调控番茄花序发育的基因及其编码蛋白与应用
文献发布时间:2024-04-18 19:58:53
技术领域
本发明属于植物生物技术领域,具体涉及调控番茄花序发育的基因及其编码蛋白与应用。
背景技术
番茄(Lycopersicon esculentum)是茄科番茄属草本植物,是一种重要的世界性蔬菜,我国是世界第一大番茄生产国,番茄产量约占世界总产量的31%。番茄产量主要由果实大小和果实数目决定,其中花序分枝数目决定了果实数目,与番茄产量直接相关。改良番茄花序形态是提高番茄产量,提升商品价值的重要环节,也是育种家进行种质创新的重点目标。然而,目前关于调控番茄花序分枝的形成机制及调控网络的研究十分有限,对于番茄花序调控基因的利用还存在局限。对番茄花序分枝调控基因的克隆及应用有助于揭示番茄花序分枝发育的遗传基础,为番茄花序形态的精准改良提供新靶点。
番茄花序发育是一个复杂而有序的过程,花序分枝数量主要是由分生组织命运决定、分生组织大小和分生组织成熟速度共同决定(Park et al.,2014;Wang et al.,2018)。开花与否是花序发育的前提,因此开花时间调控基因必然也对番茄花序形态产生影响(Quinet et al.,2006)。番茄中SINGLE FLOWER TRUSS(SFT)是开花促进因子,该基因突变不仅可以延迟番茄开花,并且导致花序发育成单花(Lifschitz et al.,2014;Cui et al.,2020)。番茄TERMINATING FLOWER(TMF)是一个开花抑制因子,该基因突变后,不仅导致番茄提前开花而且使第一花序发育为单花(MacAlister et al.,2012)。顶端分生组织的大小在一定程度上影响了番茄花序分枝数量。WUSCHEL-CLAVATA3(WUS-CLV3)反馈调控通路在分生组织干细胞维持中起到至关重要的作用(Brand et al.,2000;Kondo et al.,2006;Ogawaet al.,2008;Fletcher,2018)。番茄中fasciated(FAS)、fasciated inflorescence(FIN)、fasciated and branched(FAB)和FAB2等基因都是通过调控WUS-CLV3回路,抑制番茄花序分化。这些基因的突变能导致番茄花序分枝数目增加(Xu et al.,2015)。番茄花序形成过程中花序分生组织(inflorescence meristem,IM)的成熟速度是决定番茄花序分枝数量的最重要的因素。番茄中S基因突变会推迟IM的成熟,导致分生组织同时出现多个IM,从而导致花序分枝数量急剧增加(Lippman et al.,2008)。而AN基因促进花分生组织(floralmeristem,FM)的命运决定,因此AN突变使FM的成熟推迟,导致分生组织一直维持在IM状态,从而产生大量花序分枝(Allen and Sussex,1996;Lippman et al.,2008)。一些花器官决定基因也影响番茄的花序分枝数(Mao et al.,2000;Szymkowiak and Irish,2006;Thouetet al.,2012;Nakano et al.,2015)。
番茄花序发育受多种基因的协同调控(Prusinkiewicz et al.,2007)。尽管前人已获得了少数花序调控基因,但这些基因的遗传关系及分子机制方面的研究还很匮乏,导致这些基因在番茄花序改良中的利用存在瓶颈。因此,挖掘和克隆调控番茄花序分枝的基因并加以利用,是解决这一问题的基础和关键,对番茄高产优质育种,保证番茄高产稳产具有重要的意义和生产应用价值。
发明内容
本发明所要解决的技术问题是如何调控植物花序发育,和/或,如何调控植物花序分枝数目。所要解决的技术问题不限于所描述的技术主题,本领域技术人员通过以下描述可以清楚地理解本文未提及的其它技术主题。
为解决上述技术问题,本发明首先提供了蛋白质或调控所述蛋白质活性和/或含量的物质的应用,所述应用可为下述任一种:
D1)蛋白质或调控所述蛋白质活性和/或含量的物质在调控植物花序发育中的应用;
D2)蛋白质或调控所述蛋白质活性和/或含量的物质在制备调控植物花序发育的产品中的应用;
D3)蛋白质或调控所述蛋白质活性和/或含量的物质在培育花序分枝数目增加的植物中的应用;
D4)蛋白质或调控所述蛋白质活性和/或含量的物质在制备培育花序分枝数目增加的植物的产品中的应用;
D5)蛋白质或调控所述蛋白质活性和/或含量的物质在植物育种中的应用;
所述蛋白质名称为MIB2,可为下述任一种:
A1)氨基酸序列是SEQ ID No.1的蛋白质;
A2)将SEQ ID No.1所示的氨基酸序列经过氨基酸残基的取代和/或缺失和/或添加得到的与A1)所示的蛋白质具有80%以上的同一性且具有相同功能的蛋白质;
A3)在A1)或A2)的N端和/或C端连接标签得到的具有相同功能的融合蛋白质。
为了使A1)中的蛋白质便于纯化或检测,可在由序列表中SEQ ID No.1所示的氨基酸序列组成的蛋白质的氨基末端或羧基末端连接标签蛋白。
所述标签蛋白包括但不限于:GST(谷胱甘肽巯基转移酶)标签蛋白、His6标签蛋白(His-tag)、MBP(麦芽糖结合蛋白)标签蛋白、Flag标签蛋白、SUMO标签蛋白、HA标签蛋白、Myc标签蛋白、eGFP(增强型绿色荧光蛋白)、eCFP(增强型青色荧光蛋白)、eYFP(增强型黄绿色荧光蛋白)、mCherry(单体红色荧光蛋白)或AviTag标签蛋白。
本领域普通技术人员可以很容易地采用已知的方法,例如定向进化或点突变的方法,对本发明的编码蛋白质MIB2的核苷酸序列进行突变。那些经过人工修饰的,具有与本发明分离得到的蛋白质MIB2的核苷酸序列75%或75%以上同一性的核苷酸,只要编码蛋白质MIB2且具有蛋白质MIB2功能,均是衍生于本发明的核苷酸序列并且等同于本发明的序列。
上述75%或75%以上同一性,可为80%、85%、90%或95%以上的同一性。
本文中,同一性是指氨基酸序列或核苷酸序列的同一性。可使用国际互联网上的同源性检索站点测定氨基酸序列的同一性,如NCBI主页网站的BLAST网页。例如,可在高级BLAST2.1中,通过使用blastp作为程序,将Expect值设置为10,将所有Filter设置为OFF,使用BLOSUM62作为Matrix,将Gap existence cost,Per residue gap cost和Lambda ratio分别设置为11,1和0.85(缺省值)并进行检索以对氨基酸序列的同一性进行计算,然后即可获得同一性的值(%)。
本文中,所述80%以上的同一性可为至少80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%的同一性。
本文中,调控所述蛋白质活性和/或含量的物质可为调控基因表达的物质,所述基因编码所述蛋白质MIB2。
上文中,所述调控基因表达的物质可为进行如下6种调控中至少一种调控的物质:1)在所述基因转录水平上进行的调控;2)在所述基因转录后进行的调控(也就是对所述基因的初级转录物的剪接或加工进行的调控);3)对所述基因的RNA转运进行的调控(也就是对所述基因的mRNA由细胞核向细胞质转运进行的调控);4)对所述基因的翻译进行的调控;5)对所述基因的mRNA降解进行的调控;6)对所述基因的翻译后的调控(也就是对所述基因翻译的蛋白质的活性进行调控)。
所述调控基因表达的物质具体可为B1)-B3)中任一所述的生物材料。
上述应用中,所述蛋白质可来源于番茄(Lycopersicon esculentum)。
本发明还提供了与所述蛋白质MIB2相关的生物材料的应用,所述应用可为下述任一种:
E1)与所述蛋白质MIB2相关的生物材料在调控植物花序发育中的应用;
E2)与所述蛋白质MIB2相关的生物材料在制备调控植物花序发育的产品中的应用;
E3)与所述蛋白质MIB2相关的生物材料在培育花序分枝数目增加的植物中的应用;
E4)与所述蛋白质MIB2相关的生物材料在制备培育花序分枝数目增加的植物的产品中的应用;
E5)与所述蛋白质MIB2相关的生物材料在植物育种中的应用;
所述生物材料可为下述B1)至B7)中的任一种:
B1)编码所述蛋白质MIB2的核酸分子;
B2)含有B1)所述核酸分子的表达盒;
B3)含有B1)所述核酸分子的重组载体、或含有B2)所述表达盒的重组载体;
B4)含有B1)所述核酸分子的重组微生物、或含有B2)所述表达盒的重组微生物、或含有B3)所述重组载体的重组微生物;
B5)含有B1)所述核酸分子的转基因植物细胞系、或含有B2)所述表达盒的转基因植物细胞系;
B6)含有B1)所述核酸分子的转基因植物组织、或含有B2)所述表达盒的转基因植物组织;
B7)含有B1)所述核酸分子的转基因植物器官、或含有B2)所述表达盒的转基因植物器官。
上述应用中,B1)所述核酸分子可为下述任一种:
C1)编码序列是SEQ ID No.2的cDNA分子;
C2)核苷酸序列是SEQ ID No.2的DNA分子。
SEQ ID No.2所示的DNA分子(番茄花序发育基因MIB2)编码氨基酸序列是SEQ IDNo.1的蛋白质MIB2。
B1)所述核酸分子还可包括在SEQ ID No.2所示核苷酸序列基础上经密码子偏好性改造得到的核酸分子。
所述核酸分子还包括与SEQ ID No.2所示的核苷酸序列一致性为95%以上且来源相同种属的核酸分子。
本文所述核酸分子可以是DNA,如cDNA、基因组DNA或重组DNA;所述核酸分子也可以是RNA,如gRNA、mRNA、siRNA、shRNA、sgRNA、miRNA或反义RNA。
本文所述载体是本领域技术人员公知的,包括但不限于:质粒、噬菌体(如λ噬菌体或M13丝状噬菌体等)、黏粒(即柯斯质粒)、Ti质粒、病毒载体等。在本发明的一个实施例中,所述载体为质粒载体pCAMBIA2300_YFP_HA。
可用现有的植物表达载体构建含有MIB2基因的重组表达载体。所述植物表达载体包括但不限于如双元农杆菌载体和可用于植物微弹轰击的载体等。所述植物表达载体还可包含外源基因的3'端非翻译区域,即包含聚腺苷酸信号和任何其它参与mRNA加工或基因表达的DNA片段。所述聚腺苷酸信号可引导聚腺苷酸加入到mRNA前体的3'端,如包括但不限于农杆菌冠瘿瘤诱导(Ti)质粒基因(如胭脂合成酶Nos基因)、植物基因(如大豆贮藏蛋白基因)3'端转录的非翻译区均具有类似功能。
使用MIB2基因构建重组植物表达载体时,在其转录起始核苷酸前可加上任何一种增强型启动子或组成型启动子,包括但不限于如花椰菜花叶病毒(CaMV)35S启动子、玉米的泛素启动子(ubiquitin),它们可单独使用或与其它植物启动子结合使用;此外,使用本发明的基因构建植物表达载体时,还可使用增强子,包括翻译增强子或转录增强子,这些增强子区域可以是ATG起始密码子或邻接区域起始密码子等,但必需与编码序列的阅读框相同,以保证整个序列的正确翻译。所述翻译控制信号和起始密码子的来源是广泛的,可以是天然的,也可以是合成的。翻译起始区域可以来白转录起始区域或结构基因。
为了便于对转基因植物细胞或植物进行鉴定及筛选,可对所用植物表达载体进行加工,如加入包括但不限于可在植物中表达的编码可产生颜色变化的酶或发光化合物的基因(GUS基因、荧光素酶基因等)、具有抗性的抗生素标记物(庆大霉素标记物、卡那霉素标记物等)或是抗化学试剂标记基因(如抗除草剂基因)等。从转基因植物的安全性考虑,可不加任何选择性标记基因,直接以逆境筛选转化植株。
本文所述微生物可为酵母、细菌、藻或真菌。其中,细菌可来自埃希氏菌属(Escherichia),欧文氏菌(Erwinia),根癌农杆菌属(Agrobacterium)、黄杆菌属(Flavobacterium),产碱菌属(Alcaligenes),假单胞菌属(Pseudomonas),芽胞杆菌属(Bacillus)等。
所述重组载体具体可重组载体pCAMBIA2300 MIB2 CTP。所述重组载体pCAMBIA2300 MIB2 CTP含有核苷酸序列是SEQ ID No.3的启动子和核苷酸序列是SEQ IDNo.2的MIB2基因cDNA分子。
本发明还提供了一种增加植物花序分枝数目的方法,所述方法包括提高目的植物中所述蛋白质MIB2的含量和/或活性。
上述方法中,所述提高目的植物中所述蛋白质MIB2的含量和/或活性通过提高目的植物中所述蛋白质MIB2的编码基因的表达量实现。
上述方法中,所述提高目的植物中所述蛋白质MIB2的编码基因的表达量通过将所述蛋白质MIB2的编码基因导入所述目的植物实现。
上述方法中,所述蛋白质的编码基因可为下述任一种:
F1)编码序列是SEQ ID No.2的cDNA分子;
F2)核苷酸序列是SEQ ID No.2的DNA分子。
具体地,在本发明的一个实施方案中,所述提高目的植物中所述蛋白质MIB2的编码基因的表达量通过将SEQ ID No.2所示的DNA分子导入所述目的植物实现。
本发明还提供了一种培育花序分枝数目增加的植物的方法,所述方法包括提高目的植物中所述蛋白质MIB2的含量和/或活性,得到花序分枝数目高于所述目的植物的花序分枝数目增加的植物。
进一步地,所述培育花序分枝数目增加的植物的方法包括如下步骤:
(1)构建包含SEQ ID No.2所示DNA分子的重组载体;
(2)将步骤(1)构建的重组载体转入目的植物(如作物或番茄)中;
(3)经筛选和鉴定获得花序分枝数目高于所述目的植物的花序分枝数目增加的植物。
本文中,所述植物可为G1)或G2):
G1)单子叶植物或双子叶植物;
G2)茄科植物、禾本科植物或十字花科植物。
本文中,所述植物可为作物(如农作物)。
本发明还提供了所述增加植物花序分枝数目的方法在创制花序分枝数目增加的植物和/或植物育种中的应用。
所述植物育种可为增加作物花序分枝数目的转基因育种。
所述调控植物花序发育可为调控植物花序分枝数目。
所述调控植物花序分枝数目可为增加植物花序分枝数目或减少植物花序分枝数目。
本文中,所述花序发育可为花序分枝发育。
本文中,所述花序分枝数目增加的植物理解为不仅包含将所述MIB2基因转化目的植物得到的第一代转基因植物,也包括其子代。可以在该物种中繁殖该基因,也可用常规育种技术将该基因转移进入相同物种的其它品种,特别包括商业品种中。所述花序分枝数目增加的植物包括种子、愈伤组织、完整植株和细胞。
本发明提供了一种调控番茄花序分枝数目的基因MIB2及其应用,所述基因MIB2是番茄花序分枝的关键调控基因,可应用到番茄高产优质品种的培育中。
通过将来源于番茄(Lycopersicon esculentum)的调控植物花序分枝的MIB2基因导入到受体植物中,可得到花序分枝数目增加的转基因植株,实验证明,MIB2基因突变能导致番茄花序分枝减少,在突变体中互补该基因能增加番茄花序分枝数目。
利用任何一种可以引导外源基因在植物中表达的载体,将本发明所提供的MIB2基因或基因的片段导入植物细胞或受体植物,可获得花序分枝数目增加的转基因细胞系及转基因植株。携带MIB2基因的表达载体可通过使用Ti质粒、Ri质粒、植物病毒载体、直接DNA转化、显微注射、电导、农杆菌介导等常规生物学方法转化植物细胞或组织,并将转化的植物组织培育成植株。
与现有技术相比,本发明具有以下优点:
本发明首次提出编码SEQ ID No.1所示蛋白的基因可调控(控制)番茄花序的分枝数目,通过花序分枝数目表型鉴定表明,MIB2基因突变,导致番茄花序分枝数目减少。在突变体中转入正常的MIB2基因,可以增加番茄花序分枝数目,从而证实了该基因的功能及应用途径,为番茄育种提供了一个良好的基因资源,在番茄育种领域具有广泛的应用前景。
附图说明
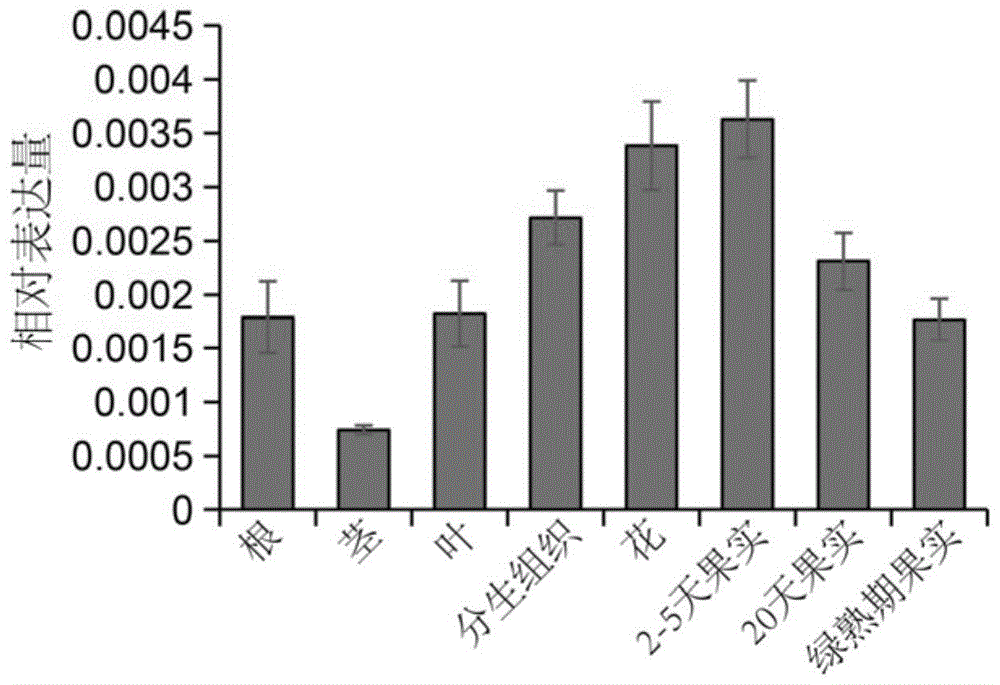
图1为MIB2基因在番茄不同组织中的表达模式。
图2为pCAMBIA2300 MIB2 CTP的质粒图谱。
图3为MIB2基因功能互补转基因植株花序表型。1,2,3,4,5,>5分别表示花序分枝数量。
图4为MIB2基因编辑植株突变类型。
图5为突变体AC/mib2-cr1和AC/mib2-cr2花序表型。1,2,3,4分别表示花序分枝数量。
具体实施方式
下面结合具体实施方式对本发明进行进一步的详细描述,给出的实施例仅为了阐明本发明,而不是为了限制本发明的范围。以下提供的实施例可作为本技术领域普通技术人员进行进一步改进的指南,并不以任何方式构成对本发明的限制。
下述实施例中的实验方法,如无特殊说明,均为常规方法,按照本领域内的文献所描述的技术或条件或者按照产品说明书进行。下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。
下述实施例中的番茄材料‘Ailsa Craig’(AC),其MIB2基因为野生型。公众可在美国加州大学戴维斯分校番茄遗传资源中心(Tomato Genetics Resource Center,网址为:https://tgrc.ucdavis.edu/)获得。Accession号为LA2838A。
下述实施例中的番茄品种Moneymaker(MM),其MIB2基因为突变型。公众可在美国加州大学戴维斯分校番茄遗传资源中心(Tomato Genetics Resource Center,网址为:https://tgrc.ucdavis.edu/)获得。Accession号为LA2706。
下述实施例中的基础质粒pCAMBIA2300_YFP_HA可从北京华越洋生物科技有限公司购得。
实施例1:MIB2基因在番茄不同组织和器官中的时空表达
本发明从番茄基因组中克隆了调控番花序分枝数目的基因Solyc02g093280(MIB2基因),经深入研究表明该基因是番茄花序分枝的关键调控基因。MIB2基因的编码序列(CDS)的核苷酸序列为SEQ ID No.2(由1137个核苷酸组成),MIB2基因编码氨基酸序列是SEQ ID No.1的蛋白质MIB2,SEQ ID No.1由378个氨基酸残基组成。
为明确MIB2基因在番茄花序发育过程中的作用,以番茄材料‘Ailsa Craig’(AC)的根、茎、叶、顶端分生组织、花芽、果实等部位的cDNA为模板,利用qR T-PCR技术检测MIB2基因的表达量变化。
用于检测MIB2基因表达量的引物序列如下:
MIB2-F:GAGAAGCAGAGCAGCTGAAGTGC;
MIB2-R:GCTGCTTAAGATACTCAATGGCTTC。
结果如图1所示,MIB2基因在花芽及果实中表达量较高,在顶端分生组织中也有一定表达,在根茎叶中表达量较低。表明MIB2基因可能在花序发育中发挥作用。
实施例2:构建MIB2基因功能互补表达载体
以番茄材料‘Ailsa Craig’(AC)的gDNA为模板,利用PCR技术扩增MIB2基因3.5kb的启动子(promoter)区(核苷酸序列为SEQ ID No.3)。
引物序列如下:
MIB2-F(P):
MIB2-R(P):
获得的PCR产物利用In-fusion无缝克隆技术(In-fusion无缝克隆试剂盒可从宝日医生物技术(北京)有限公司购得)连接至基础质粒pCAMBIA2300_YFP_HA。得到含有核苷酸序列是SEQ ID No.3的启动子的中间载体pCAMBIA2300_YFP_HA-MI B2-promoter。
以番茄材料‘Ailsa Craig’(AC)的顶端分生组织cDNA为模板,利用PCR技术扩增MIB2基因的cDNA。
引物序列如下:
MIB2-F1:AAggatcc ATGGCCAACAACAATGTTTATTATCAC;
MIB2-R1:AAggtacc GAAGTTGCTATACAGTTGTTGGGAAAG。
经PCR扩增后得到大小为1134bp的PCR产物MIB2-cDNA(对应SEQ ID No.2的第1-1134位),将该PCR产物与上述中间载体pCAMBIA2300_YFP_HA-MIB2-promot er分别利用BamHI和KpnI进行双酶切(37℃,8h),酶切产物进行胶回收纯化(胶回收试剂盒从北京全式金生物技术有限公司购得),然后利用T
质粒(重组互补表达载体pCAMBIA2300 MIB2 CTP)转化农杆菌AGL1感受态细胞,具体步骤如下:取出1支100uL农杆菌AGL1感受态(由北京全式金生物技术有限公司购得,-80℃冰箱中保存)在冰上融化;加入1uL上述质粒;冰浴30min;液氮环境1min;37℃热激5min;加入500uL无抗LB液体培养基(实验室自行配置);28℃180rpm震荡培养3h,用涂布器将上述菌液均匀涂抹在LB固体双抗培养基平板(实验室自行配置,双抗是指卡那霉素50ug/mL,利福平25ug/mL),待平板晾干后倒置于28℃环境中48h,挑取单克隆,进行阳性克隆筛选,备用。
实施例3:调查功能互补转基因株系花序分枝表型
以突变体MM(MM为番茄品种Moneymaker的简称,其MIB2基因为突变型,由美国加州大学戴维斯分校番茄遗传资源中心获得)为受体,通过根癌农杆菌AGL1介导的遗传转化法将互补质粒(pCAMBIA2300 MIB2 CTP)转入突变体MM中,通过基因型鉴定,获得两个独立的含有T-DNA插入的功能互补转基因株系,并通过后代鉴定获得稳定遗传的互补转基因纯合株系MM/MIB2-CTP1和MM/MIB2-CTP2。调查其花序分枝数目,每个株系10个植株。结果如图3所示,与对照MM相比,两个MIB2基因互补转基因株系MM/MIB2-CTP1和MM/MIB2-CTP2的花序分枝数目均有显著增加。表明MIB2基因具有调控花序发育的功能,能够增加番茄花序分枝数目。
实施例4:获得CRISPR/Cas9编辑突变体
根据Solyc02g093280基因(MIB2基因)的编码序列(SEQ ID No.2),通过CRI SPR-P(http://crispr.hzau.edu.cn/CRISPR/)设计了四个识别CRISPR/Cas9编辑靶点序列的sgRNA,所述sgRNA的靶序列为:
sgRNA1:5’-TCATCCGGATCGTAATCTA-3’(对应SEQ ID No.2的第317-335位),
sgRNA2:5’-GGGACCATAGATTACGATC-3’(对应SEQ ID No.2的第310-328位),
sgRNA3:5’-GCATTCGTGGCAGGCCAAG-3’(对应SEQ ID No.2的第625-643位),
sgRNA4:5’-TGCTGGTAGATCTTGAGAG-3’(对应SEQ ID No.2的第886-904位)。
设计引物用于扩增包含sgRNA的DNA片段,引物序列为:
MIB2-CR-F1:5’-ATATATGGTCTCGTTTGTCATCCGGATCGTAATCTAGTTTTAGAGCTAGAAATAGC-3’,
MIB2-CR-R1:5’-ATT
MIB2-CR-F2:5’-ATATATGGTCTCGGGGACCATAGATTACGATCGTTTTAGAGCTAGAAATAGC-3’,
MIB2-CR-R2:5’-ATTATTGGTCTCGATGCCAAACTACACTGTTAGATTC-3’,
MIB2-CR-F3:5’-ATATATGGTCTCGGCATTCGTGGCAGGCCAAGGTTTTAGAGCTAGAAATAGC-3’,
MIB2-CR-R3:5’-ATTATTGGTCTCGAAACTGCTGGTAGATCTTGAGAGCAAACTACACTGTTAGATTC-3’。
以pCBC_DT1T2_SlU6p质粒(该质粒记载在如下文献中:Li R,Sun S,Wang H J,Wang KT,Yu H,Zhou Z,Xin PY,Chu JF,Zhao TM,Wang HZ,Li JY,Cui X.2020.FIS1encodes a GA2-oxidase that regulates fruit firmness in to mato.NatureCommunications 11.)为模板,利用上述3对引物分别进行PCR扩增,得到三个DNA片段,然后进行胶回收纯化。与此同时,将CRISPR-Cas9载体用BsaI酶切(CRISPR-Cas9载体的名称为pTX041,来自中国科学院遗传发育研究所李传友课题组。BsaI为NEB公司产品),线性化的CRISPR-Cas9载体pTX041进行胶回收纯化。而后,在200uL RNase-free PCR管中依次加入:10×T4 DNA ligase buffer,1uL;0.1%BSA(Takara),1uL;纯化的线性载体pTX041,100ng;三个纯化的DNA片段,各30ng;BsaI酶(NEB),0.5uL;T4 DNA ligase(Promega),0.5uL;加超纯水至总体积10uL。将上述反应体系放入PCR仪中进行连接,进行如下反应:37℃,5min;16℃,10min;上述反应重复16个循环;50℃,5min;80℃,5min;10℃保存。上述连接产物转化大肠杆菌DH5α感受态细胞(方法同实施例2中的转化方法),卡那霉素抗性,进行阳性克隆筛选,测序验证序列的正确性(测序由华大基因完成),提取质粒,转化农杆菌AGL1感受态细胞(方法同实施例2中的转化方法),备用。
以野生型AC(AC为番茄品种Ailsa Craig的简称,其MIB2基因为野生型,由美国加州大学戴维斯分校番茄遗传资源中心获得)为受体进行AGL1农杆菌介导的遗传转化。对获得的8株再生植株中的Cas 9基因进行PCR检测,检测引物为Cas9-F:5’-CACTATCCTTCGCAAGACCC-3’和Cas9-R:5’-GAGATTCCCGAACAAGCCG-3’。对PCR产物进行凝胶电泳检测,有6株植物检测到Cas9基因。对含有MIB2基因的编辑位点的DNA片段进行PCR扩增,扩增引物为CR-MIB2-F:5’-CGTCATATTCTACTTCCTTCTTCGTC-3’和CR-MIB2-R:5’-GTGCACTTCAGCTGCTCTGCTTC-3’。PCR产物测序验证基因编辑的情况。最终获得2株纯合突变植株。将上述2株T0代植株单株留种获得T1代种子,播种后,于2叶期提取每个单株的DNA,利用引物Cas 9-F/R进行PCR扩增,凝胶电泳检测,选取不含Cas9的T1代植株,自交留种,获得两个独立的不同突变类型(mib2-cr1和mib2-cr2)的CRISPR株系AC/mib2-cr1和AC/mib2-cr2。
转基因株系AC/mib2-cr1和AC/mib2-cr2的扩增产物(含有MIB2基因的编辑位点的DNA片段)与野生型植株AC扩增产物的核苷酸序列比对结果如图4所示,其中突变类型mib2-cr1的第一个靶点缺失2个碱基(-AT),突变类型mib2-cr2的第一个靶点插入1个碱基(+A)。两种突变均导致蛋白翻译的提前终止,从而将MIB2基因敲除。
实施例5:调查CRISPR突变体花序分枝表型
为了鉴定CRISPR突变体材料的表型,将纯合突变且无Cas 9的CRISPR材料(AC/mib2-cr1,AC/mib2-cr2)和野生型(AC)种植于日光温室,调查花序分枝表型。结果如图5所示,与野生型(AC)相比,CRISPR突变体植株表现为花序分枝数目显著减少。该结果再次证明MIB2基因是番茄花序分枝发育重要调控基因,该基因突变导致番茄花序分枝数量减少。
本发明中所提供的花序分枝表型均进行了多年、多地观察,表型稳定。
以上对本发明进行了详述。对于本领域技术人员来说,在不脱离本发明的宗旨和范围,以及无需进行不必要的实验情况下,可在等同参数、浓度和条件下,在较宽范围内实施本发明。虽然本发明给出了特殊的实施例,应该理解为,可以对本发明作进一步的改进。总之,按本发明的原理,本申请欲包括任何变更、用途或对本发明的改进,包括脱离了本申请中已公开范围,而用本领域已知的常规技术进行的改变。
SEQUENCE LISTING
<110> 中国农业科学院蔬菜花卉研究所
<120> 调控番茄花序发育的基因及其编码蛋白与应用
<160> 3
<170> PatentIn version 3.5
<210> 1
<211> 378
<212> PRT
<213> 番茄(Lycopersicon esculentum)
<400> 1
Met Ala Asn Asn Asn Val Tyr Tyr His Asn Ala Asn Phe Ser Leu Thr
1 5 1015
Asp Pro Asp Pro Glu Pro Asp Asp Ile Ser Val Phe Leu Arg His Ile
202530
Leu Leu Pro Ser Ser Ser Ser Ser Ser Ser Ser Ser Asn Phe Met Ala
354045
Leu Lys Ser Asn Glu Met Gln Tyr Ser Ser Ser Leu Pro His Leu Met
505560
Pro Asn Asn Asn Gln Gln Gly Asn Leu Ser Ser Met Met Asn Ser Ser
65707580
Ala Cys Gly Ile Phe Ser Ser Ser Tyr Gly Val Cys Asn Gly Ala Thr
859095
Thr Val Ser Ser Ser Ser Val Gly Thr Ile Asp Tyr Asp Pro Asp Glu
100 105 110
Tyr Glu Cys Glu Ser Glu Asp Gly Thr Glu Asp Leu Gly Ala Glu Ala
115 120 125
Ser Val Gln Pro Pro Ser Arg Asn Thr Ser Lys Arg Ser Arg Ala Ala
130 135 140
Glu Val His Asn Leu Ser Glu Lys Arg Arg Arg Ser Arg Ile Asn Glu
145 150 155 160
Lys Met Lys Ala Leu Gln Lys Leu Ile Pro Asn Ser Asn Lys Thr Asp
165 170 175
Lys Ala Ser Met Leu Asp Glu Ala Ile Glu Tyr Leu Lys Gln Leu Gln
180 185 190
Leu Gln Val Gln Met Leu Thr Met Arg Asn Gly Leu Asn Met Tyr Pro
195 200 205
Leu Gly Leu Pro Arg Met Leu Gln Gln Asn Gln Leu Ser His Gln Lys
210 215 220
Val Gly Leu Cys Glu Gly Asn Ala Phe Thr Asn Ala Lys Val Ala Gly
225 230 235 240
Asn Leu Gln Val Asn Gln Asp Ala Ser Leu Asn Ala Ile Phe Asn Pro
245 250 255
Thr Glu Asn Cys Thr Glu Thr Lys Val Thr Pro Pro Ile Thr Met Ser
260 265 270
Asn Ile Asn Arg Ser Asp Ser Ala Phe Glu Leu Glu Ser Ser Met Asn
275 280 285
Ile His Leu Asp Pro Phe Gln Leu Ser Arg Ser Thr Ser Lys Glu Ile
290 295 300
Trp Arg Glu Asp Asp Leu Pro Leu Tyr Gly Met Asn Glu Leu Thr Thr
305 310 315 320
Lys Thr Ala Ser Thr Gly Ser Asn Leu Ala Phe Ser Val Pro Leu Asp
325 330 335
Thr Asp Ala Ser Asn Leu Lys Arg Ser Thr Arg Glu Ala Cys Leu Leu
340 345 350
Arg Tyr Gln Phe Gly Ala Val Asn Glu Thr Asn Leu Asp Cys Asp Gln
355 360 365
Leu Leu Ser Gln Gln Leu Tyr Ser Asn Phe
370 375
<210> 2
<211> 1137
<212> DNA
<213> 番茄(Lycopersicon esculentum)
<400> 2
atggccaaca acaatgttta ttatcacaat gctaattttt ctttgactga tcctgatcct 60
gaaccagatg atatttctgt gtttcttcgt catattctac ttccttcttc gtcttcgtct 120
tcatcttcat ctaattttat ggccctgaag agtaatgaaa tgcaatattc ctcatcatta 180
cctcatctta tgcctaataa taatcaacag ggtaatttgt catccatgat gaattcatca 240
gcttgcggca ttttctcgtc ttcctatggg gtttgtaatg gtgcaactac tgtttcttcc 300
tcatctgttg ggaccataga ttacgatccg gatgagtatg aatgtgaaag tgaggatggt 360
acagaggatt tgggggcgga agcttcagtt caaccgccat ctcgtaacac ttccaagaga 420
agcagagcag ctgaagtgca caatttgtct gaaaagagga gaagaagcag gattaatgag 480
aagatgaaag cattgcaaaa acttattcca aattcaaata agactgacaa ggcttcaatg 540
cttgatgaag ccattgagta tcttaagcag cttcagctcc aagtacagat gttgacaatg 600
agaaacgggt taaacatgta tcctcttggc ctgccacgaa tgctacaaca aaatcaactc 660
tcccatcaaa aagtaggttt gtgcgagggg aatgcattca caaatgccaa agtggctggg 720
aatctacaag ttaaccaaga cgcctcgtta aatgctattt tcaacccaac tgaaaattgt 780
acagaaacta aggtaacacc accgataact atgtcaaata taaaccggtc agacagtgcg 840
tttgagcttg agtcatcgat gaatattcac cttgatccct ttcagctctc aagatctacc 900
agcaaggaaa tttggaggga ggatgactta cctttatatg gtatgaatga gctaacgacc 960
aaaactgcat caactggatc aaatctggca ttctcagttc ccctagatac ggatgcatct 1020
aacctgaaga gaagcactcg ggaggcctgc ttgcttcgat atcagtttgg tgctgtgaat 1080
gaaaccaatc tggattgtga ccaacttctt tcccaacaac tgtatagcaa cttctga 1137
<210> 3
<211> 3582
<212> DNA
<213> 番茄(Lycopersicon esculentum)
<400> 3
gtattgctat ttggtttcca cttaagtaat tgttctatct aactttaatt tatcaaactt 60
aattcaataa aaaaaattga agataaattg cagcatgcta gtgttaaagt attactcaac 120
aatcagttga tgtgagttaa aaggactaca tattggtgaa aataataaat actatatcaa 180
ttagaataat aaattttaaa aagtaattat gtcactataa aatttattta catataaaat 240
aaatggttaa cagatataaa tatggggtgt gttttaacaa aatataaact tgtgaataca 300
ttttcgtatt aaaaaaaatc tcaaatcata atatgaaatt tctatatcat ggtttttgag 360
gaatatggaa tcaacatcta atgagacgaa atcagcataa aattacacat tcaaacacta 420
atatattata ttccctctat ttcaatttat gtaatacttt tcgttttttg agaatcaaac 480
aatttaagtt tgatcgagaa tttgcacatg aaatcttcaa tttttttgga tgaaatatat 540
atatttatac actatgtaaa aaatattata aattacaata attaagaatt taaaatattt 600
aaaaaatata acaaatttag tatcaaaatt ttactcaatt aaatcataaa atccaaaaaa 660
aatacatcat acatggccaa atatttactt agtgcgaaga gaaagatagt cagtcctctt 720
gggcggccga gagtgttatg taaaaaggac caagttgatg atcaattaat atgaaacgag 780
ggagggcgga atatagatag cctcgggaaa tcacgaaagg ctatttgctt ggcgtgtgcg 840
gaaatcagat gtgtgacggt tgatgatgcc tacttttcat acttgcagtg gtcctgcatg 900
tacaatagat atctactttt ggatacgtga tcttcttgtc caattgtgtg tagccacgtg 960
ttctattttg caagaaaaga cattgttggt atatatgtga ggagacagct actactattt 1020
acgaatcttg ggttctaaac tggaaaatac aaaagctttt gtcataggac tagtattaat 1080
aattattttc tttgtcacga tatttattag attaaaataa ttttatctta tctttaaaaa 1140
atttaacttt taatattatg acttatattt ttttatgtaa tttataaata tataaatttt 1200
atttttaaaa aaaattaaaa aaatttatat ataaattttc attcaaattt aagttgtttg 1260
attctcgaaa aaaaggaaaa atgtcacaac aattgagata aatagagtac tccatctgtt 1320
ccaattcata taataccttt tctgttttga gagtcaaaca acttaacttt gatagaaaat 1380
ttgtgtatga aatttttgat tttttaaaaa aatgaaattt atgtatttgt aaattacgta 1440
gaaagtacta cgaatcaccg gaattgatta ctcaaaatgt ttataaaata tttggaaaat 1500
ttacggtaaa aaataaactt gtttaactct tgaaattcga aaggagacat ataaaatggg 1560
acggaggtag taacataagt tacaaaaatt aaaaatttaa tgtgcaaatt ttcgatcaaa 1620
tcgaaattgt ttctcttcaa aaaaatgaaa agtttcatat aaattgagac agagagagta 1680
acatcaatat caacaagtat agagaatata tatatagaga gagagagaga gaaatctttt 1740
ggctagaaaa tttgattatt tggtcaaatt agtaaaataa catatttttt actatgatat 1800
ttttatcttt ttgaatattt tcattctttt taactctaat accataaatt taaaaataaa 1860
aaagtatcag attattttac agtaaataac aacattttac acttttgaaa tttctagtca 1920
aattctggtt atatagcatt atctatctat ttatctatat atatatatat atatatacaa 1980
aaatatgcaa atacgttaat gattaaaact atttactcac atgctaaaaa taatttatga 2040
taaaaatata ttatgtacat ataatttttg gacaaatgat attcatttga ccatttggag 2100
taacatagtt gtatcaataa agttttcaaa agattaaaaa atgaagagaa agtactacac 2160
ttaggtattt gattagctaa ctctgttttg gaaagcgact agtcattata attgaggaat 2220
caaatgattt tctgctgcta tttttcacat tacattgaat tggtaagtaa gaaaatgcag 2280
gacatcgtgt tttctttatc aaattaactc cctacacaca tagcctgcga ctataaagcc 2340
tatcaatcaa ttgaaacaag ggaatggagt tgtttggcct tgctgaaaac tatggttatt 2400
aatctgtctt aggagtacga ttgtactttt cttccttaat cagaagtttt aaatatattt 2460
tttttaatag gaagcattat cttgaataag gctttattct aacgagattg ttgtctataa 2520
attttctaaa tgttgcccaa cttattgatt ttattataaa actcattagt tttaaaaagt 2580
gatgtgtcaa aacaaaatgt caatgaacat tcaatctttt cctctgaact tcaaaaggag 2640
ttctactttt ttctttattg aatattacat ttcttatgtc ccaattttaa aatttatcat 2700
agttgcttca ataattgata aactttacga tagatctaaa gattaatgtt atattaaatt 2760
ttttaaaaaa aatatatcta aaaccttata catacactct ggtcttatcg tgccactttt 2820
tatctatgga gagtcaactt aaataacttt aaaattaaat tagactaaat aactttggta 2880
tattctaaaa tcataagttg gtacagtagt tatttcggac cagaagtggg gtaagcagca 2940
atcgtgtttt caaaaatact ttagaaaaca taaagaaaag taaaaaggag agcacaaatg 3000
gatactgaaa tggaacagtg ttcgattaaa gtgtaacgta tatactctct ctggaactga 3060
gctagttgaa gagcatgcct acaattgtgc ttcacgtgtg ccagaaagtt ggcttcacct 3120
gacactatca ctatataaaa ccactccatc aaattgaact taatttaatt cgataaaaat 3180
atttttgaat tttatgattt taattaaaaa tttgtgaaag acctatggat tttaaattga 3240
tacaacggaa gtaacaattg tactattaac cctgtcgatt tctttgattt ggttttagca 3300
gtttaccaac ttccctaatt catcagttct cagaaccaaa agtagaccag tcaaagtcta 3360
atcaacttgc tagtttaccc ctactctgtt aattcaattg gtttattcac tttaatgcga 3420
atccgcccac tctgctctat atggtatggt tggctaataa atacacactc gttcggagca 3480
tatacccaaa aaatgtcacc tactcctaca tgtaaaacag tgtgaacgta gacaaaatta 3540
acttgatctg ctcaacactg aacttgattt tgctggacta ta 3582
- ZmDRR206蛋白质及其编码基因在调控植物抗病性与生长发育中的应用
- 一种水稻花器官发育调控基因PEH1及其编码的蛋白质和应用
- 百脉根花型和花序结构调控基因、编码蛋白及其应用
- 百脉根花型和花序结构调控基因、编码蛋白及其应用