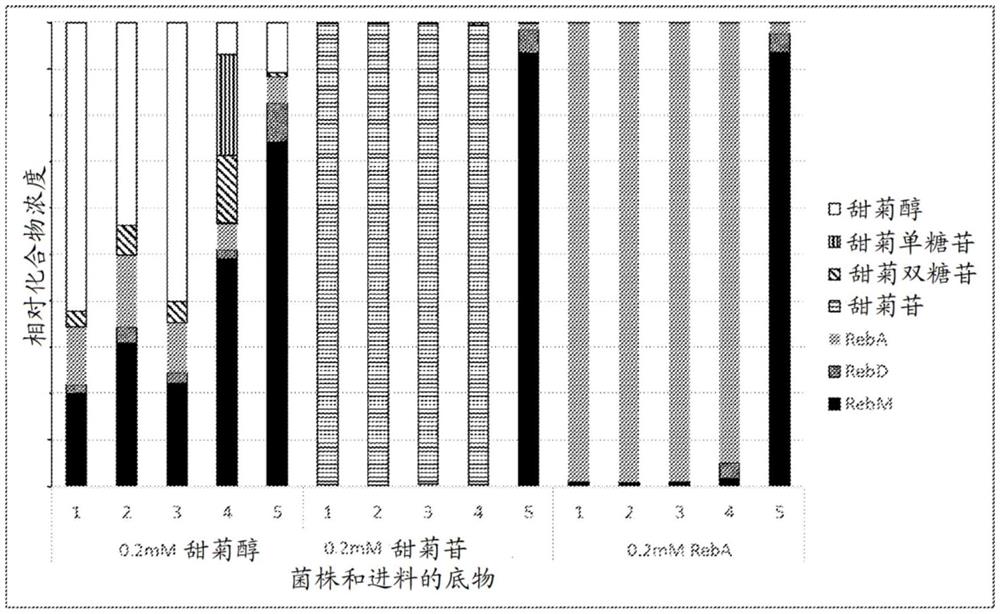
图4.甜菊双糖苷转化为RebM。对于RebM途径中的五种主要化合物,化合物浓度是相对于缺少UGT酶的对照菌株中这些化合物的总浓度示出。向菌株进料0.2mM甜菊双糖苷,并在提取之前与底物一起孵育48小时。菌株3和4显示没有甜菊双糖苷的转化,而菌株5显示实质性转化为RebD和RebM。
图5.甜菊苷和RebA转化为RebM。使用菌株1至5示出叶提取物中发现的两种市售中间体甜菊苷和RebA生物转化为RebM。还示出甜菊醇核心的生物转化。所述值报告为总甜菊醇糖苷组合物的百分比。所有菌株将甜菊醇转化为RebM表明,所有菌株都能够形成RebM。与甜菊双糖苷一样,甜菊苷和RebA两者均被转化为RebM,但仅在菌株5中。对于从甜菊苷和RebA转化,RebD:RebM的比例强烈有利于RebM(1:20)。
图6.96孔板中甜菊醇糖苷的生物转化。对于RebM途径中的五种主要化合物,在有或没有表达的UGT酶情况下示出化合物浓度。向菌株进料0.5g/L的甜菊苷/RebA叶提取物,并在取样前与底物一起孵育48小时。
图7.在生物反应器规模上甜菊苷/RebA叶提取物的生物转化。对于RebM途径中的五种主要化合物,在整个发酵过程中的不同时间点对培养基中的化合物浓度进行采样。向菌株进料2g/L的甜菊苷/RebA叶提取物,并与底物一起孵育总共30小时。
图8.遗传修饰以增加至UDP-葡萄糖的天然大肠杆菌通量。代谢物:G6P,葡萄糖-6-磷酸;G1P,葡萄糖-1-磷酸;6PGL,6-磷酸葡萄糖酸-1,5-内酯;6PG,6-磷酸葡萄糖酸酯;R5P,核糖-5-磷酸;PRPP,5-磷酸核糖基-1-焦磷酸酯;UMP,尿苷一磷酸;UDP,尿苷二磷酸;UTP,尿苷三磷酸;UDP-Glu,UDP-葡萄糖;UDP-Gal,UDP-半乳糖;UDP-GlcNAc,UDP-N-乙酰氨基葡糖;CTP,胞苷三磷酸;dUDP,脱氧尿苷-二磷酸;RebA,莱鲍迪苷A;RebM,莱鲍迪苷M;酶:1.glk,葡萄糖激酶;2.pgi,葡萄糖-6-磷酸异构酶;3.zwf,葡萄糖-6-磷酸-1-脱氢酶;4.pgl,6-磷酸葡萄糖酸内酯酶;5.edd,磷酸葡萄糖酸脱水酶;6.gnd,6-磷酸葡萄糖酸脱氢酶;7.rpiA,核糖-5-磷酸异构酶或rpe,核糖-磷酸3-差向异构酶;8.prs,核糖-磷酸二磷酸激酶;9.pyrEF,乳清酸磷酸核糖基转移酶和乳清苷-5'磷酸-脱羧酶;10.pyrK,尿苷酸激酶;11.ndk,UDP激酶;12.murG,N-乙酰葡糖胺基转移酶;13.nrdABEF,核糖核苷-二磷酸还原酶;14.pyrG,CTP合酶;15.pgm,磷酸葡萄糖变位酶;16.galU,UDP-葡萄糖焦磷酸化酶;17.galETKM,UDP-葡萄糖-4-差向异构酶/半乳糖-1-磷酸尿苷酰转移酶/半乳糖激酶/半乳糖-1-差向异构酶;18.ushA,5’-核苷酸酶/UDP-糖水解酶;19.UGT,UDP-糖基转移酶。所述示意图说明从菌株1至菌株5的顺序基因组修饰。菌株1是基础大肠杆菌菌株。菌株2缺失17,并且菌株3具有添加缺失18。通过缺失2从菌株3构建菌株4。通过过表达15和16从菌株4构建菌株5。
具体实施方式
在各个方面和实施方案,本发明提供微生物细胞和用于由低级糖基化中间体产生高级糖基化产物的方法。在各种实施方案中,所述微生物细胞表达一种或多种UDP依赖性糖基转移酶(例如,细胞内),以用于中间体的糖基化。在各种实施方案中,所述微生物细胞具有一种或多种遗传修饰,所述遗传修饰增加UDP-葡萄糖的可用性。当将微生物菌株与糖苷中间体(例如,来自植物提取物或其级分)一起孵育时,这些糖苷中间体可用于细胞,以通过细胞内表达的糖基转移酶进一步糖基化。在一些实施方案中,所述高级糖基化产物可从培养基中回收,或者在一些实施方案中,可从培养基和微生物细胞中回收。
糖苷中间体可以是任何糖基化的次级代谢物,如在植物提取物(包括糖基化萜类化合物、糖基化类黄酮、糖基化大麻素、糖基化聚酮化合物、糖基化茋类化合物和糖基化多酚等)中天然发现那些。在一些实施方案中,所述糖苷中间体是糖基化的萜类化合物,如甜菊醇糖苷或罗汉果苷。在一些实施方案中,所述糖苷中间体具有一个、两个、三个、四个或五个糖基,包括选自葡糖基、半乳糖基、甘露糖基、木糖基和鼠李糖基等的糖基。在各种实施方案中,所述糖基化产物具有至少四个糖基、或至少五个糖基、或至少六个糖基。在其他实施方案中,所述产物具有七个、八个、九个或更多个糖基。在一些实施方案中,所述产物的生物合成涉及微生物细胞对中间体的至少两次糖基化。
在一些实施方案中,所述糖苷中间体来自甜叶菊叶提取物。例如,可从甜菊醇糖苷中间体如甜菊苷、甜菊双糖苷、莱鲍迪苷A、杜克苷A、杜克苷B、莱鲍迪苷C和莱鲍迪苷F等生物合成具有五种、六种或更多种糖基化的甜菊醇糖苷(如RebD或RebM)。所述微生物细胞表达一种或多种UDP依赖性糖基转移酶,以用于这些较低价值前体的糖基化。当将微生物菌株与包含甜菊醇糖苷中间体的甜叶菊叶提取物或其级分一起孵育时,这些中间体可用于进一步糖基化为RebD或RebM或其他高级糖基化产物(例如,RebI),其可从培养基和/或微生物细胞中回收。
在各种实施方案中,所述UDP依赖性糖基转移酶“在细胞内”表达,因为所述酶不具有膜易位或分泌肽或结构域。因此,UGT酶的表达在细胞质中发生,并且这些酶不经由分泌或转运信号定向到细胞外。在各种实施方案中,所述UGT酶不含膜锚定结构域。即,在各种实施方案中,所述UGT酶不包含跨膜结构域。
在一些实施方案中,所述方法使用甜叶菊叶提取物中的高级中间体,即具有一至五种糖基化的甜菊醇(糖苷配基中间体)和甜菊醇糖苷,其可用于微生物细胞进行进一步糖基化。来自甜叶菊叶提取物的高级中间体可从甜菊醇糖苷的现有工业提取物中容易地获得。如表2所示,叶提取物可主要含有途径中间体甜菊苷和莱鲍迪苷A(RebA)。在各种实施方案中,甜叶菊叶提取物是甜菊醇糖苷的提取。在一些实施方案中,所述提取物包含甜菊苷、甜菊双糖苷和莱鲍迪苷A中的一种或多种作为主要组分。在一些实施方案中,所述提取物包含杜克苷A、杜克苷B、RebC和/或RebF中的一种或多种作为主要组分。主要组分通常占提取物或其级分中的甜菊醇糖苷的至少约10%,但在一些实施方案中,可占提取物或其级分中的甜菊醇糖苷的至少约20%、或至少约25%、或至少约30%。
RebM在图1中示出。RebM包含甜菊醇核心的六种糖基化:(1)C13 O-糖基化,(2)C131-2'糖基化,(3)C13 1-3'糖基化,(4)C19 O-糖基化,(5)C19 1-2'糖基化,和(6)C19 1-3'糖基化。尽管各种糖基化产物是可能的(图2),但是RebM可通过四种UGT酶的作用由甜菊醇合成,并且UGT糖基化酶各自能够作用于许多底物。例如,UGT91D2和OsUGT1-2两者均是1-2'糖基化酶,它们可由甜菊单糖苷(通过C13作用)产生甜菊双糖苷,也可以由RebA(通过C19处的作用)产生RebD。此外,UGT76G1是1-3'糖基化酶,可由甜菊苷(通过C13的作用)产生RebA,也可由RebD(通过C19的作用)产生RebM。
用于甜菊醇和甜菊醇糖苷的糖基化的UGT酶(包括用于RebM的生物合成)在US2017/0332673中公开,其特此以引用的方式整体并入。示例性UGT酶列于以下表1中(参考专利申请特此以引用的方式整体并入):
示例性UGT酶的氨基酸序列由本公开提供为SEQ ID NO:1-17。SEQ ID NO:1是甜叶菊UGT85C2。SEQ ID NO:13是具有P215T取代的SrUGT85C2,并且在第2位置插入Ala以增加稳定性。SEQ ID NO:2是甜叶菊UGT74G1(在第2位置插入Ala)。SEQ ID NO:8和12是基于SrUGT74G1的环状变换体(MbUGTC19和MbUGTC19-2)。SEQ ID NO:3是甜叶菊UGT76G1。具有1-3'糖基化活性的环状变换体公开为SEQ ID NO:10(MbUGT1-3)和SEQ ID NO:15、16和17(分别MbUGT1-3_1、MbUGT1-3_1.5和MbUGT1-3_2)。具有L200A取代和位置2处Ala的UGT76G1公开为SEQ ID NO:14。甜叶菊UGT91D1、UGT91D2和UGT91D2e公开为SEQ ID NO:4、5和6。水稻UGT1-2公开为SEQ ID NO:7。MbUGT1,2和MbUGT1,2.2是具有1-2'糖基化活性的环状变换体酶(SEQ ID NO:9和11)。
另外的UGT酶提供为SEQ ID NO:18至46,来自物种罗汉果(Siraitiagrosvenorii/monk fruit)、苦瓜(Momordica charantia/bitter melon)、黄瓜(Cucumissativa/Cucumber)、笋瓜(Cucurbita maxima/squash)、南瓜(Cucurbita moschata/squash/pumpkin)、碧桃(桃)、可可(Theobroma cacao/cacao)、圆果种黄麻(白黄麻)、枣(红枣)、葡萄(葡萄藤)、核桃(Juglans regia/walnut)、橡胶树(Hevea brasiliensis/rubbertree)、木薯(Manihot esculenta/cassava)、土瓶草(猪笼草)和小粒咖啡(咖啡)。可基于所需的产物和可用的中间体选择UGT酶并任选地进行工程化。
在各种实施方案中,所述微生物细胞表达至少一种、或至少两种、或至少三种、或至少四种UGT酶。在一些实施方案中,所述UGT酶使甜菊醇糖苷底物糖基化,包括表2中列出的那些。在一些实施方案中,所述微生物细胞表达使甜菊醇糖苷中间体糖基化的四种UGT酶,如13-O UGT糖基化酶、19-O UGT糖基化酶、1-2'UGT糖基化酶和1-3'UGT糖基化酶。这些通用类别的UGT酶对甜叶菊叶的糖基化中间体的作用描绘于图2和3中。
在各种实施方案中,所述微生物细胞表达1-3'糖基化UGT酶。例如,所述1-3'糖基化UGT酶可选自SrUGT76G1、MbUGT1-3_1、MbUGT1-3_1.5和MbUGT1-3_2及其衍生物。
在这些和其他实施方案中,所述微生物细胞表达1-2'糖基化UGT酶。例如,所述1-2'糖基化UGT酶可选自SrUGT91D2、SrUGT91D1、SrUGT91D2e、OsUGT1-2、MbUGT1,2、MbUGT1,2.2及其衍生物。
在这些或其它实施方案中,所述微生物细胞表达C13 O-糖基化UGT酶。例如,所述C13 O-糖基化UGT酶可选自SrUGT85C2及其衍生物(例如,MbUGTC13)。
在这些或其它实施方案中,所述微生物细胞表达C19 O-糖基化UGT酶。例如,所述C19 O-糖基化酶可选自SrUGT74G1、MbUGTc19及其衍生物(例如,MbUGTc19-2)。
在这些或其它实施方案中,所述微生物细胞表达1-3'糖基化UGT酶和1-2'糖基化UGT酶。在一些实施方案中,所述微生物细胞表达1-3'糖基化UGT酶、1-2'糖基化UGT酶和C13O-糖基化UGT酶。在一些实施方案中,所述微生物细胞表达1-3'糖基化UGT酶、1-2'糖基化UGT酶、C19 O-糖基化UGT酶和C19 O-糖基化UGT酶。在一些实施方案中,所述微生物细胞表达1-3'糖基化UGT酶、1-2'糖基化UGT酶、C19 O-糖基化UGT酶和C13 O-糖基化UGT酶。在一些实施方案中,所述微生物细胞表达SrUGT85C2或其衍生物(例如,MbUGTC13)、MbUGT1,2.2或其衍生物、SrUGT74G1或其衍生物(例如,MbUGTc19或MbUGTc19-2)以及SrUGT76G1或MbUGT1-3_1或其衍生物(例如,76G1_L200A、MbUGT1-3_1.5或MbUGT1-3_2)。不受理论束缚,工程化的UGT酶可提供增加的至RebM的碳通量(以及较高的糖基化产物),并且尤其是由于底物结合袋能够更好地容纳更多的底物,而对较低糖基化中间体的活性没有实质性损失。在这些实施方案中,UGT酶(如1-3'和1-2'糖基化UGT酶)可具有增加的活性速率(例如,底物结合和周转速率)与更高度糖基化的甜菊醇底物(如RebA或RebD)。
UGT酶的衍生物通常包含与SEQ ID NO:1至46中的一个或多个具有至少约50%同一性、至少约60%同一性、至少约70%同一性、至少约80%同一性、至少约85%同一性或至少约90%同一性、或至少约95%同一性、或至少96%、97%、98%或99%同一性的氨基酸序列。在一些实施方案中,相对于SEQ ID NO:1至46中的一个,所述衍生物具有1至20或1至10或1至5个氨基酸修饰(独立地选自氨基酸取代、插入和缺失)。在一些实施方案中,例如关于由甜菊醇糖苷中间体产生RebM和其他高级甜菊醇糖苷(例如,具有至少5个糖基),这些UGT酶的衍生物包含与SEQ ID NO:1至17中的一个或多个具有至少约50%同一性、至少约60%同一性、至少约70%同一性、至少约80%同一性、至少约85%同一性或至少约90%同一性、或至少约95%同一性、或至少96%、97%、98%或99%同一性的氨基酸序列。在一些实施方案中,相对于SEQ ID NO:1至17中的一个,所述衍生物具有1至20或1至10或1至5个氨基酸修饰(独立地选自氨基酸取代、插入和缺失)。
在一些实施方案中,所述微生物细胞表达1-3'UGT酶,所述1-3'UGT酶包含与SEQID NO:15、SEQ ID NO:16或SEQ ID NO:17的氨基酸序列至少约75%相同的氨基酸序列。在一些实施方案中,所述1-3'UGT包含与SEQ ID NO:15、16或17至少约80%相同的氨基酸序列。在一些实施方案中,所述1-3'UGT酶的氨基酸序列与SEQ ID NO:15、16或17至少约85%相同,或与SEQ ID NO:15、16或17至少约90%相同,或与SEQ ID NO:15、16或17至少约95%相同,或与SEQ ID NO:15、16或17至少约98%相同。在一些实施方案中,所述1-3'UGT酶的氨基酸序列包含SEQ ID NO:15、16或17的氨基酸。
例如,相对于氨基酸序列SEQ ID NO:15、16或17,所述氨基酸序列可具有1至20个独立地选自取代、缺失和插入的氨基酸修饰。在一些实施方案中,相对于SEQ ID NO:15、16或17的氨基酸序列,所述氨基酸序列具有1至10个(例如,1至5个)独立地选自取代、缺失和插入的氨基酸修饰。对SEQ ID NO:15、16或17的氨基酸序列的氨基酸修饰可通过可用的酶结构和同源性模型的构建来指导。示例性结构描述于例如Li等人,
在一些实施方案中,1-3'UGT酶在对应于SEQ ID NO:3(甜叶菊UGT76G1)的位置29、200、357和414的位置处包含氨基酸取代。包含在SEQ ID NO:15和17的酶中的这些位置的取代(分别在SEQ ID NO:15中的位置183、354、54和111)可提供活性的显著改进。在一些实施方案中,在对应于SEQ ID NO:15的位置183、354、54和111的位置处的氨基酸的同一性允许在其他位置处的进一步修饰。例如,在一些实施方案中,1-3'UGT酶包含与SEQ ID NO:15或17的氨基酸序列至少约60%相同的氨基酸序列,其中所述UGT酶包含:在对应于SEQ ID NO:15的位置54的位置处的甘氨酸(G)或苏氨酸(T);在对应于SEQ ID NO:15的位置111的位置处的亮氨酸(L)或异亮氨酸(I);在对应于SEQ ID NO:15的位置183的位置处的甲硫氨酸(M)或亮氨酸(L);以及在对应于SEQ ID NO:15的位置354的位置处的丙氨酸(A)、甘氨酸(G)或丝氨酸(S)。在一些实施方案中,1-3'UGT酶在对应于SEQ ID NO:15的位置183的位置处包含甲硫氨酸(M)。在一些实施方案中,1-3'UGT酶在对应于SEQ ID NO:15的位置54的位置处包含甘氨酸(G)。在一些实施方案中,1-3'UGT酶在对应于SEQ ID NO:15的位置111的位置处包含亮氨酸(L)。在一些实施方案中,1-3'UGT具有以下中的两个或三个:在对应于SEQ ID NO:15的位置183的位置处的蛋氨酸(M)、在对应于SEQ ID NO:15的位置54的位置处的甘氨酸(G)以及在对应于SEQ ID NO:15的位置111的位置处的亮氨酸(L)。这些修饰可提供对酶的活性的显著改进。
在一些实施方案中,1-3'UGT酶在对应于SEQ ID NO:15的位置155的位置之后包含5至约15个氨基酸,如6至12个氨基酸或约6或约11个氨基酸的插入。在一些实施方案中,插入是柔性和亲水性序列,所述序列主要是甘氨酸和丝氨酸残基。在一些实施方案中,所述序列是GSGGSG(SEQ ID NO:47)或GSGGSGGSG(SEQ ID NO:48)。
在各种实施方案中,相较于UGT76G1-L200A(SEQ ID NO:14),1-3'UGT酶显示出改进的甜菊苷转化为Reb A,和RebD至RebM的改进的转化。这种改进的转化在生物转化测定中得到了体现,其中将甜菊苷或RebD底物进料至表达1-3'UGT酶的微生物细胞。在与含有重组表达的1-3'UGT的细胞裂解物反应,或与纯化或部分纯化的1-3'UGT的体外反应中可证明改进的转化。
对酶的氨基酸序列的变化可改变其活性或没有可测量的影响。没有可测量的影响的沉默变化通常是保守性取代以及远离活性位点和底物结合位点的溶剂暴露表面上的小插入或缺失。相比之下,酶活性更可能受到非保守取代、大的插入或缺失以及活性位点、底物结合位点以及对于蛋白质折叠或构象重要的掩埋位置处的变化的影响。改变酶活性的变化可增加或降低反应速率或增加或降低对特定底物的亲和力或特异性。例如,增加底物结合位点大小的变化可允许酶作用于更大的底物,而使催化氨基酸侧链更靠近底物上的靶位点定位的变化可增加酶促速率。
酶的三维结构和相关活性位点、底物结合位点和其他相互作用位点的位置的知识可有助于衍生物的合理设计,并提供对特定变化的表型的机械洞察力。植物UGT具有高度保守的二级和三级结构,同时具有相对低的氨基酸序列同一性。Osmani等人,
与已知结构相比,UGT衍生物的构建可基于同源性建模进行指导。例如,根据SrUGT76G1_L200A的晶体结构分析和SrUGT76G1至MbUGT3-1_1的氨基酸序列比对,可预测甜菊苷的甜菊醇核心与MbUGT3-1_1的以下残基接近(在
所述UGT酶中的一者或多者的氨基酸序列可任选地包含在2位插入或取代的丙氨酸以降低细胞中的周转。在各种实施方案中,一种或多种UGT酶包含在位置2插入或取代的丙氨酸氨基酸残基,以在体内提供额外的稳定性。
氨基酸序列的同一性,即序列同一性百分比,可经由序列比对来测定。此类比对可用几种本领域已知的算法,诸如用Karlin和Altschul(Karlin&Altschul(1993)Proc.Natl.Acad.Sci.USA 90:5873-5877)的数学算法、hmmalign(HMMER软件包)或CLUSTAL算法(Thompson,J.D.,Higgins,D.G.和Gibson,T.J.(1994)Nucleic Acids Res.22,4673-80)进行。可使用例如BLAST、BLAT或BlastZ(或BlastX)来计算序列同一性(序列匹配)的等级。将类似算法并入到Altschul等人(1990)J.Mol.Biol.215:403-410的BLASTN和BLASTP程序中。BLAST蛋白质比对可使用BLASTP程序进行,得分=50,字长=3。为了获得空位比对用于比较目的,如Altschul等人(1997)Nucleic Acids Res.25:3389-3402中所述利用空位BLAST。利用BLAST和空位BLAST程序时,使用各个程序的默认参数。
UGT酶可整合到微生物细胞的染色体中,或者可替代地在染色体外表达。例如,UGT酶可从细菌人工染色体(BAC)或酵母人工染色体(YAC)表达。
可使用例如基因模块(例如,操纵子)或UGT酶的独立表达调节UGT酶的表达以获得最佳酶活性。例如,基因或操纵子的表达可以通过选择具有不同强度(例如强、中或弱)的启动子诸如诱导型启动子或组成型启动子来调节。具有不同强度的启动子的几个非限制性实例包括Trc、T5和T7。另外,可以通过操纵细胞中基因或操纵子的拷贝数来调节基因或操纵子的表达。在一些实施方案中,所述细胞表达每种UGT酶的单个拷贝。在一些实施方案中,可以通过操纵模块内基因的顺序来调节基因或操纵子的表达,其中先转录的基因通常以更高的水平表达。在一些实施方案中,通过将一个或多个基因或操纵子整合到染色体中来调节基因或操纵子的表达。
对UGT表达的优化也可通过选择适当的启动子和核糖体结合位点来实现。在一些实施方案中,这可以包括选择高拷贝数的质粒,或单拷贝数、低拷贝数或中拷贝数的质粒。通过引入或消除诸如茎环之类的结构,也可以靶向转录终止的步骤进行基因表达的调控。
全细胞转化需要底物(例如,糖苷中间体)可用于细胞通过所表达的酶(例如,细胞内表达的酶)进行糖基化,并且优选可从细胞外培养基中提取产物。全细胞系统具有优点,因为所述细胞提供UDP-葡萄糖辅因子再生。这与依赖于使用来自细胞外溶解或分泌的酶的过程相反,所述过程需要外源性UDP-葡萄糖供应或UDP-葡萄糖前体或UDP-葡萄糖再生机制或UDP-葡萄糖再生酶系统。在本发明的实施方案中,用活的微生物细胞进行催化(糖基化),并且使用细胞代谢来进行UDP-葡萄糖辅因子再循环,而无需酶补料或底物补料来进行UDP-葡萄糖再生。
US 2017/0332673描述过表达MEP途径酶连同下游的甜菊醇生物合成途径的以及UGT酶以驱动由葡萄糖产生RebM的大肠杆菌菌株。然而,这些菌株不进行进料的甜菊醇糖苷中间体至RebM的生物催化,这可能部分是由于宿主细胞无法接近甜菊醇糖苷底物。根据本公开,对微生物细胞的遗传修饰使得糖基化中间体可用于全细胞系统中的进一步糖基化。此外,可从细胞外培养基中回收产物,这有助于产物的下游纯化和加工。
在一些实施方案中,所述微生物细胞具有一种或多种遗传修饰,所述遗传修饰增加UDP-葡萄糖可用性。在一些实施方案中,不希望受理论束缚,这些修饰也可使细胞对葡萄糖可用性产生应力,从而导致内源性转运蛋白的表达增加,以将甜菊醇糖苷输入细胞。指数生长的大肠杆菌中的野生型UDP葡萄糖水平是约2.5mM(Bennett BD,Kimball EH,Gao M,Osterhout R,Van dien SJ,Rabinowitz JD.
在一些实施方案中,微生物细胞具有编码消耗UDP-葡萄糖的酶的基因的缺失、失活或活性或表达降低。例如,微生物细胞可具有微生物物种中ushA(UDP糖水解酶)和/或galE、galT、galK和galM(其负责来自UDP-葡萄糖的UDP-半乳糖生物合成)或其直向同源物中的一者或多者的缺失、失活或活性降低。在一些实施方案中,galETKM基因失活、缺失或表达显著降低。或者或另外地,微生物细胞具有微生物物种中大肠杆菌otsA(海藻糖-6-磷酸合酶)或其直向同源物的缺失、失活或活性或表达降低。或者或另外,微生物细胞具有微生物物种中大肠杆菌ugd(UDP-葡萄糖6-脱氢酶)或其直向同源物的缺失、失活或活性或表达降低。降低或消除otsA和ugd的活性可分别将UDP-葡萄糖槽除去或还原为海藻糖或UDP-葡萄糖醛酸。
其他可减少或消除的UDP-葡萄糖槽包括消除或降低负责脂质糖基化和LPS生物合成的基因以及负责使十一异戊烯二磷酸(UPP)糖基化的基因的活性或表达。参与使脂质糖基化或LPS生物合成的基因包括大肠杆菌waaG(脂多糖葡糖基转移酶1)大肠杆菌waaO(UDP-D-葡萄糖:(葡糖基)LPSα-1,3-葡糖基转移酶))和大肠杆菌waaJ(UDP-葡萄糖:(葡糖基)LPSα-1,2-葡糖基转移酶))。负责使十一异戊烯二磷酸(UPP)糖基化的基因包括大肠杆菌yfdG(推定的细菌萜醇连接的葡萄糖转位酶)、大肠杆菌yfdH(细菌萜醇葡糖基转移酶)、大肠杆菌yfdI(血清型特异性葡糖基转移酶)和大肠杆菌wcaJ(十一异戊烯-磷酸葡萄糖磷酸转移酶)。这些基因产物中的一种或多种(或微生物细胞中的相应直向同源物)的缺失、失活或活性或表达的降低可增加UDP-葡萄糖可用性。
在这些或其它实施方案中,微生物细胞具有编码消耗UDP-葡萄糖的前体的酶的基因的缺失、失活或活性或表达降低。例如,在一些实施方案中,微生物细胞具有宿主细胞的微生物物种中pgi(葡萄糖-6磷酸酯异构酶)或其直向同源物的缺失、失活或活性或表达降低。
在这些或其它实施方案中,所述细胞具有一种或多种编码参与将葡萄糖-6-磷酸转化为UDP-葡萄糖的酶的基因的过表达或活性增加。例如,可过表达或修饰pgm(磷酸葡萄糖变位酶)和/或galU(UTP-葡萄糖-1-磷酸尿苷酰转移酶)(或其直向同源物或衍生物)以提高酶生产率。或者或此外,可过表达或修饰大肠杆菌ycjU(β-磷酸葡萄糖变位酶)(其将葡萄糖-6-磷酸转化为葡萄糖-1-磷酸)和两歧双歧杆菌ugpA(其将葡萄糖-1-磷酸转化为UDP)或这些酶的直向同源物或衍生物以提高酶生产率。
或者或此外,所述微生物细胞具有一种或多种遗传修饰,所述遗传修饰增加至戊糖磷酸途径(PPP)的通量,如大肠杆菌zwf(或其同源物或工程化衍生物)的过表达或活性增加,其是NADP+依赖性葡萄糖-6-磷酸脱氢酶。
或者或此外,所述微生物细胞具有增加葡萄糖转运的一种或多种遗传修饰。此类修饰包括大肠杆菌galP(半乳糖:H+同向转运体)和大肠杆菌glk(葡萄糖激酶)或可替代地运动发酵单胞菌glf和大肠杆菌glk或这些基因的同源物、直向同源物或工程化的衍生物的表达或活性增加。
或者或此外,所述微生物细胞具有增加UTP产生和再循环的一种或多种遗传修饰。此类修饰包括大肠杆菌pyrH(UMP激酶)、大肠杆菌cmk(胞苷酸激酶)、大肠杆菌adk(腺苷酸激酶)或大肠杆菌ndk(核苷二磷酸激酶)或这些酶的同源物、直向同源物或工程化的衍生物的表达或活性增加。
或者或此外,所述微生物细胞具有增加UDP产生的一种或多种遗传修饰。此类修饰包括大肠杆菌upp(尿嘧啶磷酸核糖基转移酶)、大肠杆菌dctA(C4二羧酸/乳清酸:H+同向转运体)、大肠杆菌pyrE(乳清酸磷酸核糖基转移酶)和大肠杆菌pyrF(乳清苷-5'-磷酸脱羧酶),包括其同源物物、直向同源物或工程化的衍生物中的一者或多者的过表达或活性增加。例如,在一些实施方案中,所述微生物细胞过表达upp、pyrH和cmk或其同源物或工程化的衍生物,或具有增加的upp、pyrH和cmk或其同源物或工程化的衍生物活性。或者,所述微生物细胞过表达dctA、pyre、pyrH和cmk或其同源物或工程化的衍生物,或具有增加的dctA、pyre、pyrH和cmk或其同源物或工程化的衍生物活性。
或者或此外,所述微生物细胞可具有一种或多种遗传修饰以除去或减少对葡萄糖摄取的调控。例如,所述微生物细胞可具有sgrS的缺失、失活或减少的表达,sgrS是大肠杆菌中的小调控RNA。
或者或此外,所述微生物细胞可具有减少葡萄糖-1-磷酸的去磷酸化的一种或多种遗传修饰。示例性修饰包括微生物细胞中大肠杆菌agp(葡萄糖-1-磷酸酶)、大肠杆菌yihX(α-D-葡萄糖-1-磷酸磷酸酶)、大肠杆菌ybiV(糖磷酸酶)、大肠杆菌yidA(糖磷酸酶)、大肠杆菌yigL(磷酸糖磷酸酶)和大肠杆菌phoA(碱性磷酸酶)或其直向同源物中的一者或多者的缺失、失活或表达或活性降低。
或者或此外,所述微生物细胞可具有减少葡萄糖-1-磷酸至TDP-葡萄糖的转化的一种或多种遗传修饰。示例性修饰包括微生物细胞中大肠杆菌rffH(dTDP-葡萄糖焦磷酸化酶)和大肠杆菌rfbA(DTDP葡萄糖焦磷酸)或其直向同源物中的一者或多者的缺失、失活或表达或活性降低。
或者或此外,所述微生物细胞可具有减少葡萄糖-1-磷酸至ADP-葡萄糖的转化的一种或多种遗传修饰。示例性修饰包括微生物细胞中大肠杆菌gigC(葡萄糖-1-磷酸腺苷酰转移酶)或其直向同源物的缺失、失活或表达或活性降低。
在一些实施方案中,所述微生物细胞是包含遗传修饰的细菌细胞:ushA和galETKM缺失、失活或表达降低;pgi缺失、失活或表达降低;并且pgm和galU过表达或互补。
在一些实施方案中,对内源性基因进行编辑以通过改变所编码蛋白质的氨基酸序列而失活或降低酶活性,或通过表达控制序列的编辑而降低表达。编辑可修饰内源启动子、核糖体结合序列或其它表达控制序列,和/或在一些实施方案中,修饰基因调控中的反式作用因子和/或顺式作用因子。基因组编辑可以使用CRISPR/Cas基因组编辑技术,或采用锌指核酸酶和TALEN的类似技术进行。在一些实施方案中,通过同源重组置换内源性基因。
各种实施方案中的微生物细胞不表达用于产生前体(例如,包含一种或多种植物酶)的重组生物合成途径。例如,对于根据本发明的实施方案的产生RebM(和其他高级糖基化产物)的微生物细胞,不表达甜菊醇生物合成途径,如椰油酰基合酶、月桂烯合酶、月桂烯氧化酶和/或月桂烯酸羟化酶,以使得RebM和其他高级糖基化产物的产生取决于将甜菊醇糖苷中间体进料至细胞。
在各种实施方案中,微生物细胞是来自埃希氏菌属、芽孢杆菌属、红杆菌属、发酵单胞菌属或假单胞菌属的细菌。在一些实施方案中,细菌物种选自大肠杆菌、枯草芽孢杆菌、荚膜红细菌、球形红细菌、运动发酵单胞菌或恶臭假单胞菌。在一些实施方案中,细菌细胞是大肠杆菌。
在其他实施方案中,细胞是真菌细胞,如酵母细胞,例如酵母属、裂殖酵母属、毕赤酵母属、法夫酵母属、克鲁维酵母属、假丝酵母属、篮状菌属、酒香酵母属、管囊酵母属、德巴利氏酵母属、耶氏酵母属和工业多倍体酵母菌株。在一个实施方案中,酵母可以是酵母属、毕赤酵母属或耶氏酵母属的物种,包括酿酒酵母、巴斯德毕赤酵母和解脂耶氏酵母。在一些实施方案中,酵母细胞表达一种或多种细菌转运蛋白或其衍生物,其将糖苷中间体输入细胞中以进行进一步糖基化。
在各种实施方案中,所述微生物细胞具有一种或多种内源性转运蛋白的过表达(例如,与亲本微生物菌株相比),或在某些实施方案中,经修饰以表达重组和/或工程化的转运蛋白。在一些实施方案中,所述微生物细胞表达内源性转运蛋白或其衍生物的一个或多个其他拷贝。例如,转运蛋白的表达或活性可进行修饰以增加甜菊醇糖苷中间体(例如,甜菊苷、甜菊双糖苷和RebA等中的一种或多种)转运到细胞中,同时输出产物,如RebM和/或RebD和/或其他高级糖基化甜菊醇糖苷。
可过表达或工程化以改变微生物细胞中的活性或底物特异性的示例性转运蛋白包括大肠杆菌、acrAD、xylE、ascF、bglF、chbA、ptsG/crr、wzxE、rfbX以及其直向同源物或衍生物。这些蛋白质的衍生物和直向同源物通常包含与大肠杆菌转运蛋白具有至少约30%、或至少约40%、或至少约50%同一性、至少约60%同一性、至少约70%同一性、至少约80%同一性、至少约85%同一性、或至少约90%同一性、或至少约95%同一性、或至少96%、97%、98%或99%同一性的氨基酸序列。
acrAD是在Aires JR and Nikaido H.,Aminoglycosides are captured fromboth periplasm and cytoplasm by the AcrD multidrug efflux transporter ofEscherichia coli,J Bacteriol.2005;187(6):1923-9中描述的大肠杆菌多药外排泵。此外,同源物结构描述于Yu,EW等人,
xylE是与葡萄糖转运蛋白具有同源性的大肠杆菌木糖转运蛋白。xylE描述于Sumiya M,等人,
ascF由Hall BG和Xu L,
bglF由Schnetz K,等人,
chba描述于Keyhani NO,等人,
ptsG编码磷酸转移酶转运系统(PTS)的葡萄糖特异性通透酶并且描述于Meins M,等人,
wzxE及其在分子转运中的作用由Rick PD,等人,
rfbx是描述于Hong Y,等人,
其他转运蛋白包括选自细菌或内源性转运蛋白的那些,其将所需的糖苷中间体转运到细胞中,和/或将所需的产物转运出细胞。例如,转运蛋白可来自宿主物种,或另一种细菌或酵母物种,并且可被工程化以调节其对特定糖苷中间体或产物的亲和力。例如,宿主细胞可过表达与选自以下的大肠杆菌转运蛋白至少约30%、或至少约40%、或至少约50%相同的转运蛋白:ampG、araE、araJ、bcr、cynX、emrA、emrB、emrD、emrE、emrK、emrY、entS、exuT、fsr、fucP、galP、garP、glpT、gudP、gudT、hcaT、hsrA、kgtP、lacY、lgoT、lplT、lptA、lptB、lptC、lptD、lptE、lptF、lptG、mdfA、mdtD、mdtG、mdtH、mdtM、mdtL、mhpT、msbA、nanT、narK、narU、nepI、nimT、nupG、proP、setA、setB、setC、shiA、tfaP、tolC、tsgA、uhpT、xapB、xylE、yaaU、yajR、ybjJ、ycaD、ydeA、ydeF、ydfJ、ydhC、ydhP、ydjE、ydjK、ydiM、ydiN、yebQ、ydcO、yegT、yfaV、yfcJ、ygaY、ygcE、ygcS、yhhS、yhjE、yhjX、yidT、yihN、yjhB和ynfM。在一些实施方案中,所述转运蛋白与大肠杆菌转运蛋白至少约60%、至少约70%、至少约80%、至少约90%或至少约95%相同。
在一些实施方案中,微生物细胞表达与来自真核细胞如酵母、真菌或植物细胞的转运蛋白至少50%相同的转运蛋白。在一些实施方案中,转运蛋白是ABC家族转运蛋白,并且其任选地是亚类PDR(多效耐药性)转运蛋白、MDR(多药耐药性)转运蛋白、MFS家族(主要协助转运蛋白超家族)转运蛋白或SWEET(又名PQ-环、唾液或MtN3家庭)家族转运蛋白。在其他实施方案中,转运蛋白是选自以下的家族:AAAP、SulP、LCT、APC、MOP、ZIP、MPT、VIC、CPA2、ThrE、OPT、Trk、BASS、DMT、MC、AEC、Amt、Nramp、TRP-CC、ACR3、NCS1、PiT、ArsAB、IISP、GUP、MIT、Ctr和CDF。
在一些实施方案中,转运蛋白是ABC家族转运蛋白(又称ATP结合盒转运蛋白),其通常包含多个亚基,所述亚基中的一个或两个是跨膜蛋白,并且所述亚基中的一个或两个是膜相关ATP酶。ATP酶亚基利用三磷酸腺苷(ATP)结合和水解的能量来激发各种底物跨膜的易位,以摄取或输出底物。ABC家族转运蛋白可以是任何子类,包括但不限于:ABCA、ABCB、ABCC、ABCD、ABCE、ABCF和ABCG。
在一些实施方案中,转运蛋白是MFS家族转运蛋白(又称主要协助转运蛋白超家族),其是能够响应于化学渗透离子梯度转运小溶质的单多肽辅助载体。通过MFS转运蛋白转运的化合物可包括单糖、寡糖、肌醇、药物、氨基酸、核苷、有机磷酸酯、克雷布斯循环代谢物以及各种有机和无机阴离子和阳离子。作为举例,MFS转运蛋白包括XylE(来自大肠杆菌)、QacA(来自金黄色葡萄球菌)、Bmr(来自枯草芽孢杆菌)、UhpT(来自大肠杆菌)、LacY(来自大肠杆菌)、FucP(来自大肠杆菌)和ExtU(来自大肠杆菌)。
在一些实施方案中,转运蛋白是转运蛋白的SWEET(糖最终将是输出转运蛋白)家族(又称PQ-环、唾液或MtN3家族),其是糖转运蛋白家族并且是TOG超家族的成员。SWEET的真核生物家族成员具有呈3+1+3重复序列排列的7个跨膜片段(TMS)。作为举例,SWEET转运蛋白包括SWEET1、SWEET2、SWEET9、SWEET12、SWEET13和SWEET14。
在一些实施方案中,转运蛋白与来自酿酒酵母的转运蛋白至少50%相同。在一些实施方案中,转运蛋白与酿酒酵母转运蛋白至少约60%、至少约70%、至少约80%、至少约90%或至少约95%相同。示例性酿酒酵母转运蛋白包括AC1、ADP1、ANT1、AQR1、AQY3、ARN1、ARN2、ARR3、ATG22、ATP4、ATP7、ATP19、ATR1、ATX2、AUS1、AVT3、AVT5、AVT6、AVT7、AZR1、CAF16、CCH1、COT1、CRC1、CTR3、DAL4、DNF1、DNF2、DTR1、DUR3、ECM3、ECM27、ENB1、ERS1、FEN2、FLR1、FSF1、FUR4、GAP1、GET3、GEX2、GGC1、GUP1、HOL1、HCT10、HXT3、HXT5、HXT8、HXT9、HXT11、HXT15、KHA1、ITR1、LEU5、LYP1、MCH1、MCH5、MDL2、MME1、MNR2、MPH2、MPH3、MRS2、MRS3、MTM1、MUP3、NFT1、OAC1、ODC2、OPT1、ORT1、PCA1、PDR1、PDR3、PDR5、PDR8、PDR10、PDR11、PDR12、PDR15、PDR18、PDRI、PDRI 1、PET8、PHO89、PIC2、PMA2、PMC1、PMR1、PRM10、PUT4、QDR1、QDR2、QDR3、RCH1、SAL1、SAM3、SBH2、SEO1、SGE1、SIT1、SLY41、SMF1、SNF3、SNQ2、SPF1、SRP101、SSU1、STE6、STL1、SUL1、TAT2、THI7、THI73、TIM8、TIM13、TOK1、TOM7、TOM70、TPN1、TPO1、TPO2、TPO3、TPO4、TRK2、UGA4、VBA3、VBA5、VCX1、VMA1、VMA3、VMA4、VMA6、VMR1、VPS73、YEA6、YHK8、YIA6、YMC1、YMD8、YOR1、YPK9、YVC1、ZRT1、YBR241C、YBR287W、YDR061W、YDR338C、YFR045W、YGL114W、YGR125W、YIL166C、YKL050C、YMR253C、YMR279C、YNL095C、YOL075C、YPR003C和YPR011C。
在一些实施方案中,酿酒酵母转运蛋白选自以下中的一者或多者:ADP1、AQR1、ARN1、ARN2、ATR1、AUS1、AZR1、DAL4、DTR1、ENB1、FLR1、GEX2、HOL1、HXT3、HXT8、HXT11、NFT1、PDR1、PDR3、PDR5、PDR8、PDR10、PDR11、PDR12、PDR15、PDR18、QDR1、QDR2、QDR3、SEO1、SGE1、SIT1、SNQ2、SSU1、STE6、THI7、THI73、TIM8、TPN1、TPO1、TPO2、TPO3、TPO4、YHK8、YMD8、YOR1和YVC1。在一些实施方案中,酿酒酵母转运蛋白选自FLR1、PDR1、PDR3、PDR5、PDR10、PDR15、SNQ2、TPO1和YOR1中的一者或多者。
在一些实施方案中,转运蛋白与XP_013706116.1(来自欧洲油菜)、NP_001288941.1(来自芜菁)、NEC1(来自矮牵牛)和SWEET13(来自乌拉尔图小麦)至少50%、至少60%相同、至少70%相同、至少80%相同或至少90%或95%相同。
在关于RebM的生物合成的各种实施方案中,所述方法产生甜菊苷、甜菊双糖苷和RebA至RebM的至少40%转化或至少50%转化或至少75%转化。在一些实施方案中,RebM与RebD的比例是至少2:1、或至少4:1、或至少6:1、或至少8:1、或至少9:1、或至少10:1、或至少15:1、或至少20:1。
所述方法可通过分批发酵、补料-分批发酵、连续发酵或半连续发酵进行。例如,在一些实施方案中,所述方法通过分批发酵或补料-分批发酵进行,其中孵育时间小于约72小时,或者在一些实施方案中小于约48小时或小于约24小时。
虽然天然UGT酶通常是植物酶(其通常具有在20℃-24℃范围内的最佳温度),或源自植物酶,但在一些实施方案中,本公开使得能够在微生物细胞(例如,细菌细胞,如大肠杆菌)中以高收率产生糖基化产物,其中酶生产率在约24℃或更高的温度下,如约24℃至约37℃、或约27℃至约37℃、或约30℃至约37℃。
在一些实施方案中,生长期或生产期培养基可含有一种或多种洗涤剂,其量足以增强细胞渗透性,而对生长或存活力没有显著影响。示例性洗涤剂包括Tween 20、TritonX-100和SDS等。
在一些实施方案中,所述方法可扩展用于大规模生产。例如,在一些实施方案中,培养的大小是至少约100L、至少约200L、至少约500L、至少约1,000L、或至少约10,000L。
在各个实施方案中,方法还包括从细胞培养物或从细胞裂解物中回收糖基化产物。在一些实施方案中,培养产生至少约100mg/L、或至少约200mg/L、或至少约500mg/L、或至少约1g/L、或至少约2g/L、或至少约5g/L、或至少约10g/L、或至少约20g/L、或至少约30g/L、或至少约40g/L、或至少约50g/L的糖基化产物,其在一些实施方案中是从培养基提取的。
在一些实施方案中,糖基化产物(例如,RebM)是从培养基组分纯化的。因此,在一些实施方案中,所述方法包括从宿主细胞分离生长培养基,并从生长培养基分离所需的糖基化产物(例如,RebM)。在一些实施方案中,从细胞材料进一步提取诸如RebM的产物。
在一些方面,本发明提供用于制备包含糖基化产物(如RebM)的产物的方法。所述方法包括将目标甜菊醇糖苷(根据本公开产生)掺入产品,如食品、饮料、口腔护理产品、甜味剂、调味剂或其他产品中。根据本发明制备的纯化的甜菊醇糖苷可用于多种产品中,包括但不限于食品、饮料、纹理剂(例如,淀粉、纤维、树胶、脂肪和脂肪模拟物和乳化剂)、药物组合物、烟草产品、营养食品组合物、口腔卫生组合物和美容组合物。对其而言可组合使用RebM的调味剂的非限制性实例包括酸橙、柠檬、桔子、水果、香蕉、葡萄、梨、菠萝、芒果、苦杏仁、可乐果、肉桂、糖、棉花糖和香草调味剂。其它食品成分的非限制性实例包括调味剂、酸化剂和氨基酸、着色剂、增量剂、改性淀粉、树胶、调质剂、防腐剂、抗氧化剂、乳化剂、稳定剂、增稠剂和胶凝剂。
在一些方面,本发明提供了用于制备包含多种高甜度甜味剂的甜味剂产品的方法,所述多种高甜度甜味剂如包括甜菊醇糖苷、罗汉果苷、三氯蔗糖、阿斯巴甜、纽甜、爱德万甜、乙酰磺胺酸钾、糖精、环己基氨基磺酸盐、新橙皮苷二氢查尔酮、买麻藤素E和/或白皮杉醇4'-O-β-D-吡喃葡萄糖苷中的两种或更多种。所述方法还可包括将甜味剂产品掺入食品、饮料、口腔护理产品、甜味剂、调味剂或其他产品中,包括上述那些。
一种或多种目标甜菊醇糖苷(如RebM)和包含所述目标甜菊醇糖苷的甜味剂组合物可与各种生理活性物质或功能性成分组合使用。功能成分通常分为以下几类诸如:类胡萝卜素、膳食纤维、脂肪酸、皂苷、抗氧化剂、营养食品、类黄酮、异硫氰酸酯、酚、植物固醇和甾烷醇(植物甾醇和植物甾烷醇);多元醇;益生元、益生菌;植物雌激素;大豆蛋白;硫化物类/硫醇类;氨基酸;蛋白质;维生素;和矿物质。功能成分也可以基于其健康益处进行分类,诸如心血管、降胆固醇和消炎益处。
此外,根据本发明获得的一种或多种目标甜菊醇糖苷如RebM和甜味剂组合物可作为高甜度甜味剂应用以产生口味特征改善的零卡路里、低卡路里或糖尿病饮料和食物产品。它也可用于其中不能使用糖的饮料、食品、药品和其它产品中。此外,RebM和甜味剂组合物不仅可用作饮料、食品和其它专供人消耗的产品中的甜味剂,还可用作特征改善的动物饲料和草料中的甜味剂。
其中可使用目标甜菊醇糖苷和甜味剂组合物的产品的实例包括但不限于酒精饮料,诸如伏特加、葡萄酒、啤酒、烈酒和清酒等;天然果汁;清凉饮料;碳酸软饮料;减肥饮料;零卡路里饮料;低卡路里饮料和食品;酸奶饮料;速溶果汁;速溶咖啡;粉状速溶饮料;罐头产品;糖浆;发酵豆瓣酱;酱油;醋;调味品;蛋黄酱;番茄酱;咖喱;汤;速食肉汤;酱油粉;醋粉;各类饼干;大米饼干;薄脆饼干;面包;巧克力;焦糖;糖果;口香糖;果冻;布丁;果蔬脯;鲜奶油;果酱;柑橘酱;花膏;奶粉;冰淇淋;果汁冰糕;瓶装果蔬;罐装煮豆;用甜酱烹煮的肉类和食品;农业蔬菜食品;海鲜;火腿;香肠;鱼肉火腿;鱼肉香肠;鱼酱;油炸鱼制品;干海鲜产品;冷冻食品;腌制海带;腌肉;烟草;药品;及许多其它产品。
在诸如食品、饮料、药品、化妆品、桌面产品和口香糖等产品的制造过程中,可以使用常规方法诸如混合、揉捏、溶解、酸洗、渗透、渗滤、喷洒、雾化、灌注和其它方法。
本发明的实施方案在以下非限制性实施例中展示。
表2示出三批甜叶菊叶提取物的甜菊醇糖苷含量。RebM途径中的两种中间体RebA和甜菊苷是批次中的两种主要糖苷。
表2:
图4示出糖基化甜菊醇糖苷中间体的生物转化。在实验中,将0.2mM甜菊双糖苷进料至96孔板中的工程化的大肠杆菌菌株。在48小时后检查产物形成。甚至早期中间体如甜菊双糖苷的全细胞生物转化是可能的,但对于天然大肠杆菌而言,UGT酶不可获得糖基化底物(菌株5显示显著活性,而菌株3和4显示可忽略的活性)。菌株的细节在图8中描述。图4中报告的值是相对于对照的化合物浓度(对于空载体对照,每种化合物的浓度除以这些化合物的总浓度)。菌株5中所含的修饰证明甜菊双糖苷至RebM的生物转化。
菌株1-5中的每一者均包含BAC(细菌人工染色体),即单拷贝质粒,其分别表达四种UGT酶:MbUGTC13、MbUGT1.2_2、MbUGTc19_2和76G1_L200A。对照含有相同的BAC主链,但不含UGT酶。
图8示出遗传修饰以增加至UDP-葡萄糖(其是RebM途径中UGT的关键底物)的天然大肠杆菌通量。通过增加可用于UGT的底物的量,大于至UDP-葡萄糖的天然通量可允许更多的RebM形成。产生菌株5的修饰还产生细胞将进料甜菊醇糖苷转化为RebM的能力。对于菌株2和3,消耗UDP葡萄糖的酶(ushA,galETKM)缺失。通过缺失消耗UDP葡萄糖的前体,即葡萄糖-6-磷酸(G6P)的酶(pgi),从菌株3构建了菌株4。菌株5是通过过表达用于经由葡萄糖-1-磷酸(G1P)将G6P转化为UDP-葡萄糖的两种酶(pgm,galU)而从菌株4构建的。
图5示出叶提取物中发现的两种市售中间体,甜菊苷和RebA至RebM的转化。还示出甜菊醇核心的生物转化。所述值报告为总甜菊醇糖苷组合物的百分比。所有菌株将甜菊醇转化为RebM表明,所有菌株都能够形成RebM。与甜菊双糖苷的情况一样,甜菊苷和RebA均被转化为RebM,但仅在菌株5中。很可能只有菌株5使甜菊醇糖苷可用于UGT酶。对于从甜菊苷和RebA转化,RebD:RebM的比例强烈有利于RebM(1:20)。RebM被分泌到培养基中。
图6示出96孔板中甜菊醇糖苷的生物转化。对于这些研究,将76G1_L200A用MbUGT1-3_1.5替代。对于RebM途径中的五种主要化合物,在有或没有表达的UGT酶情况下示出化合物浓度。向菌株进料0.5g/L的甜菊苷/RebA叶提取物,并在取样前与底物一起孵育48小时。如所示,所述细胞几乎只产生RebM,仅含有少量RebI和RebD。
将所述菌株用于初始生物反应器实验。具体而言,将产生RebM的菌株从工作细胞库接种在50mL离心管中,所述离心管含有10mL的LB(Luria-Bertani)肉汤和合适的抗生素。将第一种子在37℃下在振荡孵育箱中培养20小时。在20小时后,然后通过使用第一种子在500mL锥形瓶中接种100mL发酵培养基来开始第二种子培养。在转移至各自含有170mL发酵培养基的生物反应器之前,将这种第二种子培养10小时。每10小时进行采样。使用LC-MS-MS和YSI对培养基中的相关代谢物进行定量。经由分光光度计在600nm的吸光度下测量细胞密度。图7示出在生物反应器规模上甜菊苷/RebA叶提取物的生物转化。对于RebM途径中的五种主要化合物,在整个发酵过程中的不同时间点对培养基中的化合物浓度进行采样。向菌株进料2g/L的甜菊苷/RebA叶提取物,并与底物一起孵育总共30小时。如图7所示,甜菊苷和RebA随时间推移而丢失,而高糖基化产物如RebM、RebI和RebD相应增加。
将途径中间体(如甜菊双糖苷、甜菊苷和RebA)转化为RebM的UGT酶在大肠杆菌细胞质中表达,并且因此需要中间体可能通过转运蛋白或通过增加的膜渗透性的作用而可用于UGT酶。
甜菊醇糖苷是可能不会被天然大肠杆菌菌株摄取的大分子。这将解释通过菌株1进行的甜菊双糖苷至RebM的可忽略的转化。菌株1-4可摄取较早的非糖基化中间体(甜菊醇)并将其转化为RebM,从而表明可忽略转化的原因是缺乏甜菊醇糖苷至细胞质的摄取,而不是在这些条件下途径中间体上UGT酶的失活。
>SrUGT85C2[甜叶菊]
(SEQ ID NO:1)
MDAMATTEKKPHVIFIPFPAQSHIKAMLKLAQLLHHKGLQITFVNTDFIHNQFLESSGPHCLDGAPGFRFETIPDGVSHSPEASIPIRESLLRSIETNFLDRFIDLVTKLPDPPTCIISDGFLSVFTIDAAKKLGIPVMMYWTLAACGFMGFYHIHSLIEKGFAPLKDASYLTNGYLDTVIDWVPGMEGIRLKDFPLDWSTDLNDKVLMFTTEAPQRSHKVSHHIFHTFDELEPSIIKTLSLRYNHIYTIGPLQLLLDQIPEEKKQTGITSLHGYSLVKEEPECFQWLQSKEPNSVVYVNFGSTTVMSLEDMTEFGWGLANSNHYFLWIIRSNLVIGENAVLPPELEEHIKKRGFIASWCSQEKVLKHPSVGGFLTHCGWGSTIESLSAGVPMICWPYSWDQLTNCRYICKEWEVGLEMGTKVKRDEVKRLVQELMGEGGHKMRNKAKDWKEKARIAIAPNGSSSLNIDKMVKEITVLARN
>SrUGT74G1[甜叶菊]
(SEQ ID NO:2)
MAEQQKIKKSPHVLLIPFPLQGHINPFIQFGKRLISKGVKTTLVTTIHTLNSTLNHSNTTTTSIEIQAISDGCDEGGFMSAGESYLETFKQVGSKSLADLIKKLQSEGTTIDAIIYDSMTEWVLDVAIEFGIDGGSFFTQACVVNSLYYHVHKGLISLPLGETVSVPGFPVLQRWETPLILQNHEQIQSPWSQMLFGQFANIDQARWVFTNSFYKLEEEVIEWTRKIWNLKVIGPTLPSMYLDKRLDDDKDNGFNLYKANHHECMNWLDDKPKESVVYVAFGSLVKHGPEQVEEITRALIDSDVNFLWVIKHKEEGKLPENLSEVIKTGKGLIVAWCKQLDVLAHESVGCFVTHCGFNSTLEAISLGVPVVAMPQFSDQTTNAKLLDEILGVGVRVKADENGIVRRGNLASCIKMIMEEERGVIIRKNAVKWKDLAKVAVHEGGSSDNDIVEFVSELIKA
>SrUGT76G1[甜叶菊]
(SEQ ID NO:3)
MENKTETTVRRRRRIILFPVPFQGHINPILQLANVLYSKGFSITIFHTNFNKPKTSNYPHFTFRFILDNDPQDERISNLPTHGPLAGMRIPIINEHGADELRRELELLMLASEEDEEVSCLITDALWYFAQSVADSLNLRRLVLMTSSLFNFHAHVSLPQFDELGYLDPDDKTRLEEQASGFPMLKVKDIKSAYSNWQILKEILGKMIKQTKASSGVIWNSFKELEESELETVIREIPAPSFLIPLPKHLTASSSSLLDHDRTVFQWLDQQPPSSVLYVSFGSTSEVDEKDFLEIARGLVDSKQSFLWVVRPGFVKGSTWVEPLPDGFLGERGRIVKWVPQQEVLAHGAIGAFWTHSGWNSTLESVCEGVPMIFSDFGLDQPLNARYMSDVLKVGVYLENGWERGEIANAIRRVMVDEEGEYIRQNARVLKQKADVSLMKGGSSYESLESLVSYISSL
>SrUGT91D1[甜叶菊]
(SEQ ID NO:4)
MYNVTYHQNSKAMATSDSIVDDRKQLHVATFPWLAFGHILPFLQLSKLIAEKGHKVSFLSTTRNIQRLSSHISPLINVVQLTLPRVQELPEDAEATTDVHPEDIQYLKKAVDGLQPEVTRFLEQHSPDWIIYDFTHYWLPSIAASLGISRAYFCVITPWTIAYLAPSSDAMINDSDGRTTVEDLTTPPKWFPFPTKVCWRKHDLARMEPYEAPGISDGYRMGMVFKGSDCLLFKCYHEFGTQWLPLLETLHQVPVVPVGLLPPEIPGDEKDETWVSIKKWLDGKQKGSVVYVALGSEALVSQTEVVELALGLELSGLPFVWAYRKPKGPAKSDSVELPDGFVERTRDRGLVWTSWAPQLRILSHESVCGFLTHCGSGSIVEGLMFGHPLIMLPIFCDQPLNARLLEDKQVGIEIPRNEEDGCLTKESVARSLRSVVVENEGEIYKANARALSKIYNDTKVEKEYVSQFVDYLEKNARAVAIDHES
>SrUGT91D2[甜叶菊]
(SEQ ID NO:5)
MATSDSIVDDRKQLHVATFPWLAFGHILPYLQLSKLIAEKGHKVSFLSTTRNIQRLSSHISPLINVVQLTLPRVQELPEDAEATTDVHPEDIPYLKKASDGLQPEVTRFLEQHSPDWIIYDYTHYWLPSIAASLGISRAHFSVTTPWAIAYMGPSADAMINGSDGRTTVEDLTTPPKWFPFPTKVCWRKHDLARLVPYKAPGISDGYRMGLVLKGSDCLLSKCYHEFGTQWLPLLETLHQVPVVPVGLLPPEVPGDEKDETWVSIKKWLDGKQKGSVVYVALGSEVLVSQTEVVELALGLELSGLPFVWAYRKPKGPAKSDSVELPDGFVERTRDRGLVWTSWAPQLRILSHESVCGFLTHCGSGSIVEGLMFGHPLIMLPIFGDQPLNARLLEDKQVGIEIPRNEEDGCLTKESVARSLRSVVVEKEGEIYKANARELSKIYNDTKVEKEYVSQFVDYLEKNTRAVAIDHES
>SrUGT91D2e[甜叶菊]
(SEQ ID NO:6)
MATSDSIVDDRKQLHVATFPWLAFGHILPYLQLSKLIAEKGHKVSFLSTTRNIQRLSSHISPLINVVQLTLPRVQELPEDAEATTDVHPEDIPYLKKASDGLQPEVTRFLEQHSPDWIIYDYTHYWLPSIAASLGISRAHFSVTTPWAIAYMGPSADAMINGSDGRTTVEDLTTPPKWFPFPTKVCWRKHDLARLVPYKAPGISDGYRMGLVLKGSDCLLSKCYHEFGTQWLPLLETLHQVPVVPVGLLPPEIPGDEKDETWVSIKKWLDGKQKGSVVYVALGSEVLVSQTEVVELALGLELSGLPFVWAYRKPKGPAKSDSVELPDGFVERTRDRGLVWTSWAPQLRILSHESVCGFLTHCGSGSIVEGLMFGHPLIMLPIFGDQPLNARLLEDKQVGIEIPRNEEDGCLTKESVARSLRSVVVEKEGEIYKANARELSKIYNDTKVEKEYVSQFVDYLEKNARAVAIDHES
>OsUGT1-2[水稻(Oryza sativa)]
(SEQ ID NO:7)
MDSGYSSSYAAAAGMHVVICPWLAFGHLLPCLDLAQRLASRGHRVSFVSTPRNISRLPPVRPALAPLVAFVALPLPRVEGLPDGAESTNDVPHDRPDMVELHRRAFDGLAAPFSEFLGTACADWVIVDVFHHWAAAAALEHKVPCAMMLLGSAHMIASIADRRLERAETESPAAAGQGRPAAAPTFEVARMKLIRTKGSSGMSLAERFSLTLSRSSLVVGRSCVEFEPETVPLLSTLRGKPITFLGLMPPLHEGRREDGEDATVRWLDAQPAKSVVYVALGSEVPLGVEKVHELALGLELAGTRFLWALRKPTGVSDADLLPAGFEERTRGRGVVATRWVPQMSILAHAAVGAFLTHCGWNSTIEGLMFGHPLIMLPIFGDQGPNARLIEAKNAGLQVARNDGDGSFDREGVAAAIRAVAVEEESSKVFQAKAKKLQEIVADMACHERYIDGFIQQLRSYKD
>MbUGTC19[环状变换体]
(SEQ ID NO:8)
MAECMNWLDDKPKESVVYVAFGSLVKHGPEQVEEITRALIDSDVNFLWVIKHKEEGKLPENLSEVIKTGKGLIVAWCKQLDVLAHESVGCFVTHCGFNSTLEAISLGVPVVAMPQFSDQTTNAKLLDEILGVGVRVKADENGIVRRGNLASCIKMIMEEERGVIIRKNAVKWKDLAKVAVHEGGSSDNDIVEFVSELIKAGSGEQQKIKKSPHVLLIPFPLQGHINPFIQFGKRLISKGVKTTLVTTIHTLNSTLNHSNTTTTSIEIQAISDGCDEGGFMSAGESYLETFKQVGSKSLADLIKKLQSEGTTIDAIIYDSMTEWVLDVAIEFGIDGGSFFTQACVVNSLYYHVHKGLISLPLGETVSVPGFPVLQRWETPLILQNHEQIQSPWSQMLFGQFANIDQARWVFTNSFYKLEEEVIEWTRKIWNLKVIGPTLPSMYLDKRLDDDKDNGFNLYKANHH
>MbUGT1,2[环状变换体]
(SEQ ID NO:9)
MAGSSGMSLAERFSLTLSRSSLVVGRSCVEFEPETVPLLSTLRGKPITFLGLMPPLHEGRREDGEDATVRWLDAQPAKSVVYVALGSEVPLGVEKVHELALGLELAGTRFLWALRKPTGVSDADLLPAGFEERTRGRGVVATRWVPQMSILAHAAVGAFLTHCGWNSTIEGLMFGHPLIMLPIFGDQGPNARLIEAKNAGLQVARNDGDGSFDREGVAAAIRAVAVEEESSKVFQAKAKKLQEIVADMACHERYIDGFIQQLRSYKDDSGYSSSYAAAAGMHVVICPWLAFGHLLPCLDLAQRLASRGHRVSFVSTPRNISRLPPVRPALAPLVAFVALPLPRVEGLPDGAESTNDVPHDRPDMVELHRRAFDGLAAPFSEFLGTACADWVIVDVFHHWAAAAALEHKVPCAMMLLGSAHMIASIADRRLERAETESPAAAGQGRPAAAPTFEVARMKLIRTK
>MbUGT1-3[环状变换体]
(SEQ ID NO:10)
MANWQILKEILGKMIKQTKASSGVIWNSFKELEESELETVIREIPAPSFLIPLPKHLTASSSSLLDHDRTVFQWLDQQPPSSVLYVSFGSTSEVDEKDFLEIARGLVDSKQSFLWVVRPGFVKGSTWVEPLPDGFLGERGRIVKWVPQQEVLAHGAIGAFWTHSGWNSTLESVCEGVPMIFSDFGLDQPLNARYMSDVLKVGVYLENGWERGEIANAIRRVMVDEEGEYIRQNARVLKQKADVSLMKGGSSYESLESLVSYISSLENKTETTVRRRRRIILFPVPFQGHINPILQLANVLYSKGFSITIFHTNFNKPKTSNYPHFTFRFILDNDPQDERISNLPTHGPLAGMRIPIINEHGADELRRELELLMLASEEDEEVSCLITDALWYFAQSVADSLNLRRLVLMTSSLFNFHAHVSLPQFDELGYLDPDDKTRLEEQASGFPMLKVKDIKSAYS
>MbUGT1,2.2[环状变换体]
(SEQ ID NO:11)
MATKGSSGMSLAERFWLTLSRSSLVVGRSCVEFEPETVPLLSTLRGKPITFLGLMPPLHEGRREDGEDATVRWLDAQPAKSVVYVALGSEVPLGVEKVHELALGLELAGTRFLWALRKPTGVSDADLLPAGFEERTRGRGVVATRWVPQMSILAHAAVGAFLTHCGWNSTIEGLMFGHPLIMLPIFGDQGPNARLIEAKNAGLQVARNDGDGSFDREGVAAAIRAVAVEEESSKVFQAKAKKLQEIVADMACHERYIDGFIQQLRSYKDDSGYSSSYAAAAGMHVVICPWLAFGHLLPCLDLAQRLASRGHRVSFVSTPRNISRLPPVRPALAPLVAFVALPLPRVEGLPDGAESTNDVPHDRPDMVELHRRAFDGLAAPFSEFLGTACADWVIVDVFHHWAAAAALEHKVPCAMMLLGSAEMIASIADERLEHAETESPAAAGQGRPAAAPTFEVARMKLIR
>MbUGTC19-2[环状变换体]
(SEQ ID NO:12)
MANHHECMNWLDDKPKESVVYVAFGSLVKHGPEQVEEITRALIDSDVNFLWVIKHKEEGKLPENLSEVIKTGKGLIVAWCKQLDVLAHESVGCFVTHCGFNSTLEAISLGVPVVAMPQFSDQTTNAKLLDEILGVGVRVKADENGIVRRGNLASCIKMIMEEERGVIIRKNAVKWKDLAKVAVHEGGSSDNDIVEFVSELIKAGSGEQQKIKKSPHVLLIPFPLQGHINPFIQFGKRLISKGVKTTLVTTIHTLNSTLNHSNTTTTSIEIQAISDGCDEGGFMSAGESYLETFKQVGSKSLADLIKKLQSEGTTIDAIIYDSMTEWVLDVAIEFGIDGGSFFTQACVVNSLYYHVHKGLISLPLGETVSVPGFPVLQRWETPLILQNHEQIQSPWSQMLFGQFANIDQARWVFTNSFYKLEEEVIEWTRKIWNLKVIGPTLPSMYLDKRLDDDKDNGFNLYKA
>MbUGTC13(甜叶菊UGT85C2,P215T)
(SEQ ID NO:13)
MADAMATTEKKPHVIFIPFPAQSHIKAMLKLAQLLHHKGLQITFVNTDFIHNQFLESSGPHCLDGAPGFRFETIPDGVSHSPEASIPIRESLLRSIETNFLDRFIDLVTKLPDPPTCIISDGFLSVFTIDAAKKLGIPVMMYWTLAACGFMGFYHIHSLIEKGFAPLKDASYLTNGYLDTVIDWVPGMEGIRLKDFPLDWSTDLNDKVLMFTTEATQRSHKVSHHIFHTFDELEPSIIKTLSLRYNHIYTIGPLQLLLDQIPEEKKQTGITSLHGYSLVKEEPECFQWLQSKEPNSVVYVNFGSTTVMSLEDMTEFGWGLANSNHYFLWIIRSNLVIGENAVLPPELEEHIKKRGFIASWCSQEKVLKHPSVGGFLTHCGWGSTIESLSAGVPMICWPYSWDQLTNCRYICKEWEVGLEMGTKVKRDEVKRLVQELMGEGGHKMRNKAKDWKEKARIAIAPNGSSSLNIDKMVKEITVLARN
>76G1_L200A[甜叶菊,L200A]
(SEQ ID NO:14)
MAENKTETTVRRRRRIILFPVPFQGHINPILQLANVLYSKGFSITIFHTNFNKPKTSNYPHFTFRFILDNDPQDERISNLPTHGPLAGMRIPIINEHGADELRRELELLMLASEEDEEVSCLITDALWYFAQSVADSLNLRRLVLMTSSLFNFHAHVSLPQFDELGYLDPDDKTRLEEQASGFPMLKVKDIKSAYSNWQIAKEILGKMIKQTKASSGVIWNSFKELEESELETVIREIPAPSFLIPLPKHLTASSSSLLDHDRTVFQWLDQQPPSSVLYVSFGSTSEVDEKDFLEIARGLVDSKQSFLWVVRPGFVKGSTWVEPLPDGFLGERGRIVKWVPQQEVLAHGAIGAFWTHSGWNSTLESVCEGVPMIFSDFGLDQPLNARYMSDVLKVGVYLENGWERGEIANAIRRVMVDEEGEYIRQNARVLKQKADVSLMKGGSSYESLESLVSYISSL
>MbUGT1-3_1[环状变换体]
(SEQ ID NO:15)
MAFLWVVRPGFVKGSTWVEPLPDGFLGERGRIVKWVPQQEVLAHGAIGAFWTHGGWNSTLESVCEGVPMIFSDFGLDQPLNARYMSDVLKVGVYLENGWERGEIANAIRRLMVDEEGEYIRQNARVLKQKADVSLMKGGSSYESLESLVSYISSLGSGGSGGSGRRRRIILFPVPFQGHINPMLQLANVLYSKGFSITIFHTNFNKPKTSNYPHFTFRFILDNDPQDERISNLPTHGPLAGMRIPIINEHGADELRRELELLMLASEEDEEVSCLITDALWYFAQSVADSLNLRRLVLMTSSLFNFHAHVSLPQFDELGYLDPDDKTRLEEQASGFPMLKVKDIKSAYSNWQIAKEILGKMIKQTKASSGVIWNSFKELEESELETVIREIPAPSFLIPLPKHLTASSSSLLDHDRTVFQWLDQQPPSSVLYVSFGSTSEVDEKDFLEIARGLVDSQS
>MbUGT1-3_1.5[环状变换体]
(SEQ ID NO:16)
MAFLWVVRPGFVKGSTWVEPLPDGFLGERGRIVKWVPQQEVLAHGAIGAFWTHGGWNSTLESVCEGVPMIFSDFGLDQPLNARYMSDVLKVGVYLENGWERGEIANAIRRLMVDEEGEYIRQNARVLKQKADVSLMKGGSSYESLESLVSYISSLGSGGSGGSGRRRRIILFPVPFQGHINPMLQLANVLYSKGFSITIFHTNFNKPKTSNYPHFTFRFILDNDPQTTHGPLAGMRIPIINEHGADELRRELELQMLASEEDEEVSCLITDALWYFAQSVADSLNLPRLVLMTSSLFNFHAHVSLPQFDELGYLDPDDKTRLEEQASGFPMLKVKDIKSAYSNWQIAKEILGKMIKQTKASSGVIWNSFKELEESELETVIREIPAPSFLIPLPKHLTASSSSLLDHDRTVFQWLDQQPPSSVLYVSFGSTSEVDEKDFLEIARGLVDSQS
>MbUGT1-3_2[环状变换体]
(SEQ ID NO:17)
MAFLWVVRPGFVKGSTWVEPLPDGFLGERGRIVKWVPQQEVLAHGAIGAFWTHGGWNSTLESVCEGVPMIFSDFGLDQPLNARYMSDVLKVGVYLENGWERGEIANAIRRLMVDEEGEYIRQNARVLKQKADVSLMKGGSSYESLESLVSYISSLGSGGSGRRRRIILFPVPFQGHINPMLQLANVLYSKGFSITIFHTNFNKPKTSNYPHFTFRFILDNDPQDERISNLPTHGPLAGMRIPIINEHGADELRRELELQMLASEEDEEVSCLITDALWYFAQSVADSLNLPRLVLMTSSLFNFHAHVSLPQFDELGYLDPDDKTRLEEQASGFPMLKVKDIKSAYSNWQIAKEILGKMIKQTKASSGVIWNSFKELEESELETVIREIPAPSFLIPLPKHLTASSSSLLEHDRTVFQWLDQQPPSSVLYVSFGSTSEVDEKDFLEIARGLVDSQS
>SgUGT720-269-1(罗汉果)
(SEQ ID NO:18)
MEDRNAMDMSRIKYRPQPLRPASMVQPRVLLFPFPALGHVKPFLSLAELLSDAGIDVVFLSTEYNHRRISNTEALASRFPTLHFETIPDGLPPNESRALADGPLYFSMREGTKPRFRQLIQSLNDGRWPITCIITDIMLSSPIEVAEEFGIPVIAFCPCSARYLSIHFFIPKLVEEGQIPYADDDPIGEIQGVPLFEGLLRRNHLPGSWSDKSADISFSHGLINQTLAAGRASALILNTFDELEAPFLTHLSSIFNKIYTIGPLHALSKSRLGDSSSSASALSGFWKEDRACMSWLDCQPPRSVVFVSFGSTMKMKADELREFWYGLVSSGKPFLCVLRSDVVSGGEAAELIEQMAEEEGAGGKLGMVVEWAAQEKVLSHPAVGGFLTHCGWNSTVESIAAGVPMMCWPILGDQPSNATWIDRVWKIGVERNNREWDRLTVEKMVRALMEGQKRVEIQRSMEKLSKLANEKVVRGINLHPTISLKKDTPTTSEHPRHEFENMRGMNYEMLVGNAIKSPTLTKK
>SgUGT94-289-3(罗汉果)
(SEQ ID NO:19)
MTIFFSVEILVLGIAEFAAIAMDAAQQGDTTTILMLPWLGYGHLSAFLELAKSLSRRNFHIYFCSTSVNLDAIKPKLPSSFSDSIQFVELHLPSSPEFPPHLHTTNGLPPTLMPALHQAFSMAAQHFESILQTLAPHLLIYDSLQPWAPRVASSLKIPAINFNTTGVFVISQGLHPIHYPHSKFPFSEFVLHNHWKAMYSTADGASTERTRKRGEAFLYCLHASCSVILINSFRELEGKYMDYLSVLLNKKVVPVGPLVYEPNQDGEDEGYSSIKNWLDKKEPSSTVFVSFGSEYFPSKEEMEEIAHGLEASEVNFIWVVRFPQGDNTSGIEDALPKGFLERAGERGMVVKGWAPQAKILKHWSTGGFVSHCGWNSVMESMMFGVPIIGVPMHVDQPFNAGLVEEAGVGVEAKRDPDGKIQRDEVAKLIKEVVVEKTREDVRKKAREMSEILRSKGEEKFDEMVAEISLLLKI
>SgUGT74-345-2(罗汉果)
(SEQ ID NO:20)
MDETTVNGGRRASDVVVFAFPRHGHMSPMLQFSKRLVSKGLRVTFLITTSATESLRLNLPPSSSLDLQVISDVPESNDIATLEGYLRSFKATVSKTLADFIDGIGNPPKFIVYDSVMPWVQEVARGRGLDAAPFFTQSSAVNHILNHVYGGSLSIPAPENTAVSLPSMPVLQAEDLPAFPDDPEVVMNFMTSQFSNFQDAKWIFFNTFDQLECKKQSQVVNWMADRWPIKTVGPTIPSAYLDDGRLEDDRAFGLNLLKPEDGKNTRQWQWLDSKDTASVLYISFGSLAILQEEQVKELAYFLKDTNLSFLWVLRDSELQKLPHNFVQETSHRGLVVNWCSQLQVLSHRAVSCFVTHCGWNSTLEALSLGVPMVAIPQWVDQTTNAKFVADVWRVGVRVKKKDERIVTKEELEASIRQVVQGEGRNEFKHNAIKWKKLAKEAVDEGGSSDKNIEEFVKTIA
>SgUGT75-281-2(罗汉果)
(SEQ ID NO:21)
MGDNGDGGEKKELKENVKKGKELGRQAIGEGYINPSLQLARRLISLGVNVTFATTVLAGRRMKNKTHQTATTPGLSFATFSDGFDDETLKPNGDLTHYFSELRRCGSESLTHLITSAANEGRPITFVIYSLLLSWAADIASTYDIPSALFFAQPATVLALYFYYFHGYGDTICSKLQDPSSYIELPGLPLLTSQDMPSFFSPSGPHAFILPPMREQAEFLGRQSQPKVLVNTFDALEADALRAIDKLKMLAIGPLIPSALLGGNDSSDASFCGDLFQVSSEDYIEWLNSKPDSSVVYISVGSICVLSDEQEDELVHALLNSGHTFLWVKRSKENNEGVKQETDEEKLKKLEEQGKMVSWCRQVEVLKHPALGCFLTHCGWNSTIESLVSGLPVVAFPQQIDQATNAKLIEDVWKTGVRVKANTEGIVEREEIRRCLDLVMGSRDGQKEEIERNAKKWKELARQAIGEGGSSDSNLKTFLWEIDLEI
>SgUGT720-269-4(罗汉果)
(SEQ ID NO:22)
MAEQAHDLLHVLLFPFPAEGHIKPFLCLAELLCNAGFHVTFLNTDYNHRRLHNLHLLAARFPSLHFESISDGLPPDQPRDILDPKFFISICQVTKPLFRELLLSYKRISSVQTGRPPITCVITDVIFRFPIDVAEELDIPVFSFCTFSARFMFLYFWIPKLIEDGQLPYPNGNINQKLYGVAPEAEGLLRCKDLPGHWAFADELKDDQLNFVDQTTASSRSSGLILNTFDDLEAPFLGRLSTIFKKIYAVGPIHSLLNSHHCGLWKEDHSCLAWLDSRAAKSVVFVSFGSLVKITSRQLMEFWHGLLNSGKSFLFVLRSDVVEGDDEKQVVKEIYETKAEGKWLVVGWAPQEKVLAHEAVGGFLTHSGWNSILESIAAGVPMISCPKIGDQSSNCTWISKVWKIGLEMEDRYDRVSVETMVRSIMEQEGEKMQKTIAELAKQAKYKVSKDGTSYQNLECLIQDIKKLNQIEGFINNPNFSDLLRV
>SgUGT94-289-2(罗汉果)
(SEQ ID NO:23)
MDAQQGHTTTILMLPWVGYGHLLPFLELAKSLSRRKLFHIYFCSTSVSLDAIKPKLPPSISSDDSIQLVELRLPSSPELPPHLHTTNGLPSHLMPALHQAFVMAAQHFQVILQTLAPHLLIYDILQPWAPQVASSLNIPAINFSTTGASMLSRTLHPTHYPSSKFPISEFVLHNHWRAMYTTADGALTEEGHKIEETLANCLHTSCGVVLVNSFRELETKYIDYLSVLLNKKVVPVGPLVYEPNQEGEDEGYSSIKNWLDKKEPSSTVFVSFGTEYFPSKEEMEEIAYGLELSEVNFIWVLRFPQGDSTSTIEDALPKGFLERAGERAMVVKGWAPQAKILKHWSTGGLVSHCGWNSMMEGMMFGVPIIAVPMHLDQPFNAGLVEEAGVGVEAKRDSDGKIQREEVAKSIKEVVIEKTREDVRKKAREMDTKHGPTYFSRSKVSSFGRLYKINRPTTLTVGRFWSKQIKMKRE
>SgUGT94-289-1(罗汉果)
(SEQ ID NO:24)
MDAQRGHTTTILMFPWLGYGHLSAFLELAKSLSRRNFHIYFCSTSVNLDAIKPKLPSSSSSDSIQLVELCLPSSPDQLPPHLHTTNALPPHLMPTLHQAFSMAAQHFAAILHTLAPHLLIYDSFQPWAPQLASSLNIPAINFNTTGASVLTRMLHATHYPSSKFPISEFVLHDYWKAMYSAAGGAVTKKDHKIGETLANCLHASCSVILINSFRELEEKYMDYLSVLLNKKVVPVGPLVYEPNQDGEDEGYSSIKNWLDKKEPSSTVFVSFGSEYFPSKEEMEEIAHGLEASEVHFIWVVRFPQGDNTSAIEDALPKGFLERVGERGMVVKGWAPQAKILKHWSTGGFVSHCGWNSVMESMMFGVPIIGVPMHLDQPFNAGLAEEAGVGVEAKRDPDGKIQRDEVAKLIKEVVVEKTREDVRKKAREMSEILRSKGEEKMDEMVAAISLFLKI
>XP_022151474.1 7-脱氧番木鳖苷酸(7-deoxyloganetic acid)葡糖基转移酶样[苦瓜]
(SEQ ID NO:25)
MAQPQTQARVLVFPYPTVGHIKPFLSLAELLADGGLDVVFLSTEYNHRRIPNLEALASRFPTLHFDTIPDGLPIDKPRVIIGGELYTSMRDGVKQRLRQVLQSYNDGSSPITCVICDVMLSGPIEAAEELGIPVVTFCPYSARYLCAHFVMPKLIEEGQIPFTDGNLAGEIQGVPLFGGLLRRDHLPGFWFVKSLSDEVWSHAFLNQTLAVGRTSALIINTLDELEAPFLAHLSSTFDKIYPIGPLDALSKSRLGDSSSSSTVLTAFWKEDQACMSWLDSQPPKSVIFVSFGSTMRMTADKLVEFWHGLVNSGTRFLCVLRSDIVEGGGAADLIKQVGETGNGIVVEWAAQEKVLAHRAVGGFLTHCGWNSTMESIAAGVPMMCWQIYGDQMINATWIGKVWKIGIERDDKWDRSTVEKMIKELMEGEKGAEIQRSMEKFSKLANDKVVKGGTSFENLELIVEYLKKLKPSN
>XP_022151546.1 7-脱氧番木鳖苷酸葡糖基转移酶样[苦瓜]
(SEQ ID NO:26)
MAQPRVLLFPFPAMGHVKPFLSLAELLSDAGVEVVFLSTEYNHRRIPDIGALAARFPTLHFETIPDGLPPDQPRVLADGHLYFSMLDGTKPRFRQLIQSLNGNPRPITCIINDVMLSSPIEVAEEFGIPVIAFCPCSARFLSVHFFMPNFIEEAQIPYTDENPMGKIEEATVFEGLLRRKDLPGLWCAKSSNISFSHRFINQTIAAGRASALILNTFDELESPFLNHLSSIFPKIYCIGPLNALSRSRLGKSSSSSSALAGFWKEDQAYMSWLESQPPRSVIFVSFGSTMKMEAWKLAEFWYGLVNSGSPFLFVFRPDCVINSGDAAEVMEGRGRGMVVEWASQEKVLAHPAVGGFLTHCGWNSTVESIVAGVPMMCCPIVADQLSNATWIHKVWKIGIEGDEKWDRSTVEMMIKELMESQKGTEIRTSIEMLSKLANEKVVKGGTSLNNFELLVEDIKTLRRPYT
>XP_022151514.1 7-脱氧番木鳖苷酸葡糖基转移酶样[苦瓜]
(SEQ ID NO:27)
MEQSDSNSDDHQHHVLLFPFPAKGHIKPFLCLAQLLCGAGLQVTFLNTDHNHRRIDDRHRRLLATQFPMLHFKSISDGLPPDHPRDLLDGKLIASMRRVTESLFRQLLLSYNGYGNGTNNVSNSGRRPPISCVITDVIFSFPVEVAEELGIPVFSFATFSARFLFLYFWIPKLIQEGQLPFPDGKTNQELYGVPGAEGIIRCKDLPGSWSVEAVAKNDPMNFVKQTLASSRSSGLILNTFEDLEAPFVTHLSNTFDKIYTIGPIHSLLGTSHCGLWKEDYACLAWLDARPRKSVVFVSFGSLVKTTSRELMELWHGLVSSGKSFLLVLRSDVVEGEDEEQVVKEILESNGEGKWLVVGWAPQEEVLAHEAIGGFLTHSGWNSTMESIAAGVPMVCWPKIGDQPSNCTWVSRVWKVGLEMEERYDRSTVARMARSMMEQEGKEMERRIAELAKRVKYRVGKDGESYRNLESLIRDIKITKSSN
>XP_004147933.2预测的:7-脱氧番木鳖苷酸葡糖基转移酶样[黄瓜]
(SEQ ID NO:28)
MGLSPTDHVLLFPFPAKGHIKPFFCLAHLLCNAGLRVTFLSTEHHHQKLHNLTHLAAQIPSLHFQSISDGLSLDHPRNLLDGQLFKSMPQVTKPLFRQLLLSYKDGTSPITCVITDLILRFPMDVAQELDIPVFCFSTFSARFLFLYFSIPKLLEDGQIPYPEGNSNQVLHGIPGAEGLLRCKDLPGYWSVEAVANYNPMNFVNQTIATSKSHGLILNTFDELEVPFITNLSKIYKKVYTIGPIHSLLKKSVQTQYEFWKEDHSCLAWLDSQPPRSVMFVSFGSIVKLKSSQLKEFWNGLVDSGKAFLLVLRSDALVEETGEEDEKQKELVIKEIMETKEEGRWVIVNWAPQEKVLEHKAIGGFLTHSGWNSTLESVAVGVPMVSWPQIGDQPSNATWLSKVWKIGVEMEDSYDRSTVESKVRSIMEHEDKKMENAIVELAKRVDDRVSKEGTSYQNLQRLIEDIEGFKLN
>XP_022978164.1 7-脱氧番木鳖苷酸葡糖基转移酶样[笋瓜]
(SEQ ID NO:29)
MELSHTHHVLLFPFPAKGHIKPFFSLAQLLCNAGLRVTFLNTDHHHRRIHDLNRLAAQLPTLHFDSVSDGLPPDEPRNVFDGKLYESIRQVTSSLFRELLVSYNNGTSSGRPPITCVITDVMFRFPIDIAEELGIPVFTFSTFSARFLFLIFWIPKLLEDGQLRYPEQELHGVPGAEGLIRWKDLPGFWSVEDVADWDPMNFVNQTLATSRSSGLILNTFDELEAPFLTSLSKIYKKIYSLGPINSLLKNFQSQPQYNLWKEDHSCMAWLDSQPRKSVVFVSFGSVVKLTSRQLMEFWNGLVNSGMPFLLVLRSDVIEAGEEVVREIMERKAEGRWVIVSWAPQEEVLAHDAVGGFLTHSGWNSTLESLAAGVPMISWPQIGDQTSNSTWISKVWRIGLQLEDGFDSSTIETMVRSIMDQTMEKTVAELAERAKNRASKNGTSYRNFQTLIQDITNIIETHI
>XP_022950128.1 7-脱氧番木鳖苷酸葡糖基转移酶样[南瓜]
(SEQ ID NO:30)
MELSPTHHLLLFPFPAKGHIKPFFSLAQLLCNAGARVTFLNTDHHHRRIHDLDRLAAQLPTLHFDSVSDGLPPDESRNVFDGKLYESIRQVTSSLFRELLVSYNNGTSSGRPPITCVITDCMFRFPIDIAEELGIPVFTFSTFSARFLFLFFWIPKLLEDGQLRYPEQELHGVPGAEGLIRCKDLPGFLSDEDVAHWKPINFVNQILATSRSSGLILNTFDELEAPFLTSLSKIYKKIYSLGPINSLLKNFQSQPQYNLWKEDHSCMAWLDSQPPKSVVFVSFGSVVKLTNRQLVEFWNGLVNSGKPFLLVLRSDVIEAGEEVVRENMERKAEGRWMIVSWAPQEEVLAHDAVGGFLTHSGWNSTLESLAAGVPMISWTQIGDQTSNSTWVSKVWRIGLQLEDGFDSFTIETMVRSVMDQTMEKTVAELAERAKNRASKNGTSYRNFQTLIQDITNIIETHI
>XP_020422423.1 7-脱氧番木鳖苷酸葡糖基转移酶[碧桃]
(SEQ ID NO:31)
MAMKQPHVIIFPFPLQGHMKPLLCLAELLCHAGLHVTYVNTHHNHQRLANRQALSTHFPTLHFESISDGLPEDDPRTLNSQLLIALKTSIRPHFRELLKTISLKAESNDTLVPPPSCIMTDGLVTFAFDVAEELGLPILSFNVPCPRYLWTCLCLPKLIENGQLPFQDDDMNVEITGVPGMEGLLHRQDLPGFCRVKQADHPSLQFAINETQTLKRASALILDTVYELDAPCISHMALMFPKIYTLGPLHALLNSQIGDMSRGLASHGSLWKSDLNCMTWLDSQPSKSIIYVSFGTLVHLTRAQVIEFWYGLVNSGHPFLWVMRSDITSGDHQIPAELENGTKERGCIVDWVSQEEVLAHKSVGGFLTHSGWNSTLESIVAGLPMICWPKLGDHYIISSTVCRQWKIGLQLNENCDRSNIESMVQTLMGSKREEIQSSMDAISKLSRDSVAEGGSSHNNLEQLIEYIRNLQHQN
>EOY07351.1 UDP-葡糖基转移酶85A3,推定[可可]
(SEQ ID NO:32)
MRQPHVLVLPFPAQGHIKPMLCLAELLCQAGLRVTFLNTHHSHRRLNNLQDLSTRFPTLHFESVSDGLPEDHPRNLVHFMHLVHSIKNVTKPLLRDLLTSLSLKTDIPPVSCIIADGILSFAIDVAEELQIKVIIFRTISSCCLWSYLCVPKLIQQGELQFSDSDMGQKVSSVPEMKGSLRLHDRPYSFGLKQLEDPNFQFFVSETQAMTRASAVIFNTFDSLEAPVLSQMIPLLPKVYTIGPLHALRKARLGDLSQHSSFNGNLREADHNCITWLDSQPLRSVVYVSFGSHVVLTSEELLEFWHGLVNSGKRFLWVLRPDIIAGEKDHNQIIAREPDLGTKEKGLLVDWAPQEEVLAHPSVGGFLTHCGWNSTLESMVAGVPMLCWPKLPDQLVNSSCVSEVWKIGLDLKDMCDRSTVEKMVRALMEDRREEVMRSVDGISKLARESVSHGGSSSSNLEMLIQELET
>XP_022155979.1β-D-葡糖基西红花酸β-1,6-葡糖基转移酶样[苦瓜]
(SEQ ID NO:33)
MDAHQQAEHTTTILMLPWVGYGHLTAYLELAKALSRRNFHIYYCSTPVNIESIKPKLTIPCSSIQFVELHLPSSDDLPPNLHTTNGLPSHLMPTLHQAFSAAAPLFEEILQTLCPHLLIYDSLQPWAPKIASSLKIPALNFNTSGVSVIAQALHAIHHPDSKFPLSDFILHNYWKSTYTTADGGASEKTRRAREAFLYCLNSSGNAILINTFRELEGEYIDYLSLLLNKKVIPIGPLVYEPNQDEDQDEEYRSIKNWLDKKEPCSTVFVSFGSEYFPSNEEMEEIAPGLEESGANFIWVVRFPKLENRNGIIEEGLLERAGERGMVIKEWAPQARILRHGSIGGFVSHCGWNSVMESIICGVPVIGVPMRVDQPYNAGLVEEAGVGVEAKRDPDGKIQRHEVSKLIKQVVVEKTRDDVRKKVAQMSEILRRKGDEKIDEMVALISLLPKG
>XP_022986080.1β-D-葡糖基西红花酸β-1,6-葡糖基转移酶样[笋瓜]
(SEQ ID NO:34)
MDAQKAVDTPPTTVLMLPWIGYGHLSAYLELAKALSRRNFHVYFCSTPVNLDSIKPNLIPPPSSIQFVDLHLPSSPELPPHLHTTNGLPSHLKPTLHQAFSAAAQHFEAILQTLSPHLLIYDSLQPWAPRIASSLNIPAINFNTTAVSIIAHALHSVHYPDSKFPFSDFVLHDYWKAKYTTADGATSEKIRRGAEAFLYCLNASCDVVLVNSFRELEGEYMDYLSVLLKKKVVSVGPLVYEPSEGEEDEEYWRIKKWLDEKEALSTVLVSFGSEYFPSKEEMEEIAHGLEESEANFIWVVRFPKGEESCRGIEEALPKGFVERAGERAMVVKKWAPQGKILKHGSIGGFVSHCGWNSVLESIRFGVPVIGVPMHLDQPYNAGLLEEAGIGVEAKRDADGKIQRDQVASLIKRVVVEKTREDIWKTVREMREVLRRRDDDMIDEMVAEISVVLKI
>XP_022156002.1β-D-葡糖基西红花酸β-1,6-葡糖基转移酶样[苦瓜]
(SEQ ID NO:35)
MDARQQAEHTTTILMLPWVGYGHLSAYLELAKALSRRNFHIYYCSTPVNIESIKPKLTIPCSSIQFVELHLPFSDDLPPNLHTTNGLPSHLMPALHQAFSAAAPLFEAILQTLCPHLLIYDSLQPWAPQIASSLKIPALNFNTTGVSVIARALHTIHHPDSKFPLSEIVLHNYWKATHATADGANPEKFRRDLEALLCCLHSSCNAILINTFRELEGEYIDYLSLLLNKKVTPIGPLVYEPNQDEEQDEEYRSIKNWLDKKEPYSTIFVSFGSEYFPSNEEMEEIARGLEESGANFIWVVRFHKLENGNGITEEGLLERAGERGMVIQGWAPQARILRHGSIGGFVSHCGWNSVMESIICGVPVIGVPMGLDQPYNAGLVEEAGVGVEAKRDPDGKIQRHEVSKLIKQVVVEKTRDDVRKKVAQMSEILRRKGDEKIDEMVALISLLLKG
>XP_022943327.1β-D-葡糖基西红花酸β-1,6-葡糖基转移酶样[南瓜]
(SEQ ID NO:36)
MDAQKAVDTPPTTVLMLPWIGYGHLSAYLELAKALSRRNFHVYFCSTPVNLDSIKPNLIPPPPSIQFVDLHLPSSPELPPHLHTTNGLPSHLKPTLHQAFSAAAQHFEAILQTLSPHLLIYDSLQPWAPRIASSLNIPAINFNTTAVSIIAHALHSVHYPDSKFPFSDFVLHDYWKAKYTTADGATSEKTRRGVEAFLYCLNASCDVVLVNSFRELEGEYMDYLSVLLKKKVVSVGPLVYEPSEGEEDEEYWRIKKWLDEKEALSTVLVSFGSEYFPPKEEMEEIAHGLEESEANFIWVVRFPKGEESSSRGIEEALPKGFVERAGERAMVVKKWAPQGKILKHGSIGGFVSHCGWNSVLESIRFGVPVIGAPMHLDQPYNAGLLEEAGIGVEAKRDADGKIQRDQVASLIKQVVVEKTREDIWKKVREMREVLRRRDDDDMMIDEMVAVISVVLKI
>XP_022996307.1β-D-葡糖基西红花酸β-1,6-葡糖基转移酶样[笋瓜]
(SEQ ID NO:37)
MSSNLFLKISIPFGRLRDSALNCSVFHCKLHLAIAIAMDAQQAANKSPTATTIFMLPWAGYGHLSAYLELAKALSTRNFHIYFCSTPVSLASIKPRLIPSCSSIQFVELHLPSSDEFPPHLHTTNGLPSRLVPTFHQAFSEAAQTFEAFLQTLRPHLLIYDSLQPWAPRIASSLNIPAINFFTAGAFAVSHVLRAFHYPDSQFPSSDFVLHSRWKIKNTTAESPTQAKLPKIGEAIGYCLNASRGVILTNSFRELEGKYIDYLSVILKKRVFPIGPLVYQPNQDEEDEDYSRIKNWLDRKEASSTVLVSFGSEFFLSKEETEAIAHGLEQSEANFIWGIRFPKGAKKNAIEEALPEGFLERAGGRAMVVEEWVPQGKILKHGSIGGFVSHCGWNSAMESIVCGVPIIGIPMQVDQPFNAGILEEAGVGVEAKRDSDGKIQRDEVAKLIKEVVVERTREDIRNKLEKINEILRSRREEKLDELATEISLLSRN
>XP_022957664.1β-D-葡糖基西红花酸β-1,6-葡糖基转移酶样[南瓜]
(SEQ ID NO:38)
MDAQQAANKSPTASTIFMLPWVGYGHLSAYLELAKALSTRNFHVYFCSTPVSLASIKPRLIPSCSSIQFVELHLPSSDEFPPHLHTTNGLPAHLVPTIHQAFAAAAQTFEAFLQTLRPHLLIYDSLQPWAPRIASSLNIPAINFFTAGAFAVSHVLRAFHYPDSQFPSSDFVLHSRWKIKNTTAESPTQVKIPKIGEAIGYCLNASRGVILTNSFRELEGKYIDYLSVILKKRVLPIGPLVYQPNQDEEDEDYSRIKNWLDRKEASSTVLVSFGSEFFLSKEETEAIAHGLEQSEANFIWGIRFPKGAKKNAIEEALPEGFLERVGGRAMVVEEWVPQGKILKHGNIGGFVSHCGWNSAMESIMCGVPVIGIPMQVDQPFNAGILEEAGVGVEAKRDSDGKIQRDEVAKLIKEVVVERTREDIRNKLEEINEILRTRREEKLDELATEISLLCKN
>OMO57892.1 UDP-葡萄糖苷酸基/UDP-葡糖基转移酶[圆果种黄麻]
(SEQ ID NO:39)
MDSKQKKMSVLMFPWLAYGHISPFLELAKKLSKRNFHTFFFSTPINLNSIKSKLSPKYAQSIQFVELHLPSLPDLPPHYHTTNGLPPHLMNTLKKAFDMSSLQFSKILKTLNPDLLVYDFIQPWAPLLALSNKIPAVHFLCTSAAMSSFSVHAFKKPCEDFPFPNIYVHGNFMNAKFNNMENCSSDDSISDQDRVLQCFERSTKIILVKTFEELEGKFMDYLSVLLNKKIVPTGPLTQDPNEDEGDDDERTKLLLEWLNKKSKSSTVFVSFGSEYFLSKEEREEIAYGLELSKVNFIWVIRFPLGENKTNLEEALPQGFLQRVSERGLVVENWAPQAKILQHSSIGGFVSHCGWSSVMESLKFGVPIIAIPMHLDQPLNARLVVDVGVGLEVIRNHGSLEREEIAKLIKEVVLGNGNDGEIVRRKAREMSNHIKKKGEKDMDELVEELMLICKMKPNSCHLS
>XP_015886141.1预测的:β-D-葡糖基西红花酸β-1,6-葡糖基转移酶样同种型X1[枣]
(SEQ ID NO:40)
MMERQRSIKVLMFPWLAHGHISPFLELAKRLTDRNFQIYFCSTPVNLTSVKPKLSQKYSSSIKLVELHLPSLPDLPPHYHTTNGLALNLIPTLKKAFDMSSSSFSTILSTIKPDLLIYDFLQPWAPQLASCMNIPAVNFLSAGASMVSFVLHSIKYNGDDHDDEFLTTELHLSDSMEAKFAEMTESSPDEHIDRAVTCLERSNSLILIKSFRELEGKYLDYLSLSFAKKVVPIGPLVAQDTNPEDDSMDIINWLDKKEKSSTVFVSFGSEYYLTNEEMEEIAYGLELSKVNFIWVVRFPLGQKMAVEEALPKGFLERVGEKGMVVEDWAPQMKILGHSSIGGFVSHCGWSSLMESLKLGVPIIAMPMQLDQPINAKLVERSGVGLEVKRDKNGRIEREYLAKVIREIVVEKARQDIEKKAREMSNIITEKGEEEIDNVVEELAKLCGM
>XP_002271587.3预测的:β-D-葡糖基西红花酸β-1,6-葡糖基转移酶样[葡萄]
(SEQ ID NO:41)
MDARQSDGISVLMFPWLAHGHISPFLQLAKKLSKRNFSIYFCSTPVNLDPIKGKLSESYSLSIQLVKLHLPSLPELPPQYHTTNGLPPHLMPTLKMAFDMASPNFSNILKTLHPDLLIYDFLQPWAPAAASSLNIPAVQFLSTGATLQSFLAHRHRKPGIEFPFQEIHLPDYEIGRLNRFLEPSAGRISDRDRANQCLERSSRFSLIKTFREIEAKYLDYVSDLTKKKMVTVGPLLQDPEDEDEATDIVEWLNKKCEASAVFVSFGSEYFVSKEEMEEIAHGLELSNVDFIWVVRFPMGEKIRLEDALPPGFLHRLGDRGMVVEGWAPQRKILGHSSIGGFVSHCGWSSVMEGMKFGVPIIAMPMHLDQPINAKLVEAVGVGREVKRDENRKLEREEIAKVIKEVVGEKNGENVRRKARELSETLRKKGDEEIDVVVEELKQLCSY
>XP_018840205.1预测的:β-D-葡糖基西红花酸β-1,6-葡糖基转移酶样[核桃]
(SEQ ID NO:42)
MDTARKRIRVVMLPWLAHGHISPFLELSKKLAKRNFHIYFCSTPVNLSSIKPKLSGKYSRSIQLVELHLPSLPELPPQYHTTKGLPPHLNATLKRAFDMAGPHFSNILKTLSPDLLIYDFLQPWAPAIAASQNIPAINFLSTGAAMTSFVLHAMKKPGDEFPFPEIHLDECMKTRFVDLPEDHSPSDDHNHISDKDRALKCFERSSGFVMMKTFEELEGKYINFLSHLMQKKIVPVGPLVQNPVRGDHEKAKTLEWLDKRKQSSAVFVSFGTEYFLSKEEMEEIAYGLELSNVNFIWVVRFPEGEKVKLEEALPEGFLQRVGEKGMVVEGWAPQAKILMHPSIGGFVSHCGWSSVMESIDFGVPIVAIPMQLDQPVNAKVVEQAGVGVEVKRDRDGKLEREEVATVIREVVMGNIGESVRKKEREMRDNIRKKGEEKMDGVAQELVQLYGNGIKNV
>XP_021652171.1β-D-葡糖基西红花酸β-1,6-葡糖基转移酶样[橡胶树]
(SEQ ID NO:43)
METLQRRKISVLMFPWLAHGHLSPFLELSKKLNKRNFHVYFCSTPVNLDSIKPKLSAEYSFSIQLVELHLPSSPELPLHYHTTNGLPPHLMKNLKNAFDMASSSFFNILKTLKPDLLIYDFIQPWAPALASSLNIPAVNFLCTSMAMSCFGLHLNNQEAKFPFPGIYPRDYMRMKVFGALESSSNDIKDGERAGRCMDQSFHLILAKTFRELEGKYIDYLSVKLMKKIVPVGPLVQDPIFEDDEKIMDHHQVIKWLEKKERLSTVFVSFGTEYFLSTEEMEEIAYGLELSKAHFIWVVRFPTGEKINLEESLPKRYLERVQERGKIVEGWAPQQKILRHSSIGGFVSHCGWSSIMESMKFGVPIIAMPMNLDQPVNSRIVEDAGVGIEVRRNKSGELEREEIAKTIRKVVVEKDGKNVSRKAREMSDTIRKKGEEEIDGVVDELLQLCDVKTNYLQ
>XP_021619073.1β-D-葡糖基西红花酸β-1,6-葡糖基转移酶样[木薯]
(SEQ ID NO:44)
MATAQTRKISVLMFPWLAHGHLSPFLELSKKLANRNFHVYFCSTPVNLDSIKPKLSPEYHFSIQFVELHLPSSPELPSHYHTTNGLPPHLMKTLKKAFDMASSSFFNILKTLNPDLLIYDFLQPWAPALASSLNIPAVNFLCSSMAMSCFGLNLNKNKEIKFLFPEIYPRDYMEMKLFRVFESSSNQIKDGERAGRCIDQSFHVILAKTFRELEGKYIDYVSVKCNKKIVPVGPLVEDTIHEDDEKTMDHHHHHHDEVIKWLEKKERSTTVFVSFGSEYFLSKEEMEEIAHGLELSKVNFIWVVRFPKGEKINLEESLPEGYLERIQERGKIVEGWAPQRKILGHSSIGGFVSHCGWSSIMESMKLGVPIIAMPMNLDQPINSRIVEAAGVGIEVSRNQSGELEREEMAKTIRKVVVEREGVYVRRKAREMSDVLRKKGEEEIDGVVDELVQLCDMKTNYL
>GAV83746.1含UDPGT结构域蛋白[土瓶草]
(SEQ ID NO:45)
MDLKRRSIRVLMLPWLAHGHISPFLELAKKLTNRNFLIYFCSTPINLNSIKPKLSSKYSFSIQLVELHLPSLPELPPHYHTTNGLPLHLMNTLKTAFDMASPSFLNILKTLKPDLLICDHLQPWAPSLASSLNIPAIIFPTNSAIMMAFSLHHAKNPGEEFPFPSININDDMVKSINFLHSASNGLTDMDRVLQCLERSSNTMLLKTFRQLEAKYVDYSSALLKKKIVLAGPLVQVPDNEDEKIEIIKWLDSRGQSSTVFVSFGSEYFLSKEEREDIAHGLELSKVNFIWVVRFPVGEKVKLEEALPNGFAERIGERGLVVEGWAPQAMILSHSSIGGFVSHCGWSSMMESMKFGVPIIAMPMHIDQPLNARLVEDVGVGLEIKRNKDGRFEREELARVIKEVLVYKNGDAVRSKAREMSEHIKKNGDQEIDGVADALVKLCEMKTNSLNQD
>小粒咖啡(Coffea Arabica)UGT_1,6
(SEQ ID NO:46)
MENHATFNVLMLPWLAHGHVSPYLELAKKLTARNFNVYLCSSPATLSSVRSKLTEKFSQSIHLVELHLPKLPELPAEYHTTNGLPPHLMPTLKDAFDMAKPNFCNVLKSLKPDLLIYDLLQPWAPEAASAFNIPAVVFISSSATMTSFGLHFFKNPGTKYPYGNAIFYRDYESVFVENLTRRDRDTYRVINCMERSSKIILIKGFNEIEGKYFDYFSCLTGKKVVPVGPLVQDPVLDDEDCRIMQWLNKKEKGSTVFVSFGSEYFLSKKDMEEIAHGLEVSNVDFIWVVRFPKGENIVIEETLPKGFFERVGERGLVVNGWAPQAKILTHPNVGGFVSHCGWNSVMESMKFGLPIIAMPMHLDQPINARLIEEVGAGVEVLRDSKGKLHRERMAETINKVMKEASGESVRKKARELQEKLELKGDEEIDDVVKELVQLCATKNKRNGLHYY
>环状变换体接头序列
(SEQ ID NO:47)
GSGGSG
>环状变换体接头序列
(SEQ ID NO:48)
GSGGSGGSG。
- 通过全细胞生物转化产生甜菊醇糖苷
- 甜菊醇、甜菊醇糖苷衍生物或其异构体之一用于刺激、修复或调节皮肤和半粘膜的细胞新陈代谢的美容用途