一种甘蓝型油菜早花基因Bnft.a02及其应用
文献发布时间:2024-01-17 01:26:37
技术领域
本发明涉及分子育种技术领域,更具体的说是涉及一种甘蓝型油菜早花基因Bnft.a02及其应用。
背景技术
甘蓝型油菜是重要的植物油和高蛋白饲用饼粕的来源。由于世界人口快速增长导致对食用油和饲料的需求亦在快速增长,这样使油料作物的高产、高品质育种显得更为重要。但传统育种耗时长,不能满足要求。高通量测序技术、分子设计育种技术应运而生,这些现代分子技术可以加速育种进程。但在芸苔属中,由于传统油菜本身的特点,如生育期过长、株型高大、转化效率低等原因,影响了油菜功能基因的挖掘与功能验证,导致油菜分子育种研究进展较慢。
另一方面,在生产实践中,我国西北青藏高原的高寒山区长期种植白菜型油菜,品质达不到双低标准(低芥酸、低硫苷),产量也极低。因此,我们提出用极早熟甘蓝型油菜取代这种白菜型油菜的想法。
在上述理论研究需求和生产实践需求的双重动力驱使下,我们成功首创了快周期甘蓝型油菜(一种生育期仅约60天的极早熟甘蓝型油菜),并为创制方法取得了专利(专利号ZL201910500867.6)。在这个专利中,我们选取亲本显蕾期作为技术指标,依靠这一指标,后续我们又选育出了一批快周期油菜。但从根本上讲,显蕾期是一种田间形态指标,易受环境效应影响,而且其中的分子机理并不清楚,从经验上讲效率也没有分子标记特别是功能基因标记的效率高。因此,有必要进一步研究快周期油菜极早花的分子机理,克隆其开花相关的功能基因并开发功能基因标记,从而更好的为高效选育快周期油菜服务。
至于开花期,采用不同的材料,不同研究人员定位、克隆出的开花基因不尽相同,其中主效基因包括BnFLC.A2(杨光圣等专利CN106834303A,2017;)、BnFT(王晶博士学位论文,2009;代书桃博士学位论文,2015)等。从这些研究中发现,同一基因,在不同材料或同一材料的不同染色体上的序列、功能都可能不同。例如,王晶等利用宁油7号和Tapidor为研究材料,克隆了BnFT基因分布在A2、A7和C6上的6个拷贝,其中BnFT.A2是新发现的促进春性背景宁油7号开花的FT等位基因。2021年,余华胜等也用极早花材料“迎春1号”锁定1个BnFLC基因及其上游10个调控基因。
综上,如何提供一种油菜开花相关的功能基因并开发功能基因标记是本领域技术人员亟需解决的问题。
发明内容
有鉴于此,本发明提供了一种甘蓝型油菜早花基因Bnft.a02及其应用。
本发明在前期发明专利(专利号ZL201910500867.6)的基础上,用快周期油菜亲本和中双11杂交产生的F2群体,采用BSA法定位了快周期甘蓝型油菜的开花基因,发现主效QTL区间锁定的Bnft.a02基因是引起快周期甘蓝型油菜极早开花的主要原因。对双亲中该基因的比较测序发现,与中双11相比,快周期油菜Bnft.a02内含子区有3个明显改变:其中第一个内含子包括15bp突变和12bp缺失,第二个内含子有288bp缺失;而外显子区则有1bp突变,导致异亮氨酸突变为亮氨酸。表达分析证实,在快周期油菜亲本中该等位基因确实较早强效表达,根据缺失位点设计的两对标记,在F2群体子代能很好的鉴定出早花表型。对两个内含子的突变位点进行了分析,推断很可能是FLC基因调控结合位点motif的改变导致FLC基因不再能抑制FT基因的表达,从而导致极早花表型。这些试验还证明,与以前王晶、代书桃等人克隆的BnFT.A2基因相比,该Bnft.a02不仅序列存在较显著的变异,而且在功能上确实是能引起更早花表型,是新的等位基因。
为了实现上述目的,本发明采用如下技术方案:
一种甘蓝型油菜早花基因Bnft.a02,其核苷酸序列如SEQ ID NO.1所示(下划线部分为外显子序列)。
甘蓝型油菜早花基因Bnft.a02的CDS序列如SEQ ID NO.13所示。
ATGTCTTTAAGTAATAGAGATCCTCTTGTGGTAGGGAGAGTTGTAGGAGACGTTCTTGAATGTTTCACAAGATCAATCGATCTAAGGGTTACTTATGGCCAAAGAGAGGTGACAAATGGGTTGGATCTAAGGCCTTCTCAAGTTCTCAACAAGCCAAGAGTTGAGATTGGTGGAGAAGACCTAAGGAACTTCTATACTTTGGTTATGGTGGATCCAGATGTTCCAAGTCCTAGCAATCCTCACCTCCGAGAATATCTTCACTGGTTGGTGACTGATATCCCAGCGACAACTGGAACAAACTTTGGCAATGAGATTGTGTCTTACGAGAGTCCAAGGCCCAACTCGGGTATTCATCGTATCGTGCTCGTATTGTTCCGACAGCTCGGTAGGCAAACAGTGTATGAACCAGGATGGCGCCAACAATTCAACACTCGTGAGTTTGCTTCCCTATACAATCTCGGCCTTCCCGTGGCTGCGGTTTTCTACAATTGTCAGAGGGAGAGTGGCTGCGGAGGACGAAGAAGTTAG,SEQ ID NO.13。
根据本领域的常规技术手段,可以采用已克隆的多核苷酸序列作为探针,从基因组或cDNA文库中筛选得到本发明的基因或同源基因。同样,采用PCR技术也可以从基因组和cDNA中扩增得到本发明的基因或任何一段感兴趣的核苷酸序列或与其同源的核苷酸序列。因此,在严格条件下,与SEQ ID NO.1所示的核苷酸序列杂交且编码相同功能蛋白质的核苷酸序列也属于快周期油菜早花基因序列。进一步的,上述任一一种核苷酸序列经核苷酸添加、删除、替换、修饰的保守性突变而获得的保守性变异体也属于甘蓝型油菜早花基因序列。
发明还提供了一种表达甘蓝型油菜早花基因Bnft.a02的蛋白,其氨基酸序列由上述任一一种核苷酸序列编码。
上述的甘蓝型油菜早花基因Bnft.a02编码的蛋白,其氨基酸序列如SEQ ID NO.2所示。
MSLSNRDPLVVGRVVGDVLECFTRSIDLRVTYGQREVTNGLDLRPSQVLNKPRVEIGGEDLRNFYTLVMVDPDVPSPSNPHLREYLHWLVTDIPATTGTNFGNEIVSYESPRPNSGIHRIVLVLFRQLGRQTVYEPGWRQQFNTREFASLYNLGLPVAAVFYNCQRESGCGGRRS,SEQ ID NO.2。
一种用于检测甘蓝型油菜开花早晚的引物组,所述引物组包括FT-F-ef、FT682-R、FT1883-F、FT2681-R,其核苷酸序列如下:
FT-F-ef:ATTTGTTTGTCTCTTATCTAAATCC,SEQ ID NO.3;
FT682-R:GCATCGAAGAATTAAAAAGAAACAAAG,SEQ ID NO.4;
FT1883-F:GAAGAGAACTATCTCTGCACAG,SEQ ID NO.5;
FT2681-R:TACGAGCACGATACGATG,SEQ ID NO.6。
一种用于检测甘蓝型油菜开花早晚的试剂盒,包括上述的引物组。
一种检测甘蓝型油菜开花早晚的方法,使用上述引物组进行PCR扩增。
进一步的,当用引物FT-F-ef和FT682-R扩增时出现150bp产物条带,且用FT1883-F和FT2681-R扩增时出现509bp产物条带,则开花早;
当用引物FT-F-ef和FT682-R扩增时无条带,且用FT1883-F和FT2681-R扩增时出现797bp产物条带,则开花晚。
上述早花基因Bnft.a02在甘蓝型油菜育种中的应用。
上述早花基因Bnft.a02在甘蓝型油菜生育期改良中的应用。
经由上述的技术方案可知,与现有技术相比,本发明取得的有益效果为:本发明用快周期油菜亲本和中双11杂交产生的F2群体,采用BSA法定位了快周期甘蓝型油菜的开花基因。本发明设计出了两对功能基因引物,在选育快周期材料时具有更准确、更高效、更经济的特点。
附图说明
为了更清楚地说明本发明实施例或现有技术中的技术方案,下面将对实施例或现有技术描述中所需要使用的附图作简单地介绍,显而易见地,下面描述中的附图仅仅是本发明的实施例,对于本领域普通技术人员来讲,在不付出创造性劳动的前提下,还可以根据提供的附图获得其他的附图。
图1为本发明鉴定、分离甘蓝型油菜早花基因Bnft.a02及推测分子机理的技术流程图;
图2为本发明BSA法定位开花期结果,其中:
a用于比较两个样品(极早池和极晚池)相同位置基因型差异的情况,对于基因型相同的位置不参与比较,样品1(极早池)基因型纯合则记为AA,此时样品2(极晚池)基因型也为纯合时记为BB,否则为AB;样品1基因型杂合则记为AB,此时样品2基因型(一定为纯合)记为AA;记录两个样品的遗传相似度(1-差异位点数/总位点数)×100%。
b为分型分析,本发明比较所有子代在这些位点的基因型,并使用图形化界面展示;蓝色区域表示的是杂合基因型,红色区域表示的是这些片段来自中双11(ZS11)的材料;红框表示目标位点位于A2染色体上。
c为sef1中克隆出的Bnft.a02等位基因与中双11中BnFT.A02的比较示意图。
图3为本发明利用PCR验证sef1与中双11杂交产生的F2群体子代单株标记与开花期的关系,其中:
a为PCR验证17个F2单株的15bp突变,白色箭头表示不含15bp突变的单株,所用引物组合为FT-F-ef/FT682-R;
b为PCR验证17个F2单株的288bp缺失,白色箭头表示不含该片段缺失,所用引物组合为FT1883-F/FT2681-R;
c为显蕾期与花期17个F2单株,箭头表示显蕾期超过60天的单株。
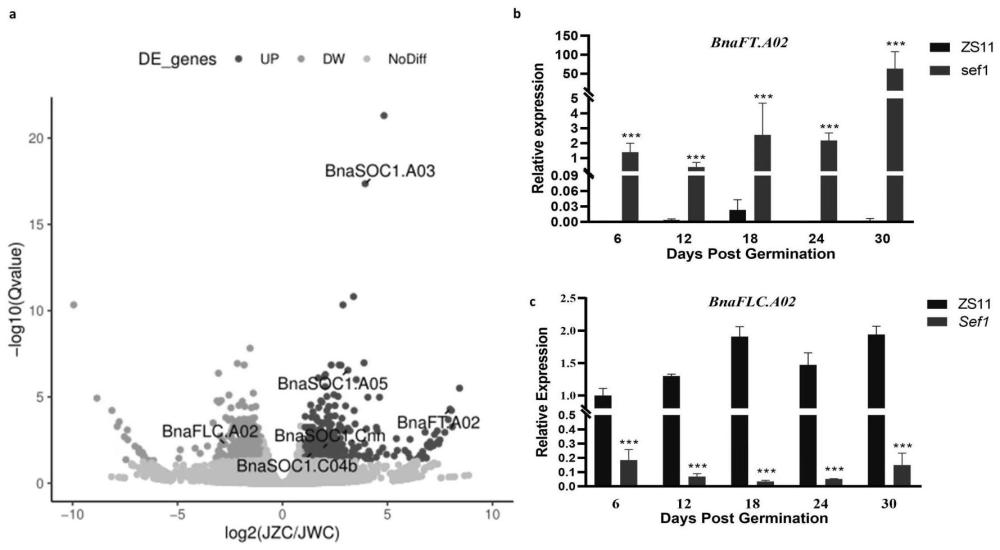
图4为利用转录组学进一步验证中双11(ZS11)和sef1中基因表达差异,其中:
a为总的基因表达差异情况;
b为发芽后30天内基因BnaFT.A02表达差异情况;
c为发芽后30天内基因BnaFLC.A02表达差异情况;
图5为本发明在JASPAR软件中输入BnaFT.A02的变异区域,选择相似性>70%,检测AtFLC的结合motif,显示扫描后的结果,其中:
上图为BnaFT.A02内含子1的15bp突变;
下图为BnaFT.A02内含子2的288bp缺失;
红标为motif可能结合位点,灰标为Bnft.a02突变区域。
具体实施方式
下面将结合本发明实施例中的附图,对本发明实施例中的技术方案进行清楚、完整地描述,显然,所描述的实施例仅仅是本发明一部分实施例,而不是全部的实施例。基于本发明中的实施例,本领域普通技术人员在没有做出创造性劳动前提下所获得的所有其他实施例,都属于本发明保护的范围。
本发明所需药剂为常规实验药剂,采购自市售渠道;未提及的实验方法为常规实验方法,在此不再一一赘述。
实施例1
构建分离群体并定位开花基因
1、实验材料
2016年早花亲本YF211和极晚花DH系SGDH284(KeL,LeiW,YangW,WangJ,GaoJ,ChengJ,SunY,FanZ,YuD(2020)Genome-wideidentification ofcoldresponsivetranscriptionfactorsinBrassicanapusL.BMCplantbiology20:62)杂交产生F2群体,该群体花期分离明显,选取其中最早的一株移入室内自交,后代定向选择最早株,最终选育出快周期油菜(详见专利ZL201910500867.6)。本发明正式将其定名为sef1,由于已将此材料用于太空极早花育种、突变体库构建等研究,因此中文名简称“太极”(一方面为“太空极早花育种”的简称,另一方面,由此材料构建突变体库,能产生更多新的资源,故而取中国道家文化中“太极生两仪,两仪生四象,四象生八卦”之意)。
中双11是生产中大面积种植的高油常规种,也是中国甘蓝型油菜基因组测序品种。
将sef1与中双11杂交,获得100个单株组成的F2分离群体,F2单株及双亲用于基因定位。
本发明中无特殊说明时,植物生长条件均为:Percival植物生长箱,22℃,16小时光/8小时暗,光强450μmolm-2s-1。
2、BSA法定位开花期
播种后第28天时,取双亲、F2各单株幼叶,置液氮中备用。记录各单株显蕾期和开花期。按显蕾期和开花期,将最早的15株的DNA混合成极早池(JZC),同理将最晚的15株混合成极晚池(JWC)。同时,各包含5个单株混样的3个早花文库与3个晚花文库提取RNA用于转录组测序。
sef1、中双11、极早池和极晚池DNA送样给武汉双绿源创芯科技研究院有限公司,采用其开发的甘蓝型油菜50K芯片分析,芯片信息参考Xiao Q,Wang H,Song N,Yu Z,ImranK,Xie W,Qiu S,Zhou F,Wen J,Dai C,Ma C,Tu J,Shen J,Fu T,Yi B(2021)TheBnapus50K array:a quick and versatile genotyping tool for Brassica napusgenomic breeding and research.G311。RNA测序由北京诺禾致源科技股份有限公司采用illumina NovaSeq平台完成,数据分析参考Ke L,Lei W,Yang W,Wang J,Gao J,Cheng J,Sun Y,Fan Z,Yu D(2020)Genome-wide identification of cold responsivetranscription factors in Brassica napus L.BMC plant biology 20:62和Sun Y,Zhang D,Zheng H,Wu Y,Mei J,Ke L,Yu D,Sun Y(2022)Biochemical and ExpressionAnalyses Revealed the Involvement of Proanthocyanidins and/or TheirDerivatives in Fiber Pigmentation of Gossypium stocksii.International journalof molecular sciences 23。
50K芯片结果分析显示,JZC和JWC之间位于A02染色体上6.18-7.06Mb区间存在显著差异(图2a,b),对比中双11测序信息,开花基因BnaFT.A02(Bna02g12130D)正好位于此区间。进一步分析转录组测序数据,P<0.05时,JZC和JWC间存在784个差异基因。其中sef1显蕾前BnaFT.A02上调表达,BnaFLC.A02则下调表达;中双11显蕾前则与此相反(图4)。具体的,sef1中开花基因BnaFT.A02在发芽6天就有显著水平的表达,而中双11则在30天时表达仍不显著(图4b);相应的,开花抑制基因BnaFLC.A02在中双11中30天内能显著表达,而sef1中表达不显著(图4c)。但由于BnaFLC.A02不在定位区间,因此排除BnaFLC.A02,仅将BnaFT.A02作为候选基因。
后续将野生型(中双11)中BnaFT.A02命名为BnFT.A02,sef1中BnaFT.A02命名为Bnft.a02。
3.候选基因验证与全基因克隆
在锁定BnaFT.A02为目标候选基因后,我们克隆并测序sef1中Bnft.a02从起始密码子至终止密码子的全长序列(SEQ ID NO.1)。与中双11中该位点相比,内含子区存在明显的三处变化:第一个内含子包含一处15bp突变和一处12bp缺失,第二个内含子则包含一处长达288bp的缺失。此外,其外显子还包含1bp突变,导致由异亮氨酸突变为亮氨酸(图2c)。
ATGTCTTTAAGTAATAGAGATCCTCTTGTGGTAGGGAGAGTTGTAGGAGACGTTCTTGAATGTTTCACAAGATCAATCGATCTAAGGGTTACTTATGGCCAAAGAGAGGTGACAAATGGGTTGGATCTAAGGCCTTCTCAAGTTCTCAACAAGCCAAGAGTTGAGATTGGTGGAGAAGACCTAAGGAACTTCTATACTTTGGTTCTTACATTTAAACTCTCTTTTGTCTCATCTCTTCTTCATTTCTTCATTATTCGTCTTCTACTATAACTTGGCCATGCAGACCCATAAGGAGGCCTTAAGATAATATTTCCTTTGTTCTATTTTCTTACGATTTATTGGTTTAACTTCTAGATACCCATGTTAAGAAGTATTTCTTTTTCACTTTTTACCATAAAGTATTATACCCATGAATCATTTCATCAAATACCAAACTAAATGTAAACAAGTTTTTGTTTTTATTAGAACAAACAATTTTATATTCTTTCGCAATTATTATTATTTGTTTGTCTCTTATCTAAATCCCAGATGACTTTTTGCAAACTCATTCTTTTATTTTGGGTAAGAATCTTGGTTTTTTCTTTGTTCCTCCAAAAAATTAGATTGTATCTTTGTTTCTTTTTAATTCTTCGATGCTTTTAAAAAGGCAAAAAAATAATATCTGGGTTCGAATATGCAAAATAATAATAATAAAGAAACTCAATGGTCAtTCACACAAATATATAAtGTTCATTTTATTATCTCTATTGTATGACTGTTTGAATCGGTATATTCAAGTCAGTCTTGTAAATACTCTGCTATACAAGACATATAGCTAGGACCTACCTTTTTCTATTTATCTTTCTTCTTCTTCCTTTTTTGTGTTATCTCGTTTCTCAAACTTCAAAAAGTAACTTCTGCTTTCTTGATAAGTTTGAGAATGATAATCTTATATTTTCATTCTTCTTCGTCTTATTTGTAACGGATTTTAGTGTTCATGGGTTTAATGTAGGTTATGGTGGATCCAGATGTTCCAAGTCCTAGCAATCCTCACCTCCGAGAATATCTTCACTGGTTTGTGCACTAATTCACCTCTTTTATTCAAGTTTTTTTTATCCACAAACATGTTTGCTTTAGTTAGATATAAATATATACACTGAACCACACATGCAACATGATATGTATATCAGTTGGAGACTTTGAAATTTTGTAAATCAGTTATAACAAAAGTCAAACATCTTCCGTTTAGTAGACCAGAACATGTACAATCAGGTTTAAGAAAGCGTTTTATCACTTTTTCTTCTAACAAGTCCCATACACCAAGGATAAACTCAAGGGATTTCAAAAGCTTTAGATGTGCATATAAGATATGTATAAATGTTATGAAAACCCTTTTTTTTAAGATATTACATATATATATATATATATATTATTTTTTTTCTTAACAGAGCTACACAATGATTCTATTAAGTATATAGGACGTGACTTATACTAAGATTTGGTTTTCCACTTTAAAGTTGTTTGTATGAACATTCTTAGCCTAATGACATATCTTTATGACTCGAAGTGAGTAGGACTAGGTCTTCTGTTCAATCACAGCTACTACACAACACATCTCGTTTTTAGTCTCAGCTTATATATATATATATATATGCTTTAATACATAGTCTCACTTTTATATATGCATGCACAATATAAAACTTTCAAAGACTCGATTTAAATGTTGATGCATGTTTTGGAGTCACTTAGAGCAGCTCCATTAGTAGTATCAAAACAAGCATCTCATAAATACCAAGGATTACAAAAACAAAAAGTTGGAGAGATAAAGAAGAGAACTATCTCTGCACAGATGTTTTTAGAAAAGGATCAAGCCAAATGTCCATGTTTGAATGGTTCAAATTAAATCTCCAAATTATAAATTAAGTTTTCAACAAAACTACAATATAATATTAATATAAAACCTGACAGATACTTATTTGGAGATATTGTCCAATAATGTTGCTCTTACACCGGCAATCTATTAGTTTAAAATTACAATAAAACCAAGGATCTCATTAGCACCAATAGTCTATTGGTTACGTTATTATCATTTAAAAAAAAACTTTATGTTTATGTGCAATATCTTAGAAAATTATATGGGTACACTTATTACAGGTTGGTGACTGATATCCCAGCGACAACTGGAACAAACTTTGGTGAGTTTTATTCTATAGATTGATCGCTAGACAGTCGATAGAACGAATTCACATGTGTTTGATGTTGTATTTACAGGCAATGAGATTGTGTCTTACGAGAGTCCAAGGCCCAACTCGGGTATTCATCGTATCGTGCTCGTATTGTTCCGACAGCTCGGTAGGCAAACAGTGTATGAACCAGGATGGCGCCAACAATTCAACACTCGTGAGTTTGCTTCCCTATACAATCTCGGCCTTCCCGTGGCTGCGGTTTTCTACAATTGTCAGAGGGAGAGTGGCTGCGGAGGACGAAGAAGTTAG,SEQ ID NO.1。
为验证内含子变异是否关联开花表型,我们根据sef1两个内含子区变异设计了两对引物(表1),在sef1、中双11、JZC和JWC中扩增PCR。引物组合FT-F-ef/FT682-R在sef1和JZC中检测到150bp产物条带,但中双11和JWC无条带。而用FT1883-F/FT2681-R引物组合扩增时,中双11扩增出797bp产物条带,sef1扩增出509bp产物条带,两者刚好相差288bp。为进一步验证,将这两组引物用于100个单株的F2群体扩增,检验标记与花时的连锁关系。结果,在出苗后26天前所有显蕾了的35个单株中,引物组合FT-F-ef/FT682-R都扩增出了150bp条带,而出苗后60天仍未显蕾的22个单株未扩出产物;引物组合FT1883-F/FT2681-R扩增同样单株时,60天后仍未显蕾的单株扩增出797bp产物条带,26天前显蕾的单株扩增出509bp产物条带(图3)。结果还发现,15bp突变和288bp缺失在sef1中同时发生,F2子代中纯合与杂合突变位点均表现早花,而除了7株显蕾期在26~60天间外,其余大部分无288bp缺失的子代显蕾期晚于60天(表2)。综合这些结果,可以肯定BnaFT.A02的变异是导致极早花表型的首要因素。
表1本发明中实验所用引物
表2大群体验证Bnft.a02功能基因标记与显蕾期、开花期的关系
注:mutation表示有突变片段,nomutation表示无突变片段;nodeletion表示无288bp缺失,deletion表示288bp缺失,heterozygote单株表示该位点杂合状态。
4.BnaFT.A02变异在快周期油菜(sef1)中导致极早花表型的分子机理分析
在双子叶越冬植物中,春化前FLC编码1个MADS-box结构的蛋白,直接抑制成花素基因FT及FD表达,从而防止植物在过冬前或越冬时开花。但在快周期油菜sef1中,在22℃恒温培养条件下,BnaFLC.A02一直处于低丰度表达,反而是BnaFT.A02处于较高表达水平(图4)。这说明在sef1中,这一传统现象失效。根据近来研究,FT蛋白可能反馈抑制FLC基因表达从而控制开花(ChenandPenfield,2018;Luoetal.2019)。因此,我们可以推断在快周期油菜中BnaFT.A02的变异提高了FT基因表达水平,抑制了FLC基因表达,从而导致了sef1的极早花表型。同时,由于BnaFT.A02内含子的变异,导致FLC转录因子MADs-box不能有交结合到BnaFT.A02从而不能抑制BnaFT.A02表达。为验证这一设想,我们将BnaFT.A02序列提交到转录因子分析工具JASPAR(
本说明书中各个实施例采用递进的方式描述,每个实施例重点说明的都是与其他实施例的不同之处,各个实施例之间相同相似部分互相参见即可。
对所公开的实施例的上述说明,使本领域专业技术人员能够实现或使用本发明。对这些实施例的多种修改对本领域的专业技术人员来说将是显而易见的,本文中所定义的一般原理可以在不脱离本发明的精神或范围的情况下,在其它实施例中实现。因此,本发明将不会被限制于本文所示的这些实施例,而是要符合与本文所公开的原理和新颖特点相一致的最宽的范围。
- 一种甘蓝型油菜昆虫抗性和抗病性BnGPAT基因及其应用
- 一种基于高分辨率溶解曲线鉴定梨树早花的SNP标记及其应用
- 一种甘蓝型油菜高芥酸基因的分子标记及选育方法
- 一种利用玉米ZmDPS10-2基因选育早花玉米的方法及其应用
- 一种利用玉米ZmDPS10‑2基因选育早花玉米的方法及其应用