一种肺纤维化标志物及其应用
文献发布时间:2023-06-19 11:22:42
技术领域
本发明涉及生物技术领域,具体涉及一种肺纤维化标志物及其应用。
背景技术
肺纤维化(PF)是许多弥漫性实质性肺疾病的终末期。其特征是过多的基质形成导致正常肺部结构的破坏并最终可以导致死亡。在PF期间,成纤维细胞显示出较强的增殖能力,侵袭成纤维细胞并促使其向肌成纤维细胞转化,最终导致肺实质ECM(细胞外基质)的产生,称之为成纤维细胞的活化。
据报道,多种调节剂参与了成纤维细胞活化的启动和维持,包括在PF患者中与肺肌成纤维细胞活化相关的蛋白质编码或非编码RNA(ncRNA)的表达失调。非编码RNA(ncRNA)是指一大批没有蛋白质编码能力的内源RNA分子,分为两大类,即MicroRNA(miRNA;大约22个核苷酸)和长lncRNA(lncRNA;长于200个核苷酸))。PF中已广泛报道了miRNA的关键作用,包括miR-29,miR-101,miR-26a和miR-18a。已有研究证明miR-627/HMGB1的轴与RAGE/NF-κB信号形成调节环,以调节TGFβ1诱导的正常人原代肺成纤维细胞(NHLF)的体外活化。在NHLF中,TGFβ1刺激显著下调了miR-627的表达;在PF组织样本中,miR-627的表达显著下调。然而,尚不清楚在NHLF活化和PF过程中导致miR-627失调的上游因素。
越来越多的证据表明,lncRNAs在表观遗传,转录和转录后水平上参与了各种病理生理过程,例如基因组印迹,细胞命运的决定,选择性剪接以及作为ceRNA(竞争性内源性RNA)起作用。
发明内容
本发明提供一种检测MIR155HG基因或其部分片段表达水平的物质在制备诊断或辅助诊断肺纤维化的产品中的应用。
可选的,所述检测MIR155HG基因或其部分片段表达水平的物质为如下1)或2);
1)由序列表中SEQ ID NO:2所示的单链DNA分子和序列表中序列3所示的单链DNA分子组成的引物对A;
2)由序列A所示的单链DNA分子和序列B所示的单链DNA分子组成的引物对B;
所述序列A为将SEQ ID NO:2删除或增加或改变一个或几个核苷酸,且与SEQ IDNO:2具有相同功能的核苷酸;
所述序列B为将序列3删除或增加或改变一个或几个核苷酸,且与序列3具有相同功能的核苷酸。
一种检测或辅助检测待测患者是否为肺纤维化患者的试剂盒,包括检测MIR155HG基因或其部分片段表达水平的物质。
可选的,所述检测MIR155HG基因或其部分片段表达水平的物质为如下1)或2)或3):
1)由序列表中SEQ ID NO:2所示的单链DNA分子和序列表中序列3所示的单链DNA分子组成的引物对A;
2)由序列A所示的单链DNA分子和序列B所示的单链DNA分子组成的引物对B;
所述序列A为将SEQ ID NO:2删除或增加或改变一个或几个核苷酸,且与SEQ IDNO:2具有相同功能的核苷酸;
所述序列B为将序列3删除或增加或改变一个或几个核苷酸,且与序列3具有相同功能的核苷酸;
3)含有1)所述引物对A或2)所述引物对B的PCR试剂。
上述的试剂盒在诊断或辅助诊断肺纤维化中的应用。
MIR155HG基因或其部分片段作为标志物在诊断或辅助诊断肺纤维化中的应用。
调节MIR155HG基因表达量的物质在制备调节成纤维细胞增殖和细胞外基质沉积产品中的应用。
可选的,所述调节MIR155HG基因表达量为促进MIR155HG基因表达或抑制MIR155HG基因表达。
可选的,所述促进MIR155HG基因表达为在细胞中导入表达MIR155HG基因的物质;
抑制MIR155HG基因表达为在细胞中导入靶向MIR155HG基因的shRNA或能够转录成所述shRNA的DNA。
所述shRNA是由表达shRNA的慢病毒表达得到的;
表达shRNA的慢病毒的制备方法为:
F:GATCCGCATTCACATGGAACAAATTGCTCGAGCAATTTGTTCCATGTGAATGCTTTTTG和R:AATTCAAAAAGCATTCACATGGAACAAATTGCTCGAGCAATTTGTTCCATGTGAATGCG退火,得到Sh1-MIR155HG-Oligo DNA;BamHI和EcoRI酶切载体PLVX-shRNA2-puro和Sh1-MIR155HG-OligoDNA,酶切后的PLVX-shRNA2-puro和Sh1-MIR155HG-Oligo DNA连接,得到Sh1-MIR155HGPLVX-shRNA2-puro,该重组慢病毒载体转入HEK293A细胞,包装得到表达shRNA的慢病毒。
可选的,所述表达MIR155HG基因的物质为如下A)或B)
A)重组载体,将核苷酸序列如SEQ ID NO:29所示DNA分子插入动物表达载体得到的重组载体;
B)慢病毒颗粒,将核苷酸序列如SEQ ID NO:29所示DNA分子插入PLVX-shRNA2-puro慢病毒载体,所述慢病毒载体包装成的慢病毒颗粒。
本发明技术方案,具有如下优点:
1、在本研究中,发明人选择了数据集GSE121241,该数据集报告了由IL-1β(一种NF-κB信号激活剂)诱导的PF中差异表达的lncRNA。在7个显著上调的lncRNA中使用LncTar在线工具进行预测,发现了了3个可靶向miR-627的lncRNA。然后,用TGFβ1诱导NHLF活化并检测三种lncRNA的表达,分别使三种lncRNA沉默,并检测miR-627的表达。最后,选择了MIR155HG。验证了MIR155HG与miR-627的靶向结合。研究了MIR155HG的特异性作用以及MIR155HG/miR-627轴对TGFβ1诱导的NHLF活化的作用。接下来,为了研究NF-κB/MIR155HG/miR-627/HMGB1调节环,发明人研究了miR-627对p65磷酸化的影响以及NF-κB对MIR155HG表达的影响。最后,检测验证了组织样品中MIR155HG,miR-627和HMGB1的表达及其相关性。检测验证了HMGB1和p-65磷酸化的蛋白水平。本申请证明了NF-κB/MIR155HG/miR-627/HMGB1调控环可以调节TGFβ1诱导的NHLF活化。
2、MIR155HG过表达增强TGFβ1诱导的NHLF活化,实验结果证明MIR155HG沉默可减弱TGFβ1诱导的NHLF增殖和ECM沉积,而MIR155HG过表达可增强TGFβ1诱导的NHLF增殖和ECM沉积。
附图说明
为了更清楚地说明本发明具体实施方式或现有技术中的技术方案,下面将对具体实施方式或现有技术描述中所需要使用的附图作简单地介绍,显而易见地,下面描述中的附图是本发明的一些实施方式,对于本领域普通技术人员来讲,在不付出创造性劳动的前提下,还可以根据这些附图获得其他的附图。
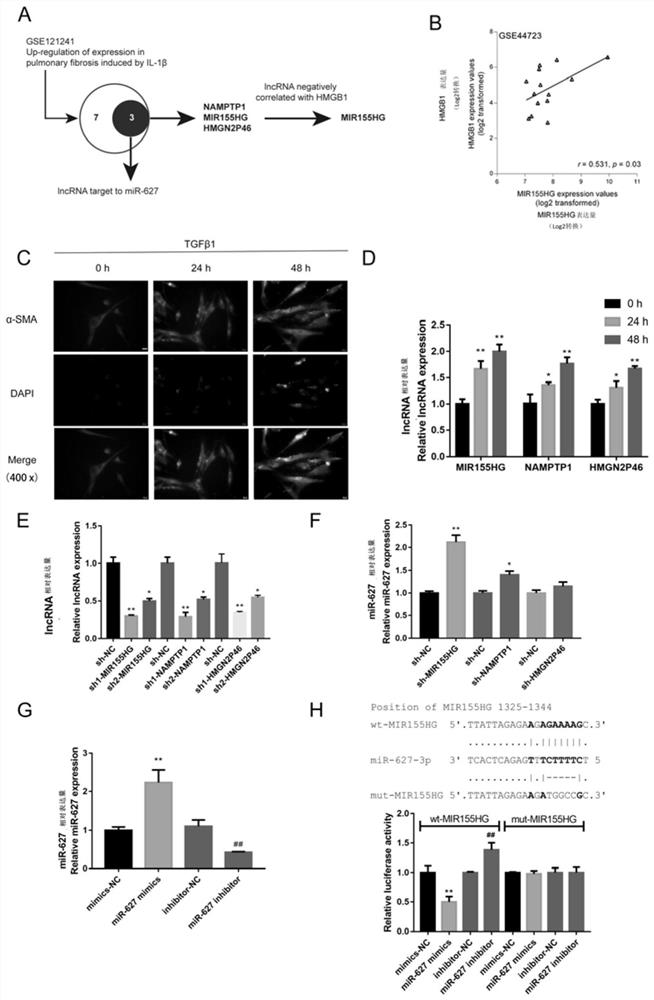
图1选择与miR-627和HMGB1相关的引起NF-κB改变的lncRNA
(A)示意图显示选择在PF发病过程中可能与miR-627和HMGB1相关的IL-1β改变的lncRNA,发现了LncRNA MIR155HG;
(B)根据GSE44723分析了MIR155HG与HMGB1表达之间的相关性;
(C)正常人肺成纤维细胞(NHLFs)用10ng/mL TGFβ1分别刺激0、24或48h,并通过免疫荧光(IF)染色检测α-SMA的蛋白质含量和分布;图中标记α-SMA表示单克隆抗体α-SMA与细胞中α-SMA反应,通过荧光标记二抗结合产生绿色荧光;图中标记DAPI表示荧光染料DAPI对肺纤维化细胞的细胞核染色结果;图中标记Merge表示DAPI及α-SMA融合免疫荧光结果;
(D)正常人肺成纤维细胞(NHLFs)用10ng/mL TGFβ1分别刺激0、24或48h,通过RT-qPCR检测lncRNA MIR155HG,NAMPTP1和HMGN2P46表达;
(E)通过转染MIR155HG,NAMPTP1或HMGN2P46的“短发夹RNA”(sh1/2-MIR155HG,sh1/2-NAMPTP1或sh1/2)在NHLF中实现MIR155HG,NAMPTP1或HMGN2P46的沉默;sh-NC为阴性对照;通过RT-qPCR验证转染效率;
(F)用sh-MIR155HG,sh-NAMPTP1或sh-HMGN2P46转染NHLF,并通过RT-qPCR检测miR-627的表达;
(G)miR-627过表达或抑制通过转染miR-627mimics或miR-627inhibitor在NHLF中实现;RT-qPCR验证转染效率;
(H)构建MIR155HG野生型和突变型荧光素酶报道质粒,并用miR-627mimics或miR-627inhibitor转染293T细胞;测定萤光素酶活性;*P<0.05,**P<0.01,##P<0.01。
图2MIR155HG的过表达增强了TGFβ1诱导的NHLF活化;
(A)通过转染过表达MIR155HG的慢病毒(lv-MIR155HG),在NHLF中实现了MIR155HG的过表达;lv-NC为阴性对照,通过RT-qPCR验证转染效率;
(B)通过lv-MIR155HG转染NHLF,并通过RT-qPCR检测miR-627的表达;
(C)用sh-MIR155HG转染NHLF或用lv-MIR155HG转染NHLF,用10ng/mL TGFβ1刺激48小时,并通过免疫印迹检测HMGB1,p-65和p-p65的蛋白水平;将左图的结果进行统计学分析做柱状图,得到右图;
(D)用sh-MIR155HG转染NHLF或用lv-MIR155HG转染NHLF,用10ng/mL TGFβ1刺激48小时,通过EdU分析细胞增殖能力,图中标记EdU代表细胞中DNA的复制活性及合成能力;图中标记DAPI表示荧光染料DAPI对肺纤维化细胞的细胞核染色结果;图中标记Merge表示DAPI及EdU融合免疫荧光结果图片;
(E)用sh-MIR155HG转染NHLF或用lv-MIR155HG转染NHLF,用10ng/mL TGFβ1刺激48小时,通过IF染色检测α-SMA的蛋白质含量和分布;图中标记α-SMA表示单克隆抗体α-SMA与细胞中α-SMA反应,通过荧光标记二抗结合产生绿色荧光;图中标记DAPI表示荧光染料DAPI对肺纤维化细胞的细胞核染色结果;图中标记Merge表示DAPI及α-SMA融合免疫荧光结果;
(F)用sh-MIR155HG转染NHLF或用lv-MIR155HG转染NHLF,用10ng/mLTGFβ1刺激48小时,通过免疫印迹检测胶原I,α-SMA和E-cadherin的蛋白水平;右图为将作图结果作为柱状图所得;**P<0.01,##P<0.01;
图3MIR155HG和miR-627对TGFβ1刺激的NHLF的动态影响
(A)用lv-MIR155HG和miR-627mimics共转染NHLF,通过RT-qPCR检测miR-627的表达;
(B)用lv-MIR155HG和miR-627mimics共转染NHLF,用10ng/mLTGFβ1刺激48小时,并通过免疫印迹检测HMGB1,p-65和p-p65的蛋白水平;将作图进行统计学分析做柱状图,得到右图;
(C)用lv-MIR155HG和miR-627mimics共转染NHLF,用10ng/mLTGFβ1刺激48小时,通过EdU分析NHLF的DNA合成能力;
(D)用lv-MIR155HG和miR-627mimics共转染NHLF,用10ng/mL TGFβ1刺激48小时,IF染色检测α-SMA的蛋白质含量和分布;图中标记α-SMA表示单克隆抗体α-SMA与细胞中α-SMA反应,通过荧光标记二抗结合产生绿色荧光;图中标记DAPI表示荧光染料DAPI对肺纤维化细胞的细胞核染色结果;图中标记Merge表示DAPI及α-SMA融合免疫荧光结果;
(E)通过免疫印迹检测胶原I,α-SMA和E-cadherin的蛋白水平;右图为左图分析后所做柱状图;**P<0.01,##P<0.01。
图4MIR155HG/miR-627轴与NF-κB形成调节环
(A)用miR-627inhibitor转染NHLFs,与10ng/ml TGFβ1和/或JSH-23(NF-κB抑制剂)处理,并用免疫印迹法检测p-65和p-p65的蛋白水平;
(B)用miR-627inhibitor转染NHLFs,与10ng/ml TGFβ1和/或JSH-23(NF-κB抑制剂)处理RT-qPCR检测MIR155HG的表达;
(C)用过表达NF-κB(lv-NF-κB)的慢病毒转染NHLF以获得NF-κB在NHLF的过度表达,lv-NC为阴性对照;通过RT-qPCR验证转染效率;
(D)用lv-NF-κB和/或JSH-23处理转染NHLF,并通过RT-qPCR检测MIR155HG的表达,**P<0.01,##P<0.01;
图5NF-κB/MIR155HG/miR-627/HMGB1调控环在组织样品中的表达及相关性
(A)通过RT-qPCR测定19例PF组织和正常组织中MIR155HG的表达;
(B)通过皮尔逊相关系数分析组织样品中MIR155HG,miR-627和HMGB1的表达相关性;
(C)通过免疫印迹法测定19例PF和正常组织中HMGB1,p-p65和p65的蛋白水平;将作图进行统计学分析后做柱状图得到右图;
(D)通过H&E染色检测PF和正常组织的组织病理学特征;
(E)通过免疫组织化学(IHC)染色测定组织样品中HMGB1和NF-κB的蛋白质含量和分布,**P<0.01。
具体实施方式
方法
(1)细胞来源
正常人原代肺成纤维细胞NHLF购自Lonza Ltd.(目录号:CC-2512;瑞士巴塞尔)。在本研究中使用了第3和第8段之间的NHLF。在含有5%(g/ml)胎牛血清(FCS,Sigma-Aldrich)、100U/ml青霉素和100mg/ml链霉素的DMEM/Ham's F-12培养基(Sigma-Aldrich,St.Louis,MO,USA)中培养细胞。在5%CO
表1引物序列
表2
表3
(2)细胞转染
通过转染Sh1-MIR155HGPLVX-shRNA2-puro、Sh2-MIR155HGPLVX-shRNA2-puro、Sh1-HMGN2P46PLVX-shRNA2-puro、Sh2-HMGN2P46PLVX-shRNA2-puro、Sh1-NAMPTP1PLVX-shRNA2-puro、Sh2-NAMPTP1PLVX-shRNA2-puro质粒在NHLF中实现MIR155HG,NNAMPTP1或HMGN2P46的干扰;将sh-NC(空载体PLVX-shRNA2-puro质粒)转染作为阴性对照。
Sh1-MIR155HGPLVX-shRNA2-puro能够表达shRNA干扰MIR155HG表达,Sh1-MIR155HGPLVX-shRNA2-puro重组慢病毒载体的构建过程为:经过Oligo DNA(F:GATCCGCATTCACATGGAACAAATTGCTCGAGCAATTTGTTCCATGTGAATGCTTTTTG和R:AATTCAAAAAGCATTCACATGGAACAAATTGCTCGAGCAATTTGTTCCATGTGAATGCG)的退火,得到Sh1-MIR155HG-Oligo DNA;BamHI和EcoRI酶切载体PLVX-shRNA2-puro和Sh1-MIR155HG-Oligo DNA,连接,得到Sh1-MIR155HGPLVX-shRNA2-puro。
Sh2-MIR155HGPLVX-shRNA2-puro能够表达shRNA干扰MIR155HG表达,经过OligoDNA(F:GATCCGGTTCAAGTTCAATAGCTTAGCTCGAGCTAAGCTATTGAACTTGAACCTTTTTG;R:AATTCAAAAAGGTTCAAGTTCAATAGCTTAGCTCGAGCTAAGCTATTGAACTTGAACCG)的退火,得到Sh2-MIR155HG-Oligo DNA,BamHI和EcoRI酶切载体PLVX-shRNA2-puro和Sh2-MIR155HG-Oligo DNA,将Sh2-MIR155HG-Oligo DNA与酶切后的载体连接得到Sh2-MIR155HGPLVX-shRNA2-puro。
同理得到Sh1-HMGN2P46 PLVX-shRNA2-puro和Sh2-HMGN2P46PLVX-shRNA2-puro重组载体,能够表达shRNA干扰HMGN2P46的表达;干扰NAMPTP1表达的Sh1-NAMPTP1 PLVX-shRNA2-puro和Sh2-NAMPTP1 PLVX-shRNA2-puro重组载体。所用的引物序列见表2。酶切所用酶为BamHI和EcoRI。
将上述获得的重组慢病毒载体转入HEK293A细胞,包装得到表达对应shRNA的慢病毒。
使用Lipofectamine 3000试剂(Thermo Fisher Scientific,美国马萨诸塞州沃尔瑟姆)进行转染。将过表达MIR155HG的慢病毒(lv-MIR155HG)转染NHLF,以实现MIR155HG过表达;将过表达NF-kB的慢病毒(lv-NF-kB)转染NHLF,以实现NF-kB过表达;lv-NC(PLVX-puro空质粒)为阴性对照。所有转染质粒和慢病毒均购自GenePharma(中国上海)。根据制造商的说明,使用ViraPower慢病毒表达系统(Invitrogen)进行慢病毒转染。将MIR155HG的核苷酸分子(SEQ ID NO:29)插入PLVX-puro慢病毒载体,形成Lv-MIR155HG pLVX-puro,该重组慢病毒载体过表达MIR155HG基因;同理,将NF-kB的核酸分子(SEQ ID NO:30)插入PLVX-puro慢病毒载体,形成Lv-NF-kBpLVX-puro,该重组慢病毒载体过表达NF-kB蛋白。将上述获得的重组慢病毒载体转入HEK293A细胞,包装得到过表达NF-kB的慢病毒(lv-NF-kB),过表达MIR155HG的慢病毒(lv-MIR155HG)。
(3)免疫荧光(IF)实验
用10ng/mL TGFβ1刺激NHLFs指定时间,用PBS洗涤3次,并在4%甲醛中固定30分钟。然后将细胞在4℃下于含有0.1%(v/v)Triton和0.1%(g/ml)柠檬酸钠的缓冲液中孵育5分钟,以提高细胞通透性,然后与单克隆抗体α-SMA(55135-1-AP;Proteintech,Rosemont,IL,USA)一起孵育。然后,将细胞与标记有Alexa Fluor 488山羊抗兔IgG(Invitrogen,Carlsbad,CA,USA)的二抗孵育,以进行蛋白质可视化。DAPI用于细胞核染色。获得并显示了代表性图像。绿色荧光代表α-SMA,蓝色代表核。
(4)RT-qPCR
将NHLF以1×10
(4)双荧光素酶报告基因检测
为了验证与miR-627-3p结合的MIR155HG,进行了双荧光素酶报告基因测定。使用NHLF细胞系的基因组DNA通过PCR扩增MIR155HG,并将其克隆到Renilla psiCHECK2载体(Promega,Madison,WI,USA)中的Renilla荧光素酶开放阅读框的下游。所用的引物为Mut-MIR155G引物(SEQ ID NO:31和SEQ ID NO:32),将突变引入到MIR155HG中的miR-627-3p结合位点的种子区域(seed region),并将该载体命名为Mut-MIR155GpsiCHECK-2reportervectors载体。
将上述步骤中的Mut-MIR155G引物替换为Wt-MIR155HG引物进行实验,获得Wt-MIR155HGpsiCHECK-2reporter vectors载体。
将293T细胞接种到96孔板中,mut-MIR155HG/wt-MIR155HG psiCHECK-2reportervectors载体与miR-627mimics/inhibitor共转染到293T细胞中。转染48小时后,根据制造商的说明,使用Dual-Luciferase Reporter Assay System(Promega)测量荧光素酶活性。将值对荧光素酶活性和用空psiCHECK-2对照载体转染的细胞进行双重标准化。
(5)免疫印迹
裂解细胞,并将裂解物用于SDS-PAGE以分离蛋白质。将分离的蛋白质电印迹到聚偏二氟乙烯(PVDF)膜(Thermo Fisher Scientific)上。用含0.05%(v/v)Tween 20(PBS–Tween)和3%(g/mL)脱脂牛奶的PBS封闭后,膜分别用适当的一抗探测2h,然后与辣根过氧化物酶结合的抗山羊或-兔IgG(在PBS–Tween中以1:5000稀释)孵育2h,并用ECL Western-blot观察系统(Sigma-Aldrich)。使用的一抗如下:anti-HMGB1(ab77302,Abcam,Cambridge,MA,USA),anti-p65(10745-1-AP,Proteintech),anti-p-p65(ab194726,Abcam),anti-胶原I(ab90395,Abcam),anti-α-SMA(14395-1-AP,Proteintech),anti-E-cadherin(20874-1-AP,Proteintech)。
(6)基于EdU分析DNA合成能力
使用Click-IT EdU Alexa Fluor 647试剂盒(Thermo Fisher,Waltham,MA,USA)进行EdU分析。进行了阿波罗染色和用于核染色的DAPI染色。在荧光显微镜下,蓝色荧光代表被DAPI染色的细胞核,红色荧光代表被EdU染色的新合成的DNA。EdU的阳性率等于EdU阳性细胞(红色)/DAPI阳性细胞(蓝色)的比率。
(7)临床样本
如先前的研究所述进行临床采样[Li,J.,et al.,miR-627/HMGB1/NF-kappaBregulatory loop modulates TGF-beta1-induced pulmonary fibrosis.J CellBiochem,2019.120(3):p.2983-2993.]。从接受肺活检的患者中获得19个PF组织。从肺癌切除术切除的周围肺的正常区域获得了19个对照肺组织。经江西省伦理委员会批准(批准文号:2019(5)),所有组织均在江西省胸科医院省胸科医院收集,是根据赫尔辛基宣言实施的。每位登记患者均签署知情同意书。
(8)基于HE染色的组织学分析
将组织标本在室温下用4%多聚甲醛固定10分钟,包埋在石蜡中,切成4-μm的切片。将切片用苏木精/伊红(H&E)染色以观察形态变化。
(9)免疫组化
将组织标本在室温下用4%多聚甲醛固定10分钟,包埋在石蜡中,切成4-μm的切片。用二甲苯孵育液分别进行10分钟的去石蜡作用,然后在浓度递减的乙醇系列和水中再水化。在98℃的10mM柠檬酸钠缓冲液(Dako,Carpentaria,CA,美国)中进行抗原提取20分钟。然后将组织切片在室温下冷却30分钟,并用0.1%Triton-X溶液(Sigma Aldrich,密苏里州圣路易斯)渗透15分钟。然后,将切片用1%牛血清白蛋白BSA溶液洗涤,在室温下于10%山羊血清(Gibco:货号16210064)中孵育60分钟以阻断非特异性结合,以阻断非特异性结合。吸出封闭液后,将切片与anti-HMGB1(ab77302,Abcam)或anti-NF-κB(10745-1-AP,Proteintech)在4℃孵育过夜。然后,将切片用1%的水洗涤3次每次5分钟,然后与生物素偶联的抗IgG(Boster,中国)和SABC溶液一起孵育。根据产品说明,通过在光学显微镜下孵育二氨基联苯胺(DAB)(中国,Boster)。
(10)统计分析
使用配对的双尾t检验进行两组比较,或使用单向方差分析与Tukey检验进行多次比较,进行统计分析。所有数据均以均值±标准差(SD)表示。使用GraphPad(Graph PadSoftware,美国加利福尼亚州圣地亚哥)进行统计分析。P值小于0.05被认为具有统计学差异。
实施例1
1、选择可能与miR-627和HMGB1相关的lncRNA
为了选择在IL-1β诱导的肺成纤维细胞活化过程中被失调的lncRNA,发明人下载并分析了GSE121241,发现总共7种lncRNA(AL138828.1,XLOC_405581,XLOC_387302,MIR155HG,BISPR,NAMPTP1,HMGN2P46)被IL-1β显著上调(p<0.0001,fdr<0.01,log2FC>1.5)(图1A)。其中,lncTar预测MIR155HG,NAMPTP1和HMGN2P46靶向hsa-miR-627-3p(图1A)。根据对人成纤维细胞(GSE44723)的基因表达微阵列分析,在三种lncRNA中,MIR155HG与HMGB1正相关(图1A-B)。接下来,发明人用10ng/ml TGFβ1持续0、24或48h刺激正常的人肺成纤维细胞(NHLF)诱导NHLF活化;如通过IF染色所揭示的,通过TGFβ1刺激持续24或48小时,α-SMA的蛋白质含量增加了(图1C),表明TGFβ1激活了NHLF。同时,TGFβ1刺激24或48小时,lncRNAMIR155HG,NAMPTP1和HMGN2P46的表达显著上调,MIR155HG上调更明显(图1D)。接下来,通过转染MIR155HG,NAMPTP1或HMGN2P46的sh-RNA(sh1/2-MIR155HG,sh1/2-NAMPTP1或sh1/2-HMGN2P46)在NHLF中实现MIR155HG,NAMPTP1或HMGN2P46的干扰,sh-NC为阴性对照;如RT-qPCR所示,MIR155HG所用的引物为RT-PCRMIR155HG,NAMPTP1所用的引物为RT-PCRNAMPTP1,HMGN2P46所用的引物为RT-PCRHMGN2P46,结果见图1。在MIR155HG、NAMPTP1或HMGN2P46干扰的NHLF中,MIR155HG干扰的NHLFs的miR-627表达显著上调(图1F)。考虑到所有上述发现,选择了MIR155HG进行进一步的实验。
然后,通过以下方法在NHLF中实现了miR-627过表达或抑制作用:
RT-qPCR证实,通过转染miR-627mimics/inhibitor(即表中的miR-627-3p mimics和miR-627-3p inhibitor)可在NHLF中实现miR-627的过表达或抑制(图1G)。对于双荧光素酶报道基因测定,相应地构建了两种不同类型的MIR155HG荧光素酶报道基因质粒,即野生型和突变型,并与miR-627mimics/inhibitor共转染到293T细胞中。如图1H所示,miR-627的过表达被抑制,而miR-627的抑制增强了wt-MIR155HG荧光素酶的活性。当与mut-MIR155HG共转染时,miR-627的过表达或抑制作用无法改变mut-MIR155HG的荧光素酶活性。这些数据表明,MIR155HG靶向结合miR-627-3p。
2、MIR155HG过表达增强TGFβ1诱导的NHLF活化
在确认了MIR155HG与miR-617之间的直接结合后,接下来,实现了MIR155HG在NHLF中的过表达,以检测其对TGFβ1诱导的NHLF活化的特异性作用。将过表达MIR155HG的慢病毒(lv-MIR155HG)转染NHLF;lv-NC为阴性对照,通过RT-qPCR检测(图2A)。在NHLF中,MIR155HG的过表达显著下调了miR-627的表达(图2B)。然后,用sh-MIR155HG转染NHLF或用lv-MIR155HG转染NHLF,用10ng/ml TGFβ1刺激48小时,并检测NHLF的活化。在TGFβ1刺激的NHLF中,MIR155HG干扰降低了HMGB1的蛋白水平并抑制了p65磷酸化,而MIR155HG的过表达发挥了相反的作用,这表明MIR155HG积极调节HMGB1的表达和NF-κB的活化(图2C)。在TGFβ1刺激下,MIR155HG干扰受到抑制,而MIR155HG的过表达促进了NHLF DNA的合成能力(图2D)。如IF染色所示,MIR155HG干扰减弱,而MIR155HG的过表达增强了α-SMA蛋白的荧光强度(图2E)。此外,MIR155HG干扰可降低胶原蛋白I和α-SMA并增加E-cadherin,而MIR155HG的过表达对这些蛋白产生相反的作用(图2F)。这些数据表明,MIR155HG干扰减弱,而MIR155HG过表达增强TGFβ1诱导的NHLF增殖和ECM沉积。
3、MIR155HG和miR-627对TGFβ1刺激的NHLF的动态影响
由于MIR155HG直接结合miR-627,因此用lv-MIR155HG并用miR-627mimics共转染,并检测miR-627表达以研究miR-627是否可以逆转MIR155HG的作用。如图3A所示,MIR155HG的过表达显著下调了miR-627的表达,而miR-627的过表达则明显逆转了MIR155HG的表达。然后,用lv-MIR155HG转染NHLF,并用miR-627mimics转染,用10ng/mL TGFβ1刺激48小时,并检测相关指标。在TGFβ1刺激下,MIR155HG的过表达增加,而miR-627的过表达减少HMGB1蛋白和p-65磷酸化。miR-627过表达可逆转MIR155HG过表达的作用(图3B)。MIR155HG过度表达作用得到强化,而miR-627则抑制DNA的合成能力;miR-627过表达可逆转MIR155HG过表达的作用(图3C)。MIR155HG的过表达增强,而miR-627的过表达减弱了代表α-SMA蛋白的荧光强度。miR-627过表达可逆转MIR155HG过表达的作用(图3D)。与此相一致地,MIR155HG的过表达增加了I型胶原和α-SMA的表达,并降低了E-cadherin的表达,而miR-627对这些蛋白的作用却相反。miR-627过表达可逆转MIR155HG过表达的作用(图3E)。这些数据表明,MIR155HG的过表达增强,而miR-627的过表达减弱了TGFβ1诱导的NHLF激活;miR-627过表达可逆转MIR155HG过表达对TGFβ1诱导的NHLF活化的影响。
4、MIR155HG/miR-627轴与NF-κB形成调节环
先前的研究表明NF-κB可以直接激活通过靶向MIR155HG近端启动子区。此外,在之前的研究中,证明了miR-627/HMGB1和RAGE/NF-κB信号通路可以形成一个调节环,以调节TGFβ1刺激的NHLF体外活化。接下来,发明人继续研究MIR155HG/miR-627/HMGB1轴是否可以与NF-κB形成调节环。用miR-627inhibitor转染NHLF,并与10ng/mLTGFβ1和/或JSH-23(NF-κB抑制剂)共同处理,测定p-65和p-p65蛋白以及MIR155HG的表达。在TGFβ1刺激下,miR-627抑制作用显著增加,而JSH-23处理则降低了p65的磷酸化和MIR155HG的表达。JSH-23治疗显著减弱了miR-627的作用抑制(图4A-B)。然后,用过表达NF-κB的慢病毒(lv-NF-κB)转染NHLF,以达到NF-κB的过表达。如RT-qPCR所证实,lv-NC被转染为阴性对照(图4C)。用lv-NF-κB在有或没有JSH-23处理的条件下转染NHLF,并检测其MIR155HG的表达。如图4D所示,NF-κB的过表达明显上调,而JSH-23处理下调了MIR155HG的表达;JSH-23处理可逆转NF-κB过表达的作用。这些数据表明NF-κB积极调节MIR155HG表达。
5、NF-κB/MIR155HG/miR-627/HMGB1的表达和相关性组织样本中的调节环
检测19例PF及正常组织中MIR155HG的表达。如图5A所示,与正常组织样品相比,PF组织样品中MIR155HG的表达显著上调。在组织样品中,MIR155HG与miR-627负相关,而MIR155HG与HMGB1正相关(图5B)。在PF组织样品中,与正常组织相比,HMGB1,p-p65和p65蛋白含量显著升高(图5C)。从H&E染色可知,在PF组织中观察到致密的纤维化病变分布不均,胶原蛋白和ECM过多沉积(图5D)。一致地,如通过IHC染色所揭示的那样,PF组织中HMGB1和NF-κB的蛋白质含量增加了(图5E)
显然,上述实施例仅仅是为清楚地说明所作的举例,而并非对实施方式的限定。对于所属领域的普通技术人员来说,在上述说明的基础上还可以做出其它不同形式的变化或变动。这里无需也无法对所有的实施方式予以穷举。而由此所引伸出的显而易见的变化或变动仍处于本发明创造的保护范围之中。
序列表
<110> 江西胸科医院
<120> 一种肺纤维化标志物及其应用
<160> 40
<170> SIPOSequenceListing 1.0
<210> 1
<211> 24
<212> DNA
<213> 人工序列
<400> 1
gcttgctgta ggctgtatgc tgtt 24
<210> 2
<211> 22
<212> DNA
<213> 人工序列
<400> 2
cttctttgtc atcctcccac gg 22
<210> 3
<211> 25
<212> DNA
<213> 人工序列
<400> 3
gagttattca aggggatgga gtaga 25
<210> 4
<211> 20
<212> DNA
<213> 人工序列
<400> 4
gcaaacctcc accagaaccg 20
<210> 5
<211> 21
<212> DNA
<213> 人工序列
<400> 5
gcctatgtcc caccactgtc g 21
<210> 6
<211> 25
<212> DNA
<213> 人工序列
<400> 6
ttttatctcc tttagcatcc ccttc 25
<210> 7
<211> 20
<212> DNA
<213> 人工序列
<400> 7
tatggcaaaa gcggacaagg 20
<210> 8
<211> 21
<212> DNA
<213> 人工序列
<400> 8
cttcgcaaca tcaccaatgg a 21
<210> 9
<211> 20
<212> DNA
<213> 人工序列
<400> 9
gaagcacgaa tgacagaggc 20
<210> 10
<211> 21
<212> DNA
<213> 人工序列
<400> 10
gcttggcgga ttagctcttt t 21
<210> 11
<211> 20
<212> DNA
<213> 人工序列
<400> 11
ttccagcctt ccttcctggg 20
<210> 12
<211> 20
<212> DNA
<213> 人工序列
<400> 12
ttgcgctcag gaggagcaat 20
<210> 13
<211> 18
<212> DNA
<213> 人工序列
<400> 13
gccgtctttt ctttgaga 18
<210> 14
<211> 16
<212> DNA
<213> 人工序列
<400> 14
cagtgcgtgt cgtgga 16
<210> 15
<211> 17
<212> DNA
<213> 人工序列
<400> 15
ctcgcttcgg cagcaca 17
<210> 16
<211> 20
<212> DNA
<213> 人工序列
<400> 16
aacgcttcac gaatttgcgt 20
<210> 17
<211> 59
<212> DNA
<213> 人工序列
<400> 17
gatccgcatt cacatggaac aaattgctcg agcaatttgt tccatgtgaa tgctttttg 59
<210> 18
<211> 59
<212> DNA
<213> 人工序列
<400> 18
aattcaaaaa gcattcacat ggaacaaatt gctcgagcaa tttgttccat gtgaatgcg 59
<210> 19
<211> 59
<212> DNA
<213> 人工序列
<400> 19
gatccggttc aagttcaata gcttagctcg agctaagcta ttgaacttga acctttttg 59
<210> 20
<211> 59
<212> DNA
<213> 人工序列
<400> 20
aattcaaaaa ggttcaagtt caatagctta gctcgagcta agctattgaa cttgaaccg 59
<210> 21
<211> 59
<212> DNA
<213> 人工序列
<400> 21
gatccgagca attgcacaaa cacttgctcg agcaagtgtt tgtgcaattg ctctttttg 59
<210> 22
<211> 59
<212> DNA
<213> 人工序列
<400> 22
aattcaaaaa gagcaattgc acaaacactt gctcgagcaa gtgtttgtgc aattgctcg 59
<210> 23
<211> 21
<212> RNA
<213> 人工序列
<400> 23
uucuccgaac gugucacguu u 21
<210> 24
<211> 21
<212> RNA
<213> 人工序列
<400> 24
acgugacacg uucggagaau u 21
<210> 25
<211> 20
<212> RNA
<213> 人工序列
<400> 25
ucuuuucuuu gagacucacu 20
<210> 26
<211> 20
<212> RNA
<213> 人工序列
<400> 26
ugagucucaa agaaaagauu 20
<210> 27
<211> 21
<212> RNA
<213> 人工序列
<400> 27
caguacuuuu guguaguaca a 21
<210> 28
<211> 20
<212> RNA
<213> 人工序列
<400> 28
agugagucuc aaagaaaaga 20
<210> 29
<211> 1500
<212> DNA
<213> 人工序列
<400> 29
gccgagcccg ggcccagcgc cgcctgcagc ctcgggaagg gagcggatag cggagccccg 60
agccgcccgc agagcaagcg cggggaacca aggagacgct cctggcactg cagataactt 120
gtctgcattt caagaacaac ctaccagaga ccttacctgt caccttggct ctcccaccca 180
atggagatgg ctctaatggt ggcacaaacc aggaagggga aatctgtggt ttaaattctt 240
tatgcctcat cctctgagtg ctgaaggctt gctgtaggct gtatgctgtt aatgctaatc 300
gtgatagggg tttttgcctc caactgactc ctacatatta gcattaacag tgtatgatgc 360
ctgttactag cattcacatg gaacaaattg ctgccgtggg aggatgacaa agaagcatga 420
gtcaccctgc tggataaact tagacttcag gctttatcat ttttcaatct gttaatcata 480
atctggtcac tgggatgttc aaccttaaac taagttttga aagtaaggtt atttaaaaga 540
tttatcagta gtatcctaaa tgcaaacatt ttcatttaaa tgtcaagccc atgtttgttt 600
ttatcattaa cagaaaatat attcatgtca ttcttaattg caggttttgg cttgttcatt 660
ataatgttca taaacacctt tgattcaact gttagaaatg tgggctaaac acaaatttct 720
ataatatttt tgtagttaaa aattagaagg actactaacc tccagttata tcatggattg 780
tctggcaacg ttttttaaaa gatttagaaa ctggtacttt cccccaggta acgattttct 840
gttcaggcaa cttcagttta aaattaatac ttttatttga ctcttaaagg gaaactgaaa 900
ggctatgaag ctgaattttt ttaatgaaat atttttaaca gttagcaggg taaataacat 960
ctgacagcta atgagatatt ttttccatac aagataaaaa gatttaatca aaaaatttca 1020
tatttgaaat gaagtcccaa atctaggttc aagttcaata gcttagccac ataatacggt 1080
tgtgcgagca gagaatctac ctttccactt ctaagcctgt ttcttcctcc atatggggat 1140
aatactttac aaggttgttg tgaggcttag atgagataga gaattattcc ataagataat 1200
caagtgctac attaatgtta tagttagatt aatccaagaa ctagtcaccc tactttatta 1260
gagaagagaa aagctaatga tttgatttgc agaatattta aggtttggat ttctatgcag 1320
tttttctaaa taaccatcac ttacaaatat gtaaccaaac gtaattgtta gtatatttaa 1380
tgtaaacttg ttttaacaac tcttctcaac attttgtcca ggttattcac tgtaaccaaa 1440
taaatctcat gagtctttag ttgatttaaa ataaaaaaaa aaaaaaaaaa aaaaaaaaaa 1500
<210> 30
<211> 4055
<212> DNA
<213> 人工序列
<400> 30
gaagtgcacc agcgagccgg ggcaggaaga ggaggtttcg ccaccggagc ggcccggcga 60
cgcgctgaca gcttcccctg cccttcccgt cggtcgggcc gccagccgcc gcagccctcg 120
gcctgcacgc agccaccggc cccgctcccg gagcccagcg ccgccgaggc cgcagccgcc 180
cggccagtaa ggcggcgccg ccgcccggcc accgcgcgcc ctgcgcttcc ctccgcccgc 240
gctgcggcca tggcgcggcg ctgactggcc tggcccggcc ccgccgcgct cccgctcgcc 300
ccgacccgca ctcgggcccg cccgggctcc ggcctgccgc cgcctcttcc ttctccagcc 360
ggcaggcccg cgccgcttag gagggagagc ccacccgcgc caggaggccg aacgcggact 420
cgccacccgg cttcagaatg gcagaagatg atccatattt gggaaggcct gaacaaatgt 480
ttcatttgga tccttctttg actcatacaa tatttaatcc agaagtattt caaccacaga 540
tggcactgcc aacagcagat ggcccatacc ttcaaatatt agagcaacct aaacagagag 600
gatttcgttt ccgttatgta tgtgaaggcc catcccatgg tggactacct ggtgcctcta 660
gtgaaaagaa caagaagtct taccctcagg tcaaaatctg caactatgtg ggaccagcaa 720
aggttattgt tcagttggtc acaaatggaa aaaatatcca cctgcatgcc cacagcctgg 780
tgggaaaaca ctgtgaggat gggatctgca ctgtaactgc tggacccaag gacatggtgg 840
tcggcttcgc aaacctgggt atacttcatg tgacaaagaa aaaagtattt gaaacactgg 900
aagcacgaat gacagaggcg tgtataaggg gctataatcc tggactcttg gtgcaccctg 960
accttgccta tttgcaagca gaaggtggag gggaccggca gctgggagat cgggaaaaag 1020
agctaatccg ccaagcagct ctgcagcaga ccaaggagat ggacctcagc gtggtgcggc 1080
tcatgtttac agcttttctt ccggatagca ctggcagctt cacaaggcgc ctggaacccg 1140
tggtatcaga cgccatctat gacagtaaag cccccaatgc atccaacttg aaaattgtaa 1200
gaatggacag gacagctgga tgtgtgactg gaggggagga aatttatctt ctttgtgaca 1260
aagttcagaa agatgacatc cagattcgat tttatgaaga ggaagaaaat ggtggagtct 1320
gggaaggatt tggagatttt tcccccacag atgttcatag acaatttgcc attgtcttca 1380
aaactccaaa gtataaagat attaatatta caaaaccagc ctctgtgttt gtccagcttc 1440
ggaggaaatc tgacttggaa actagtgaac caaaaccttt cctctactat cctgaaatca 1500
aagataaaga agaagtgcag aggaaacgtc agaagctcat gcccaatttt tcggatagtt 1560
tcggcggtgg tagtggtgct ggagctggag gcggaggcat gtttggtagt ggcggtggag 1620
gagggggcac tggaagtaca ggtccagggt atagcttccc acactatgga tttcctactt 1680
atggtgggat tactttccat cctggaacta ctaaatctaa tgctgggatg aagcatggaa 1740
ccatggacac tgaatctaaa aaggaccctg aaggttgtga caaaagtgat gacaaaaaca 1800
ctgtaaacct ctttgggaaa gttattgaaa ccacagagca agatcaggag cccagcgagg 1860
ccaccgttgg gaatggtgag gtcactctaa cgtatgcaac aggaacaaaa gaagagagtg 1920
ctggagttca ggataacctc tttctagaga aggctatgca gcttgcaaag aggcatgcca 1980
atgccctttt cgactacgcg gtgacaggag acgtgaagat gctgctggcc gtccagcgcc 2040
atctcactgc tgtgcaggat gagaatgggg acagtgtctt acacttagca atcatccacc 2100
ttcattctca acttgtgagg gatctactag aagtcacatc tggtttgatt tctgatgaca 2160
ttatcaacat gagaaatgat ctgtaccaga cgcccttgca cttggcagtg atcactaagc 2220
aggaagatgt ggtggaggat ttgctgaggg ctggggccga cctgagcctt ctggaccgct 2280
tgggtaactc tgttttgcac ctagctgcca aagaaggaca tgataaagtt ctcagtatct 2340
tactcaagca caaaaaggca gcactacttc ttgaccaccc caacggggac ggtctgaatg 2400
ccattcatct agccatgatg agcaatagcc tgccatgttt gctgctgctg gtggccgctg 2460
gggctgacgt caatgctcag gagcagaagt ccgggcgcac agcactgcac ctggctgtgg 2520
agcacgacaa catctcattg gcaggctgcc tgctcctgga gggtgatgcc catgtggaca 2580
gtactaccta cgatggaacc acacccctgc atatagcagc tgggagaggg tccaccaggc 2640
tggcagctct tctcaaagca gcaggagcag atcccctggt ggagaacttt gagcctctct 2700
atgacctgga tgactcttgg gaaaatgcag gagaggatga aggagttgtg cctggaacca 2760
cgcctctaga tatggccacc agctggcagg tatttgacat attaaatggg aaaccatatg 2820
agccagagtt tacatctgat gatttactag cacaaggaga catgaaacag ctggctgaag 2880
atgtgaagct gcagctgtat aagttactag aaattcctga tccagacaaa aactgggcta 2940
ctctggcgca gaaattaggt ctggggatac ttaataatgc cttccggctg agtcctgctc 3000
cttccaaaac acttatggac aactatgagg tctctggggg tacagtcaga gagctggtgg 3060
aggccctgag acaaatgggc tacaccgaag caattgaagt gatccaggca gcctccagcc 3120
cagtgaagac cacctctcag gcccactcgc tgcctctctc gcctgcctcc acaaggcagc 3180
aaatagacga gctccgagac agtgacagtg tctgcgacag cggcgtggag acatccttcc 3240
gcaaactcag ctttaccgag tctctgacca gtggtgcctc actgctaact ctcaacaaaa 3300
tgccccatga ttatgggcag gaaggacctc tagaaggcaa aatttagcct gctgacaatt 3360
tcccacaccg tgtaaaccaa agccctaaaa ttccactgcg ttgtccacaa gacagaagct 3420
gaagtgcatc caaaggtgct cagagagccg gcccgcctga atcattctcg atttaactcg 3480
agaccttttc aacttggctt cctttcttgg ttcataaatg aattttagtt tggttcactt 3540
acagatagta tctagcaatc acaacactgg ctgagcggat gcatctgggg atgaggttgc 3600
ttactaagct ttgccagctg ctgctggatc acagctgctt tctgttgtca ttgctgttgt 3660
ccctctgcta cgttcctatt gtcattaaag gtatcacggt cgccacctgg cattccttct 3720
gaccacagca tcattttgca ttcaaattaa gggttaagaa aagagatatt ttaaaatgag 3780
agtcacttga tgtgccattt taaaaaaaaa ggcatattgc tttttctaat gtggttattt 3840
ctctgatttg caaaaaaaaa aaaaaaaaaa atacttgtca atatttaaac atggttacaa 3900
tcattgctga aaatggtatt ttcccccttt tctgcatttt gctattgtaa atatgttttt 3960
tagatcaaat actttaaagg aaaaaatgtt ggatttataa atgctatttt ttattttact 4020
tttataataa aaggaaaagc aaattgatga cctca 4055
<210> 31
<211> 42
<212> DNA
<213> 人工序列
<400> 31
gagaagatgg ccgctaatga tttgatttgc agaatattta ag 42
<210> 32
<211> 43
<212> DNA
<213> 人工序列
<400> 32
ttagcggcca tcttctctaa taaagtaggg tgactagttc ttg 43
<210> 33
<211> 44
<212> DNA
<213> 人工序列
<400> 33
aattctaggc gatcgctcga gagcctgttt cttcctccat atgg 44
<210> 34
<211> 51
<212> DNA
<213> 人工序列
<400> 34
attttattgc ggccagcggc cgctgttgag aagagttgtt aaaacaagtt t 51
<210> 35
<211> 59
<212> DNA
<213> 人工序列
<400> 35
gatccgtgtt agttgcaaag atatgcctcg aggcatatct ttgcaactaa cactttttg 59
<210> 36
<211> 59
<212> DNA
<213> 人工序列
<400> 36
aattcaaaaa gtgttagttg caaagatatg cctcgaggca tatctttgca actaacacg 59
<210> 37
<211> 59
<212> DNA
<213> 人工序列
<400> 37
gatccggtta ctcactataa acaatactcg agtattgttt atagtgagta acctttttg 59
<210> 38
<211> 59
<212> DNA
<213> 人工序列
<400> 38
aattcaaaaa ggttactcac tataaacaat actcgagtat tgtttatagt gagtaaccg 59
<210> 39
<211> 59
<212> DNA
<213> 人工序列
<400> 39
gatccgttac tcactataaa caatatctcg agatattgtt tatagtgagt aactttttg 59
<210> 40
<211> 59
<212> DNA
<213> 人工序列
<400> 40
aattcaaaaa gttactcact ataaacaata tctcgagata ttgtttatag tgagtaacg 59
- 一种肺纤维化早期诊断标志物在制备肺纤维化诊断试剂中的应用
- 一种与肺纤维化辅助诊断相关的外周血circRNA标志物及其应用