一种α-1,3-葡萄糖苷酶在黑曲霉低聚糖制备中的应用
文献发布时间:2023-06-19 11:40:48
技术领域
本发明涉及一种α-1,3-葡萄糖苷酶在黑曲霉低聚糖制备中的应用,属于基因工程和发酵工程技术领域。
背景技术
黑曲霉低聚糖(Nigerooligosaccharides,NOS)是一类含有α-1,3键的α-D-葡萄糖寡聚物,由于这种糖最早在日本的发酵清酒中被发现有少量存在,又称Sakebiose,其成分主要包括黑曲霉二糖、黑曲霉三糖、黑曲霉糖基葡萄糖和黑曲霉糖基麦芽糖等。与其他低聚糖相比,黑曲霉低聚糖具有特殊醇香风味、低黏度、低甜度(约是蔗糖的45%)等特点,是清酒、豆酱等发酵型产品具有特殊的良好口感的主要原因。它可代替砂糖加入各种饮料中以增加厚味,降低高甜度甜味剂的异味、苦味,同时具有协调咸味和在低盐下保持与增强鲜味的效果,对以鲜味为主体的各种饮料食品有味质调节效果。此外,黑曲霉低聚糖还具备预防龋齿、促进双歧杆菌增殖调节肠道健康、改善脂质代谢等生理功效,以及使T淋巴球和B淋巴球等免疫细胞活性化的免疫激活作用。
黑曲霉低聚糖的生产方法主要有化学合成、多糖水解法和酶催化转糖苷法3种。α-葡萄糖苷酶催化的葡萄糖基转移反应是食品工业生产低聚异麦芽糖和黑曲霉低聚糖的重要反应,GH31家族α-葡萄糖苷酶是具有高转苷活性的一类α-葡萄糖苷酶,但目前该家族α-葡萄糖苷酶的应用研究主要集中于催化合成α-1,6糖苷键制备低聚异麦芽糖方面,对α-1,3糖苷键具有高度选择性的α-葡萄糖苷酶报道较少。目前已报道的具有α-1,3糖苷键合成能力的α-葡萄糖苷酶主要来源于真菌,如Acremoniun sp.S4G13分泌转化酶以麦芽糖为底物催化制备黑曲霉低聚糖的转化率可达37%,但其糖浆产品整体浓度仅20g/L(NOS约7.4g/L),生产效率较为低下,且合成产物主要为黑曲霉二糖和黑曲霉糖基麦芽糖,而黑曲霉三糖、黑曲霉糖基葡萄糖以及DP≥4的黑曲霉糖基麦芽低聚糖则含量较低。本发明的目的在于利用一种细菌来源的α-1,3-葡萄糖苷酶提高黑曲霉低聚糖的产量及转化率,同时提升所制备黑曲霉低聚糖组分中黑曲霉三糖、黑曲霉糖基葡萄糖的含量比例。
发明内容
α-葡萄糖苷酶(EC3.2.1.20)可催化麦芽低聚糖、异麦芽糖、α-葡聚糖等底物的非还原末端释放α-D-葡萄糖,或通过转糖基作用将葡萄糖基从供体底物转移至糖基受体生成低聚糖或糖苷衍生物。
本发明提供了一种重组大肠杆菌,表达Thermoplasmaacidophilum来源的α-1,3-葡萄糖苷酶,所述α-1,3-葡萄糖苷酶的氨基酸序列如SEQ ID NO.1所示。
在一种实施方式中,以pET24a为表达载体。
在一种实施方式中,以大肠杆菌BL21(DE3)为宿主。
本发明提供了一种制备黑曲霉低聚糖的方法,所述方法是利用氨基酸序列如SEQIDNO.1所示的α-1,3-葡萄糖苷酶,或所述重组大肠杆菌表达的α-1,3-葡萄糖苷酶,以麦芽糖、葡萄糖或麦芽糊精中的一种或几种为底物,合成黑曲霉低聚糖。
在一种实施方式中,以麦芽糖和葡萄糖的混合物为底物,麦芽糖的浓度为100-200g/L,葡萄糖的浓度为100-200g/L。
在一种实施方式中,麦芽糖和葡萄糖的浓度均为100g/L,或葡萄糖的浓度为100g/L、麦芽糖的浓度为200g/L。
在一种实施方式中,α-1,3-葡萄糖苷酶的添加量为0.5-2U/mL。
在一种实施方式中,α-1,3-葡萄糖苷酶的添加量不低于1U/mL。
在一种实施方式中,在pH5-7、70-90℃下反应。
在一种实施方式中,反应时间为1-6h。
在一种实施方式中,反应时间为3-6h。
本发明提供了所述重组大肠杆菌在制备黑曲霉低聚糖中的应用。
在一种实施方式中,所述黑曲霉低聚糖包括黑曲霉二糖、黑曲霉三糖、黑曲霉糖基葡萄糖和黑曲霉糖基麦芽糖。
本发明的有益效果:本发明将来源于Thermoplasmaacidophilumα-1,3-葡萄糖苷酶异源表达到大肠杆菌BL21(DE3)中,得到一种高产α-1,3-葡萄糖苷酶基因工程菌。利用本发明的α-1,3-葡萄糖苷酶TaAglA以10%葡萄糖和20%麦芽糖为底物,α-1,3-葡萄糖苷酶加酶量为1U/mL,在pH 6、温度80℃下进行酶反应,反应3h,黑曲霉低聚糖产量最高能达到88.5g/L,转化率为29.5%。以10%麦芽糖和10%麦芽糊精为底物,α-1,3-葡萄糖苷酶加酶量为1U/mL,在pH 6、温度80℃下进行酶反应,反应6h,黑曲霉低聚糖产量最高能达到47.4g/L,转化率为23.7%。本专利所获得的黑曲霉低聚糖(NOS)产量远高于目前已有报道的Acremoniun sp.S4G1 3α-葡萄糖苷酶制备产量7.4g/L。同时,本专利提出以10%麦芽糖和10%麦芽糊精为底物,同样获得了较高的黑曲霉低聚糖(NOS)转化率和产量,有利于拓宽黑曲霉低聚糖的底物范围和降低生产成本。
附图说明
图1为α-1,3-葡萄糖苷酶TaAglA重组基因工程菌发酵酶液SDS-PAGE电泳图;
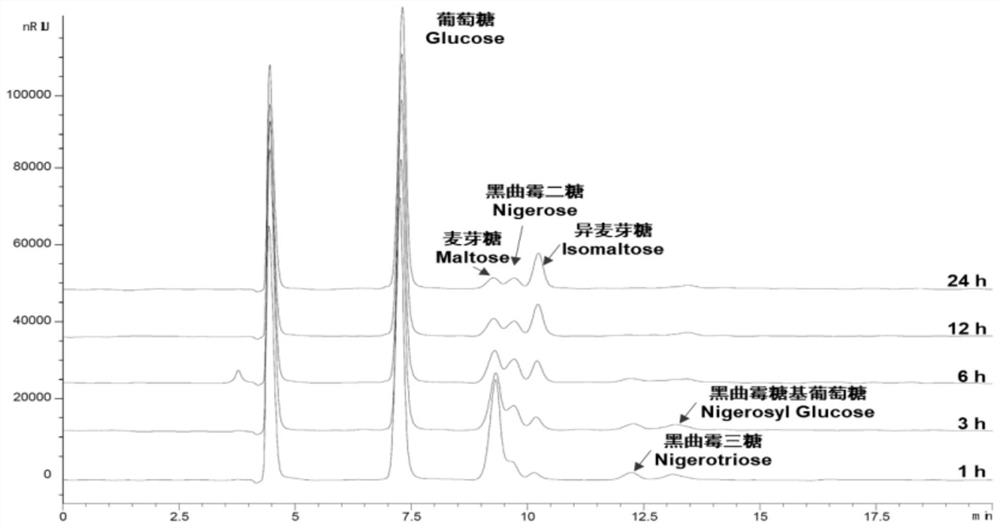
图2为反应时间对α-1,3-葡萄糖苷酶TaAglA制备黑曲霉低聚糖转化率的影响图;
图3为加酶量对α-1,3-葡萄糖苷酶TaAglA制备黑曲霉低聚糖转化率的影响图;
图4为以不同底物进行反应黑曲霉低聚糖产物组分的质量浓度图。
具体实施方式
(一)培养基:
(1)LB培养基:酵母粉5.0g/L,蛋白胨10g/L,NaCl10g/L;固体培养基添加20g/L琼脂。
(2)TB培养基:蛋白胨12g/L,酵母粉24g/L,甘油5g/L,K
(二)α-1,3-葡萄糖苷酶活力分析:
(1)酶活单位定义
每分钟水解pNP-α-G产生1μmol的对硝基苯酚的酶活力为一个酶活单位。
(2)酶活力测定步骤
反应体系为1.0mL,pH6的醋酸缓冲液960μL,加入适度稀释的粗酶液20μL,再加入20μL 100mmol/L的pNP-α-G,在90℃eppendorfThermoMixer恒温混匀仪中预热5~10min,用秒表精确计时,10min后立即加入200μL的1mol/L的Na
实施例1:基因工程菌E.coli BL21(DE3)/pET24a-TaAglA的构建
将合成的α-1,3-葡萄糖苷酶基因TaAglA(核苷酸序列如SEQ ID NO.2所示)通过Nde I酶和EcoRI酶切位点,插入表达载体pET24a获得重组质粒pET24a-TaAglA,电击转化至大肠杆菌E.coli BL21(DE3)中。涂布LB四环素固体培养基,挑单菌落接种LB液体培养基培养10h,酶切验证正确后甘油保菌,获得基因工程菌E.coli BL21(DE3)/pET24a-TaAglA。
将上述基因工程菌在LB培养基中进行摇瓶诱导发酵,诱导产酶条件为30℃,0.4mMIPTG,诱导表达时间24h。收集菌体,用磷酸盐缓冲液重悬后超声破碎细胞,所得细胞破碎上清液即为粗酶液。粗酶液采用Ni2+亲和层析柱纯化,洗脱所得纯化酶液的SDS-PAGE凝胶电泳分析如图1所示。
分别测定α-1,3-葡萄糖苷酶TaAglA的最适pH和最适温度(通过调节酶活测定时醋酸缓冲液的pH来找到最适pH,通过调节在eppendorfThermoMixer恒温混匀仪中的温度来找到最适温度),本发明菌株生产的α-1,3-葡萄糖苷酶TaAglA的最适pH为6、最适温度为80℃,重组酶摇瓶表达水平达0.2g/L。在最适反应pH条件下,α-1,3-葡萄糖苷酶TaAglA在50-90℃反应温度下可保持60%以上酶活;在最适反应温度条件下,α-1,3-葡萄糖苷酶TaAglA在pH4.0-9.0下可保持50%以上酶活。
实施例2:反应时间对α-1,3-葡萄糖苷酶TaAglA制备黑曲霉低聚糖(NOS)的影响
以20%(200g/L)麦芽糖为底物,添加1U/mLα-1,3-葡萄糖苷酶TaAglA,在pH 6.0柠檬酸-磷酸盐缓冲液、80℃反应条件下反应0-24h,分别设置时间1h、3h、6h、12h、24h。
采用高效液相色谱分析酶转化产物组分,液相检测方法如下:使用Agilent1260Infinity II液相色谱系统的RID示差检测器检测酶反应产生的葡萄糖、麦芽糖、曲二糖、黑曲霉二糖、黑曲霉三糖、黑曲霉糖基葡萄糖为标准品,确定产物的峰面积和出峰时间。所用色谱柱型号为HYPERSIL APS2(250×4.6mm,5μm),参比池温度和柱温度设定为35℃,流动相为78%(v·v
检测结果如图2所示,可见α-1,3-葡萄糖苷酶TaAglA转苷合成的α-1,3糖苷键黑曲霉低聚糖(NOS)主要为黑曲霉二糖、黑曲霉三糖、黑曲霉糖基葡萄糖,且在酶反应3h时NOS产量达到最高,继续进行反应,NOS组分逐渐被水解,生成葡萄糖和异麦芽糖。因此最佳反应时间位3h。
实施例3:α-1,3-葡萄糖苷酶TaAglA加酶量对黑曲霉低聚糖(NOS)制备转化率的影响
以20%(200g/L)麦芽糖为底物,在pH 6、80℃反应条件,设置不同加酶量0.25U/mL、0.5U/mL、1U/mL、1.5U/mL、2U/mL,探究以20%麦芽糖为底物利用α-1,3-葡萄糖苷酶TaAglA转苷活性合成黑曲霉低聚糖(NOS)加酶量对转化率的影响。结果如图3所示,在一定范围内,底物的转化率随着加酶量的增加不断提高,当加酶量达到1U/mL时,黑曲霉低聚糖(NOS)转化率可达到17.8%;进一步增加酶量,转化率几乎保持不变。综合考虑,选择1U/mL的加酶量,此时底物转化率可达到17.8%,低聚龙胆糖的产量能达到35.6g/L。
实施例4:α-1,3-葡萄糖苷酶TaAglA以不同底物进行反应制备黑曲霉低聚糖(NOS)
在pH 6,80℃,加酶量为1U/mL的反应条件下,以不同底物组分进行酶转化反应,通过改变底物的组成和浓度来找到适宜的底物组分。结果如图4所示,以10%(100g/L)葡萄糖和20%(200g/L)麦芽糖(10%G+20%M)为底物时,黑曲霉低聚糖(NOS)转化率最高,可达到29.5%,产量达88.5g/L。而以10%麦芽糖和20%麦芽糊精为底物时,转化率最高为23.7%,产量达47.4g/L。
虽然本发明已以较佳实施例公开如上,但其并非用以限定本发明,任何熟悉此技术的人,在不脱离本发明的精神和范围内,都可做各种的改动和修饰,因此本发明的保护范围应该以权利要求书所界定的为准。
SEQUENCE LISTING
<110> 江南大学
<120> 一种α-1,3-葡萄糖苷酶在黑曲霉低聚糖制备中的应用
<130> BAA210281A
<160> 2
<170> PatentIn version 3.3
<210> 1
<211> 749
<212> PRT
<213> Thermoplasma acidophilum
<400> 1
Met Leu Asp Asp Ile Arg Gly Phe Met Asn Thr Phe Ser Glu Thr Met
1 5 10 15
Phe Met Asp Val Ile Asn Tyr Ser Leu Thr Arg Asp Arg Tyr Asp Ser
20 25 30
Val Phe Leu Gln Arg Gln Asn Tyr Arg Asp Leu Gly Gln Leu Lys Glu
35 40 45
Ile Leu Thr Asn Asp Asp Gly Ile Lys Ile Lys Phe Asp Gly Tyr Asp
50 55 60
Leu Lys Ile Gln Gly Thr Ala Asp Gly Thr Ile Lys Phe Thr Trp Thr
65 70 75 80
Asp Gly Pro Glu Ile Pro Val Thr Asp Pro Glu Phe Val Lys Pro Arg
85 90 95
Leu Asp Asn Gly Val Tyr Glu Phe Gly Asp Tyr Arg Leu Leu Ile Glu
100 105 110
Lys Asp Gly Phe Ser Ile Arg Asp Val Glu Gly Thr Ile Leu His Arg
115 120 125
Glu Phe Phe Pro Ser Phe Gly Glu Asn Ile Thr His Ala Phe Glu Leu
130 135 140
Asn Gln Gly Asp Ile Ile Ala Gly Leu Gly Glu Lys Ala Ala Pro Ile
145 150 155 160
Asn Met Ile Gly His Val Phe Arg Leu Trp Asn His Asp Ala Asn Gly
165 170 175
Ser Tyr Gly Pro Asp Ser Asp Pro Leu Tyr Val Asn Val Pro Ile Met
180 185 190
Leu His Gly His Ala Gly Arg Phe Ile Leu Ile Leu Tyr Val Asn Ala
195 200 205
Gly Asp Ala Thr Val Asp Val Gly Tyr Ser Asp Glu His Arg Val Ser
210 215 220
Ser Ser Phe Lys Ser Lys Pro Leu Ala Tyr Tyr Val Ile Thr Gly Asn
225 230 235 240
Leu Asp Thr Ile Tyr Glu Lys Leu Ser Leu Ile Thr Gly Lys Pro Gln
245 250 255
Lys Pro Pro Tyr Trp Ala Phe Glu Phe Gln Gln Ser Arg Tyr Ser Tyr
260 265 270
Met Asp Thr Lys Glu Val Arg Asp Leu Val Asp Gly Phe Ala Ser Arg
275 280 285
Gly Ile Pro Leu Gly Ala Val Tyr Leu Asp Ile Asp Tyr Met Asp Arg
290 295 300
Phe Lys Met Phe Thr Phe Asp Pro Gln Arg Phe Gly Asp Val Lys Gln
305 310 315 320
Leu Thr Glu Tyr Met Glu Gln Lys Gly Val Lys Leu Ile Thr Ile Met
325 330 335
Glu Pro Ser Ile Lys Met Glu His Gly Phe Asp Leu Tyr Glu Glu Gly
340 345 350
Leu Lys Gly Gly Tyr Phe Val Lys Tyr Pro Asp Gly Asn Val Met Tyr
355 360 365
Ala Pro Val Trp Pro Glu Met Ala Ala Phe Pro Asp Phe Thr Asp Glu
370 375 380
Lys Ala Arg Glu Trp Tyr Ala Ser Lys Tyr Asp Phe Met Arg Ser Met
385 390 395 400
Gly Val Ser Gly Phe Trp His Asp Met Asn Glu Pro Ala Ile Phe Val
405 410 415
Gly Trp Gly Asp Asn Thr Met Pro Arg Ser Ala Val His Arg Ile Gly
420 425 430
Arg His Glu Glu Val His Asn Leu Tyr Gly Tyr Tyr Met Asp Lys Ala
435 440 445
Ala Tyr Asp His Leu Ser Lys Val Glu Arg Pro Phe Ile Leu Ser Arg
450 455 460
Ser Gly Trp Ala Gly Ile Ser Arg Tyr Gly Trp Ile Trp Thr Gly Asp
465 470 475 480
Thr Glu Thr Ser Trp Lys Glu Leu Lys Gln Asn Ile Ile Thr Ile Met
485 490 495
His Met Ser Met Ser Gly Ile Thr Leu Thr Gly Cys Asp Ile Gly Gly
500 505 510
Phe Thr Gly Ser Pro Thr Pro Glu Leu Phe Ile Arg Trp Leu Gln Ala
515 520 525
Ser Leu Phe Phe Pro Leu Tyr Arg Val His Ser Asp Lys Lys Ser Lys
530 535 540
Arg Arg Glu Pro Trp Ala Phe Gly Ser His Glu Lys Glu Ile Ile Glu
545 550 555 560
Ile Ile Arg Leu Arg His Ser Phe Val Pro His Ile Tyr Ser Glu Ala
565 570 575
Ile Ser Ser Ser Ile Thr Gly Leu Pro Leu Val Arg Pro Val Phe Trp
580 585 590
Ala Asp Pro Ser Arg Asn Asp Leu Met Ser Val Asp Asp Glu Tyr Thr
595 600 605
Phe Gly Gly Ser Ile Leu Ile Ala Pro Ile Val Glu Glu His Ala Val
610 615 620
Ile Arg Arg Ile Ile Leu Pro Ser Gly Arg Trp Tyr Asn Ile Ser Asp
625 630 635 640
Asp Arg Ile Val Glu Gly Ser Leu Ser Ile Asn Val Asp Leu Ser Thr
645 650 655
Val Pro Ile Phe Val Arg Glu Gly Ser Ala Ile Leu Arg Glu Asn Asp
660 665 670
Gly Ile Glu Leu His Leu Tyr Leu Ala Asn Glu Arg Arg Lys Ser Ile
675 680 685
Leu Tyr Ile Ser Asp Gly Asn Asp Asp Ile Lys Val Glu Val Glu Phe
690 695 700
Asp Ser Lys Ser Val Asp Ile Asn Pro Gln Lys Val Pro Lys Ile Lys
705 710 715 720
Ala Ile Val Met His Gly Phe Arg Glu Asp Glu Leu Ile Leu Asn Gly
725 730 735
Glu Pro Ile Lys Thr Ser Asp Gly Ile Ile Arg Leu Lys
740 745
<210> 2
<211> 2268
<212> DNA
<213> 人工序列
<400> 2
atgttggacg acattcgcgg gtttatgaat acattttcag agaccatgtt catggatgtt 60
attaactatt cattaacgcg cgaccgctac gactcggttt tcttacagcg ccaaaactat 120
cgtgacctgg gacagttgaa agaaatcctg accaacgacg acggaattaa aattaaattc 180
gatgggtacg atctgaagat ccaaggaaca gccgatggga caatcaagtt tacttggact 240
gatggtcccg aaatccctgt cactgacccg gagttcgtga aaccgcgtct ggacaatgga 300
gtgtatgagt tcggggatta tcgtctgctg attgagaaag acggattctc cattcgtgac 360
gtagaaggga cgattttgca ccgcgagttt tttccttcgt tcggagagaa tatcacgcat 420
gcgttcgaac ttaatcaggg tgacatcatt gctgggctgg gggagaaggc agcgcccatt 480
aacatgattg gccacgtatt tcgtctttgg aaccacgacg cgaatggctc ttacggaccc 540
gattcggacc cgttatacgt taacgtacca attatgctgc atggccatgc aggccgcttc 600
atcttgatcc tttatgtaaa tgctggtgac gcgactgtcg atgtaggcta ttcggacgaa 660
caccgcgtat catcaagttt caagagcaaa ccattggcat actatgtcat cacaggaaac 720
cttgatacaa tttatgagaa actttcactt attacgggta aacctcaaaa gcctccttac 780
tgggcgttcg agtttcaaca atcccgctat tcttatatgg atacgaaaga agtccgtgac 840
ttggtagatg gttttgcgtc gcgtggtatc ccgttaggtg ctgtttacct tgacattgac 900
tacatggacc gctttaagat gttcacattt gatccgcagc gttttggcga cgttaagcaa 960
ttgacggagt acatggaaca aaagggcgtc aaattaatca cgatcatgga accgagcatc 1020
aagatggagc atggttttga cctttacgaa gagggtttaa aaggcggtta ttttgtaaag 1080
taccccgacg gcaatgtaat gtatgcacct gtatggcctg aaatggcggc attcccggat 1140
tttaccgatg agaaggctcg tgaatggtac gcatctaaat atgacttcat gcgtagcatg 1200
ggagtaagcg gcttttggca tgatatgaat gagccagcga tctttgtagg atggggtgac 1260
aacaccatgc cgcgtagcgc ggtacatcgt atcggccgcc acgaggaagt gcacaacctg 1320
tacggatact atatggacaa ggcagcgtac gaccacctgt cgaaagttga acgtcctttt 1380
attctgtctc gctcgggatg ggcgggcatc tcccgttacg gttggatttg gacaggtgac 1440
acggagacat cttggaaaga gttaaagcag aacattatca ccatcatgca catgtccatg 1500
tctggaatta cccttaccgg ctgtgatatc ggaggtttca cggggagtcc taccccggaa 1560
ttgttcattc gttggttgca ggcatcatta ttttttcccc tgtatcgtgt gcactctgac 1620
aaaaaaagca agcgtcgcga gccttgggcc tttgggtcac acgagaagga aatcatcgaa 1680
attatccgtt tgcgtcactc attcgtgccg catatctatt cagaagcaat cagttcgtcg 1740
attactggtc tgccgttagt acgccctgtt ttctgggctg accctagtcg taacgatctt 1800
atgagcgtgg atgacgaata cacattcggc ggcagtattt taatcgcacc gattgtcgaa 1860
gagcatgcgg taattcgtcg tatcatcttg ccctcaggtc gctggtataa tatttcggac 1920
gaccgtatcg ttgaagggtc gttaagtatc aacgtggacc ttagcactgt acctatcttc 1980
gttcgcgagg ggtcggcaat tcttcgtgag aacgacggca tcgagctgca cctttactta 2040
gcaaacgagc gtcgcaagag tattttatat atttcggacg gtaacgatga catcaaagtc 2100
gaggttgagt tcgactcaaa atcggttgac atcaacccac agaaagtacc taagatcaaa 2160
gcgatcgtca tgcacgggtt tcgtgaagat gaacttatcc tgaatggtga gccgattaag 2220
acttcagatg gtattattcg tttaaagcat catcaccatc accactaa 2268
- 一种α-1,3-葡萄糖苷酶在黑曲霉低聚糖制备中的应用
- 一种利用固定化α-葡萄糖苷酶制备黑曲霉低聚糖的方法