一种高效异源合成罗汉果甜苷V前体的方法
文献发布时间:2023-06-19 12:08:44
技术领域
本发明涉及一种高效异源合成罗汉果甜苷V前体的方法,属于生物工程领域。
背景技术
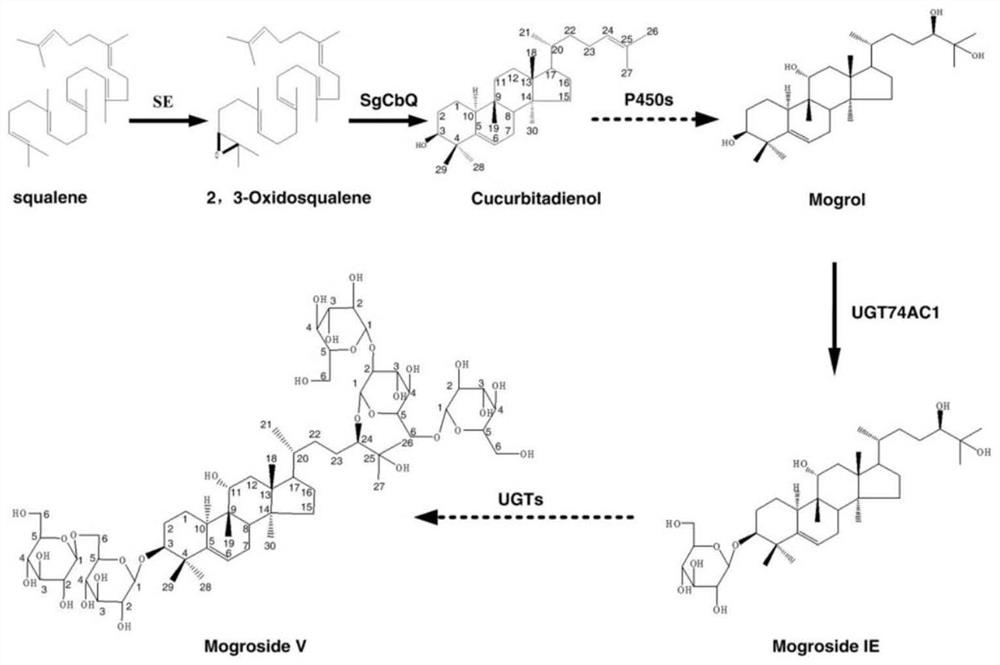
高糖饮食,不管是天然还是人工合成的糖类,都会造成一系列现代健康问题,如肥胖、糖尿病、心血管疾病等(Sievenpiper,2014)。因此,植物来源的天然非糖物质因其高甜度、低热量、安全度高等优势,成为满足甜味需求的新一代甜味剂。其中罗汉果的主要活性物质罗汉果甜苷V甜度是蔗糖的250-300倍,热量低,且具有止咳祛痰、降血糖、抗癌、抗氧化等药理活性(Kasai et al.,1989)。因此,罗汉果甜苷V可代替蔗糖作为甜味剂广泛应用于各种食品,作为肥胖症或糖尿病患者的代糖。但罗汉果生长周期长,栽培存在诸多困难;且果实中罗汉果甜苷V含量低;传统获取方法已经难以满足日益增长的需求(Makapugay etal.,1985)。而近年来快速发展的组学促进了关键合成通路的解析与关键功能基因的发掘。代谢工程和合成生物学的快速发展为植物天然产物的异源生物合成提供了可持续生产思路。2018年,Iktin等人根据罗汉果转录组和基因组数据,解析了罗汉果甜苷V合成通路,为利用合成生物学构建细胞工程、实现体外合成奠定了基础(Itkin,2018)。
科学家们对罗汉果甜苷V的异源生物合成开展了一些初步探究,主要表现在以下几方面。一,底盘菌的选择和优化。合适的底盘细胞是天然产物高效生产的基础。例如酿酒酵母的MVA途径能够提供罗汉果甜苷异源合成的前体2,3-环氧角鲨烯,但酵母中大部分2,3-环氧角鲨烯会被羊毛甾醇合酶(ERG7)分流进入麦角固醇通路。科学家们通过ERG7抑制剂R048-8072处理或利用缺乏ERG7的酿酒酵母菌株(GIL77)作为底盘菌,增加了2,3-环氧角鲨烯到罗汉果甜苷V的代谢流(Dai et al.,2015)。二,葫芦二烯醇的生物合成。李守连等在酵母中过表达三萜化合物合成关键酶以及氧化鲨烯环化酶SgCDS,并进一步改善不同基因表达比例,葫芦二烯醇产量可达1724.10mg/L。虽然葫芦二烯醇不是罗汉果甜苷合成的骨架物,但可利用体外P450氧化酶、糖基转移酶的催化,生成罗汉果甜苷V(罗祖良et al.,2016)。三,体外糖基化修饰。天津工业生物所孙媛霞课题组鉴定到了糖基转移酶UGT74AC1,从大肠杆菌中纯化的UGT74AC1可以在体外以甜甙的共同前体物质(罗汉果醇)为底物,对其C3-OH进行糖基化生成甜甙IE(Dai et al.,2015)。
科学家们已经探索了一条罗汉果甜苷的异源生物合成途径(如图1)。我们发现目前获得罗汉果甜苷V前体主要依赖代谢工程的方法,将途径中所需关键酶进行异源组装,并没有对这些酶在原核系统中进行适应性改造,这将导致关键酶的酶活和稳定性受到一定程度的影响,不能发挥最大功能。因此,关键酶的适应性改造对于天然产物的高效生产至关重要。另一方面,我们可以发现现有合成方法依赖于原有途径,基本上是在原有途径的基础上逐个实现每一步反应,虽然有可能实现异源生物合成,但罗汉果甜苷V合成路径复杂,这将是一个漫长的过程。为了高效快速合成罗汉果甜苷V,需要缩短路径并对关键酶进行功能改良。11H-Cuol是葫芦二烯醇到罗汉果醇的合成过程的中间化合物,是罗汉果甜苷V的前体物质(如图2)。11H-Cuol合成关键酶属于细胞色素P450s,催化效率低且在异源宿主中的表达量很低,这些因素限制了罗汉果甜苷V前体的生产效率。现有技术也尚未对该酶进行适应性改造,导致其酶活和稳定性受到一定程度影响,进一步影响了罗汉果甜苷V的生产效率。因此,本项目将专注于实现葫芦二烯醇到罗汉果醇的合成路径中关键酶细胞色素P450s的功能改良。
细胞色素P450s是植物最大的酶蛋白家族之一,能够催化合成大量结构特异的代谢物。这些代谢物不仅在植物的生长发育、胁迫反应等过程中发挥作用,同时它们也可作为珍贵原料合成新的药物(Alonso-Gutierrez et al.,2013)。因此,P450s通常被选作代谢工程的靶标,利用蛋白工程改变P450s的酶活性、蛋白稳定性等因素提高相关代谢物的合成效率。近年来,利用基因组挖掘以及现代生物学技术,科学家成功鉴定并阐明了不同代谢途径中P450s的催化机制。P450s催化植物特异的代谢反应,催化效率低(kcat<5s
发明内容
为了解决上述发明问题,本发明提供一种高效异源合成罗汉果甜苷V前体的方法。
在一种实施方式中,本发明首先提供了一种高效异源合成罗汉果甜苷V前体的酶及其核酸编码序列,蛋白序列如SEQ ID No.5所示,其一种核酸序列如SEQ ID No.7所示。
其次,本发明还提供了一种高效异源合成罗汉果甜苷V前体的方法,将含有所述核酸的宿主在能够产生罗汉果甜苷V前体的条件下进行培养,然后从代谢产物中分离出罗汉果甜苷V前体即可。
本发明还提供了一种制备罗汉果甜苷V的方法,其特征在于,将含有所述核酸的宿主在能够产生罗汉果甜苷V前体的条件下进行培养,然后以该前体为底物合成罗汉果甜苷V即可。
所述宿主选自原核生物、真菌、植物细胞或动物细胞,优选的,所述宿主为酵母。
所述异源合成中进行表达的宿主为原核生物,例如,细菌,具体的,如大肠杆菌。
所述异源合成中进行表达的宿主为真核生物,例如,真菌、植物细胞或动物细胞,具体的,如酵母。
所述罗汉果甜苷V前体为11H-Cuol。
一种蛋白,其序列如SEQ ID No.5所示。
一种核酸,其序列如SEQ ID No.7所示。
含有所述核酸的生物材料,为下述A1)至A4)中的任一种:
A1)含有所述核酸的表达盒;
A2)含有所述核酸的重组载体;
A3)含有所述核酸的重组微生物;
A4)含有所述核酸的转基因植物细胞系。
所述的蛋白、或所述的核酸、或所述的生物材料在制备罗汉果甜苷V前体、或罗汉果甜苷V中的应用。
所述葫芦二烯醇的结构式如下:
在一种实施方式中,所述罗汉果甜苷V前体为11H-Cuol。
所述11H-Cuol(即11-hydroxy-cucurbitadienol)的结构式如下:
本发明的P450s酶优于现有技术的方面在于:
一、能高效异源合成罗汉果甜苷V前体。
二、通过提高前体的合成效率间接提高罗汉果甜苷V的产量
三、能高效异源表达。
可见,本发明十分难得地兼顾了P450s的酶的异源高表达和对目标产物特异的、高效的催化活性,显著超出了现有技术的水平,具有十分广阔的工业实用前景。
附图说明
图1罗汉果甜苷V异源生物合成路径。
图2从葫芦二烯醇到罗汉果醇的合成路径。
图3P450s及其突变体的代谢物检测。
图4P450s及其突变体的表达量检测。
图5验证V2为基础的突变体催化活性及蛋白表达。其中:
A:利用HPLC-qTOF检测酵母体系中相关代谢物的产量;
B:利用western blotting检测V2突变体蛋白的表达。
图6验证V3为基础的突变体催化活性及蛋白表达。其中:
A:利用HPLC-qTOF检测酵母体系中相关代谢物的产量;
B:利用western blotting检测V3突变体蛋白的表达。
图7验证混合突变体催化活性及蛋白表达。其中:
A:利用HPLC-qTOF检测酵母体系中相关代谢物的产量;
B:利用western blotting检测混合突变体蛋白的表达。
注:上述代谢物检测图中,代谢物从左到右依次为11H-Cuol(彩图为蓝色)、11C-Cuol(彩图为红色)、11C-20H-Cuol(彩图为绿色)。
具体实施方式
下述实施例中所使用的实验方法如无特殊说明,均为常规方法。下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。
实施例1、蛋白表达与检测
对P450进行突变,获得多个突变体,例如V1(L48F-S49A-I61F-L120T-T352K-L356P)、V2(L48F-S49A-I61F-L120T-K352I-L356P)、V2-I46L-A49L等,并最终获得了可以高效产生罗汉果甜苷前体11H-Cuol的突变体V4(I46L-L48F-S49L-I61F-L120T-C343Y-T352I-L356P)。
将这些突变体转化到酵母菌表达菌株INVSc1中,诱导表达12h,利用三氯乙酸-丙酮法提取酵母总蛋白,利用western blotting检测突变体蛋白的表达。三氯乙酸-丙酮法检测方法如下:取5OD酵母菌。用1mL碱裂解液(0.25M NaOH+1%β-巯基乙醇)重悬,冰上放置10min;加入160μL 50%TCA,混匀,冰上放置10min,14000g离心10min;充分弃上清,加入1mL预冷的丙酮重悬沉淀,冰上放置10min后14000g离心10min;弃上清,将丙酮充分挥发,加入100μL 1×蛋白上样缓冲液,煮沸10min,取10μL上样检测(EV为对照)。
实施例2、代谢物检测
首先,为保证足够的底物葫芦二烯醇,对酵母菌种INVSc1进行改造。HMGR、ERG20、ERG9、ERG1这四种关键酶在2,3-氧化角鲨烯的生物合成路径中至关重要,将这四个基因整合到酵母菌株染色体上过表达(提高葫芦二烯醇的含量)。在此基础上,转入环氧鲨烯合酶以及P450氧化还原伴侣CPR(Csa1G423150),获得底盘菌。
然后,将酶突变体转化到底盘酵母菌中,诱导48h,取相同质量酵母菌,利用碱裂解法提取酵母总代谢物,利HPLC-qTOF检测合成通路中化合物(目标产物11H-Cuol及副产物11C-Cuol、11C-20H-Cuol)的产量。代谢物重要检测参数如下:Q-TOF系统和APCI离子源检测正离子;HHS T3 3.5μm,4.6×150mm column;柱温35℃;流动相为A相:0.1%(v/v)甲酸水溶液,B相:0.1%(v/v)甲酸-甲醇溶液;流速为1mL/min;梯度如下B(tmin,B%):(1,90),(3,100),(10,100),(12,90),and(15,90);MS参数如下:corona current=4μA;capillaryvoltage=5.0kV;skimmer voltage=65V;segmented fragmentor=135V;gastemperature=350℃;vaporizer temperature=400℃;drying gas flow rate=8L/min;nebulizer pressure=60psi;dynamic range=150–1,000m/z。
代谢产物检测结果显示:
突变体V4产生11H-Cuol的产量平均是野生型的25-28倍左右,每克鲜重酵母产生11H-Cuol的量可达约0.28mg,目标产物11H-Cuol的产量占总产物的97.85%,酶对底物的选择特异性明显提高(表1,图3)。
而且,在酵母体系中V4突变体蛋白的表达水平与WT(野生)相比,也明显提高(图4)。
表1 每种产物占总产物的比率
实施例3、其他突变体的效果
本发明的P450s突变体是基于黄瓜(cucumis sativus)的CPY87D20的氨基酸序列,如SEQ ID No.1所示,并先后获得了包括V4在内的多个突变体,例如:突变体V1,氨基酸序列如SEQ ID No.2所示;突变体V2,氨基酸序列如SEQ ID No.3所示;突变体V3,氨基酸序列如SEQ ID No.4所示。
再比如,在上述突变体的基础上进一步突变的突变体:V2-I46L-A49L、V2-W119I-L125D、V2-R385Y、V2-W399K、V2-I439H、V2-E463P、V2-I46L-A49L-C343Y、V3-C343Y、V3-C343Y-S49L、V3-C343Y-I46L、V3-K73Y、V3-F89D、V3-Y432E、V3-L125D、V3-R383T、V3-W399D等,在实施例1-2的相同条件下分批检测。发现了若干个突变体,无论是目标产物11H-Cuol的产率、酶的底物选择特异性,还是蛋白表达水平都明显高于野生型(见图5-7)。
综上,本发明提供了一种高效异源合成罗汉果甜苷V前体的方法,同时提供了多种P450酶的突变体,不仅优化了其在酵母中的表达性能,且明显提高了目标产物11H-Cuol的产率和酶的底物选择特异性。
本发明已经尽力阐述了全部关键的技术细节,实施例仅仅为了示例本发明的构思,不应当反而成为限制本发明保护范围的理由。
SEQUENCE LISTING
<110> 云南师范大学
<120> 一种高效异源合成罗汉果甜苷V前体的方法
<130> 20210515
<160> 7
<170> PatentIn version 3.5
<210> 1
<211> 474
<212> PRT
<213> 人工合成
<400> 1
Met Trp Thr Ile Leu Leu Gly Leu Ala Thr Leu Ala Ile Ala Tyr Tyr
1 5 10 15
Ile His Trp Val Asn Lys Trp Lys Asp Ser Lys Phe Asn Gly Val Leu
20 25 30
Pro Pro Gly Thr Met Gly Leu Pro Leu Ile Gly Glu Thr Ile Gln Leu
35 40 45
Ser Arg Pro Ser Asp Ser Leu Asp Val His Pro Phe Ile Gln Arg Lys
50 55 60
Val Lys Arg Tyr Gly Pro Ile Phe Lys Thr Cys Leu Ala Gly Arg Pro
65 70 75 80
Val Val Val Ser Thr Asp Ala Glu Phe Asn His Tyr Ile Met Leu Gln
85 90 95
Glu Gly Arg Ala Val Glu Met Trp Tyr Leu Asp Thr Leu Ser Lys Phe
100 105 110
Phe Gly Leu Asp Thr Glu Trp Leu Lys Ala Leu Gly Leu Ile His Lys
115 120 125
Tyr Ile Arg Ser Ile Thr Leu Asn His Phe Gly Ala Glu Ser Leu Arg
130 135 140
Glu Arg Phe Leu Pro Arg Ile Glu Glu Ser Ala Arg Glu Thr Leu His
145 150 155 160
Tyr Trp Ser Thr Gln Thr Ser Val Glu Val Lys Glu Ser Ala Ala Ala
165 170 175
Met Val Phe Arg Thr Ser Ile Val Lys Met Phe Ser Glu Asp Ser Ser
180 185 190
Lys Leu Leu Thr Glu Gly Leu Thr Lys Lys Phe Thr Gly Leu Leu Gly
195 200 205
Gly Phe Leu Thr Leu Pro Leu Asn Leu Pro Gly Thr Thr Tyr His Lys
210 215 220
Cys Ile Lys Asp Met Lys Gln Ile Gln Lys Lys Leu Lys Asp Ile Leu
225 230 235 240
Glu Glu Arg Leu Ala Lys Gly Val Lys Ile Asp Glu Asp Phe Leu Gly
245 250 255
Gln Ala Ile Lys Asp Lys Glu Ser Gln Gln Phe Ile Ser Glu Glu Phe
260 265 270
Ile Ile Gln Leu Leu Phe Ser Ile Ser Phe Ala Ser Phe Glu Ser Ile
275 280 285
Ser Thr Thr Leu Thr Leu Ile Leu Asn Phe Leu Ala Asp His Pro Asp
290 295 300
Val Val Lys Glu Leu Glu Ala Glu His Glu Ala Ile Arg Lys Ala Arg
305 310 315 320
Ala Asp Pro Asp Gly Pro Ile Thr Trp Glu Glu Tyr Lys Ser Met Asn
325 330 335
Phe Thr Leu Asn Val Ile Cys Glu Thr Leu Arg Leu Gly Ser Val Thr
340 345 350
Pro Ala Leu Leu Arg Lys Thr Thr Lys Glu Ile Gln Ile Lys Gly Tyr
355 360 365
Thr Ile Pro Glu Gly Trp Thr Val Met Leu Val Thr Ala Ser Arg His
370 375 380
Arg Asp Pro Glu Val Tyr Lys Asp Pro Asp Thr Phe Asn Pro Trp Arg
385 390 395 400
Trp Lys Glu Leu Asp Ser Ile Thr Ile Gln Lys Asn Phe Met Pro Phe
405 410 415
Gly Gly Gly Leu Arg His Cys Ala Gly Ala Glu Tyr Ser Lys Val Tyr
420 425 430
Leu Cys Thr Phe Leu His Ile Leu Phe Thr Lys Tyr Arg Trp Arg Lys
435 440 445
Leu Lys Gly Gly Lys Ile Ala Arg Ala His Ile Leu Arg Phe Glu Asp
450 455 460
Gly Leu Tyr Val Asn Phe Thr Pro Lys Glu
465 470
<210> 2
<211> 474
<212> PRT
<213> 人工合成
<400> 2
Met Trp Thr Ile Leu Leu Gly Leu Ala Thr Leu Ala Ile Ala Tyr Tyr
1 5 10 15
Ile His Trp Val Asn Lys Trp Lys Asp Ser Lys Phe Asn Gly Val Leu
20 25 30
Pro Pro Gly Thr Met Gly Leu Pro Leu Ile Gly Glu Thr Ile Gln Phe
35 40 45
Ala Arg Pro Ser Asp Ser Leu Asp Val His Pro Phe Phe Gln Arg Lys
50 55 60
Val Lys Arg Tyr Gly Pro Ile Phe Lys Thr Cys Leu Ala Gly Arg Pro
65 70 75 80
Val Val Val Ser Thr Asp Ala Glu Phe Asn His Tyr Ile Met Leu Gln
85 90 95
Glu Gly Arg Ala Val Glu Met Trp Tyr Leu Asp Thr Leu Ser Lys Phe
100 105 110
Phe Gly Leu Asp Thr Glu Trp Thr Lys Ala Leu Gly Leu Ile His Lys
115 120 125
Tyr Ile Arg Ser Ile Thr Leu Asn His Phe Gly Ala Glu Ser Leu Arg
130 135 140
Glu Arg Phe Leu Pro Arg Ile Glu Glu Ser Ala Arg Glu Thr Leu His
145 150 155 160
Tyr Trp Ser Thr Gln Thr Ser Val Glu Val Lys Glu Ser Ala Ala Ala
165 170 175
Met Val Phe Arg Thr Ser Ile Val Lys Met Phe Ser Glu Asp Ser Ser
180 185 190
Lys Leu Leu Thr Glu Gly Leu Thr Lys Lys Phe Thr Gly Leu Leu Gly
195 200 205
Gly Phe Leu Thr Leu Pro Leu Asn Leu Pro Gly Thr Thr Tyr His Lys
210 215 220
Cys Ile Lys Asp Met Lys Gln Ile Gln Lys Lys Leu Lys Asp Ile Leu
225 230 235 240
Glu Glu Arg Leu Ala Lys Gly Val Lys Ile Asp Glu Asp Phe Leu Gly
245 250 255
Gln Ala Ile Lys Asp Lys Glu Ser Gln Gln Phe Ile Ser Glu Glu Phe
260 265 270
Ile Ile Gln Leu Leu Phe Ser Ile Ser Phe Ala Ser Phe Glu Ser Ile
275 280 285
Ser Thr Thr Leu Thr Leu Ile Leu Asn Phe Leu Ala Asp His Pro Asp
290 295 300
Val Val Lys Glu Leu Glu Ala Glu His Glu Ala Ile Arg Lys Ala Arg
305 310 315 320
Ala Asp Pro Asp Gly Pro Ile Thr Trp Glu Glu Tyr Lys Ser Met Asn
325 330 335
Phe Thr Leu Asn Val Ile Cys Glu Thr Leu Arg Leu Gly Ser Val Lys
340 345 350
Pro Ala Leu Pro Arg Lys Thr Thr Lys Glu Ile Gln Ile Lys Gly Tyr
355 360 365
Thr Ile Pro Glu Gly Trp Thr Val Met Leu Val Thr Ala Ser Arg His
370 375 380
Arg Asp Pro Glu Val Tyr Lys Asp Pro Asp Thr Phe Asn Pro Trp Arg
385 390 395 400
Trp Lys Glu Leu Asp Ser Ile Thr Ile Gln Lys Asn Phe Met Pro Phe
405 410 415
Gly Gly Gly Leu Arg His Cys Ala Gly Ala Glu Tyr Ser Lys Val Tyr
420 425 430
Leu Cys Thr Phe Leu His Ile Leu Phe Thr Lys Tyr Arg Trp Arg Lys
435 440 445
Leu Lys Gly Gly Lys Ile Ala Arg Ala His Ile Leu Arg Phe Glu Asp
450 455 460
Gly Leu Tyr Val Asn Phe Thr Pro Lys Glu
465 470
<210> 3
<211> 474
<212> PRT
<213> 人工合成
<400> 3
Met Trp Thr Ile Leu Leu Gly Leu Ala Thr Leu Ala Ile Ala Tyr Tyr
1 5 10 15
Ile His Trp Val Asn Lys Trp Lys Asp Ser Lys Phe Asn Gly Val Leu
20 25 30
Pro Pro Gly Thr Met Gly Leu Pro Leu Ile Gly Glu Thr Ile Gln Phe
35 40 45
Ala Arg Pro Ser Asp Ser Leu Asp Val His Pro Phe Phe Gln Arg Lys
50 55 60
Val Lys Arg Tyr Gly Pro Ile Phe Lys Thr Cys Leu Ala Gly Arg Pro
65 70 75 80
Val Val Val Ser Thr Asp Ala Glu Phe Asn His Tyr Ile Met Leu Gln
85 90 95
Glu Gly Arg Ala Val Glu Met Trp Tyr Leu Asp Thr Leu Ser Lys Phe
100 105 110
Phe Gly Leu Asp Thr Glu Trp Thr Lys Ala Leu Gly Leu Ile His Lys
115 120 125
Tyr Ile Arg Ser Ile Thr Leu Asn His Phe Gly Ala Glu Ser Leu Arg
130 135 140
Glu Arg Phe Leu Pro Arg Ile Glu Glu Ser Ala Arg Glu Thr Leu His
145 150 155 160
Tyr Trp Ser Thr Gln Thr Ser Val Glu Val Lys Glu Ser Ala Ala Ala
165 170 175
Met Val Phe Arg Thr Ser Ile Val Lys Met Phe Ser Glu Asp Ser Ser
180 185 190
Lys Leu Leu Thr Glu Gly Leu Thr Lys Lys Phe Thr Gly Leu Leu Gly
195 200 205
Gly Phe Leu Thr Leu Pro Leu Asn Leu Pro Gly Thr Thr Tyr His Lys
210 215 220
Cys Ile Lys Asp Met Lys Gln Ile Gln Lys Lys Leu Lys Asp Ile Leu
225 230 235 240
Glu Glu Arg Leu Ala Lys Gly Val Lys Ile Asp Glu Asp Phe Leu Gly
245 250 255
Gln Ala Ile Lys Asp Lys Glu Ser Gln Gln Phe Ile Ser Glu Glu Phe
260 265 270
Ile Ile Gln Leu Leu Phe Ser Ile Ser Phe Ala Ser Phe Glu Ser Ile
275 280 285
Ser Thr Thr Leu Thr Leu Ile Leu Asn Phe Leu Ala Asp His Pro Asp
290 295 300
Val Val Lys Glu Leu Glu Ala Glu His Glu Ala Ile Arg Lys Ala Arg
305 310 315 320
Ala Asp Pro Asp Gly Pro Ile Thr Trp Glu Glu Tyr Lys Ser Met Asn
325 330 335
Phe Thr Leu Asn Val Ile Cys Glu Thr Leu Arg Leu Gly Ser Val Ile
340 345 350
Pro Ala Leu Pro Arg Lys Thr Thr Lys Glu Ile Gln Ile Lys Gly Tyr
355 360 365
Thr Ile Pro Glu Gly Trp Thr Val Met Leu Val Thr Ala Ser Arg His
370 375 380
Arg Asp Pro Glu Val Tyr Lys Asp Pro Asp Thr Phe Asn Pro Trp Arg
385 390 395 400
Trp Lys Glu Leu Asp Ser Ile Thr Ile Gln Lys Asn Phe Met Pro Phe
405 410 415
Gly Gly Gly Leu Arg His Cys Ala Gly Ala Glu Tyr Ser Lys Val Tyr
420 425 430
Leu Cys Thr Phe Leu His Ile Leu Phe Thr Lys Tyr Arg Trp Arg Lys
435 440 445
Leu Lys Gly Gly Lys Ile Ala Arg Ala His Ile Leu Arg Phe Glu Asp
450 455 460
Gly Leu Tyr Val Asn Phe Thr Pro Lys Glu
465 470
<210> 4
<211> 474
<212> PRT
<213> 人工合成
<400> 4
Met Trp Thr Ile Leu Leu Gly Leu Ala Thr Leu Ala Ile Ala Tyr Tyr
1 5 10 15
Ile His Trp Val Asn Lys Trp Lys Asp Ser Lys Phe Asn Gly Val Leu
20 25 30
Pro Pro Gly Thr Met Gly Leu Pro Leu Ile Gly Glu Thr Ile Gln Leu
35 40 45
Ser Arg Pro Ser Asp Ser Leu Asp Val His Pro Phe Ile Gln Arg Lys
50 55 60
Val Lys Arg Tyr Gly Pro Ile Phe Lys Thr Cys Leu Ala Gly Arg Pro
65 70 75 80
Val Val Val Ser Thr Asp Ala Glu Phe Asn His Tyr Ile Met Leu Gln
85 90 95
Glu Gly Arg Ala Val Glu Met Trp Tyr Leu Asp Thr Phe Ser Lys Phe
100 105 110
Leu Gly Leu Asp Thr Glu Trp Leu Lys Ala Leu Gly Leu Ile His Lys
115 120 125
Tyr Ile Arg Ser Ile Thr Leu Asn His Phe Gly Ala Glu Ser Leu Arg
130 135 140
Glu Arg Phe Leu Pro Arg Ile Glu Glu Ser Ala Arg Glu Thr Leu His
145 150 155 160
Tyr Trp Ser Thr Gln Thr Ser Val Glu Val Lys Glu Ser Ala Ala Ala
165 170 175
Met Val Phe Arg Thr Ser Ile Val Lys Met Phe Ser Glu Asp Ser Ser
180 185 190
Lys Leu Leu Thr Glu Gly Leu Thr Lys Lys Phe Thr Gly Leu Leu Gly
195 200 205
Gly Phe Leu Thr Leu Pro Leu Asn Leu Pro Gly Thr Thr Tyr His Lys
210 215 220
Cys Ile Lys Asp Met Lys Gln Ile Gln Lys Lys Leu Lys Asp Ile Leu
225 230 235 240
Glu Glu Arg Leu Ala Lys Gly Val Lys Ile Asp Glu Asp Phe Leu Gly
245 250 255
Gln Ala Ile Lys Asp Lys Glu Ser Gln Gln Phe Ile Ser Glu Glu Phe
260 265 270
Ile Ile Gln Leu Leu Phe Ser Ile Ser Phe Ala Ser Phe Ala Ser Ile
275 280 285
Ser Thr Thr Leu Thr Leu Ile Leu Asn Phe Leu Ala Asp His Pro Asp
290 295 300
Val Val Lys Glu Leu Glu Ala Glu His Glu Ala Ile Arg Lys Ala Arg
305 310 315 320
Ala Asp Pro Asp Gly Pro Ile Thr Trp Glu Glu Tyr Lys Ser Met Asn
325 330 335
Phe Thr Leu Asn Val Ile Cys Glu Thr Leu Arg Leu Gly Ser Val Thr
340 345 350
Pro Ala Leu Leu Arg Lys Thr Thr Lys Glu Ile Gln Ile Lys Gly Tyr
355 360 365
Thr Ile Pro Glu Gly Trp Thr Val Met Leu Val Thr Ala Ser Arg His
370 375 380
Arg Asp Pro Glu Val Tyr Lys Asp Pro Asp Thr Phe Asn Pro Trp Arg
385 390 395 400
Trp Lys Glu Leu Asp Ser Ile Thr Ile Gln Lys Asn Phe Met Pro Phe
405 410 415
Gly Gly Gly Leu Arg His Cys Ala Gly Ala Glu Tyr Ser Lys Val Tyr
420 425 430
Leu Cys Thr Phe Leu His Ile Leu Phe Thr Lys Tyr Arg Trp Arg Lys
435 440 445
Leu Lys Gly Gly Lys Ile Ala Arg Ala His Ile Leu Arg Phe Glu Asp
450 455 460
Gly Leu Tyr Val Asn Phe Thr Pro Lys Glu
465 470
<210> 5
<211> 474
<212> PRT
<213> 人工合成
<400> 5
Met Trp Thr Ile Leu Leu Gly Leu Ala Thr Leu Ala Ile Ala Tyr Tyr
1 5 10 15
Ile His Trp Val Asn Lys Trp Lys Asp Ser Lys Phe Asn Gly Val Leu
20 25 30
Pro Pro Gly Thr Met Gly Leu Pro Leu Ile Gly Glu Thr Leu Gln Phe
35 40 45
Leu Arg Pro Ser Asp Ser Leu Asp Val His Pro Phe Phe Gln Arg Lys
50 55 60
Val Lys Arg Tyr Gly Pro Ile Phe Lys Thr Cys Leu Ala Gly Arg Pro
65 70 75 80
Val Val Val Ser Thr Asp Ala Glu Phe Asn His Tyr Ile Met Leu Gln
85 90 95
Glu Gly Arg Ala Val Glu Met Trp Tyr Leu Asp Thr Leu Ser Lys Phe
100 105 110
Phe Gly Leu Asp Thr Glu Trp Thr Lys Ala Leu Gly Leu Ile His Lys
115 120 125
Tyr Ile Arg Ser Ile Thr Leu Asn His Phe Gly Ala Glu Ser Leu Arg
130 135 140
Glu Arg Phe Leu Pro Arg Ile Glu Glu Ser Ala Arg Glu Thr Leu His
145 150 155 160
Tyr Trp Ser Thr Gln Thr Ser Val Glu Val Lys Glu Ser Ala Ala Ala
165 170 175
Met Val Phe Arg Thr Ser Ile Val Lys Met Phe Ser Glu Asp Ser Ser
180 185 190
Lys Leu Leu Thr Glu Gly Leu Thr Lys Lys Phe Thr Gly Leu Leu Gly
195 200 205
Gly Phe Leu Thr Leu Pro Leu Asn Leu Pro Gly Thr Thr Tyr His Lys
210 215 220
Cys Ile Lys Asp Met Lys Gln Ile Gln Lys Lys Leu Lys Asp Ile Leu
225 230 235 240
Glu Glu Arg Leu Ala Lys Gly Val Lys Ile Asp Glu Asp Phe Leu Gly
245 250 255
Gln Ala Ile Lys Asp Lys Glu Ser Gln Gln Phe Ile Ser Glu Glu Phe
260 265 270
Ile Ile Gln Leu Leu Phe Ser Ile Ser Phe Ala Ser Phe Glu Ser Ile
275 280 285
Ser Thr Thr Leu Thr Leu Ile Leu Asn Phe Leu Ala Asp His Pro Asp
290 295 300
Val Val Lys Glu Leu Glu Ala Glu His Glu Ala Ile Arg Lys Ala Arg
305 310 315 320
Ala Asp Pro Asp Gly Pro Ile Thr Trp Glu Glu Tyr Lys Ser Met Asn
325 330 335
Phe Thr Leu Asn Val Ile Tyr Glu Thr Leu Arg Leu Gly Ser Val Ile
340 345 350
Pro Ala Leu Pro Arg Lys Thr Thr Lys Glu Ile Gln Ile Lys Gly Tyr
355 360 365
Thr Ile Pro Glu Gly Trp Thr Val Met Leu Val Thr Ala Ser Arg His
370 375 380
Arg Asp Pro Glu Val Tyr Lys Asp Pro Asp Thr Phe Asn Pro Trp Arg
385 390 395 400
Trp Lys Glu Leu Asp Ser Ile Thr Ile Gln Lys Asn Phe Met Pro Phe
405 410 415
Gly Gly Gly Leu Arg His Cys Ala Gly Ala Glu Tyr Ser Lys Val Tyr
420 425 430
Leu Cys Thr Phe Leu His Ile Leu Phe Thr Lys Tyr Arg Trp Arg Lys
435 440 445
Leu Lys Gly Gly Lys Ile Ala Arg Ala His Ile Leu Arg Phe Glu Asp
450 455 460
Gly Leu Tyr Val Asn Phe Thr Pro Lys Glu
465 470
<210> 6
<211> 474
<212> PRT
<213> 人工合成
<400> 6
Met Trp Thr Ile Leu Leu Gly Leu Ala Thr Leu Ala Ile Ala Tyr Tyr
1 5 10 15
Ile His Trp Val Asn Lys Trp Lys Asp Ser Lys Phe Asn Gly Val Leu
20 25 30
Pro Pro Gly Thr Met Gly Leu Pro Leu Ile Gly Glu Thr Ile Gln Phe
35 40 45
Ala Arg Pro Ser Asp Ser Leu Asp Val His Pro Phe Phe Gln Arg Lys
50 55 60
Val Lys Arg Tyr Gly Pro Ile Phe Lys Thr Cys Leu Ala Gly Arg Pro
65 70 75 80
Val Val Val Ser Thr Asp Ala Glu Phe Asn His Tyr Ile Met Leu Gln
85 90 95
Glu Gly Arg Ala Val Glu Met Trp Tyr Leu Asp Thr Leu Ser Lys Phe
100 105 110
Phe Gly Leu Asp Thr Glu Ile Thr Lys Ala Leu Gly Asp Ile His Lys
115 120 125
Tyr Ile Arg Ser Ile Thr Leu Asn His Phe Gly Ala Glu Ser Leu Arg
130 135 140
Glu Arg Phe Leu Pro Arg Ile Glu Glu Ser Ala Arg Glu Thr Leu His
145 150 155 160
Tyr Trp Ser Thr Gln Thr Ser Val Glu Val Lys Glu Ser Ala Ala Ala
165 170 175
Met Val Phe Arg Thr Ser Ile Val Lys Met Phe Ser Glu Asp Ser Ser
180 185 190
Lys Leu Leu Thr Glu Gly Leu Thr Lys Lys Phe Thr Gly Leu Leu Gly
195 200 205
Gly Phe Leu Thr Leu Pro Leu Asn Leu Pro Gly Thr Thr Tyr His Lys
210 215 220
Cys Ile Lys Asp Met Lys Gln Ile Gln Lys Lys Leu Lys Asp Ile Leu
225 230 235 240
Glu Glu Arg Leu Ala Lys Gly Val Lys Ile Asp Glu Asp Phe Leu Gly
245 250 255
Gln Ala Ile Lys Asp Lys Glu Ser Gln Gln Phe Ile Ser Glu Glu Phe
260 265 270
Ile Ile Gln Leu Leu Phe Ser Ile Ser Phe Ala Ser Phe Glu Ser Ile
275 280 285
Ser Thr Thr Leu Thr Leu Ile Leu Asn Phe Leu Ala Asp His Pro Asp
290 295 300
Val Val Lys Glu Leu Glu Ala Glu His Glu Ala Ile Arg Lys Ala Arg
305 310 315 320
Ala Asp Pro Asp Gly Pro Ile Thr Trp Glu Glu Tyr Lys Ser Met Asn
325 330 335
Phe Thr Leu Asn Val Ile Cys Glu Thr Leu Arg Leu Gly Ser Val Ile
340 345 350
Pro Ala Leu Pro Arg Lys Thr Thr Lys Glu Ile Gln Ile Lys Gly Tyr
355 360 365
Thr Ile Pro Glu Gly Trp Thr Val Met Leu Val Thr Ala Ser Arg His
370 375 380
Arg Asp Pro Glu Val Tyr Lys Asp Pro Asp Thr Phe Asn Pro Trp Arg
385 390 395 400
Trp Lys Glu Leu Asp Ser Ile Thr Ile Gln Lys Asn Phe Met Pro Phe
405 410 415
Gly Gly Gly Leu Arg His Cys Ala Gly Ala Glu Tyr Ser Lys Val Tyr
420 425 430
Leu Cys Thr Phe Leu His Ile Leu Phe Thr Lys Tyr Arg Trp Arg Lys
435 440 445
Leu Lys Gly Gly Lys Ile Ala Arg Ala His Ile Leu Arg Phe Glu Asp
450 455 460
Gly Leu Tyr Val Asn Phe Thr Pro Lys Glu
465 470
<210> 7
<211> 1425
<212> DNA
<213> 人工合成
<400> 7
atgtggacga tcttgctcgg tttggcgacg ttggcaattg cctactatat tcattgggtt 60
aacaaatgga aggattctaa attcaacgga gttttgccgc cgggcaccat ggggctgccc 120
ctcatcggag aaacccttca atttcttcgc cctagtgact cccttgatgt tcatcctttc 180
tttcaacgca aagttaaaag atatggaccg atcttcaaga cttgtttggc gggaaggccg 240
gtggtggttt caacggatgc agagtttaac cattacataa tgctccaaga aggaagggcc 300
gtagaaatgt ggtatttgga tacactctct aaattctttg gccttgacac tgaatggacc 360
aaagcccttg gcctcatcca caaatacatt agaagcatta ctttgaacca ctttggtgct 420
gagtcccttc gtgagcgttt ccttcctcgt atcgaagaat ccgctcgaga aacccttcat 480
tattggtcaa ctcaaaccag cgttgaagtc aaggaatcag ccgctgcgat ggttttcaga 540
acttcgattg ttaagatgtt tagtgaagat tctagtaaat tactgacaga aggtctcact 600
aagaagttca caggacttct cggaggtttt ctcaccttgc ctctaaattt gcctggcact 660
acctatcata aatgcataaa ggacatgaag caaatccaaa agaagctaaa agacatttta 720
gaggaaagat tggctaaagg ggttaaaatt gatgaagatt tcttggggca agccattaaa 780
gataaagaat ctcaacaatt catttcagag gaattcatta tccagttgtt gttttccatc 840
agctttgcta gctttgagtc catctctacc actcttactt tgattctcaa cttcctcgcc 900
gatcaccccg acgtagtgaa agaattggag gctgagcatg aggctattag aaaggcaagg 960
gcagatccag atggaccaat cacttgggaa gaatacaaat ccatgaattt cacactcaat 1020
gtcatctatg aaacacttag gttgggaagt gtaatacctg ctttgccgag gaagacaacc 1080
aaggaaattc aaataaaagg atacacaatt ccagaaggat ggacagtaat gcttgtgacc 1140
gcttctcgtc atagagatcc agaagtgtac aaggatcccg ataccttcaa tccatggcgt 1200
tggaaggagt tggactcaat tactattcaa aagaacttca tgccatttgg gggaggctta 1260
aggcattgtg ctggtgctga atactctaaa gtctatttgt gcactttcct tcatatcctt 1320
ttcaccaaat acagatggag aaaactaaag ggaggaaaga ttgcaagggc tcatatattg 1380
aggtttgaag atgggttata tgtgaacttc actcccaagg aatga 1425
- 一种高效异源合成罗汉果甜苷V前体的方法
- 一种高效异源合成罗汉果甜苷V前体的方法