包含抗FAP克隆212的双特异性抗原结合分子
文献发布时间:2023-06-19 11:21:00
技术领域
本发明涉及新型双特异性抗原结合分子,其包含至少一个能够与成纤维细胞活化蛋白(FAP)特异性结合的抗原结合结构域、至少一个能够与CD40特异性结合的抗原结合结构域以及由能够稳定缔合的第一亚基和第二亚基组成的Fc结构域。本发明进一步涉及一种新型抗FAP克隆212。本发明的进一步方面为生产这些分子的方法以及使用所述分子的方法。
背景技术
在产生有效的适应性免疫应答期间,需要多个分子信号。信号一涉及T细胞抗原受体(TCR)与存在于抗原呈递细胞(APC)表面的同源抗原的结合。信号二由共刺激受体与在T细胞和APC之间的其各自配体的接合组成。研究最深入的以及最重要的共刺激效应子之一是肿瘤坏死因子受体(TNFR)家族成员CD40及其配体CD40L(Elgueta R.等人,ImmunolRev.2009;229(1):152-72)。初始T细胞活化后,TNFR家族的几个成员(包括CD40)起维持APC和T细胞应答的作用,并且因此在免疫系统的组织和功能中起关键作用(Watts T.H.(2005)Annu.Rev.Immunol.23,23-68)。不同共刺激TNFR家族成员的组合允许对APC和T细胞活化和存活进行顺序和瞬时调节,从而导致免疫应答增强,同时保持对APC和T细胞功能的严格控制。根据疾病状况,经由共刺激TNF家族成员的刺激可以加剧或改善疾病。TNFR家族共刺激物的活化或阻断显示了在多个领域(包括癌症、传染病、移植和自身免疫)中的几种治疗应用的前景。
在几个共刺激分子中,TNFR家族成员CD40通过诱导APC的成熟、APC的存活、APC的抗原呈递、APC的细胞因子生产和APC的共刺激分子的表达在触发免疫应答中起关键作用,其然后通过促炎性细胞因子驱动抗原特异性T细胞应答和NK细胞活化。CD40调节针对感染、肿瘤和自身抗原的免疫应答,并且其表达已在APC(诸如B细胞、树突细胞(DC)、单核细胞、巨噬细胞以及血小板)和非造血来源的细胞(诸如成肌纤维细胞、成纤维细胞、上皮细胞和内皮细胞)的表面上得到证实(Elgueta R.等人,Immunol Rev.2009;229(1):152-72)。CD40配体CD40L在活化的CD4
I型跨膜受体CD40通过其天然配体CD40L(II型跨膜蛋白)或通过激动性抗体的接合促进CD40簇聚并且诱导衔接蛋白募集到胞质受体结构域。这些称为TNF受体相关因子(TRAF)的衔接蛋白的募集导致促分裂原激活蛋白激酶(MAPK)、磷脂酰肌醇3激酶(PI3K)以及规范性和非规范性核因子κB(NFκB)信号传导通路的协同活化(Elgueta R.等人,ImmunolRev.2009;229(1):152-72)。继而,这导致APC的成熟和活化,其然后使抗原特异性T细胞应答最大化。最近的研究显示了激动性CD40抗体在利用抗肿瘤免疫中的两种不同作用方式。除了通过活化适应性免疫系统介导肿瘤细胞杀伤的间接作用方式之外,激动性CD40抗体还可以通过诱导表达CD40的实体瘤细胞凋亡来诱导直接肿瘤细胞杀伤(Eliopoulos AG.等人,Mol Cell Biol.2000;20(15):5503-15)。直接CD40抗体介导的肿瘤细胞杀伤可以提供肿瘤抗原来源,其可通过经由抗CD40抗体的CD40接合同时活化的APC进行处理和呈递,然后这可以诱导肿瘤抗原特异性T细胞,一种称为内源性疫苗接种的假定的机制。鉴于CD40接合可以促进有效的抗癌免疫应答,因此激动性CD40抗体已成功用于多种临床前肿瘤模型,其既可以作为单一药剂也可以与化学疗法联用(Vonderheide RH和Glennie MJ.,ClinCancer Res.2013;19(5):1035-43)。
迄今为止,六种CD40 mAb在临床试验中进行研究:Chi Lob 7/4(CD40激动性IgG1嵌合mAb;Cancer Research UK;Chowdhury F.等人,Cancer Immunol Res.2013;2:229–40);ADC1013(完全人CD40激动性IgG1抗体;Alligator Bioscience和Johnson&Johnson;Mangsbo SM.等人,Clin Cancer Res.2015Mar 1;21(5):1115-26);APX-005(完全人源化CD40激动性IgG1 mAb;Apexigen;Bjorck P.等人,J Immunother Cancer.2015;3(Suppl2):P198);SEA-CD40(CD40激动性IgG1嵌合mAb;Seattle Genetics;Gardai SJ.等人,AACR106th Annual Meeting 2015;April 18-22,摘要2472);及正在临床I期研究中进行研究的RO7009789(完全人,CD40超级激动性IgG2 mAb);以及正在临床II期研究中进行研究的达西珠单抗(CD40部分激动性IgG1嵌合mAb;Seattle Genetics;Khubchandani S.等人,CurrOpin Investig Drugs.2009;10,579–87)。这些研究的符合条件患者患有实体瘤、经典型霍奇金淋巴瘤(HL)、弥漫性大B细胞淋巴瘤(DLBCL)或惰性淋巴瘤(包括滤泡性淋巴瘤)。这些CD40激动性抗体已显示出多种活性,所述多种活性范围从经由补体介导的细胞毒性(CMC)或抗体依赖性细胞毒性(ADCC)的CD40
激动性CD40抗体的临床开发已提供了有希望的初步结果。在第一临床试验中,CP-870,893已显示出对晚期癌症患者的临床功效。29例晚期癌症患者中的4例在接受单次静脉输注CP-870,893后表现出部分应答(Vonderheide RH.,J Clin Oncol.2007Mar 1;25(7):876-83)。在一年半的时间内用9个后续剂量的CP-870,893治疗的四例患者中的一例保持完全缓解超过5年。然而,CP-870,893的最常见副作用是细胞因子释放综合征和血栓栓塞事件,使得在使用的剂量时间表和施用途径的情况下,超过140例患者的1期临床研究的合并数据仅指示有限的临床功效,并且建议局部施用抗体(Vonderheide RH,Glennie M,ClinCancer Res.2013,19(5),1035-1043)。缺乏单一药剂应答的部分原因是由于CD40广泛表达引起的严重的上靶/离靶肿瘤效应,这导致剂量限制性毒性(例如,细胞因子释放综合征)。当将CD40通过肿瘤特异性靶标交联时,特异性活化APC的激动性CD40抗体的开发可以减少副作用并降低剂量限制,提供新治疗选择,并有可能产生有效的持久抗癌免疫力。
现有的临床前和临床数据清楚地证明,临床上迫切需要能够诱导和增强对癌症有效的内源性免疫应答的有效CD40激动剂。然而,这类作用几乎永远不会局限于单一类型的细胞或经由单一机制起作用,并且设计成阐明细胞间和细胞内信号传导机制的研究揭示了复杂性水平的提高。由于剂量限制性毒性(诸如,细胞因子释放综合征和血小板/内皮细胞活化),已知的CD40抗体只能以相对低的剂量施用,从而导致靶APC上的途径活化不足和窄治疗指数。因此,需要优选地作用于单一类型细胞的“靶向”激动剂。
本发明涉及能够与CD40和成纤维细胞活化蛋白(FAP)特异性结合的新型双特异性抗原结合分子,并且因此将能够与FAP结合的部分与能够激动性结合CD40的部分相结合,其中通过在肿瘤基质细胞上表达的FAP以及潜在地也通过在次级淋巴组织中中间表达的FAP交联提供了CD40对APC的活化。与能够与CD40以及与活化T细胞上的免疫检查点受体(诸如,CTLA-4或PD-1)特异性结合的双特异性抗原结合分子相反,靶向肿瘤靶标诸如FAP可使CD40介导的APC活化主要在肿瘤基质和肿瘤引流淋巴结中发生,其中与其他组织相比,成纤维细胞表达的FAP水平升高。因此,本发明的抗原结合分子不仅能够有效地而且还能够非常有选择性地在所需位点触发CD40受体,同时克服了FcγR交联交联的需求,从而减少了副作用。新型双特异性抗原结合分子的特征还在于包含一种新型FAP抗原结合结构域,当该FAP抗原结合结构域融合至Fc结构域的C末端时不失去其优异的结合特性。
发明内容
本发明涉及双特异性抗原结合分子,这些双特异性抗原结合分子将至少一个能够与共刺激TNF受体家族成员CD40特异性结合的抗原结合结构域与至少一个靶向成纤维细胞活化蛋白(FAP)的抗原结合结构域结合,其包含新型抗人FAP克隆212及其人源化变体。这些双特异性抗原结合分子是有利的,因为它们优选在表达FAP的肿瘤相关部位激活共刺激CD40受体,因为它们能够以高亲和力与FAP结合。
在一个方面,本发明提供了一种双特异性抗原结合分子,其包含:
(a)至少一个能够与CD40特异性结合的抗原结合结构域;以及
(b)至少一个能够与成纤维细胞活化蛋白(FAP)特异性结合的抗原结合结构域,其包含:重链可变区(V
(i)CDR-H1,其包含SEQ ID NO:3的氨基酸序列,(ii)CDR-H2,其包含选自由SEQ IDNO:4、SEQ ID NO:11和SEQ ID NO:12组成的组的氨基酸序列,和(iii)CDR-H3,其包含SEQID NO:5的氨基酸序列;以及轻链可变区(V
(vi)CDR-L3,其包含SEQ ID NO:8的氨基酸序列。
在某一特定方面,双特异性抗原结合分子包含:(a)至少一个能够与CD40特异性结合的抗原结合结构域;(b)至少一个能够与FAP特异性结合的抗原结合结构域,该FAP包含:重链可变区(V
在一个方面,所述至少一个能够与FAP特异性结合的抗原结合结构域:重链可变区(V
在另一方面,能够与FAP特异性结合的抗原结合结构域包含重链可变区(V
在一个方面,能够与CD40特异性结合的抗原结合结构域结合到包含SEQ ID NO:1的氨基酸序列的多肽或由其组成的多肽。
在进一步方面,提供了一种双特异性抗原结合分子,其中至少一个能够与CD40特异性结合的抗原结合结构域包含:重链可变区(V
在一个方面,提供了如上文所定义的双特异性抗原结合分子,其中至少一个能够与CD40特异性结合的抗原结合结构域包含
(i)重链可变区(V
(ii)轻链可变区(V
在另一方面,提供了如上文所定义的双特异性抗原结合分子,其中至少一个能够与CD40特异性结合的抗原结合结构域包含:(i)重链可变区(V
(ii)轻链可变区(V
此外,提供了如上文所定义的双特异性抗原结合分子,其中至少一个能够与CD40特异性结合的抗原结合结构域包含:
(a)VH,其包含SEQ ID NO:37的氨基酸序列,和VL,其包含SEQ ID NO:41的氨基酸序列,或者
(b)VH,其包含SEQ ID NO:37的氨基酸序列,和VL,其包含SEQ ID NO:42的氨基酸序列,或者
(c)VH,其包含SEQ ID NO:37的氨基酸序列,和VL,其包含SEQ ID NO:43的氨基酸序列,或者
(d)VH,其包含SEQ ID NO:37的氨基酸序列,和VL,其包含SEQ ID NO:44的氨基酸序列,或者
(e)VH,其包含SEQ ID NO:38的氨基酸序列,和VL,其包含SEQ ID NO:41的氨基酸序列,或者
(f)VH,其包含SEQ ID NO:38的氨基酸序列,和VL,其包含SEQ ID NO:42的氨基酸序列,或者
(g)VH,其包含SEQ ID NO:38的氨基酸序列,和VL,其包含SEQ ID NO:43的氨基酸序列,或者
(h)VH,其包含SEQ ID NO:38的氨基酸序列,和VL,其包含SEQ ID NO:44的氨基酸序列,或者
(i)VH,其包含SEQ ID NO:39的氨基酸序列,和VL,其包含SEQ ID NO:41的氨基酸序列,或者
(j)VH,其包含SEQ ID NO:39的氨基酸序列,和VL,其包含SEQ ID NO:42的氨基酸序列,或者
(k)VH,其包含SEQ ID NO:39的氨基酸序列,和VL,其包含SEQ ID NO:43的氨基酸序列,或者
(l)VH,其包含SEQ ID NO:39的氨基酸序列,和VL,其包含SEQ ID NO:44的氨基酸序列,或者
(m)VH,其包含SEQ ID NO:40的氨基酸序列,和VL,其包含SEQ ID NO:41的氨基酸序列,或者
(n)VH,其包含SEQ ID NO:40的氨基酸序列,和VL,其包含SEQ ID NO:42的氨基酸序列,或者
(o)VH,其包含SEQ ID NO:40的氨基酸序列,和VL,其包含SEQ ID NO:43的氨基酸序列,或者
(p)VH,其包含SEQ ID NO:40的氨基酸序列,和VL,其包含SEQ ID NO:44的氨基酸序列。
在某一特定方面,提供了一种双特异性抗原结合分子,其中至少一个能够与CD40特异性结合的抗原结合结构域包含:VH,其包含SEQ ID NO:37的氨基酸序列,和VL,其包含SEQ ID NO:41的氨基酸序列。
在进一步方面,提供了如上文所定义的双特异性抗原结合分子,其中至少一个能够与CD40特异性结合的抗原结合结构域包含:
(a)VH,其包含SEQ ID NO:45的氨基酸序列,和VL,其包含SEQ ID NO:51的氨基酸序列,或者
(b)VH,其包含SEQ ID NO:46的氨基酸序列,和VL,其包含SEQ ID NO:51的氨基酸序列,或者
(c)VH,其包含SEQ ID NO:47的氨基酸序列,和VL,其包含SEQ ID NO:51的氨基酸序列,或者
(d)VH,其包含SEQ ID NO:48的氨基酸序列,和VL,其包含SEQ ID NO:51的氨基酸序列,或者
(e)VH,其包含SEQ ID NO:45的氨基酸序列,和VL,其包含SEQ ID NO:52的氨基酸序列,或者
(f)VH,其包含SEQ ID NO:46的氨基酸序列,和VL,其包含SEQ ID NO:52的氨基酸序列,或者
(g)VH,其包含SEQ ID NO:47的氨基酸序列,和VL,其包含SEQ ID NO:52的氨基酸序列,或者
(h)VH,其包含SEQ ID NO:48的氨基酸序列,和VL,其包含SEQ ID NO:52的氨基酸序列,或者
(i)VH,其包含SEQ ID NO:49的氨基酸序列,和VL,其包含SEQ ID NO:53的氨基酸序列,或者
(j)VH,其包含SEQ ID NO:50的氨基酸序列,和VL,其包含SEQ ID NO:53的氨基酸序列,或者
(k)VH,其包含SEQ ID NO:49的氨基酸序列,和VL,其包含SEQ ID NO:54的氨基酸序列,或者
(l)VH,其包含SEQ ID NO:50的氨基酸序列,和VL,其包含SEQ ID NO:54的氨基酸序列。
在进一步特定方面,提供了一种双特异性抗原结合分子,其中至少一个能够与CD40特异性结合的抗原结合结构域包含:VH,其包含SEQ ID NO:45的氨基酸序列,和VL,其包含SEQ ID NO:51的氨基酸序列,或者其中能够与CD40特异性结合的抗原结合结构域包含:VH,其包含SEQ ID NO:48的氨基酸序列,和VL,其包含SEQ ID NO:51的氨基酸序列。
更具体地,提供了一种双特异性抗原结合分子,其包含:
(i)至少一个能够与CD40特异性结合的抗原结合结构域,其包含:重链可变区(V
(ii)至少一个能够与FAP特异性结合的抗原结合结构域,其包含:重链可变区(V
在一个方面,双特异性抗原结合分子为人源化或嵌合抗体。在进一步方面,双特异性抗原结合分子包含IgG Fc区,特别是IgG1 Fc区或IgG4Fc区。特别地,Fc区包含降低抗体与Fc受体的结合亲和力和/或效应子功能的一个或多个氨基酸取代。在某一特定方面,提供了一种双特异性抗原结合分子,其中Fc区属于人IgG1亚类,具有氨基酸突变L234A、L235A和P329G(根据Kabat EU索引编号)。
在另一方面,提供了如上文所定义的双特异性抗原结合分子,其中根据杵臼结构方法,Fc区的第一亚基包含突起并且Fc区的第二亚基包含孔。特别地,提供了一种双特异性抗原结合分子,其中(i)所述Fc区的第一亚基包含氨基酸取代S354C和T366W(根据Kabat EU索引编号),并且所述Fc区的第二亚基包含氨基酸取代Y349C、T366S和Y407V(根据Kabat EU索引编号),或(ii)所述Fc区的第一亚基包含氨基酸取代K392D和K409D(根据Kabat EU索引编号),并且所述Fc区的第二亚基包含氨基酸取代E356K和D399K(根据Kabat EU索引编号)。更特别地,提供了一种双特异性抗原结合分子,其中Fc区的第一亚基包含氨基酸取代S354C和T366W(根据Kabat EU索引编号),并且Fc区的第二亚基包含氨基酸取代Y349C、T366S和Y407V(根据Kabat EU索引编号)。
在进一步方面,提供了一种双特异性抗原结合分子,其中双特异性抗原结合分子包含:
(a)至少两个能够与CD40特异性结合的Fab片段,其连接至Fc区,以及
(b)一个能够与FAP特异性结合的抗原结合结构域,其连接至所述Fc区的C末端。
因此,提供了一种双特异性抗原结合分子,该双特异性抗原结合分子提供了与CD40的二价结合以及与FAP的单价结合。
在另一方面,提供了一种双特异性抗原结合分子,其中双特异性抗原结合分子包含:
(a)至少两个能够与CD40特异性结合的Fab片段,其融合至Fc区,以及
(b)一个能够与FAP特异性结合的抗原结合结构域,其融合至Fc区的C末端。
在某一特定方面,能够与FAP特异性结合的抗原结合结构域为交叉fab片段,该抗原结合结构域连接至Fc区的C末端。因此,提供了一种双特异性抗原结合分子,其中双特异性抗原结合分子包含:
(a)至少两个能够与CD40特异性结合的Fab片段,其融合至Fc区,以及
(b)能够与FAP特异性结合的交叉fab片段,其融合至所述Fc区的C末端。
在进一步方面,双特异性抗原结合分子包含四个能够与CD40特异性结合的Fab片段。因此,提供了一种双特异性抗原结合分子,该双特异性抗原结合分子提供了与CD40的四价结合以及与FAP的单价结合。
在一个方面,提供了一种双特异性抗原结合分子,其包含:
(a)抗体的两个轻链和两个重链,该抗体包含两个能够与CD40特异性结合的Fab片段以及Fc区;以及
(b)能够与FAP特异性结合的抗原结合结构域的VH和VL,其中VH融合至(a)的两个重链中的一个的C末端,并且其中VL融合至(a)的两个重链中的另一个的C末端。
在另一方面,提供了一种双特异性抗原结合分子,其包含:
(a)抗体的两个轻链和两个重链,该抗体包含两个能够与CD40特异性结合的Fab片段以及Fc区;以及
(b)能够与FAP特异性结合的交叉fab片段,其中VH-Cκ链融合至(a)的两个重链中的一个的C末端。
在又一方面,提供了一种双特异性抗原结合分子,其包含:
(a)抗体的两个轻链和两个重链,该抗体包含两个能够与CD40特异性结合的Fab片段以及Fc区;以及
(b)能够与FAP特异性结合的交叉fab片段,其中VL-CH1链融合至(a)的两个重链中的一个的C末端。
此外,提供了一种双特异性抗原结合分子,其包含:
(a)抗体的两个轻链和两个重链,该抗体包含两个能够与CD40特异性结合的Fab片段以及Fc区;以及
(b)两个能够与FAP特异性结合的Fab片段,其中Fab片段中的一个连接至(a)的两个重链中的一个的C末端,并且Fab片段中的另一个连接至(a)的两个重链中的另一个的C末端。
在另一方面,提供了一种双特异性抗原结合分子,其中双特异性抗原结合分子包含:
(a)两个重链,其中每个重链包含能够与CD40特异性结合的Fab片段的VH结构域和CH1结构域以及Fc区亚基;
(b)两个轻链,其中每个轻链包含能够与CD40特异性结合的Fab片段的VL和CL结构域;以及
(c)一个能够与FAP特异性结合的Fab片段,其中Fab片段连接至(a)的两个重链中的一个的C末端。
在另一方面,能够与FAP特异性结合的Fab片段为交叉fab片段,其包含VL-CH1链和VH-Cκ链,并且其中VH-Cκ链或VL-CH1链连接至(a)的两个重链中的一个的C末端。
在一个方面,提供了一种双特异性抗原结合分子,其中双特异性抗原结合分子包含四个能够与CD40特异性结合的Fab片段。在某一特定方面,提供了一种双特异性抗原结合分子,其中如上文所定义的(a)的两个重链中的每个包含能够与CD40特异性结合的Fab片段的两条VH-CH1链,这两条VH-CH1链彼此连接,并且任选地通过肽连接基连接。
因此,在一个方面,本发明提供了一种双特异性抗原结合分子,其包含:
(a)两个重链,其中每个重链包含能够与CD40特异性结合的Fab片段的两条VH-CH1链以及Fc区亚基,这两条VH-CH1链彼此连接,并且任选地通过肽连接基连接;
(b)四个轻链,其中每个轻链包含能够与CD40特异性结合的Fab片段的VL和Cκ结构域;以及
(c)能够与FAP特异性结合的交叉fab片段,其包含VL-CH1链和VH-Cκ链,其中VH-Cκ链或VL-CH1链连接至(a)的两个重链中的一个的C末端,并且任选地通过肽连接基连接。
在另一方面,提供了一种双特异性抗原结合分子,其包含:
(a)两个重链,其中每个重链包含能够与CD40特异性结合的Fab片段的两条VH-CH1链以及Fc区亚基,这两条VH-CH1链彼此连接,并且任选地通过肽连接基连接;
(b)四个轻链,其中每个轻链包含能够与CD40特异性结合的Fab片段的VL和CL结构域;以及
(c)能够与FAP特异性结合的交叉fab片段,其中所述交叉fab片段的VH-CL链连接至(a)的两个重链中的一个的C末端。
在又一方面,提供了一种双特异性抗原结合分子,其包含:
(a)两个重链,其中每个重链包含能够与CD40特异性结合的Fab片段的两条VH-CH1链以及Fc区亚基,这两条VH-CH1链彼此连接,并且任选地通过肽连接基连接;
(b)四个轻链,其中每个轻链包含能够与CD40特异性结合的Fab片段的VL和CL结构域;以及
(c)能够与FAP特异性结合的交叉fab片段,其中所述交叉fab片段的VL-CH1链连接至(a)的两个重链中的一个的C末端。
在另一方面,提供了一种与FAP特异性结合的抗体,其中所述抗体包含:重链可变区(V
在进一步方面,提供了一种抗体,该抗体包含:(a)重链可变区(V
根据本发明的另一方面,提供了一种编码如上文所述的双特异性抗原结合分子的分离的核酸。另外提供了编码如上文所述的分离的核酸。本发明进一步提供了一种载体,特别是表达载体,其包含本发明的分离的核酸;并且提供了一种宿主细胞,其包含本发明的分离的核酸或表达载体。在一些方面,宿主细胞是真核细胞,特别是哺乳动物细胞。在另一方面,提供了一种生产如上文所述的双特异性抗原结合分子或抗体的方法,其包括在适合于表达双特异性抗原结合分子或抗体的条件下培养如上所述的宿主细胞,并且分离所述双特异性抗原结合分子或抗体。本发明还涵盖通过本发明的方法生产的与CD40和FAP特异性结合的双特异性抗原结合分子或与FAP特异性结合的抗体。
本发明进一步提供了一种药物组合物,其包含如上文所述的双特异性抗原结合分子或如上文所述的抗体以及药用运载体。在一个方面,药物组合物包含附加治疗剂。
另外,本发明涵盖如上文所述的双特异性抗原结合分子或抗体或包含该双特异性抗原结合分子的药物组合物,其用作药物。
在一个方面,提供了如上文所述的双特异性抗原结合分子或本发明的药物组合物,用于
(i)通过表达CD40的抗原呈递细胞(APC)诱导免疫刺激,
(ii)刺激肿瘤特异性T细胞应答,
(iii)引起肿瘤细胞凋亡,
(iv)治疗癌症,
(v)延缓癌症发展,
(vi)延长癌症患者的存活期,
(vii)治疗感染。
在一个具体方面,提供了如上文所述的双特异性抗原结合分子或本发明的药物组合物,其用于治疗癌症。在另一具体方面,本发明提供了如上文所述的双特异性抗原结合分子,其用于治疗癌症,其中双特异性抗原结合分子与化学治疗剂、放射和/或用于癌症免疫疗法的其他药剂联合施用。在一个方面,如本文所述的双特异性抗原结合分子用于治疗癌症,其中双特异性抗原结合分子用于与阻断PD-L1/PD-1相互作用的药剂联合施用。在另一方面,提供了如上文所述的双特异性抗原结合分子或本发明的药物组合物,用于上调或延长细胞毒性T细胞活性。在进一步方面,提供了如上文所述的抗体,其用于治疗癌症。
在进一步方面,本发明提供了一种抑制个体中肿瘤细胞生长的方法,其包括向个体施用有效量的如上文所述的双特异性抗原结合分子或本发明的药物组合物,以抑制肿瘤细胞的生长。在另一方面,本发明提供了一种治疗或延缓个体癌症的方法,其包括向个体施用有效量的如上文所述的双特异性抗原结合分子或本发明的药物组合物。
另外,提供了如上文所述的双特异性抗原结合分子在制备用于治疗有此需要的个体的疾病的药物中的用途,特别是在制造用于治疗癌症的药物中的用途,以及一种治疗个体的疾病的方法,其包括向所述个体施用治疗有效量的包含本发明的双特异性抗原结合分子的组合物,所述组合物呈药用形式。在一个特定方面,疾病为癌症。在上述方面中的任何方面,个体为哺乳动物,特别是人。
附图说明
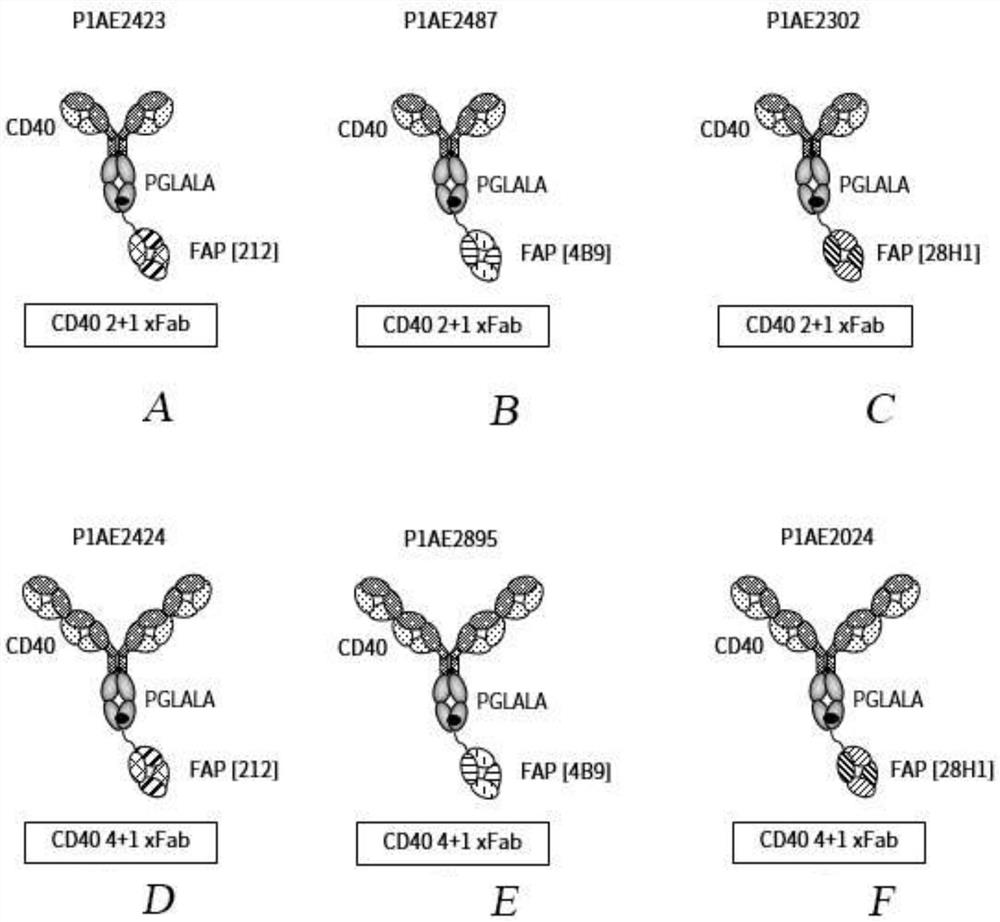
图1A至图F显示了与人CD40和FAP特异性结合的双特异性抗原结合分子的示意图。图1A显示了2+1形式的双特异性FAP-CD40抗体的示意图,其由与一个作为交叉fab片段的FAP(212)结合部分组合的两个CD40结合部分组成,其中VL-CH1链在Fc突起链的C末端融合(对于CD40为二价,而对于FAP为单价)。图1B显示了2+1形式的双特异性FAP-CD40抗体的示意图,其由与一个作为交叉fab片段的FAP(4B9)结合部分组合的两个CD40结合部分组成,其中VH-Cκ链在Fc突起链的C末端融合(对于CD40为二价,而对于FAP为单价)。图1C显示了2+1形式的双特异性FAP-CD40抗体的示意图,其由与一个作为交叉fab片段的FAP(28H1)结合部分组合的两个CD40结合部分组成,其中VL-CH1链在Fc突起链的C末端融合(对于CD40为二价,而对于FAP为单价)。图1D显示了4+1形式的双特异性FAP-CD40抗体的示意图,其由与一个作为交叉fab片段的FAP(212)结合部分组合的四个CD40结合部分组成,其中VL-CH1链在Fc突起链的C末端融合(对于CD40为四价,而对于FAP为单价)。图1E显示了4+1形式的双特异性FAP-CD40抗体的示意图,其由与一个作为交叉fab片段的FAP(4B9)结合部分组合的四个CD40结合部分组成,其中VL-CH1链在Fc突起链的C末端融合(对于CD40为四价,而对于FAP为单价)。图1F显示了4+1形式的双特异性FAP-CD40抗体的示意图,其由与一个作为交叉fab片段的FAP(28H1)结合部分组合的四个CD40结合部分组成,其中VL-CH1链在Fc突起链的C末端融合(对于CD40为四价,而对于FAP为单价)。黑点表示杵臼突变。替代性地,在所有分子中,crossfab的VH-Cκ链可融合至Fc突起链的C末端。
图2A和图2B显示了在与FAP克隆4B9和28H1竞争中,免疫接种衍生的FAP克隆与在转染的HEK细胞上表达的人FAP的细胞结合。图2A显示了所有测试的杂交瘤衍生的鼠克隆(命名为209、210、211、212、213、214、215、216、217和218)均没有竞争与抗FAP抗体4B9结合,并且图2B显示相同的克隆没有竞争与抗FAP抗体28H1结合。通过流式细胞术测量MFI。x轴显示了FAP抗体的浓度。
图3A和图3B显示了抗体构建体的示意图,该抗体构建体用于确定当抗FAP克隆经C末端融合至Fc结构域时是否不丧失其结合特性。图3A显示了包含Fc突起链和Fc孔链的构建体,其中VH结构域与Fc突起链的C末端融合,并且VL结构域与Fc孔链的C末端融合(C末端VH/VL融合)。图3B显示了包含Fc突起链和Fc孔链的构建体,其中整个Fab以其VH结构域与Fc突起链的C末端融合(C末端Fab融合)。图3C显示了使用基于Biacore T200仪器的表面等离子体共振(SPR)测定进行的表位分级的设置(参见实例1.9)。
图4显示了单价地靶向FAP(212)、FAP(4B9)或FAP(28H1)的人四价或二价抗CD40抗体与人FAP阳性NIH/3T3细胞的结合。转基因修饰的小鼠胚胎成纤维细胞NIH/3T3-hFAP细胞系表达高水平的人成纤维细胞活化蛋白(huFAP)。所有描绘的具有FAP结合部分的抗CD40抗原结合分子均与NIH/3T3-hFAP细胞有效结合,但与NIH/3T3-hFAP细胞的结合强度(EC
图5显示了单价地靶向FAP(212)或FAP(4B9)的人四价或二价抗CD40抗体与具有高表面表达水平的人CD40的原代人B细胞的结合。所有描绘的构建体均与CD40结合,但其与CD40阳性B细胞的结合强度(EC
图6A和6B显示了在存在FAP包被的(图6A)或未经包被的
图7A和7B显示了在存在FAP包被的(图7A)或未经包被的
图8A和图8B显示了在存在FAP(图8A)或不存在FAP(图8B)的情况下,通过靶向FAP的抗CD40结合分子活化的OVA脉冲的DC的T细胞引发。从huCD40转基因小鼠中分离的、用DEC205-OVA缀合物处理的、并受FAP依赖性双特异性抗CD40抗体以及FAP包被的珠粒刺激的DC诱导抗原特异性T细胞的强烈增殖。相反,在不存在FAP(未经包被的珠粒)的情况下,通过用靶向FAP的抗CD40抗体刺激的DC没有诱导T细胞增殖。通过用具有两个或四个CD40和一个FAP(212)或FAP(4B9)结合部分的人双特异性抗原结合分子刺激的DC诱导的T细胞增殖是相当的。用大量SIINFEKL而不是DEC205-OVA缀合物脉冲的DC诱导了强烈的T细胞增殖。显示了在存在OVA的情况下,用以指示的滴定抗体预孵育的huCD40 tg DC共培养的增殖(CFSE低)活CFSE标记的鼠CD3
图9显示了双特异性FAP x CD40抗体同时结合在实例5.3中所述的两个细胞系桥接试验中的作用。显示了不同形式的双特异性抗体的光密度(OD)与浓度的关系。
在图10中,显示了不同形式的FAP x CD40抗体激活CD40受体(与FAP无关)的潜力。在报告细胞系试验中,信号转导诱导NFκB依赖性产生分泌型胚胎碱性磷酸酶(SEAP),并且测量了SEAP的活性。相对于分子绘制n=6的光密度(OD)的平均值和3x STDEV。
图11显示了体内小鼠研究的研究设计,该研究用于在皮下MC38-FAP肿瘤模型中评估FAP靶向的抗CD40结合分子相比于非靶向抗CD40抗体的安全性、药代动力学和药效学特征。
图12显示了用指定的单抗体或联合抗体疗法治疗的小鼠在第1次和第2次(再激发)MC38-FAP肿瘤细胞注射后的肿瘤生长。箭头指示治疗注射和再激发的天数。y轴表示以mm
图13A为比较第28天(治疗后第6天)所有治疗组的统计表,图13B为比较第31天(治疗后第9天)所有治疗组的统计表。
图14显示了4+1和2+1FAP靶向抗CD40结合分子以及非靶向亲本抗CD40抗体在注射表达FAP的鼠结肠腺癌肿瘤细胞系(MC38-FAP)的小鼠中的药代动力学特征。y轴表示血清中的剂量归一化浓度,x轴表示注射抗体后的时间。观察到非靶向抗CD40抗体的清除率最高。与非靶向抗CD40分子相比,4+1形式的靶向FAP的抗CD40结合分子的清除率较低,并且观察到2+1形式的靶向FAP的抗CD40结合分子的清除率最低。
图15、16和17显示了接受表达FAP的鼠结肠腺癌肿瘤细胞系(MC38-FAP)注射并且单独接受非靶向抗CD40(P1AE2301)、抗CD40-FAP 4+1(P1AE2024)、抗CD40-FAP 2+1(P1AE2302)或媒介物节治疗的小鼠的引流淋巴结(LN,图15A、图16A和图17A)、非引流淋巴结(图15B、图16B和图17B)、脾脏(图15C、图16C和图17C)和肿瘤(图15D、图16D和图17D)中的DC、T细胞和B细胞活化。与接受媒介物治疗的动物相比,接受抗CD40-FAP 2+1(P1AE2302)治疗的动物的肿瘤中的DC和T细胞活化在注射治疗后四天显著增加(图15D和16D)。在所有其他分析的组织(引流LN、非引流LN和脾脏)中,与媒介物组相比,非靶向抗CD40、抗CD40-FAP4+1和抗CD40-FAP 2+1诱导了显著DC和T细胞活化。相比之下,与媒介物对照组相比,在所有分析的组织中仅非靶向抗CD40介导了显著的B细胞活化(图17A-17D,*p<0.05,**p<0.01,***p<0.001,****p<0.0001,非配对双尾学生检验)。
图18显示了注射表达FAP的鼠结肠腺癌肿瘤细胞系(MC38-FAP)并且接受与抗PD-L1抗体(P1AE0099)联合注射或不联合注射的非靶向抗CD40(P1AE2301)、抗CD40-FAP 4+1(P1AE2024)、抗CD40-FAP 2+1(P1AE2302)或媒介物治疗的小鼠的体重。y轴表示治疗前体重的百分比,x轴表示治疗注射后的天数。仅在单独接受非靶向抗CD40抗体或接受与抗PD-L1的联合治疗的小鼠中,观察到明显的体重下降。
具体实施方式
定义
除非另有定义,否则本文使用的所有技术术语和科技术语都具有如在本发明所属领域中通常使用的相同含义。出于解释本说明书的目的,将应用以下定义,并且在适当时,以单数形式使用的术语也将包括复数,反之亦然。
如本文所用,术语“抗原结合分子”在其最广义上是指特异性结合抗原决定簇的分子。抗原结合分子的示例是抗体、抗体片段和支架抗原结合蛋白。
如本文所用,术语“能够与靶细胞抗原特异性结合的抗原结合结构域”或“能够与靶细胞抗原特异性结合的部分”是指与抗原决定簇特异性结合的多肽分子。在一个方面,抗原结合结构域能够通过其靶细胞抗原活化信号传导。在一个特定方面,抗原结合结构域能够将与其附接的实体(例如CD40激动剂)引导至靶位点,例如引导至携带抗原决定簇的特定类型的肿瘤细胞或肿瘤基质。能够与靶细胞抗原特异性结合的抗原结合结构域包括如本文进一步定义的抗体及其片段。另外,能够与靶细胞抗原特异性结合的抗原结合结构域包括如本文进一步定义的支架抗原结合蛋白,例如基于设计的重复序列蛋白或设计的重复序列结构域的结合结构域(参见例如WO 2002/020565)。
关于抗体或其片段,术语“能够与靶细胞抗原特异性结合的抗原结合结构域”是指分子的一部分,其包含与抗原的一部分或全部特异性结合并且与之互补的区域。可通过例如一种或多种抗体可变结构域(也称为抗体可变区)来提供能够特异性抗原结合的抗原结合结构域。具体地,能够特异性抗原结合的抗原结合结构域包括抗体轻链可变区(VL)和抗体重链可变区(VH)。在另一方面,“能够与靶细胞抗原特异性结合的抗原结合结构域”也可以是Fab片段或交叉Fab片段。
本文的术语“抗体”以最广义使用并且涵盖各种抗体结构,包括但不限于单克隆抗体、多克隆抗体、单特异性和多特异性抗体(例如,双特异性抗体),以及抗体片段,只要它们表现出所需的抗原结合活性即可。
如本文所用的术语“单克隆抗体”是指从基本上同质的抗体群体获得的抗体,即,除了可能的变异抗体(例如含有天然存在的突变或在单克隆抗体制剂的生产过程中产生,此类变体通常以少量存在)之外,包含所述群体的各个抗体是相同的和/或结合相同的表位。与通常包括针对不同决定簇(表位)的不同抗体的多克隆抗体制剂相反,单克隆抗体制剂中的每种单克隆抗体针对抗原上的单一决定簇。
如本文所用,术语“单特异性”抗体表示具有一个或多个结合位点的抗体,每个结合位点与相同抗原的相同表位结合。术语“双特异性”意指抗原结合分子能够与至少两种独特的抗原决定簇特异性结合。通常,双特异性抗原结合分子包含两个抗原结合位点,这两个抗原结合位点中的每个抗原结合位点对不同的抗原决定簇具有特异性。在某些实施例中,双特异性抗原结合分子能够同时结合两种抗原决定簇,特别是在两种独特细胞上表达的两种抗原决定簇。如本文所述的双特异性抗原结合分子还可形成多特异性抗体的一部分。
本申请中所用的术语“价态”表示在对一种独特抗原决定簇具有特异性的抗原结合分子中存在特定数量的对一种独特抗原决定簇具有特异性的结合位点。因此,术语“二价”、“四价”和“六价”分别表示在抗原结合分子中存在对特定抗原决定簇具有特异性的两个结合位点、四个结合位点和六个结合位点。在本发明的特定方面,根据本发明的双特异性抗原结合分子对特定抗原决定簇可以是单价的,意味着它们对所述抗原决定簇仅具有一个结合位点,或者对特定抗原决定簇可以是二价或四价的,意味着它们对所述抗原决定簇分别具有两个结合位点或四个结合位点。
术语“全长抗体”、“完整抗体”和“全抗体”在本文中可互换使用地指代具有与天然抗体结构基本上相似的结构的抗体。“天然抗体”是指具有不同结构的天然免疫球蛋白分子。例如,天然IgG类抗体为约150,000道尔顿的异四聚体糖蛋白,由二硫键键合的两条轻链和两条重链组成。从N末端到C末端,每条重链具有可变区(VH)(也称为可变重链结构域或重链可变结构域),后接三个恒定结构域(CH1、CH2和CH3)(也称为重链恒定区)。类似地,从N末端到C末端,每条轻链具有可变区(VL)(也称为可变轻链结构域或轻链可变结构域),后接轻链恒定结构域(CL)(也称为轻链恒定区)。抗体的重链可以分配为五种类型中的一种,所述五种类型被称为α(IgA)、δ(IgD)、ε(IgE)、γ(IgG)或μ(IgM),它们中的一些可以进一步分为亚型,例如γ1(IgG1)、γ2(IgG2)、γ3(IgG3)、γ4(IgG4)、α1(IgA1)和α2(IgA2)。抗体的轻链基于其恒定结构域的氨基酸序列,可以归属于两种类型中的一种,这两种类型称为卡帕(κ)和兰姆达(λ)。
“抗体片段”是指完整抗体以外的分子,其包含完整抗体的一部分,所述部分结合完整抗体所结合的抗原。抗体片段的示例包括但不限于Fv、Fab、Fab'、Fab'-SH、F(ab')
木瓜蛋白酶消化完整抗体产生两个称为“Fab”片段的相同的抗原结合片段,每个“Fab”片段含有重链可变结构域和轻链可变结构域以及轻链的恒定结构域和重链的第一恒定结构域(CH1)。因此,如本文所用,术语“Fab片段”是指包含轻链片段以及重链的VH结构域和第一恒定结构域(CH1)的抗体片段,所述轻链片段包含VL结构域和轻链恒定结构域(CL)。Fab'片段与Fab片段的不同之处在于Fab'片段在重链CH1结构域的羧基末端添加了一些残基,这些残基包括来自抗体铰链区的一个或多个半胱氨酸。Fab’-SH是Fab’片段,其中恒定结构域的半胱氨酸残基具有游离巯基。胃蛋白酶处理产生F(ab')
术语“交叉Fab片段”或“xFab片段”或“交换型Fab片段”是指这样的Fab片段,其中重链和轻链的可变区或恒定区被交换。交换型Fab分子的两种不同链组成是可能的,并且包含在本发明的双特异性抗体中:在一个方面,Fab重链和轻链的可变区被交换,即交换型Fab分子包含由轻链可变区(VL)和重链恒定区(CH1)组成的肽链,以及由重链可变区(VH)和轻链恒定区(CL)组成的肽链。该交换型Fab分子也称为CrossFab
“单链Fab片段”或“scFab”是由抗体重链可变结构域(VH)、抗体恒定结构域1(CH1)、抗体轻链可变结构域(VL)、抗体轻链恒定结构域(CL)和接头组成的多肽,其中所述抗体结构域和所述接头在N-末端至C-末端方向上具有以下顺序中的一种:a)VH-CH1-接头-VL-CL,b)VL-CL-接头-VH-CH1,c)VH-CL-接头-VL-CH1,或d)VL-CH1-接头-VH-CL;并且其中所述接头是至少30个氨基酸,优选是32个至50个氨基酸的多肽。所述单链Fab片段经由CL结构域与CH1结构域之间的天然二硫键而稳定化。另外,这些单链Fab分子可以通过经由插入半胱氨酸残基(例如根据Kabat编号的可变重链中的44位和可变轻链中的100位)产生链间二硫键,而进一步稳定化。
“交换型单链Fab片段”或“x-scFab”是由抗体重链可变结构域(VH)、抗体恒定结构域1(CH1)、抗体轻链可变结构域(VL)、抗体轻链恒定结构域(CL)和接头组成的多肽,其中所述抗体结构域和所述接头在N末端至C末端方向上具有以下顺序中的一种:a)VH-CL-接头-VL-CH1和b)VL-CH1-接头-VH-CL;其中VH和VL一起形成与抗原特异性结合的抗原结合位点,并且其中所述接头是至少30个氨基酸的多肽。另外,这些x-scFab分子可以通过经由插入半胱氨酸残基(例如根据Kabat编号的可变重链中的44位和可变轻链中的100位)产生链间二硫键,而进一步稳定化。
“单链可变片段(scFv)”是抗体的重链可变区(V
“支架抗原结合蛋白”是本领域已知的,例如纤连蛋白和设计的锚蛋白重复序列蛋白(DARPin)已被用作抗原结合结构域的替代支架,参见例如Gebauer和Skerra,Engineeredprotein scaffolds as next-generation antibody therapeutics.Curr Opin ChemBiol 13:245-255(2009)和Stumpp等人,Darpins:A new generation of proteintherapeutics.Drug Discovery Today13:695-701(2008)。在本发明的一个方面中,支架抗原结合蛋白选自由以下项组成的组:CTLA-4(Evibody)、脂质运载蛋白(Anticalin)、蛋白A衍生的分子(诸如蛋白A的Z结构域(亲和体))、A结构域(Avimer/巨型抗体)、血清转铁蛋白(反式体);设计的锚蛋白重复序列蛋白(DARPin)、抗体轻链或重链的可变结构域(单结构域抗体,sdAb)、抗体重链的可变结构域(纳米抗体,aVH)、V
作为参考分子的“结合相同表位的抗原结合分子”是指这样的抗原结合分子,在竞争测定中所述抗原结合分子使参考分子与其抗原的结合被阻断50%或更多,并且相反地,在竞争测定中参考分子使抗原结合分子与其抗原的结合被阻断50%或更多。作为参考分子的“不结合相同表位的抗原结合分子”是指这样的抗原结合分子,在竞争测定中所述抗原结合分子不使参考分子与其抗原的结合被阻断50%或更多,并且相反地,在竞争测定中参考分子不使抗原结合分子与其抗原的结合被阻断50%或更多。
术语“抗原结合结构域”或“抗原结合位点”是指抗原结合分子的一部分,其包含与抗原的一部分或全部特异性结合并互补的区域。在抗原很大的情况下,抗原结合分子可以仅结合抗原的特定部分,其称为表位。抗原结合结构域可以由例如一个或多个可变结构域(也称为可变区)提供。优选地,抗原结合结构域包含抗体轻链可变区(VL)和抗体重链可变区(VH)。
如本文所用,术语“抗原决定簇”与“抗原”和“表位”同义,并且是指多肽大分子上的位点(例如一段连续的氨基酸或由非连续氨基酸的不同区域组成的构象构型),抗原结合部分与所述位点结合,从而形成抗原结合部分-抗原复合物。有用的抗原决定簇可以在例如肿瘤细胞的表面上、病毒感染细胞的表面上、其他患病细胞的表面上、免疫细胞的表面上、血清中的游离物和/或细胞外基质(ECM)中找到。除非另有说明,否则本文中用作抗原的蛋白质可以是来自任何脊椎动物来源的任何天然形式的蛋白质,该脊椎动物来源包括哺乳动物诸如灵长类动物(例如人类)和啮齿动物(例如小鼠和大鼠)。在一个具体实施例中,抗原是人蛋白质。当提及本文中的特定蛋白质时,该术语涵盖“全长”、未加工的蛋白质,以及由细胞内加工而产生的任何形式的蛋白质。该术语还涵盖天然存在的蛋白质变体,例如剪接变体或等位基因变体。
“特异性结合”是指结合对于抗原具有选择性,并且可以与不需要的或非特异性的相互作用区分开。抗原结合分子与特定抗原结合的能力可以通过酶联免疫吸附测定(ELISA)或本领域技术人员熟悉的其他技术(例如表面等离子体共振(SPR)技术(在BIAcore仪器上分析)(Liljeblad等人,Glyco J 17,323-329(2000))以及传统的结合测定(Heeley,Endocr Res28,217-229(2002))来测量。在一个实施例中,例如如通过SPR所测得的,抗原结合分子与不相关蛋白的结合程度小于所述抗原结合分子与抗原的结合程度的约10%。在某些实施例中,与抗原结合的分子的解离常数(Kd)为≤1μM、≤100nM、≤10nM、≤1nM、≤0.1nM、≤0.01nM或≤0.001nM(例如10
“亲和力”或“结合亲和力”是指分子(例如抗体)的单个结合位点与其结合配偶体(例如抗原)之间的非共价相互作用的总和的强度。除非另有说明,否则如本文所用的“结合亲和力”是指内在结合亲和力,其反映了结合对的成员(例如抗体和抗原)之间的1:1相互作用。分子X对其配偶体Y的亲和力通常可以用解离常数(Kd)表示,解离常数(Kd)是解离速率常数与缔合速率常数(分别为koff和kon)的比率。因此,等效亲和力可以包括不同的速率常数,只要速率常数的比率保持相同即可。亲和力可以通过本领域已知的常规方法测量,包括本文所述的那些方法。测量亲和力的特定方法是表面等离子体共振(SPR)。
“亲和力成熟的”的抗体是指在一个或多个高变区(HVR)中具有一个或多个改变的抗体,与不具有此类改变的亲本抗体相比,此类改变导致了抗体对抗原的亲和力的改善。
如本文所用,“靶细胞抗原”是指存在于靶细胞表面上的抗原决定簇,特别地肿瘤中的靶细胞(诸如癌细胞或肿瘤基质的细胞)。因此,靶细胞抗原是肿瘤相关抗原。特别地,肿瘤靶细胞抗原为成纤维细胞活化蛋白(FAP)。
术语“成纤维细胞活化蛋白(FAP)”也称为脯氨酰内肽酶FAP或Seprase(EC3.4.21),除非另有说明,否则该术语是指来自任何脊椎动物来源的任何天然FAP,该脊椎动物来源包括哺乳动物诸如灵长类动物(例如人)、非人灵长类动物(例如食蟹猴)和啮齿动物(例如小鼠和大鼠)。该术语包括“全长”的未加工FAP,以及通过细胞中加工产生的任何形式的FAP。该术语还涵盖FAP的天然存在变体,例如剪接变体或等位基因变体。在一个实施例中,本发明的抗原结合分子能够与人、小鼠和/或食蟹猴FAP特异性结合。人FAP的氨基酸序列示出于UniProt(www.uniprot.org)登录号Q12884(149版,SEQ ID NO:2)或NCBI(www.ncbi.nlm.nih.gov/)RefSeq NP_004451.2中。人FAP的细胞外结构域(ECD)从26位的氨基酸延伸至760位的氨基酸。His标记的人FAP ECD的氨基酸序列示出于SEQ ID NO 78中。小鼠FAP的氨基酸序列示出于UniProt登录号P97321(126版,SEQ ID NO:79)或NCBI RefSeqNP_032012.1中。小鼠FAP的细胞外结构域(ECD)从26位的氨基酸延伸至761位的氨基酸。SEQID NO:80示出His标记的小鼠FAP ECD的氨基酸。SEQ ID NO:81示出His标记的食蟹猴FAPECD的氨基酸。优选地,本发明的抗FAP结合分子结合到FAP的细胞外结构域。
术语“可变区”或“可变结构域”是指参与抗原结合分子与抗原结合的抗体重链或轻链的结构域。天然抗体的重链和轻链的可变结构域(分别为VH和VL)通常具有相似的结构,其中每个结构域包含四个保守框架区(FR)和三个高变区(HVR)。参见例如,Kindt等人,Kuby Immunology,第6版,W.H.Freeman and Co.,第91页(2007)。单个VH或VL结构域可足以赋予抗原结合特异性。
如本文所用,术语“高变区”或“HVR”是指抗体可变结构域中在序列上高变并确定抗原结合特异性的各个区域,例如“互补决定区”(“CDR”)。
通常,抗体包含六个CDR;三个在VH中(CDR-H1、CDR-H2、CDR-H3),并且三个在VL中的(CDR-L1、CDR-L2、CDR-L3)。本文中的示例性CDR包括:
(a)在氨基酸残基26-32(L1)、50-52(L2)、91-96(L3)、26-32(H1)、53-55(H2)和96-101(H3)处发生的高可变环(Chothia和Lesk,J.Mol.Biol.196:901-917(1987));
(b)存在于氨基酸残基24-34(L1)、50-56(L2)、89-97(L3)、31-35b(H1)、50-65(H2)和95-102(H3)处的CDR(Kabat等人,Sequences of Proteins of ImmunologicalInterest,第5版,Public Health Service,National Institutes of Health,Bethesda,MD(1991));以及
(c)存在于氨基酸残基27c-36(L1)、46-55(L2)、89-96(L3)、30-35b(H1)、47-58(H2)和93-101(H3)处的抗原接触点(MacCallum等人,J.Mol.Biol.262:732-745(1996))。
除非另有说明,否则CDR根据Kabat等人所述的方法(同上)确定。本领域技术人员将理解,也可以根据Chothia(同上)、McCallum(同上)所述的方法或任何其他在科学上接受的命名系统来确定CDR名称。
“框架”或“FR”是指除互补决定区(CDR)之外的可变结构域残基。可变结构域的FR通常由以下四个FR结构域组成:FR1、FR2、FR3和FR4。因此,CDR和FR序列通常在VH(或VL)中以如下序列出现:FR1-CDR-H1(CDR-L1)-FR2-CDR-H2(CDR-L2)-FR3-CDR-H3(CDR-L3)-FR4。
术语“嵌合”抗体是指这样的抗体,在所述抗体中重链和/或轻链的一部分来源于特定来源或物种,而重链和/或轻链的其余部分来源于不同的来源或物种。
抗体的“类别”是指抗体的重链所具有的恒定结构域或恒定区的类型。存在五大类抗体:IgA、IgD、IgE、IgG和IgM,并且这些类别中的若干可以进一步分为亚类(同种型),例如IgG
“人源化”抗体是指包含来自非人HVR的氨基酸残基和来自人FR的氨基酸残基的嵌合抗体。在某些实施例中,人源化抗体将基本上包含所有中的至少一个可变结构域,通常是两个可变结构域,其中所有或基本上所有HVR(例如CDR)对应于非人抗体的HVR,并且所有或基本上所有的FR对应于人抗体的FR。人源化抗体任选地可以包含来源于人抗体的抗体恒定区的至少一部分。“人源化形式”的抗体,例如非人抗体,是指已经经历进行过人源化的抗体。本发明涵盖的其他形式的“人源化抗体”是这样的抗体,相对于原始抗体,所述抗体中的恒定区已经经过了另外修饰或改变,以产生根据本发明的特性,特别是关于C1q结合和/或Fc受体(FcR)结合的特性。
本文中的术语“Fc结构域”或“Fc区”用于定义含有恒定区的至少一部分的抗体重链C-末端区域。该术语包括天然序列Fc区和变体Fc区。IgG Fc区包含IgG CH2结构域和IgGCH3结构域。人IgG Fc区的“CH2结构域”通常从大致231位的氨基酸残基延伸至大致340位的氨基酸残基。在一个实施例中,碳水化合物链附接至CH2结构域。本文的CH2结构域可以是天然序列CH2结构域或变体CH2结构域。“CH3结构域”包含Fc区中CH2结构域C-末端的一段残基(即,从IgG的约341位的氨基酸残基到约447位的氨基酸残基)。本文的CH3区可以是天然序列CH3结构域或变体CH3结构域(例如在一条链中具有引入的“凸起”(“突起”)而在另一条链中具有相应的引入的“空腔”(“孔”)的CH3结构域;参见以引用方式明确并入本文的美国专利5,821,333)。此类变体CH3结构域可用于促进如本文所述的两条不相同的抗体重链的异二聚化。在一个实施例中,人IgG重链Fc区从Cys226或从Pro230延伸至重链的羧基末端。然而,Fc区的C末端赖氨酸(Lys447)可以存在或不存在。除非本文另外规定,否则Fc区或恒定区中氨基酸残基的编号是根据EU编号系统,EU编号系统也称为EU索引,如在Kabat等人,Sequences of Proteins of Immunological Interest,第5版,Public Health Service,National Institutes of Health,Bethesda,MD,1991中所述。
“杵臼结构(knob-into-hole)”技术描述于例如US 5,731,168;US7,695,936;Ridgway等人,Prot Eng 9,617-621(1996)和Carter,J Immunol Meth 248,7-15(2001)中。通常,该方法涉及在第一多肽的界面处引入凸起(“突起”)并在第二多肽的界面中引入相应的空腔(“孔”),使得所述凸起可以定位在所述空腔中,以便促进异二聚体的形成并阻碍同二聚体的形成。凸起是通过用较大侧链(例如酪氨酸或色氨酸)取代来自第一多肽的界面的小氨基酸侧链而构建的。具有与凸起相同或相似大小的补偿空腔是通过用较小的氨基酸侧链(例如丙氨酸或苏氨酸)取代大氨基酸侧链而在第二多肽的界面中创建的。凸起和空腔可以通过改变编码多肽的核酸来制备,例如通过位点特异性诱变或通过肽合成。在一个具体实施例中,突起修饰包含Fc结构域的两个亚基中的一个中的氨基酸取代T366W,而孔修饰包含Fc结构域的两个亚基中的另一个中的氨基酸取代T366S、L368A和Y407V。在另一个具体实施例中,包含突起修饰的Fc结构域的亚基另外包含氨基酸取代S354C,而包含孔修饰的Fc结构域的亚基另外包含氨基酸取代Y349C。引入这两个半胱氨酸残基导致在Fc区的两个亚基之间形成二硫桥,从而进一步稳定化二聚体(Carter,J Immunol Methods 248,7-15(2001))。
“与免疫球蛋白的Fc区等同的区域”旨在包括免疫球蛋白的Fc区的天然存在的等位基因变体,以及具有产生取代、添加或缺失但基本上不降低免疫球蛋白介导效应子功能(诸如抗体依赖性细胞毒性)的能力的修改的变体。例如,可以使一个或多个氨基酸从免疫球蛋白的Fc区的N-末端或C-末端缺失,而基本上不丧失生物学功能。可以根据本领域中已知的一般规则来选择此类变体,以便对活性具有最小的影响(参见例如Bowie,J.U.等人,Science 247:1306-10(1990))。
术语“效应子功能”是指可归因于抗体的Fc区、随着抗体同种型而变化的那些生物学活性。抗体效应子功能的示例包括:C1q结合和补体依赖性细胞毒性(CDC)、Fc受体结合、抗体依赖性细胞介导的细胞毒性(ADCC)、抗体依赖性细胞吞噬作用(ADCP)、细胞因子分泌、免疫复合物介导的抗原呈递细胞的抗原摄取、下调细胞表面受体(例如B细胞受体),以及B细胞活化。
Fc受体结合依赖性效应子功能可通过抗体的Fc区与Fc受体(FcR)的相互作用来介导,Fc受体是造血细胞上的特异性细胞表面受体。Fc受体属于免疫球蛋白超家族,并且已被证明可通过吞噬免疫复合物来介导去除抗体包被的病原体,并且通过抗体依赖性细胞介导的细胞毒性(ADCC)来裂解涂有相应抗体的红细胞及其他各种细胞靶点(例如,肿瘤细胞)(参见例如Van de Winkel,J.G.和Anderson,C.L.,J.Leukoc.Biol.49(1991)511-524)。FcRs由其对免疫球蛋白同种型的特异性来限定:IgG抗体的Fc受体称为FcγR。Fc受体结合描述于例如:Ravetch,J.V.和Kinet,J.P.,Annu.修订版,Immunol.9(1991)457-492;Capel,P.J.等人,Immunomethods 4(1994)25-34;de Haas,M.等人,J.Lab.Clin.Med.126(1995)330-341;以及Gessner,J.E.等人,Ann.Hematol.76(1998)231-248。
IgG抗体(FcγR)Fc区受体的交联触发多种效应子功能,包括吞噬作用、抗体依赖性细胞细胞毒性、炎症介质的释放以及免疫复合物清除和抗体产生的调节。已在人体中鉴定出三类FcγR,其中包括:
FcγRI(CD64)以高亲和力结合单体IgG,并且在巨噬细胞、单核细胞、中性粒细胞和嗜酸性粒细胞上表达。Fc区IgG中在至少一个氨基酸残基E233-G236、P238、D265、N297、A327和P329(根据Kabat的EU索引编号)的修饰,降低与FcγRI的结合。在233-236位的IgG2残基被IgG1和IgG4取代,使得与FcγRI的结合力降低10
FcγRII(CD32)以中等至低亲和力结合复合IgG,并得到广泛表达。该受体可分为两种亚型,即FcγRIIA和FcγRIIB。FcγRIIA存在于许多参与杀伤的细胞(例如巨噬细胞、单核细胞、中性粒细胞)中,并且似乎能够活化杀伤过程。FcγRIIB似乎在抑制过程中起作用,并且存在于B细胞、巨噬细胞以及肥大细胞和嗜酸性粒细胞中。在B细胞上,它似乎起到抑制免疫球蛋白进一步产生以及同种型转换为例如IgE类的作用。在巨噬细胞上,FcγRIIB用于抑制由FcγRIIA介导的吞噬作用。在嗜酸性粒细胞和肥大细胞上,B型可能通过IgE与其单独受体的结合而有助于抑制这些细胞的活化。发现例如抗体(包含在至少一个氨基酸残基E233-G236、P238、D265、N297、A327、P329、D270、Q295、A327、R292和K414(根据Kabat的EU索引编号)的突变的IgG Fc区)对FcγRIIA的结合力降低。
FcγRIII(CD16)以中等至低亲和力结合IgG,并且包括两种类型。Fc RIIIA存在于NK细胞、巨噬细胞、嗜酸性粒细胞以及一些单核细胞和T细胞上,并且介导ADCC。FcγRIIIB在中性粒细胞上的表达水平高。发现例如抗体(包含在至少一个氨基酸残基E233-G236、P238、D265、N297、A327、P329、D270、Q295、A327、S239、E269、E293、Y296、V303、A327、K338和D376(根据Kabat的EU索引编号)的突变的IgG Fc区)对Fc RIIIA的结合力降低。
Shields,R.L.等人(J.Biol.Chem.276(2001)6591-6604)描述了人IgG1上与Fc受体的结合位点的定位、上述突变位点以及测量与FcγRI和FcγRIIA结合的方法。
术语“ADCC”或“抗体依赖性细胞毒性”是由Fc受体结合介导的功能,并且是指在效应子细胞存在下本文所报道的抗体对靶细胞的裂解。通过测量抗体与表达Fcγ受体的细胞(诸如重组表达FcγRI和/或FcγRIIA或NK细胞(在本质上表达FcγRIIIA)的细胞)的结合来研究抗体诱导介导ADCC的初始步骤的能力。具体地,测量与NK细胞上与FcγR的结合。
“活化Fc受体”是这样的Fc受体,其在抗体的Fc区接合后,引起刺激携带受体的细胞执行效应子功能的信号传导事件。活化Fc受体包括FcγRIIIa(CD16a)、FcγRI(CD64)、FcγRIIa(CD32)和FcαRI(CD89)。特定的活化Fc受体是人FcγRⅢa(参见UniProt登录号P08637,版本141)。
除非另外指明,否则如本文所用的术语“CD40”是指来自任何脊椎动物来源的任何天然CD40,该脊椎动物来源包括哺乳动物诸如灵长类动物(例如人)和啮齿动物(例如小鼠和大鼠)。该术语包括“全长”的未加工CD40,以及通过细胞中加工产生的任何形式的CD40。该术语还涵盖CD40的天然存在变体,例如剪接变体或等位基因变体。示例性人CD40的氨基酸序列示出于SEQ ID NO:1(Uniprot P25942,版本200)中,并且示例性小鼠CD40的氨基酸序列示出于SEQ ID NO:146(Uniprot P27512,版本160)中。CD40抗原是50kDa细胞表面糖蛋白,其属于肿瘤坏死因子受体(TNF-R)家族。(Stamenkovic等人(1989),EMBO J.8:1403-10)。CD40在许多正常和肿瘤细胞类型中表达,其包括B淋巴细胞、树突细胞、单核细胞、巨噬细胞、胸腺上皮细胞、内皮细胞、成纤维细胞和平滑肌细胞。CD40在所有B淋巴瘤中和70%的所有实体瘤中表达,并通过成熟信号(诸如IFN-γ和GM-CSF)在抗原呈递细胞(APC)中上调。CD40活化还诱导单核细胞分化为功能性树突细胞(DC),并通过APC-CD40诱导的细胞因子增强NK细胞的溶细胞活性。因此,CD40通过诱导APC的成熟,辅助细胞因子的分泌,共刺激分子的上调以及效应子功能的增强,在引发和增强免疫应答中起着至关重要的作用。
如本文所用,术语“CD40激动剂”包括使CD40/CD40L相互作用激动的任何部分。如上下文所用,CD40优选是指人CD40,因此CD40激动剂优选为人CD40的激动剂。通常,该部分为激动性CD40抗体或抗体片段。
术语“抗CD40抗体”、“抗CD40”、“CD40抗体”和“与CD40特异性结合的抗体”是指能够以足够亲和力结合CD40的抗体,以便该抗体用作在靶向CD40中的诊断剂和/或治疗剂。在一个方面,例如通过放射免疫测定法(RIA)或流式细胞术(FACS)所测得的,抗CD40抗体与不相关的非CD40蛋白的结合程度小于该抗体与CD40结合程度的约10%。在某些实施例中,与CD40结合的抗体的解离常数(K
术语“肽接头”是指包含一个或多个氨基酸,通常约2至20个氨基酸的肽。肽接头是本领域中已知的或在本文中描述的。合适的非免疫原性连接肽是例如(G
如本申请中所用的术语“氨基酸”表示包括以下项的天然存在的羧基α-氨基酸的组:丙氨酸(三字母代码:ala,单字母代码:A)、精氨酸(arg,R)、天冬酰胺(asn,N)、天冬氨酸(asp,D)、半胱氨酸(cys,C)、谷氨酰胺(gln,Q)、谷氨酸(glu,E)、甘氨酸(gly,G)、组氨酸(his,H)、异亮氨酸(ile,I)、亮氨酸(leu,L)、赖氨酸(lys,K)、蛋氨酸(met,M)、苯丙氨酸(phe,F)、脯氨酸(pro,P)、丝氨酸(ser,S)、苏氨酸(thr,T)、色氨酸(trp,W)、酪氨酸(tyr,Y),以及缬氨酸(val,V)。
“融合”或“连接”意指组分(例如,抗体和Fab片段的重链)直接地或经由一个或多个肽接头而通过肽键连接。
相对于参考多肽(蛋白质)序列的“氨基酸序列一致性百分比(%)”被定义为在比对候选序列中的氨基酸残基与参考多肽序列中的氨基酸残基并引入缺口(如果需要的话)以实现最大的序列一致性百分比后,并且在不将任何保守取代考虑为所述序列一致性的一部分的情况下,与所述参考多肽序列中的氨基酸残基相同的所述候选序列中的氨基酸残基的百分比。用于确定氨基酸序列一致性百分比的比对可以以本领域技术范围内的各种方式实现,例如使用公众可获得的计算机软件,诸如BLAST、BLAST-2、ALIGN.SAWI或Megalign(DNASTAR)软件。本领域技术人员可确定用于比对序列的适当参数,包括在所比较的序列的全长上实现最大比对所需的任何算法。然而,为了本文的目的,使用序列比较计算机程序ALIGN-2来生成氨基酸序列一致性%的值。ALIGN-2序列比较计算机程序由Genentech,Inc.编写,并且源代码已经与用户文档一起提交到U.S.Copyright Office,Washington D.C.,20559,在那里以美国版权登记号TXU510087注册。ALIGN-2程序可从Genentech,Inc.,SouthSan Francisco,California公开获得,或者可以从所述源代码编译。ALIGN-2程序应经编译以在UNIX操作系统上使用,所述UNIX操作系统包括数字UNIX V4.0D。所有序列比较参数均由ALIGN-2程序设置并且不变。在采用ALIGN-2进行氨基酸序列比较的情况下,给定氨基酸序列A与给定氨基酸序列B的氨基酸序列一致性%(其可以替代地表达为给定氨基酸序列A具有或包含与给定氨基酸序列B的某一氨基酸序列一致性%)计算如下:
100乘以分数X/Y
其中X是由序列比对程序ALIGN-2在该程序对A和B的比对中评分为相同匹配的氨基酸残基的数目,而其中Y是B中氨基酸残基的总数。应当理解,在氨基酸序列A的长度不等于氨基酸序列B的长度的情况下,A与B的氨基酸序列一致性%将不等于B与A的氨基酸序列一致性%。除非另外特别指明,否则本文所使用的所有氨基酸序列一致性%的值是如前一段中所述使用ALIGN-2计算机程序获得的。
在某些实施例中,考虑了本文提供的双特异性抗原结合分子的氨基酸序列变体。例如,可能需要改善含TNF配体三聚体的抗原结合分子的结合亲和力和/或其他生物学特性。含TNF配体三聚体的抗原结合分子的氨基酸序列变体可以通过向编码分子的核苷酸序列中引入适当的修饰或通过肽合成来制备。此类修饰包括例如抗体氨基酸序列内残基的缺失、和/或插入和/或取代。可以进行缺失、插入和取代的任何组合以实现最终构建体,前提条件是最终构建体具有所需特征,例如抗原结合。用于取代诱变的感兴趣的位点包括HVR和框架(FR)。保守性取代在表B中表头“优选的取代”下提供,并在下文中参考氨基酸侧链类别(1)至(6)进一步描述。可以将氨基酸取代引入感兴趣的分子中,并对产物进行所需活性(例如保留/改善的抗原结合、降低的免疫原性,或改善的ADCC或CDC)筛选。
表A
可根据共同的侧链特性将氨基酸分组:
(1)疏水性;正亮氨酸、Met、Ala、Val、Leu、Ile;
(2)中性亲水性:Cys、Ser、Thr、Asn、Gln;
(3)酸性:Asp、Glu;
(4)碱性:His、Lys、Arg;
(5)影响链取向的残基:Gly、Pro;
(6)芳香族:Trp、Tyr、Phe。
非保守性取代将需要用这些类别中的一个的成员交换另一类别。
术语“氨基酸序列变体”包括实质性变体,其中在亲本抗原结合分子(例如人源化或人抗体)的一个或多个高变区残基中存在氨基酸取代。通常,相对于亲本抗原结合分子,选为用于进一步研究的一个或多个所得变体将在某些生物学特性方面(例如,亲和力增加、免疫原性降低)有改变(例如,改善)和/或将基本上保留亲本抗原结合分子的某些生物学特性。示例性取代变体是亲和力成熟抗体,其可例如使用诸如本文所述的那些基于噬菌体展示的亲和力成熟技术方便地生成。简而言之,将一个或多个HVR残基突变并且将变体抗原结合分子展示在噬菌体上并针对特定生物活性(例如结合亲和力)进行筛选。在某些实施例中,取代、插入或缺失可发生在一个或多个HVR内,只要此类改变基本上不降低抗原结合分子的抗原结合能力即可。例如,可在HVR中进行基本上不降低结合亲和力的保守性改变(例如,如本文提供的保守性取代)。可用于鉴别可被靶向诱变的抗体残基或区域的方法称作“丙氨酸扫描诱变”,如Cunningham和Wells(1989)Science,244:1081-1085所述。在此方法中,鉴别残基或一组靶残基(例如,带电残基,诸如Arg、Asp、His、Lys和Glu)并用中性或带负电的氨基酸(例如,丙氨酸或多丙氨酸)替换以确定抗体与抗原的相互作用是否受到影响。可在对初始取代展现功能敏感性的氨基酸位置引入其他取代。另选地或另外地,利用抗原-抗原结合分子复合物的晶体结构鉴别抗体与抗原之间的接触点。可靶向或消除作为取代的候选的此类接触残基和相邻残基。可筛选变体以确定它们是否具备期望的特性。
氨基酸序列插入包括长度范围为一个残基至含有一百个或更多个残基的多肽的氨基和/或羧基末端融合,以及一个或多个氨基酸残基的序列内插入。末端插入的示例包括具有N末端甲硫氨酰残基的本发明的双特异性抗原结合分子。分子的其他插入变体包括与增加双特异性抗原结合分子的血清半衰期的多肽的N末端或C末端的融合。
在某些实施例中,改变本文提供的双特异性抗原结合分子以增加或降低抗体糖基化的程度。可通过改变氨基酸序列,使得产生或去除一个或多个糖基化位点,从而便利地获得分子的糖基化变体。当含TNF配体三聚体的抗原结合分子包含Fc区时,附接于其上的碳水化合物可以被改变。由哺乳动物细胞产生的天然抗体通常包含具有支链的双触角寡糖,所述双触角寡糖通常通过N-连接附接于Fc区的CH2结构域的Asn297。参见,例如,Wright等人TIBTECH 15:26-32(1997)。寡糖可包括各种碳水化合物,例如,甘露糖、N-乙酰基葡糖胺(GlcNAc)、半乳糖和唾液酸,以及附接于双触角寡糖结构的“主干”中的GlcNAc的岩藻糖。在一些实施例中,可以对含TNF家族配体三聚体的抗原结合分子中的寡糖进行修饰,以产生具有某些改善的特性的变体。在一个方面,提供了本发明的双特异性抗原结合分子或抗体的变体,其具有缺乏附接(直接或间接)于Fc区的岩藻糖的碳水化合物结构。此类岩藻糖基化变体可具有改善的ADCC功能,参见例如美国专利公开文本No.US 2003/0157108(Presta,L.)或US2004/0093621(Kyowa Hakko Kogyo Co.,Ltd)。在另一方面,本发明的双特异性抗原结合分子或抗体提供有两分型寡糖的变体,例如其中附接于Fc区的双触角寡糖是通过GlcNAc两分的。此类变体可具有降低的岩藻糖基化和/或改善的ADCC功能,参见例如WO2003/011878(Jean-Mairet等人);美国专利号6,602,684(Umana等人);以及US 2005/0123546(Umana等人)。还提供了在附接于Fc区的寡糖中具有至少一个半乳糖残基的变体。此类抗体变体可具有改善的CDC功能并描述于例如WO1997/30087(Patel等人);WO 1998/58964(Raju,S.);以及WO1999/22764(Raju,S.)中。
在某些方面,可期望产生本发明的双特异性抗原结合分子的半胱氨酸工程化改造的变体,例如“thioMAb”,其中分子的一个或多个残基被半胱氨酸残基取代。在特定方面,取代的残基存在于分子的可接近位点。通过用半胱氨酸取代那些残基,反应性硫醇基团由此定位于抗体的可接近位点,并且可用于将抗体与其他部分,诸如药物部分或接头-药物部分缀合,以产生免疫缀合物。在某些方面,可用半胱氨酸取代下列残基中的任何一个或多个:轻链的V205(Kabat编号);重链的A118(EU编号);以及重链Fc区的S400(EU编号)。可如例如美国专利号7,521,541中所述形成半胱氨酸工程化改造的抗原结合分子。
术语“核酸”或“多核苷酸”包括包含核苷酸聚合物的任何化合物和/或物质。每个核苷酸由碱基组成,特别是嘌呤或嘧啶碱基(即胞嘧啶(C)、鸟嘌呤(G)、腺嘌呤(A)、胸腺嘧啶(T)或尿嘧啶(U))、糖(即脱氧核糖或核糖)和磷酸基团。通常,核酸分子通过碱基序列进行描述,由此所述碱基代表核酸分子的一级结构(线性结构)。碱基序列通常表示为从5'至3'。在本文中,术语核酸分子涵盖脱氧核糖核酸(DNA)(包括例如互补DNA(cDNA)和基因组DNA)、核糖核酸(RNA)(特别是信使RNA(mRNA))、DNA或RNA的合成形式,以及包含这些分子中的两种或更多种的混合聚合物。核酸分子可以是线性的或环状的。另外,术语核酸分子包括有义链和反义链,以及单链和双链形式。此外,本文所描述的核酸分子可含有天然存在的或非天然存在的核苷酸。非天然存在的核苷酸的示例包括具有衍生化的糖或磷酸主链键或经化学修饰的残基的经修饰的核苷酸碱基。核酸分子还涵盖适合作为用于本发明的抗体的体外和/或体内(例如,在宿主或患者体内)直接表达的载体的DNA和RNA分子。此类DNA(例如cDNA)或RNA(例如mRNA)载体可以是未修饰的或经修饰的。例如,可以对mRNA进行化学修饰以增强RNA载体的稳定性和/或编码分子的表达,使得可以将mRNA注射到受试者体内以产生体内抗体(参见例如Stadler等人,Nature Medicine 2017,在线发表于2017年6月12日,doi:10.1038/nm.4356或EP 2 101 823B1)。
“分离的”核酸是指已从其天然环境的组分中分开的核酸分子。分离的核酸包括这样的核酸分子,其包含在通常含有所述核酸分子的细胞中,但所述核酸分子存在于染色体外或与其天然染色体位置不同的染色体位置处。
“编码双特异性抗原结合分子或抗体的分离的核酸”是指编码双特异性抗原结合分子或抗体重链和轻链(或其片段)的一种或多种核酸分子,包括在单个载体或单独载体中的此类核酸分子,以及存在于宿主细胞中的一个或多个位置处的此类核酸分子。
关于与本发明的参考核苷酸序列具有至少例如95%“相同”的核苷酸序列的核酸或多核苷酸,是指除了多核苷酸序列可包括参考核苷酸序列的每100个核苷酸至多五个点突变之外,多核苷酸的核苷酸序列与参考序列是相同的。换句话讲,为了获得具有与参考核苷酸序列至少95%相同的核苷酸序列的多核苷酸,参考序列中至多5%的核苷酸可缺失或被另外的核苷酸取代,或参考序列中总核苷酸的至多5%的数量的核苷酸可插入到参考序列中。参考序列的这些改变可发生在参考核苷酸序列的5’或3’末端位置或那些末端位置之间的任意位置,或单个地散布在参考序列的残基之中,或以一个或多个连续的组散布在参考序列内。作为一种实际情况,可以使用已知的计算机程序,诸如上文针对多肽所讨论的程序(例如ALIGN-2),常规确定任何特定多核苷酸序列是否与本发明的核苷酸序列至少80%、85%、90%、95%、96%、97%、98%或99%相同。
术语“表达盒”是指通过重组或合成生成的多核苷酸,其具有允许特定核酸在靶细胞中转录的一系列特定核酸元件。重组表达盒可以掺入质粒、染色体、线粒体DNA、质粒DNA、病毒或核酸片段中。典型地,表达载体的重组表达盒部分除其他序列之外还包括待转录的核酸序列和启动子。在某些实施例中,本发明的表达盒包含编码本发明的双特异性抗原结合分子或其片段的多核苷酸序列。
术语“载体”或“表达载体”与“表达构建体”是同义的并且是指用于将特定基因引入与其可操作地关联的靶细胞中并指导所述基因的表达的DNA分子。该术语包括作为自我复制核酸结构的载体,以及整合入其已被引入的宿主细胞的基因组中的载体。本发明的表达载体包含表达盒。表达载体允许大量稳定mRNA的转录。一旦表达载体处于靶细胞内部,即通过细胞转录和/或翻译机制产生由该基因编码的核糖核酸分子或蛋白质。在一个实施例中,本发明的表达载体包含表达盒,所述表达盒包含编码本发明的双特异性抗原结合分子或其片段的多核苷酸序列。
术语“宿主细胞”“宿主细胞系”和“宿主细胞培养物”可互换使用,并且是指外源核酸已被引入其中的细胞,包括此类细胞的子代。宿主细胞包括“转化体”和“转化细胞”,其包括原代转化细胞和来源于所述原代转化细胞的子代,不考虑传代次数。子代可能不与亲本细胞的核酸内容物完全一致,而是可能含有突变。本文包括如在原始转化细胞中筛选或选择的具有相同功能或生物活性的突变子代。宿主细胞是可以用于生成本发明的双特异性抗原结合分子的任何类型的细胞系统。宿主细胞包括培养的细胞,举几个例子,例如培养的哺乳动物细胞,诸如CHO细胞、BHK细胞、NS0细胞、SP2/0细胞、YO骨髓瘤细胞、P3X63小鼠骨髓瘤细胞、PER细胞、PER.C6细胞或杂交瘤细胞、酵母细胞、昆虫细胞和植物细胞,以及包括在转基因动物、转基因植物或培养的植物或动物组织中的细胞。
药剂的“有效量”是指在其所施用的细胞或组织中产生生理学变化所需的量。
药剂(例如药物组合物)的“治疗有效量”是指在必需的剂量和时段上有效实现期望的治疗或预防结果的量。治疗有效量的药剂例如消除、减少、延迟、最小化或预防疾病的不良影响。
“个体”或“受试者”是哺乳动物。哺乳动物包括但不限于驯养的动物(例如牛、绵羊、猫、犬和马)、灵长类动物(例如人和非人灵长类动物,诸如猴)、兔以及啮齿类动物(例如小鼠和大鼠)。具体地,个体或受试者是人。
术语“药物组合物”或“药物制剂”是指处于允许包含在其中的活性成分的生物活性有效的形式,并且不含对于将被施用药物组合物的受试者具有不可接受的毒性的另外组分的制剂。
“药用运载体”是指药物组合物或制剂中除活性成分外的对受试者无毒的成分。药学上可接受的运载体包括但不限于缓冲剂、赋形剂、稳定剂,或防腐剂。
术语“包装插页”用于指治疗产品的商业包装中通常包括的说明书,其含有涉及此类治疗产品的使用的有关适应症、用法、剂量、施用、联合疗法、禁忌症和/或警告的信息。
如本文所用,“治疗(treatment)”(及其语法变型,诸如“治疗(treat)”或“治疗(treating)”)是指试图改变所治疗个体的自然进程的临床干预,并且可以是为了预防或在临床病理学的进程中进行。治疗的期望效果包括但不限于预防疾病的发生或复发、减轻症状、削弱疾病的任何直接或间接病理学后果、预防转移、降低疾病进展的速率、改善或减轻疾病状态,以及缓解或改善预后。在一些实施例中,本发明的分子用于延迟疾病的发展或用于减缓疾病的进展。
如本文所用的术语“癌症”是指增生性疾病,诸如淋巴瘤、淋巴细胞性白血病、肺癌、非小细胞肺(NSCL)癌、支气管肺泡细胞肺癌、骨癌、胰腺癌、皮肤癌、头或颈癌、皮肤或眼内黑素瘤、子宫癌、卵巢癌、直肠癌、肛区癌、胃癌(stomach cancer)、胃癌(gastriccancer)、结肠癌、乳腺癌、子宫癌、输卵管癌、子宫内膜癌、子宫颈癌、阴道癌、外阴癌、霍奇金病、食管癌、小肠癌、内分泌系统癌、甲状腺癌、甲状旁腺癌、肾上腺癌、软组织肉瘤、尿道癌、阴茎癌、前列腺癌、膀胱癌、肾癌或输尿管癌、肾细胞癌、肾盂癌、间皮瘤、肝细胞癌、胆管癌、中枢神经系统(CNS)肿瘤、脊椎轴肿瘤、脑干胶质瘤、多形性成胶质细胞瘤、星形细胞瘤、神经鞘瘤、室管膜瘤、成髓细胞瘤、脑膜瘤、鳞状细胞癌、垂体腺瘤和尤文氏肉瘤,包括以上癌症中的任一种的难治性型式,或一种或多种以上癌症的组合。
如本文所用,术语“化学治疗剂”是指可用于治疗癌症的化合物。在一个方面,化学治疗剂为抗代谢物。在一个方面,抗代谢物选自由以下项组成的组:氨基蝶呤、甲氨蝶呤、培美曲塞、雷替曲塞、克拉屈滨、氯法拉滨、氟达拉滨、巯基嘌呤、喷司他丁、硫鸟嘌呤、卡培他滨、阿糖胞苷、氟尿嘧啶、氟尿苷和吉西他滨。在一个特定方面,抗代谢物为卡培他滨或吉西他滨。在另一方面,抗代谢物为氟尿嘧啶。在一个方面,化学治疗剂是影响微管形成的药剂。在一个方面,影响微管形成的药剂选自由以下项组成的组:紫杉醇、多西他赛、长春新碱、长春碱、长春地辛、长春瑞滨、泰素帝、依托泊苷和替尼泊苷。在另一方面,化学治疗剂为烷化剂诸如环磷酰胺。在一个方面,化学治疗剂为细胞毒性抗生素,诸如拓扑异构酶II抑制剂。在一个方面,拓扑异构酶II抑制剂为多柔比星。
本发明的双特异性抗体
本发明提供了双特异性抗原结合分子,这些双特异性抗原结合分子包含新型抗FAP抗体(克隆212)。包含该新型抗FAP抗体的双特异性抗原结合分子具有特别有利的特性,诸如可生产性、稳定性、结合亲和力、生物学活性、靶向效率、减少的内化、优异的药代动力学(PK)特性、降低的毒性、可以给予患者的扩展剂量范围以及从而可能增强的功效。
示例性双特异性抗原结合分子
在一个方面,本发明提供了双特异性抗原结合分子,其特征在于与CD40的靶向激动性结合。特别地,双特异性抗原结合分子为靶向FAP的CD40激动剂。在另一特定方面,本发明的双特异性抗原结合分子包含由能够稳定缔合的第一亚基和第二亚基组成的Fc区,其包含降低效应子功能的突变。使用包含减少或消除效应子功能的突变的Fc区将通过经由Fc受体进行交联来防止非特异性激动作用,并防止CD40
因此,这些双特异性抗原结合分子的特征在于与CD40的靶向FAP的激动性结合。在存在表达FAP的细胞时,双特异性抗原结合分子能够活化抗原呈递细胞(APC),活化人B细胞(实例5.1.2)、人Daudi细胞(实例5.1.1)和人单核细胞衍生的树突细胞(moDC)。
在一个方面,提供了一种双特异性抗原结合分子,其包含:
(a)至少一个能够与CD40特异性结合的抗原结合结构域;以及
(b)至少一个能够与成纤维细胞活化蛋白(FAP)特异性结合的抗原结合结构域,其包含:重链可变区(V
(i)CDR-H1,其包含SEQ ID NO:3的氨基酸序列,(ii)CDR-H2,其包含选自由SEQ IDNO:4、SEQ ID NO:11和SEQ ID NO:12组成的组的氨基酸序列,和(iii)CDR-H3,其包含SEQID NO:5的氨基酸序列;以及轻链可变区(V
(vi)CDR-L3,其包含SEQ ID NO:8的氨基酸序列。
在某一特定方面,提供了一种双特异性抗原结合分子,其包含:a)至少一个能够与CD40特异性结合的抗原结合结构域;以及(b)至少一个能够与成纤维细胞活化蛋白(FAP)特异性结合的抗原结合结构域,该FAP包含:重链可变区(V
在一个方面,提供了一种双特异性抗原结合分子,其中能够与FAP特异性结合的抗原结合结构域包含:重链可变区(V
在另一方面,提供了一种双特异性抗原结合分子,其中能够与FAP特异性结合的抗原结合结构域包含:
重链可变区(V
在一个方面,双特异性抗原结合分子包含
(a)重链可变区(V
(b)重链可变区(V
(c)重链可变区(V
(d)重链可变区(V
在某一特定方面,双特异性抗原结合分子包含:重链可变区(V
在另一方面,提供了一种双特异性抗原结合分子,其中能够与CD40特异性结合的抗原结合结构域包含:重链可变区(V
在进一步方面,提供了一种双特异性抗原结合分子,其中能够与CD40特异性结合的抗原结合结构域包含
(i)重链可变区(V
(ii)轻链可变区(V
在一个方面,提供了一种双特异性抗原结合分子,其中能够与CD40特异性结合的抗原结合结构域包含:
(a)VH,其包含SEQ ID NO:37的氨基酸序列,和VL,其包含SEQ ID NO:41的氨基酸序列,或者
(b)VH,其包含SEQ ID NO:37的氨基酸序列,和VL,其包含SEQ ID NO:42的氨基酸序列,或者
(c)VH,其包含SEQ ID NO:37的氨基酸序列,和VL,其包含SEQ ID NO:43的氨基酸序列,或者
(d)VH,其包含SEQ ID NO:37的氨基酸序列,和VL,其包含SEQ ID NO:44的氨基酸序列,或者
(e)VH,其包含SEQ ID NO:38的氨基酸序列,和VL,其包含SEQ ID NO:41的氨基酸序列,或者
(f)VH,其包含SEQ ID NO:38的氨基酸序列,和VL,其包含SEQ ID NO:42的氨基酸序列,或者
(g)VH,其包含SEQ ID NO:38的氨基酸序列,和VL,其包含SEQ ID NO:43的氨基酸序列,或者
(h)VH,其包含SEQ ID NO:38的氨基酸序列,和VL,其包含SEQ ID NO:44的氨基酸序列,或者
(i)VH,其包含SEQ ID NO:39的氨基酸序列,和VL,其包含SEQ ID NO:41的氨基酸序列,或者
(j)VH,其包含SEQ ID NO:39的氨基酸序列,和VL,其包含SEQ ID NO:42的氨基酸序列,或者
(k)VH,其包含SEQ ID NO:39的氨基酸序列,和VL,其包含SEQ ID NO:43的氨基酸序列,或者
(l)VH,其包含SEQ ID NO:39的氨基酸序列,和VL,其包含SEQ ID NO:44的氨基酸序列,或者
(m)VH,其包含SEQ ID NO:40的氨基酸序列,和VL,其包含SEQ ID NO:41的氨基酸序列,或者
(n)VH,其包含SEQ ID NO:40的氨基酸序列,和VL,其包含SEQ ID NO:42的氨基酸序列,或者
(o)VH,其包含SEQ ID NO:40的氨基酸序列,和VL,其包含SEQ ID NO:43的氨基酸序列,或者
(p)VH,其包含SEQ ID NO:40的氨基酸序列,和VL,其包含SEQ ID NO:44的氨基酸序列。
在某一特定方面,能够与CD40特异性结合的抗原结合结构域包含:VH,其包含SEQID NO:37的氨基酸序列,和VL,其包含SEQ ID NO:41的氨基酸序列。
在进一步方面,提供了一种双特异性抗原结合分子,其中能够与CD40特异性结合的抗原结合结构域包含
(i)重链可变区(V
(ii)轻链可变区(V
在一个方面,提供了一种双特异性抗原结合分子,其中能够与CD40特异性结合的抗原结合结构域包含:
(a)VH,其包含SEQ ID NO:45的氨基酸序列,和VL,其包含SEQ ID NO:51的氨基酸序列,或者
(b)VH,其包含SEQ ID NO:46的氨基酸序列,和VL,其包含SEQ ID NO:51的氨基酸序列,或者
(c)VH,其包含SEQ ID NO:47的氨基酸序列,和VL,其包含SEQ ID NO:51的氨基酸序列,或者
(d)VH,其包含SEQ ID NO:48的氨基酸序列,和VL,其包含SEQ ID NO:51的氨基酸序列,或者
(e)VH,其包含SEQ ID NO:45的氨基酸序列,和VL,其包含SEQ ID NO:52的氨基酸序列,或者
(f)VH,其包含SEQ ID NO:46的氨基酸序列,和VL,其包含SEQ ID NO:52的氨基酸序列,或者
(g)VH,其包含SEQ ID NO:47的氨基酸序列,和VL,其包含SEQ ID NO:52的氨基酸序列,或者
(h)VH,其包含SEQ ID NO:48的氨基酸序列,和VL,其包含SEQ ID NO:52的氨基酸序列,或者
(i)VH,其包含SEQ ID NO:49的氨基酸序列,和VL,其包含SEQ ID NO:53的氨基酸序列,或者
(j)VH,其包含SEQ ID NO:50的氨基酸序列,和VL,其包含SEQ ID NO:53的氨基酸序列,或者
(k)VH,其包含SEQ ID NO:49的氨基酸序列,和VL,其包含SEQ ID NO:54的氨基酸序列,或者
(l)VH,其包含SEQ ID NO:50的氨基酸序列,和VL,其包含SEQ ID NO:54的氨基酸序列。
在某一特定方面,提供了一种双特异性抗原结合分子,其包含:(i)至少一个能够与CD40特异性结合的抗原结合结构域,其包含:重链可变区(V
与CD40和FAP结合的双特异性抗原结合分子
在另一方面,提供了一种双特异性抗原结合分子,其包含:
(i)至少一个能够与CD40特异性结合的抗原结合结构域,其包含:重链可变区(V
(ii)至少一个能够与FAP特异性结合的抗原结合结构域,其包含:重链可变区(V
在进一步方面,提供了一种双特异性抗原结合分子,其包含:
(i)至少一个能够与CD40特异性结合的抗原结合结构域,其包含:重链可变区(V
(ii)至少一个能够与FAP特异性结合的抗原结合结构域,其包含:重链可变区(V
在某一特定方面,提供了一种双特异性抗原结合分子,其包含:
(i)至少一个能够与CD40特异性结合的抗原结合结构域,其包含:重链可变区(V
(ii)至少一个能够与FAP特异性结合的抗原结合结构域,其包含:重链可变区(V
在另一个特定方面,提供了一种双特异性抗原结合分子,其包含:
(i)至少一个能够与CD40特异性结合的抗原结合结构域,其包含:重链可变区(V
(ii)至少一个能够与FAP特异性结合的抗原结合结构域,其包含:重链可变区(V
在进一步方面,提供了一种双特异性抗原结合分子,其中(i)能够与CD40特异性结合的抗原结合结构域包含:重链可变区(V
(ii)能够与FAP特异性结合的抗原结合结构域包含:重链可变区VH,其包含SEQ IDNO:15的氨基酸序列,以及轻链可变区,其包含SEQ ID NO:21的氨基酸序列。
此外,提供了一种双特异性抗原结合分子,其中
(i)能够与CD40特异性结合的抗原结合结构域包含:重链可变区(V
(ii)能够与FAP特异性结合的抗原结合结构域包含:重链可变区VH,其包含SEQ IDNO:15的氨基酸序列,以及轻链可变区,其包含SEQ ID NO:21的氨基酸序列。
在某一特定方面,提供了一种双特异性抗原结合分子,其包含:(i)至少一个能够与CD40特异性结合的抗原结合结构域,其包含:重链可变区(V
双特异性单价抗原结合分子(1+1形式)
在一个方面,本发明涉及双特异性抗原结合分子,其包含:
(a)一个能够与CD40特异性结合的抗原结合结构域:
(b)一个能够与成纤维细胞活化蛋白(FAP)特异性结合的抗原结合结构域,其包含:重链可变区(V
(i)CDR-H1,其包含SEQ ID NO:3的氨基酸序列,(ii)CDR-H2,其包含选自由SEQ IDNO:4、SEQ ID NO:11和SEQ ID NO:12组成的组的氨基酸序列,和(iii)CDR-H3,其包含SEQID NO:5的氨基酸序列;以及轻链可变区(V
在一个特定方面,提供了一种双特异性抗原结合分子,其中所述分子包含:(a)能够与CD40特异性结合的第一Fab片段;(b)能够与FAP特异性结合的第二Fab片段,其包含:重链可变区(V
在一个特定方面,本发明提供了一种双特异性抗原结合分子,其包含:(a)第一重链,其包含SEQ ID NO:63的氨基酸序列;第二重链,其包含SEQ ID NO:98的氨基酸序列;第一轻链,其包含SEQ ID NO:61的氨基酸序列;第二轻链,其包含SEQ ID NO:62的氨基酸序列。
与CD40二价结合并且与靶细胞抗原单价结合的双特异性抗原结合分子(2+1形式)
在另一方面,本发明提供了一种双特异性抗原结合分子,其包含:
(a)两个能够与CD40特异性结合的抗原结合结构域;
(b)一个能够与FAP特异性结合的抗原结合结构域,其包含:重链可变区(V
(c)Fc结构域,其由能够稳定缔合的第一亚基和第二亚基组成。
因此,提供了一种双特异性抗原结合分子,其中双特异性抗原结合分子与CD40双价结合并且与FAP单价结合。
在一个方面,双特异性抗原结合分子包含
(a)抗体的两个轻链和两个重链,该抗体包含两个能够与CD40特异性结合的Fab片段以及Fc结构域;以及
(b)能够与靶细胞抗原特异性结合的VH结构域和VL结构域,其中VH结构域和VL结构域各自经由肽连接基连接至两个重链的C末端中的一个。
在进一步方面,提供了一种双特异性抗原结合分子,其包含:
(a)两个重链,其中每个重链包含能够与CD40特异性结合的Fab片段的VH结构域和CH1结构域以及Fc区亚基;
(b)两个轻链,其中每个轻链包含能够与CD40特异性结合的Fab片段的VL和CL结构域;以及
(c)能够与FAP特异性结合的交叉fab片段,其包含VL-CH1链和VH-CL链,其中VH-CL链连接至(a)的两个重链中的一个的C末端。
在一个方面,VH-CL(VH-Cκ)链连接至Fc突起重链的C末端。
在另一方面,提供了一种双特异性抗原结合分子,其包含:
(a)两个重链,其中每个重链包含能够与CD40特异性结合的Fab片段的VH结构域和CH1结构域以及Fc区亚基;
(b)两个轻链,其中每个轻链包含能够与CD40特异性结合的Fab片段的VL和CL结构域;以及
(c)能够与FAP特异性结合的交叉fab片段,其包含VL-CH1链和VH-CL链,其中VL-CH1链连接至(a)的两个重链中的一个的C末端。
在一个方面,VL-CH1链连接至Fc突起重链的C末端。
在一个特定方面,本发明提供了一种双特异性抗原结合分子,其包含:
(a)两个轻链,其各自包含SEQ ID NO:62的氨基酸序列;一个轻链,其包含SEQ IDNO:61的氨基酸序列;第一重链,其包含SEQ ID NO:63的氨基酸序列;以及第二重链,其包含SEQ ID NO:64的氨基酸序列;或者
(b)两个轻链,其各自包含SEQ ID NO:66的氨基酸序列;一个轻链,其包含SEQ IDNO:65的氨基酸序列;第一重链,其包含SEQ ID NO:67的氨基酸序列;以及第二重链,其包含SEQ ID NO:68的氨基酸序列。
在一个特定方面,本发明提供了一种双特异性抗原结合分子,其包含:
(a)两个轻链,其各自包含SEQ ID NO:62的氨基酸序列;一个轻链,其包含SEQ IDNO:61的氨基酸序列;第一重链,其包含SEQ ID NO:63的氨基酸序列;以及第二重链,其包含SEQ ID NO:64的氨基酸序列。
头到尾形式(2+1)的双特异性抗原结合分子
在另一方面,提供了一种双特异性抗原结合分子,其包含:
(a)重链,其包含能够与CD40特异性结合的Fab片段的VH结构域和CH1结构域以及Fc区亚基;
(b)重链,其包含能够与CD40特异性结合的Fab片段的VH结构域和CH1结构域、能够与FAP特异性结合的Fab片段的VL结构域和CH1结构域以及Fc区亚基;
(c)两个轻链,其中每个轻链包含能够与CD40特异性结合的Fab片段的VL和CL结构域;以及
(d)轻链,其包含能够与FAP特异性结合的Fab片段的VH和CL结构域。
与CD40二价结合并且与靶细胞抗原二价结合的双特异性抗原结合分子(2+2形式)
在另一方面,本发明提供了一种双特异性抗原结合分子,其包含:
(a)两个能够与CD40特异性结合的抗原结合结构域;
(b)两个能够与FAP特异性结合的抗原结合结构域,其包含:(i)CDR-H1,其包含SEQID NO:3的氨基酸序列,(ii)CDR-H2,其包含选自由SEQ ID NO:4、SEQ ID NO:11和SEQ IDNO:12组成的组的氨基酸序列,和(iii)CDR-H3,其包含SEQ ID NO:5的氨基酸序列;以及轻链可变区(V
(c)Fc结构域,其由能够稳定缔合的第一亚基和第二亚基组成。
因此,提供了一种双特异性抗原结合分子,其中双特异性抗原结合分子与CD40二价结合并且与FAP单价结合。
在一个方面,提供了一种双特异性抗原结合分子,其中双特异性抗原结合分子包含:
(a)两个重链,其中每个重链包含能够与CD40特异性结合的Fab片段的VH结构域和CH1结构域以及Fc区亚基;
(b)两个轻链,其中每个轻链包含能够与CD40特异性结合的Fab片段的VL和CL结构域;以及
(c)两个能够与FAP特异性结合的Fab片段,其中Fab片段中的一个连接至(a)的两个重链中的一个的C末端,并且Fab片段中的另一个连接至(a)的两个重链中的另一个的C末端。
与CD40四价结合并且与靶细胞抗原单价结合的双特异性抗原结合分子(4+1形式)
在另一方面,本发明提供了一种双特异性抗原结合分子,其包含:
(a)四个能够与CD40特异性结合的抗原结合结构域;
(b)一个能够与FAP特异性结合的抗原结合结构域,其包含:(i)CDR-H1,其包含SEQID NO:3的氨基酸序列,(ii)CDR-H2,其包含选自由SEQ ID NO:4、SEQ ID NO:11和SEQ IDNO:12组成的组的氨基酸序列,和(iii)CDR-H3,其包含SEQ ID NO:5的氨基酸序列;以及轻链可变区(V
(c)Fc结构域,其由能够稳定缔合的第一亚基和第二亚基组成。
因此,提供了一种双特异性抗原结合分子,其中双特异性抗原结合分子与CD40四价结合并且与FAP单价结合。
在一个方面,提供了一种双特异性抗原结合分子,其中四个能够与CD40特异性结合的抗原结合结构域为Fab片段,并且其中各有两个抗原结合结构域彼此融合至重链处,任选地经由肽连接基融合。在某一特定方面,肽连接基包含SEQ ID NO:83的氨基酸序列。更具体地,抗原结合分子包含两个重链,这两个重链各自包含VHCH1-肽连接基-VHCH1片段。在某一特定方面,肽连接基具有SEQ ID NO:83的氨基酸序列。
在某一特定方面,双特异性抗原结合分子包含;
(a)四个轻链,其中每个轻链包含能够与CD40特异性结合的Fab片段的VL和CL结构域;
(b)两个重链,其中每个重链包含能够与CD40特异性结合的Fab片段的VH结构域和CH1结构域,其融合至能够与CD40特异性结合的第二Fab片段的VH结构域和CH1结构域,以及Fc区亚基;以及
(c)能够与FAP特异性结合的交叉fab片段,其中VH结构域经由肽连接基连接至重链中的一个的C末端。
在另一方面,双特异性抗原结合分子包含
(a)四个轻链,其中每个轻链包含能够与CD40特异性结合的Fab片段的VL和CL结构域;
(b)两个重链,其中每个重链包含能够与CD40特异性结合的Fab片段的VH结构域和CH1结构域,其融合至能够与CD40特异性结合的第二Fab片段的VH结构域和CH1结构域,以及Fc区亚基;以及
(c)能够与FAP特异性结合的交叉fab片段,其中VL结构域经由肽连接基连接至重链中的一个的C末端。
在某一特定方面,肽连接基包含选自由SEQ ID NO:82、SEQ ID NO:83、SEQ ID NO:87和SEQ ID NO:88组成的组的氨基酸序列。更具体地,肽连接基包含SEQ ID NO:83。
在某一特定方面,本发明提供了一种双特异性抗原结合分子,其包含:
(a)四个轻链,其各自包含SEQ ID NO:62的氨基酸序列;一个轻链,其包含SEQ IDNO:61的氨基酸序列;第一重链,其包含SEQ ID NO:69的氨基酸序列;以及第二重链,其包含SEQ ID NO:70的氨基酸序列。
与CD40四价结合并且与靶细胞抗原二价结合的双特异性抗原结合分子(4+2形式)
在另一方面,本发明提供了一种双特异性抗原结合分子,其包含:
(a)四个能够与CD40特异性结合的抗原结合结构域;
(b)两个能够与FAP特异性结合的抗原结合结构域,其包含:
(i)CDR-H1,其包含SEQ ID NO:3的氨基酸序列,(ii)CDR-H2,其包含选自由SEQ IDNO:4、SEQ ID NO:11和SEQ ID NO:12组成的组的氨基酸序列,和(iii)CDR-H3,其包含SEQID NO:5的氨基酸序列;以及轻链可变区(V
(c)Fc结构域,其由能够稳定缔合的第一亚基和第二亚基组成:
因此,提供了一种双特异性抗原结合分子,其中双特异性抗原结合分子与CD40四价结合并且与FAP二价结合。
在一个方面,提供了一种双特异性抗原结合分子,其中四个能够与CD40特异性结合的抗原结合结构域为Fab片段,并且其中各有两个抗原结合结构域彼此融合,任选地经由肽连接基融合。在某一特定方面,肽连接基包含SEQ ID NO:83的氨基酸序列。更具体地,抗原结合分子包含两个重链,这两个重链各自包含VHCH1-肽连接基-VHCH1片段。在某一特定方面,肽连接基具有SEQ ID NO:83的氨基酸序列。
在另一方面,提供了一种双特异性抗原结合分子,其中能够与靶细胞抗原特异性结合的抗原结合结构域为Fab片段,并且其中第一Fab片段经由肽连接基连接至Fc结构域的第一亚基的C末端,并且第二Fab片段经由肽连接基连接至Fc结构域的第二亚基的C末端。在一个方面,能够与靶细胞抗原特异性结合的两个Fab片段为交叉Fab片段,其各自包含VL-CH1链和VH-CL链,并且其中VL-CH1链连接至两个重链中的一个的C末端。
降低Fc受体结合和/或效应子功能的Fc结构域修饰
本发明的双特异性抗原结合分子进一步包含由能够稳定结合的第一亚基和第二亚基组成的Fc结构域。
在某些方面,一个或多个氨基酸修饰可引入本文提供的抗体的Fc区中,从而生成Fc区变体。Fc区变体可包含人Fc区序列(例如人IgG1、IgG2、IgG3或IgG4 Fc区),其在一个或多个氨基酸位置上包含氨基酸修饰(例如取代)。
Fc结构域为本发明的双特异性抗体赋予有利的药代动力学特性,包括有助于靶组织中的良好积聚的长血清半衰期和有利的组织-血液分配比。然而,与此同时,可能导致本发明的双特异性抗体不期望地靶向表达Fc受体的细胞,而不是优选的抗原携带细胞。因此,在特定实施例中,与天然IgG Fc结构域,特别是IgG1 Fc结构域或IgG4 Fc结构域相比,本发明的双特异性抗体的Fc结构域表现出降低的对Fc受体的结合亲和力和/或降低的效应子功能。更具体地,Fc结构域是IgG1 Fc结构域。
在一个此类方面,与天然IgG1 Fc结构域(或包含天然IgG1 Fc结构域的本发明的双特异性抗原结合分子)相比,该Fc结构域(或包含该Fc结构域的本发明的双特异性抗原结合分子)表现出对Fc受体的结合亲和力的小于50%、优选地少小20%、更优选地小于10%且最优选地小于5%;和/或与天然IgG1 Fc结构域(或包含天然IgG1 Fc结构域的本发明的双特异性抗原结合分子)相比,该Fc结构域(或包含该Fc结构域的本发明的双特异性抗原结合分子)表现出效应子功能的小于50%、优选地小于20%、更优选地小于10%且最优选地小于5%。在一个方面,Fc结构域(或包含该Fc结构域的本发明的双特异性抗原结合分子)不显著结合Fc受体和/或诱导效应子功能。在一个特定方面,Fc受体是Fcγ受体。在一个方面,Fc受体是人Fc受体。在一个方面,Fc受体是活化的Fc受体。在一个具体方面,Fc受体是活化的人Fcγ受体,更具体地是人FcγRIIIa、FcγRI或FcγRIIa,最具体地是人FcγRIIIa。在一个方面,Fc受体是抑制性Fc受体。在一个具体方面,Fc受体是抑制性人Fcγ受体,更具体地是人FcγRIIB。在一个方面,效应子功能是CDC、ADCC、ADCP和细胞因子分泌中的一个或多个。在一个特定方面,效应子功能是ADCC。在一个方面,与天然IgG1 Fc结构域相比,Fc结构域表现出对新生儿Fc受体(FcRn)基本上类似的结合亲和力。当Fc结构域(或包含所述Fc结构域的本发明的双特异性抗原结合分子)表现出天然IgG1 Fc结构域(或包含天然IgG1 Fc结构域的本发明的双特异性抗原结合分子)对FcRn的结合亲和力的大于约70%、具体地大于约80%、更具体地大于约90%时,实现了与FcRn基本上类似的结合。
在一个特定方面,Fc结构域经过工程化改造,以与非工程化改造的Fc结构域相比具有降低的对Fc受体的结合亲和力和/或降低的效应子功能。在一个特定方面,本发明的双特异性抗原结合分子的Fc结构域包含降低Fc结构域对Fc受体的结合亲和力和/或效应子功能的一个或多个氨基酸突变。典型地,相同的一个或多个氨基酸突变存在于Fc结构域的两个亚基中的每一个中。在一个方面,氨基酸突变降低Fc结构域对Fc受体的结合亲和力。在另一方面,氨基酸突变将Fc结构域对Fc受体的结合亲和力降低至少2倍、至少5倍或至少10倍。在一个方面,与包含非工程化改造的Fc结构域的本发明的双特异性抗体相比,包含工程化改造的Fc结构域的本发明的双特异性抗原结合分子表现出对Fc受体的结合亲和力的小于20%、具体地小于10%、更具体地小于5%。在一个特定方面,Fc受体是Fcγ受体。在其他方面,Fc受体是人Fc受体。在一个方面,Fc受体是抑制性Fc受体。在一个具体方面,Fc受体是抑制性人Fcγ受体,更具体地是人FcγRIIB。在一些方面,Fc受体是活化的Fc受体。在一个具体方面,Fc受体是活化的人Fcγ受体,更具体地是人FcγRIIIa、FcγRI或FcγRIIa,最具体地是人FcγRIIIa。优选地,与这些受体中的每一个的结合力降低。在一些方面,对互补组分的结合亲和力,对C1q的特异性结合亲和力也降低。在一个方面,对新生儿Fc受体(FcRn)的结合亲和力不降低。当Fc结构域(或包含Fc结构域的本发明的双特异性抗原结合分子)表现出Fc结构域的非工程化改造形式(或包含Fc结构域的非工程化改造形式的本发明的双特异性抗原结合分子)对FcRn的结合亲和力的大于约70%时,实现了基本上类似的与FcRn的结合,即实现了Fc结构域对该受体的结合亲和力的保留。Fc结构域或包含该Fc结构域的本发明的双特异性抗原结合分子可表现出这种亲和力的大于约80%或甚至大于约90%。在某些实施例中,本发明的双特异性抗原结合分子的Fc结构域被工程化改造,以相比于非工程化改造的Fc结构域具有降低的效应子功能。降低的效应子功能可以包括但不限于以下中的一个或多个:降低的补体依赖性细胞毒性(CDC)、降低的抗体依赖性细胞介导的细胞毒性(ADCC)、降低的抗体依赖性细胞吞噬(ADCP)、减少的细胞因子分泌、减少的免疫复合物介导的抗原呈递细胞的抗原摄取、减少的与NK细胞的结合、减少的与巨噬细胞的结合、减少的与单核细胞的结合、减少的与多形核细胞的结合、减少的直接信号传导诱导性细胞凋亡、降低的树突细胞成熟或减少的T细胞引发。
具有降低的效应子功能的抗体包括具有一个或多个Fc区残基238、265、269、270、297、327和329的取代的那些(美国专利号6,737,056)。此类Fc突变体包括在第265、269、270、297和327位氨基酸的两个或多个处具有取代的Fc突变体,包括所谓的“DANA”Fc突变体,其残基265和297被取代为丙氨酸(美国专利号7,332,581)。描述了具有改善的或降低的与FcR的结合的某些抗体变体。(例如美国专利号6,737,056;WO 2004/056312,以及Shields,R.L.等人,J.Biol.Chem.276(2001)6591-6604)。
在本发明的一个方面,Fc结构域在E233、L234、L235、N297、P331和P329位处包含氨基酸取代。在一些方面,Fc结构域包含氨基酸取代L234A和L235A(“LALA”)。在一个此类实施例中,Fc结构域是IgG1 Fc结构域,特别是人IgG1 Fc结构域。在一个方面,Fc结构域在P329位处包含氨基酸取代。在一个更具体的方面,氨基酸取代是P329A或P329G,特别地是P329G。在一个实施例中,Fc结构域在位置P329处包含氨基酸取代并且包含选自由E233P、L234A、L235A、L235E、N297A、N297D或P331S组成的组的另外的氨基酸取代。在更特定的实施例中,Fc结构域包含氨基酸突变L234A、L235A和P329G(“P329G LALA”)。氨基酸取代的“P329GLALA”组合几乎完全消除人IgG1 Fc结构域的Fcγ受体结合,如PCT专利申请号WO 2012/130831 A1中所述。所述文档还描述了制备此类突变Fc结构域的方法和用于确定其性质(诸如Fc受体结合或效应子功能)的方法。此类抗体是具有突变L234A和L235A或具有突变L234A、L235A和P329G的IgG1(根据Kabat等人的EU索引编号,参见:Kabat等人,Sequencesof Proteins of Immunological Interest,第5版Public Health Service,NationalInstitutes of Health,Bethesda,MD,1991)。
在一个方面,Fc结构域为IgG4 Fc结构域。在一个更具体的实施例中,Fc结构域是在S228位处(Kabat编号)包含氨基酸取代,具体地是氨基酸取代S228P的IgG4 Fc结构域。在一个更具体的实施例中,Fc结构域是包含氨基酸取代L235E和S228P以及P329G的IgG4 Fc结构域。这种氨基酸取代减少IgG4抗体的体内Fab臂交换(参见Stubenrauch等人,DrugMetabolism and Disposition 38,84-91(2010))。
具有延长的半衰期和改善的与新生儿Fc受体(FcRn)结合、负责将母体IgG转移至胎儿(Guyer,R.L.等人,J.Immunol.117(1976)587-593,以及Kim,J.K.等人,J.Immunol.24(1994)2429-2434)的抗体描述于US 2005/0014934中。那些抗体包含这样的Fc区,所述Fc区中具有改善Fc区与FcRn的结合的一个或多个取代。此类Fc变体包括在以下Fc区残基中的一处或多处具有取代的Fc变体:238、256、265、272、286、303、305、307、311、312、317、340、356、360、362、376、378、380、382、413、424或434,例如对Fc区残基434的取代(美国专利号7,371,826)。关于Fc区变体的其他示例,还参见Duncan,A.R.和Winter,G.,Nature 322(1988)738-740;US 5,648,260;US 5,624,821;以及WO 94/29351。
与Fc受体的结合可以例如通过ELISA或通过表面等离子体共振(SPR)使用标准仪器(诸如BIAcore仪器(GE Healthcare))容易地确定,并且Fc受体诸如可以通过重组表达获得。本文描述了合适的此类结合测定。另选地,可以使用已知表达特定Fc受体的细胞系(诸如表达FcγIIIa受体的人NK细胞)来评估Fc结构域或包含Fc结构域的细胞活化双特异性抗原结合分子对Fc受体的结合亲和力。Fc结构域的效应子功能,或本发明的包含Fc结构域的双特异性抗原结合分子,可以通过本领域已知的方法来测量。本文描述了用于测量ADCC的合适的测定法。用于评定感兴趣的分子的ADCC活性的体外测定的其他示例描述于美国专利号5,500,362;Hellstrom等人,Proc Natl Acad Sci USA 83,7059-7063(1986)和Hellstrom等人,Proc Natl Acad Sci USA 82,1499-1502(1985);美国专利号5,821,337;Bruggemann等人,J Exp Med 166,1351-1361(1987)。另选地,可以使用非放射性测定方法(参见例如,用于流式细胞术的ACTI
以下部分描述了包含降低Fc受体结合和/或效应子功能的Fc结构域修饰的本发明的双特异性抗原结合分子的优选方面。在一个方面,本发明涉及双特异性抗原结合分子,其包含:(a)至少一个能够与CD40特异性结合的抗原结合结构域;(b)至少一个能够与FAP特异性结合的抗原结合结构域;以及(c)由能够稳定缔合的第一亚基和第二亚基组成的Fc结构域,其中Fc结构域包含一个或多个氨基酸取代,该氨基酸取代降低抗体与Fc受体的结合亲和力,特别是对Fc受体的结合亲和力。在另一方面,本发明涉及双特异性抗原结合分子,其包含:(a)至少一个能够与CD40特异性结合的抗原结合结构域;(b)至少一个能够与靶细胞抗原特异性结合的抗原结合结构域;以及(c)由能够稳定缔合的第一亚基和第二亚基组成的Fc结构域,其中Fc结构域包含一个或多个氨基酸取代,该氨基酸取代降低效应子功能。在特定方面,Fc结构域属于人IgG1亚类,其具有氨基酸突变L234A、L235A和P329G(根据KabatEU索引编号)。
促进异源二聚化的Fc结构域修饰
本发明的双特异性抗原结合分子包含与Fc结构域的两个亚基中的一个或另一个融合的不同抗原结合位点,因此该Fc结构域的两个亚基可包含在两个不相同的多肽链中。这些多肽的重组共表达和随后的二聚化导致了两种多肽的几种可能的组合。为了在重组生产中提高本发明的双特异性抗原结合分子的产率和纯度,因此在本发明的双特异性抗原结合分子的Fc结构域中引入促进所需多肽的缔合的修饰将是有利的。
因此,在特定方面,本发明涉及双特异性抗原结合分子,其包含:(a)至少一个能够与CD40特异性结合的抗原结合结构域;(b)至少一个能够与FAP特异性结合的抗原结合结构域;以及(c)由能够稳定缔合的第一亚基和第二亚基组成的Fc结构域,其中Fc结构域包含促进Fc结构域的第一亚基和第二亚基的缔合的修饰。人IgG Fc结构域的两个亚基之间最广泛的蛋白质间相互作用位点在Fc结构域的CH3结构域中。因此,在一个方面,所述修饰在Fc结构域的CH3结构域中。
在一个具体方面,所述修饰是所谓的“突起进入孔”修饰,所述修饰包括Fc结构域的两个亚基中的一个中的“突起”修饰和Fc结构域的两个亚基中的另一个中的“孔”修饰。因此,本发明涉及双特异性抗原结合分子,其包含:(a)至少一个能够与CD40特异性结合的抗原结合结构域,(b)至少一个能够与靶细胞抗原特异性结合的抗原结合结构域,以及(c)由能够稳定缔合的第一亚基和第二亚基组成的Fc结构域,其中根据杵臼结构方法,Fc结构域的第一亚基包含突起并且Fc结构域的第二亚基包含孔。在一个特定方面,Fc结构域的第一亚基包含氨基酸取代S354C和T366W(EU编号)而Fc结构域的第二亚基包含氨基酸取代Y349C、T366S和Y407V(根据Kabat EU索引编号)。
杵臼结构技术描述于例如US 5,731,168;US 7,695,936;Ridgway等人,Prot Eng9,617-621(1996)和Carter,J Immunol Meth 248,7-15(2001)中。通常,该方法涉及在第一多肽的界面处引入凸起(“突起”)并在第二多肽的界面中引入相应的空腔(“孔”),使得所述凸起可以定位在所述空腔中,以便促进异二聚体的形成并阻碍同二聚体的形成。凸起是通过用较大侧链(例如酪氨酸或色氨酸)取代来自第一多肽的界面的小氨基酸侧链而构建的。具有与凸起相同或相似大小的补偿空腔是通过用较小的氨基酸侧链(例如丙氨酸或苏氨酸)取代大氨基酸侧链而在第二多肽的界面中创建的。
因此,在一个方面,在本发明的双特异性抗原结合分子的Fc结构域的第一亚基的CH3结构域中,某一氨基酸残基被具有较大侧链体积的氨基酸残基取代,从而在第一亚基的CH3结构域内产生凸起,所述凸起可定位于第二亚基的CH3结构域内的空腔中;而在Fc结构域的第二亚基的CH3结构域中,某一氨基酸残基被具有较小侧链体积的氨基酸残基取代,从而在第二亚基的CH3结构域内产生空腔,该第一亚基的CH3结构域内的所述凸起可定位在该空腔内。凸起和空腔可以通过改变编码多肽的核酸来制备,例如通过位点特异性诱变或通过肽合成。在一个具体方面,在Fc结构域的第一亚基的CH3结构域中,366位处的苏氨酸残基被色氨酸残基(T366W)取代,而在Fc结构域的第二亚基的CH3结构域中,407位处的酪氨酸残基被缬氨酸残基(Y407V)取代。在一个方面中,另外在Fc结构域的第二亚基中,366位处的苏氨酸残基被丝氨酸残基(T366S)取代,并且368位处的亮氨酸残基被丙氨酸残基(L368A)取代。
在另一个方面,另外在Fc结构域的第一亚基中,354位处的丝氨酸残基被半胱氨酸残基(S354C)取代,并且另外在Fc结构域的第二亚基中,349位的酪氨酸残基被半胱氨酸残基(Y349C)取代。引入这两个半胱氨酸残基导致在Fc结构域的两个亚基之间形成二硫桥,从而进一步稳定该二聚体(Carter(2001),J Immunol Methods 248,7-15)。在一个特定方面,Fc结构域的第一亚基包含氨基酸取代S354C和T366W(EU编号)而Fc结构域的第二亚基包含氨基酸取代Y349C、T366S和Y407V(根据Kabat EU索引编号)。
在另一方面,促进Fc结构域的第一亚基和第二亚基缔合的修饰包括介导静电转向效应的修饰,例如如在PCT公开WO 2009/089004中所述。通常,该方法涉及用带电荷的氨基酸残基取代两个Fc结构域亚基的界面处的一个或多个氨基酸残基,使得同源二聚体形成变得在静电上不利,但异源二聚化在静电上有利。
如本文报道的双特异性抗体的重链的C末端可以是以氨基酸残基PGK结束的完整C末端。重链的C末端可以是缩短的C末端,在所述缩短的C末端中已经去除了一个或两个C末端氨基酸残基。在一个优选的方面,重链的C末端是以PG结束的缩短的C末端。在本文报道的所有方面中的一个方面,如本文所指定的包含含有C末端CH3结构域的重链的双特异性抗体,包含C末端甘氨酸-赖氨酸二肽(G446和K447,根据Kabat EU索引编号)。在本文报道的所有方面的一个实施例中,如本文所指定的包含含有C末端CH3结构域的重链的双特异性抗体,包含C末端甘氨酸残基(G446,根据Kabat EU索引编号)。
Fab结构域中的修饰
在一个方面,本发明涉及双特异性抗原结合分子,其包含:(a)能够与CD40特异性结合的第一Fab片段;(b)能够与FAP特异性结合的第二Fab片段;以及(c)由能够稳定缔合的第一亚基和第二亚基组成的Fc结构域,其中在Fab片段之一中,可变结构域VH和VL或恒定结构域CH1和CL被交换。根据Crossmab技术制备双特异性抗体。
在WO2009/080252和Schaefer,W.等人,PNAS,108(2011)11187-1191中详细描述了在一个结合臂中具有结构域置换/交换的多特异性抗体(CrossMabVH-VL或CrossMabCH-CL)。它们明显减少了由抗第一抗原的轻链与抗第二抗原的错误重链的错配导致的副产物(与没有此类结构域交换的方法相比)。
在一个方面,本发明涉及双特异性抗原结合分子,其包含:(a)能够与CD40特异性结合的第一Fab片段;(b)能够与FAP特异性结合的第二Fab片段;以及(c)由能够稳定缔合的第一亚基和第二亚基组成的Fc结构域,其中在Fab片段之一中,恒定结构域CL和CH1被彼此替换,使得CH1结构域是轻链的一部分,并且CL结构域是重链的一部分。更特别地,在能够与靶细胞抗原特异性结合的第二Fab片段中,恒定结构域CL和CH1被彼此替换,使得CH1结构域是轻链的一部分,并且CL结构域是重链的一部分。
在某一特定方面,本发明涉及一种双特异性抗原结合分子,其包含:(a)能够与CD40特异性结合的第一Fab片段;(b)能够与FAP特异性结合的第二Fab片段,其中恒定结构域CL和CH1被彼此替换,使得CH1结构域是轻链的一部分,并且CL结构域是重链的一部分。此类分子与CD40和FAP均单价结合。
在另一方面,本发明涉及一种双特异性抗原结合分子,其包含:(a)包含两个能够与CD40特异性结合的Fab片段和Fc结构域的抗体的两个轻链和两个重链;以及(b)两个能够与FAP特异性结合的另外的Fab片段,其中所述另外的Fab片段各自经由肽连接基连接至(a)的重链的C末端。在某一特定方面,另外的Fab片段是Fab片段,其中可变结构域VL和VH被彼此替换,使得VH结构域是轻链的一部分并且VL结构域是重链的一部分。
因此,在某一特定方面,本发明包含一种双特异性抗原结合分子,其包含:(a)包含两个能够与CD40特异性结合的Fab片段和Fc结构域的抗体的两个轻链和两个重链;以及(b)两个能够与FAP特异性结合的另外的Fab片段,其中能够与靶细胞抗原特异性结合的所述两个另外的Fab片段是交叉Fab片段,其中可变结构域VL和VH被彼此取代,并且VL-CH链各自经由肽连接基连接至(a)重链的C末端。
在另一方面,并进一步改善正确的配对,双特异性抗原结合分子可能含有不同的带电荷的氨基酸取代(所谓的“带电荷的残基”),其包含:(a)第一Fab片段,其能够与CD40特异性结合;(b)第二Fab片段,其能够与FAP特异性结合;以及(c)Fc结构域,由能够稳定缔合的第一亚基和第二亚基组成。将这些修饰引入交叉或非交叉的CH1和CL结构域中。在某一特定方面,本发明涉及一种双特异性抗原结合分子,其中在CL结构域之一中,123位(EU编号)的氨基酸已被精氨酸(R)取代并且/或者其中124位(EU编号)的氨基酸已被赖氨酸(K)取代;并且其中在CH1结构域之一中,147位(EU编号)和/或213位(EU编号)的氨基酸已被谷氨酸(E)取代。
本发明的示例性抗体
在一个方面,本发明提供了与FAP特异性结合的新型抗体和抗体片段。这些抗体与已知的FAP抗体4B)或28H1结合至不同的表位,这使其尤其适合掺入可与其他FAP靶向分子结合使用的双特异性抗原结合分子中。这些新型抗体的特征进一步在于,它们能够以高含量生产并且具有高滴度,表现出高的热稳定性(通过聚集温度T
在一个方面,提供了一种与FAP特异性结合的抗体(克隆212),其中所述抗体包含:重链可变区(V
在一个方面,提供了一种与FAP特异性结合的人源化抗体,其中所述抗体包含:重链可变区(V
在另一方面,提供了一种抗体,该抗体与特异性结合FAP的抗体竞争结合,其中所述抗体包含重链可变区(V
在一个方面,提供了一种抗体,该抗体与特异性结合FAP的抗体竞争结合,其中所述抗体包含:重链可变区VH,其包含SEQ ID NO:15的氨基酸序列,以及轻链可变区,其包含SEQ ID NO:21的氨基酸序列。
在进一步方面,提供了一种与FAP特异性结合的抗体,其中所述抗体包含:
(a)重链可变区(V
(b)重链可变区(V
(c)重链可变区(V
(d)重链可变区(V
在进一步方面,提供了一种与FAP特异性结合的抗体,其包含:重链可变区(V
多核苷酸
本发明进一步提供了编码如本文所述的双特异性抗原结合分子或其片段的分离的核酸或编码如本文所述的抗体的分离的核酸。
编码本发明的双特异性抗原结合分子的分离的多核苷酸可以表达为编码完整抗原结合分子的单个多核苷酸,或表达为共表达的多个(例如两个或更多个)多核苷酸。由共表达的多核苷酸编码的多肽可以经由例如二硫键或其他手段缔合以形成功能性抗原结合分子。例如,免疫球蛋白的轻链部分可以由来自免疫球蛋白的重链部分的单独多核苷酸编码。当共表达时,重链多肽将与轻链多肽缔合以形成免疫球蛋白。
在一些方面中,分离的多核苷酸编码如本文所述的包含在根据本发明的双特异性分子中的多肽。
在一个方面,本发明涉及编码双特异性抗原结合分子的分离的多核苷酸,该双特异性抗原结合分子包含:(a)至少一个能够与CD40特异性结合的抗原结合结构域;(b)至少一个能够与FAP特异性结合的抗原结合结构域,该FAP包含:重链可变区(V
在某些实施例中,多核苷酸或核酸是DNA。在其他实施例中,本发明的多核苷酸是RNA,例如以信使RNA(mRNA)的形式。本发明的RNA可以是单链或双链的。
重组方法
本发明的双特异性抗原结合分子可通过例如重组生产获得。对于重组生产,提供了一种或多种编码双特异性抗原结合分子或其多肽片段的多核苷酸。将一种或多种编码双特异性抗原结合分子的多核苷酸分离并插入至一种或多种载体中以用于在宿主细胞中进一步克隆和/或表达。此类多核苷酸可使用常规方法容易地分离并且测序。在本发明的一个方面,提供了一种载体,优选为表达载体,该载体包含本发明的多核苷酸中的一种或多种多核苷酸。可以使用本领域技术人员熟知的方法来构建含有双特异性抗原结合分子(片段)的编码序列以及适当转录/翻译控制信号的表达载体。这些方法包括体外重组DNA技术、合成技术和体内重组/遗传重组。参见例如以下文献所述的技术:Maniatis等人,MOLECULARCLONING:A LABORATORY MANUAL,Cold Spring Harbor Laboratory,N.Y.(1989);和Ausubel等人,CURRENT PROTOCOLS IN MOLECULAR BIOLOGY,Greene PublishingAssociates and Wiley Interscience,N.Y.(1989)。表达载体可以是质粒、病毒的一部分,或者可以是核酸片段。表达载体包括表达盒,编码双特异性抗原结合分子或其多肽片段的多核苷酸(即编码区)与启动子和/或其他转录或翻译控制元件可操作缔合地克隆至所述表达盒中。如本文所用,“编码区”是核酸的一部分,该部分由翻译成氨基酸的密码子组成。尽管“终止密码子”(TAG、TGA或TAA)未被翻译成氨基酸,但其(如果存在的话)可被认为是编码区的一部分,而任何侧翼序列,例如启动子、核糖体结合位点、转录终止子、内含子、5'和3'非翻译区等不是编码区的一部分。两个或更多个编码区可存在于单个多核苷酸构建体中(例如在单个载体上),或在单独的多核苷酸构建体中(例如在单独的(不同的)载体上)。此外,任何载体可含有单一编码区,或可包含两个或更多个编码区,例如本发明的载体可以编码一种或多种多肽,该一种或多种多肽在翻译后或翻译时通过蛋白水解切割分离成最终的蛋白质。另外,本发明的载体、多核苷酸或核酸可编码异源编码区,所述异源编码区与编码本发明的双特异性抗原结合分子或其多肽片段的多核苷酸或其变体或衍生物融合或不融合。异源编码区包括但不限于特化元件或基序,诸如分泌信号肽或异源功能结构域。可操作缔合是当基因产物(例如多肽)的编码区以某种方式与一个或多个调控序列缔合,以使基因产物的表达处于调控序列的影响或控制下。如果启动子功能的诱导导致编码所需基因产物的mRNA的转录,并且如果两个DNA片段之间的连接性质不干扰表达调控序列指导基因产物表达的能力或干扰待转录的基因模板的能力,则该两个DNA片段(诸如多肽编码区和与其相关的启动子)是“可操作地缔合的”。因此,如果启动子能够影响该核酸的转录,则启动子区域将与编码多肽的核酸可操作地缔合。启动子可以是细胞特异性启动子,该细胞特异性启动子仅在预定细胞中指导DNA的实质转录。除启动子外,其他转录控制元件,例如增强子、操纵子、阻遏物和转录终止信号,可以与多核苷酸可操作地缔合以指导细胞特异性转录。
本文公开了合适的启动子和其他转录控制区。多种转录控制区是本领域技术人员已知的。这些转录控制区包括但不限于在脊椎动物细胞中起作用的转录控制区,诸如但不限于来自巨细胞病毒的启动子和增强子区段(例如立即早期启动子结合内含子-A)、猿猴病毒40(例如早期启动子)和逆转录病毒(诸如例如劳氏肉瘤病毒)。其他转录控制区包括来源于脊椎动物基因(诸如肌动蛋白、热休克蛋白、牛生长激素和兔
本发明的多核苷酸和核酸编码区可以与编码分泌肽或信号肽的附加编码区缔合,所述附加编码区指导由本发明的多核苷酸编码的多肽的分泌。例如,如果需要分泌双特异性抗原结合分子或其多肽片段,可以将编码信号序列的DNA置于编码本发明的本发明的双特异性抗原结合分子或其多肽片段的核酸的上游。根据信号假设,由哺乳动物细胞分泌的蛋白质具有信号肽或分泌前导序列,一旦已经起始跨粗面内质网输出生长的蛋白质链,该信号肽或分泌前导序列就被从成熟蛋白质上切割下来。本领域普通技术人员知道由脊椎动物细胞分泌的多肽通常具有与所述多肽的N末端融合的信号肽,所述信号肽被从翻译的多肽上切割下来以产生所述多肽的分泌或“成熟”形式。在某些实施例中,使用天然信号肽(例如免疫球蛋白重链或轻链信号肽),或保留指导与其可操作地缔合的多肽分泌的能力的该序列的功能性衍生物。另选地,可以使用异源哺乳动物信号肽或其功能衍生物。例如,野生型前导序列可以被人组织纤溶酶原活化剂(TPA)或小鼠β葡糖醛酸酶的前导序列取代。
DNA编码可用于促进后续纯化的短蛋白序列(例如组氨酸标签)或帮助标记融合蛋白的DNA可包含在编码本发明的双特异性抗原结合分子或其多肽片段的多核苷酸的内部或末端。
在本发明的另一方面,提供了包含一种或多种本发明的多核苷酸的宿主细胞。在某些方面,提供了包含一种或多种本发明的载体的宿主细胞。多核苷酸和载体可以单独或组合地渗入本文中分别关于多核苷酸和载体描述的任何特征。在一个方面,宿主细胞包含(例如,已转化或转染)载体,该载体包含编码本发明的双特异性抗原结合分子(的一部分)的多核苷酸。如本文所用,术语“宿主细胞”是指可以被工程化改造以产生本发明的融合蛋白或其片段的任何种类的细胞系统。适于复制和支持抗原结合分子表达的宿主细胞是本领域中熟知的。此类细胞可以用特定的表达载体适当地转染或转导,并且可以生长大量含有载体的细胞以用于接种大规模发酵罐来获得足够量的抗原结合分子用于临床应用。合适的宿主细胞包括原核微生物,诸如大肠杆菌,或各种真核细胞,诸如中国仓鼠卵巢细胞(CHO)、昆虫细胞等。例如,多肽可以在细菌中产生,特别是当不需要糖基化时。多肽在表达后可以在可溶性级分中从细菌细胞糊状物中分离,并可以进一步纯化。除原核生物外,诸如丝状真菌或酵母之类的真核微生物也是用于编码多肽的载体的合适克隆或表达宿主,包括这样的真菌和酵母菌株,该真菌和酵母菌株的糖基化途径已经被“人源化”,从而导致产生具有部分或完全人糖基化模式的多肽。参见Gerngross,Nat Biotech 22,1409-1414(2004)和Li等人,Nat Biotech 24,210-215(2006)。
用于表达(糖基化)多肽的合适宿主细胞还来源于多细胞生物(无脊椎动物和脊椎动物)。无脊椎动物细胞的实例包括植物细胞和昆虫细胞。已经鉴定出了可以与昆虫细胞结合使用,特别是用于转染草地夜蛾(Spodoptera frugiperda)细胞的许多杆状病毒株。植物细胞培养物也可用作宿主。参见例如美国专利号5,959,177、6,040,498、6,420,548、7,125,978和6,417,429(描述了用于在转基因植物中产生抗体的PLANTIBODIES
在一个方面,提供了一种生产本发明的双特异性抗原结合分子或其多肽片段的方法,其中所述方法包括在适于表达本发明的双特异性抗原结合分子或其多肽片段的条件下培养包含编码如本文提供的本发明的双特异性抗原结合分子或其多肽片段多核苷酸的宿主细胞,以及从宿主细胞(或宿主细胞培养基)中回收本发明的双特异性抗原结合分子或其多肽片段。
如本文所述制备的本发明的双特异性分子可通过本领域已知的技术纯化,这些技术为诸如高效液相色谱法、离子交换色谱法、凝胶电泳法、亲和色谱法、尺寸排阻色谱法等。用于纯化特定蛋白质的实际条件将部分取决于诸如净电荷、疏水性、亲水性等因素,并且对于本领域技术人员而言将是显而易见的。对于亲和色谱法纯化,可以使用双特异性抗原结合分子结合的抗体、配体、受体或抗原。例如,对于本发明融合蛋白的亲和色谱纯化,可以使用具有蛋白A或蛋白G的基质。基本上如实例中所述,可使用顺序蛋白A或G亲和色谱和尺寸排阻色谱来分离抗原结合分子。双特异性抗原结合分子或其片段的纯度可以通过各种熟知的分析方法中任一者来确定,所述熟知的分析方法包括凝胶电泳法、高压液相色谱法等。例如,实例中所述的表达的双特异性抗原结合分子被显示为得到完整并且适当组装,如还原和非还原SDS-PAGE所示。
测定
本文提供的抗原结合分子可以通过本领域已知的各种测定来表征其结合特性和/或生物学活性。特别地,它们的特征在于实例中更详细描述的测定。
1.结合测定
可以例如通过使用表达人成纤维细胞活化蛋白(FAP)的鼠成纤维细胞系和流式细胞术(FACS)分析来评估本文提供的双特异性抗原结合分子与相应靶标表达细胞的结合。如实例4.2中所述,可以通过使用初级B细胞来确定本文提供的双特异性抗原结合分子与CD40的结合。
2.活性测定
测试本发明的双特异性抗原结合分子的生物学活性。生物学活性可以包括双特异性抗原结合分子的功效和特异性。通过与靶抗原结合后CD40受体的激动性信号传导的测定证明了其功效和特异性。此外,使用已经与双特异性抗原结合分子一起孵育的树突细胞(DC)进行体外T细胞引发测定。
药物组合物、制剂和施用途径
在进一步方面,本发明提供了包含本文提供的双特异性抗原结合分子中任一者的药物组合物,其例如用于以下治疗方法中的任一者中。在一个实施例中,药物组合物包含本文所提供的双特异性抗原结合分子中的任何一者以及至少一种药用赋形剂。在另一实施例中,药物组合物包含本文提供的双特异性抗原结合分子中任一者以及如下文所述的至少一种附加治疗剂。
本发明的药物组合物包含溶于或分散于药用运载体中的治疗有效量的一种或多种双特异性抗原结合分子。术语“药学上或药理学上可接受的”是指分子实体和组合物在所采用的剂量和浓度下通常对接受者无毒,即当视情况而定施用于动物(例如人)时不产生不利的、过敏的或其他不良反应。含有根据本发明所述的至少一种双特异性抗原结合分子和任选地附加活性成分的药物组合物的制备将是鉴于本公开而为本领域的技术人员已知的,如Remington's Pharmaceutical Sciences(第18版,Mack Printing Company,1990)所例示,所述文献以引用方式并入本文。具体地,组合物为冻干制剂或水溶液。如本文所用,“药学上可接受的赋形剂”包括任何和所有溶剂、缓冲剂、分散介质、包衣、表面活性剂、抗氧化剂、防腐剂(例如抗细菌剂、抗真菌剂)、等渗剂、盐、稳定剂及它们的组合,如对本领域的普通技术人员所知。
肠胃外组合物包括设计用于注射(例如,皮下、皮内、病灶内、静脉内、动脉内、肌内、鞘内或腹膜内注射)的那些组合物。为了注射,可在水溶液中配制本发明的双特异性抗原结合分子,优选在生理相容性缓冲剂诸如Hanks溶液、林格氏溶液或生理盐水中配制。溶液可含有配制剂(formulatory agent),诸如悬浮剂、稳定剂和/或分散剂。另选地,双特异性抗原结合分子可以是粉末形式,用于在使用前用合适的媒介物(例如无菌无热原水)构建。根据需要,通过将本发明的抗原结合分子以所需的量与下面列举的各种其他成分一起掺入适当的溶剂中,来制备无菌可注射溶液。例如,无菌可以通过无菌过滤膜过滤而容易地实现。通常,通过将各种灭菌的活性成分掺入含有基础分散介质和/或其他成分的无菌媒介物中来制备分散体。在用于制备无菌可注射溶液、悬浮液或乳液的无菌粉末的情况下,优选的制备方法是真空干燥或冻干技术,所述真空干燥或冻干技术产生来自先前无菌过滤的液体介质的活性成分加上任何附加所需成分的粉末。如果需要的话,液体介质应适当缓冲,并且在注射之前应首先使用足够的盐水或葡萄糖来使液体稀释剂等渗。该组合物必须是在制造和贮存条件下稳定的,并且保存为抗诸如细菌和真菌等微生物的污染作用。应当理解,内毒素污染应以例如低于0.5ng/mg蛋白的安全水平保持最低。合适的药学上可接受的赋形剂包括但不限于:缓冲剂,诸如磷酸盐、柠檬酸盐和其他有机酸;抗氧化剂,包括抗坏血酸和蛋氨酸;防腐剂(诸如十八烷基二甲基苄基氯化铵;氯化六甲双铵;苯扎氯铵;苄索氯铵;苯酚、丁醇或苄醇;对羟基苯甲酸烷基酯,诸如对羟基苯甲酸甲酯或对羟基苯甲酸丙酯;儿茶酚;间苯二酚;环己醇;3-戊醇;间甲酚);低分子量(少于约10个残基)多肽;蛋白质,诸如血清白蛋白、明胶或免疫球蛋白;亲水性聚合物,诸如聚乙烯吡咯烷酮;氨基酸,诸如甘氨酸、谷氨酰胺、天冬酰胺、组氨酸、精氨酸或赖氨酸;单糖、二糖和其他碳水化合物,包括葡萄糖、甘露糖或糊精;螯合剂,诸如EDTA;糖,诸如蔗糖、甘露醇、海藻糖或山梨糖醇;成盐抗衡离子,诸如钠;金属络合物(例如锌蛋白络合物);和/或非离子表面活性剂,诸如聚乙二醇(PEG)。水性注射悬浮液可含有增加悬浮液粘度的化合物,诸如羧甲基纤维素钠、山梨糖醇、葡聚糖等。任选地,悬浮液还可含有合适的稳定剂或增大化合物溶解度的试剂,以允许制备高浓度溶液。另外,活性化合物的悬浮液可以制备成适当的油性注射悬浮液。合适的亲脂性溶剂或载体包括脂肪油,诸如芝麻油;或合成脂肪酸酯,诸如油酸乙酯或甘油三酯;或脂质体。
活性成分可以包埋在例如通过凝聚技术或通过界面聚合而制备的微胶囊(例如分别为羟甲基纤维素或明胶微胶囊和聚(甲基丙烯酸甲酯)微胶囊)中;在胶体药物递送系统(例如,脂质体、白蛋白、微球、微乳液、纳米粒子和纳米胶囊)中;或在粗乳液中。此类技术在Remington's Pharmaceutical Sciences(第18版,Mack Printing Company,1990)中公开。可以制备缓释制备物。缓释制备物的合适示例包括含有多肽的固态疏水聚合物的半透性基质,所述基质是例如膜或微胶囊等成型制品的形式。在特定实施例中,可注射组合物的延长吸收可以通过在所述组合物中使用延迟吸收的试剂(例如单硬脂酸铝、明胶或它们的组合)来实现。
本文的示例性药学上可接受的赋形剂还包括间质药物分散剂,诸如可溶中性活性透明质酸酶糖蛋白(sHASEGP),例如人可溶性PH-20透明质酸酶糖蛋白,诸如rHuPH20(
示例性的冻干抗体制剂描述于美国专利号6,267,958中。水性抗体制剂包括在美国专利号6,171,586和WO2006/044908中描述的那些,后一者中的制剂包含组氨酸-乙酸盐缓冲剂。
除了先前描述的组合物之外,抗原结合分子也可以配制成长效制备物。此类长效制剂可以通过植入(例如皮下或肌内植入)或通过肌内注射施用。因此,例如,融合蛋白可以用合适的聚合或疏水材料(例如配制成可接受的油中的乳剂)或离子交换树脂配制,或配制成微溶衍生物(例如配制成微溶盐)。
包含本发明的双特异性抗原结合分子的药物组合物可通过常规的混合、溶解、乳化、包封、包埋或冻干工艺进行生产。药物组合物可以使用一种或多种生理上可接受的运载体、稀释剂、赋形剂或助剂以常规方式配制,所述运载体、稀释剂、赋形剂或助剂有助于将蛋白质加工成可以在药学上使用的制备物。合适的制剂取决于所选择的施用途径。
双特异性抗原结合分子可以配制成以游离酸或碱、中性或盐形式的组合物。药学上可接受的盐是基本上保留游离酸或游离碱的生物活性的盐。这些药学上可接受的盐包括酸加成盐,例如用蛋白质性质组合物的游离氨基形成的酸加成盐,或用无机酸(诸如盐酸或磷酸)或有机酸(诸如乙酸、草酸、酒石酸或扁桃酸)形成的酸加成盐。用游离羧基形成的盐也可以衍生自无机碱,诸如氢氧化钠、氢氧化钾、氢氧化铵、氢氧化钙或氢氧化铁;或者有机碱,诸如异丙胺、三甲胺、组氨酸或普鲁卡因。与相应的游离碱形式相比,药用盐倾向于更易溶于水性和其他质子溶剂中。
本文的组合物还可含有多于一种对于所治疗的特定适应症是必需的活性成分,优选是具有不会彼此不利地影响的互补活性的活性成分。此类活性成分适当地以对预期目的有效的量组合存在。
待用于体内施用的制剂通常是无菌的。例如,无菌可以通过无菌过滤膜过滤而容易地实现。
治疗方法和组合物
本文提供的双特异性抗原结合分子中任一者可用于治疗方法中。为了用于治疗方法中,本发明的双特异性抗原结合分子可按照符合良好医学实践的方式配制、投配和施用。在这种情况下需要考虑的因素包括所治疗的特定疾患、所治疗的特定哺乳动物、个体患者的临床病症、疾患的原因、药剂的递送部位、施用方法、施用的时间安排,以及执业医师已知的其他因素。
在一个方面,将本发明的双特异性抗原结合分子提供为用作药物。
在进一步方面,提供了本发明的双特异性抗原结合分子,其用于(i)诱导通过CD40+抗原呈递细胞(APC)的免疫刺激,(ii)刺激肿瘤特异性T细胞应答,(iii)引起肿瘤细胞凋亡,(iv)治疗癌症,(v)延迟癌症进展,(vi)延长癌症患者的生存期,(vii)治疗感染。在某一特定方面,提供了用于治疗疾病特别是用于治疗癌症的本发明的双特异性抗原结合分子。
在某些方面,提供了用于治疗方法中的本发明的双特异性抗原结合分子。在一个方面,本发明提供了一种如本文所述的双特异性抗原结合分子,其用于治疗有此需要的个体的疾病。在某些方面,本发明提供了一种用于治疗患有疾病的个体的方法中的双特异性抗原结合分子,该方法包括向个体施用治疗有效量的双特异性抗原结合分子。在某些方面,待治疗的疾病是癌症。需要治疗的受试者、患者或“个体”通常是哺乳动物,更特别地是人。
在一个方面,提供了一种方法,其用于i)诱导通过CD40+抗原呈递细胞(APC)的免疫刺激,(ii)刺激肿瘤特异性T细胞应答,(iii)引起肿瘤细胞凋亡,(iv)治疗癌症,(v)延迟癌症进展,(vi)延长癌症患者的生存期或(vii)治疗感染,其中该方法包括向有此需要的个体施用治疗有效量的本发明的双特异性抗原结合分子。
在进一步方面,本发明提供了本发明的双特异性抗原结合分子用于生产或制备用于治疗有此需要的个体的疾病的药物的用途。在一个方面,药物用于治疗疾病的方法中,该方法包括向具有所述疾病的个体施用治疗有效量的所述药物。在某些方面中,待治疗的疾病是增殖性病症,特别是癌症。癌症的示例包括但不限于膀胱癌、脑癌、头颈癌、胰腺癌、肺癌、乳腺癌、卵巢癌、子宫癌、宫颈癌、子宫内膜癌、食道癌、结肠癌、结直肠癌、直肠癌、胃癌、前列腺癌、血癌、皮肤癌、鳞状细胞癌、骨癌以及肾癌。癌症的其他示例包括恶性肿瘤、淋巴瘤(例如,霍奇金淋巴瘤和非霍奇金淋巴瘤)、母细胞瘤、肉瘤和白血病。可以使用本发明的双特异性抗原结合分子或抗体治疗的其他细胞增殖疾患包括但不限于位于以下部位中的肿瘤:腹部、骨骼、乳腺、消化系统、肝脏、胰腺、腹膜、内分泌腺(肾上腺、甲状旁腺、垂体、睾丸、卵巢、胸腺、甲状腺)、眼、头颈部、神经系统(中枢神经系统和外周神经系统)、淋巴系统、骨盆、皮肤、软组织、脾脏、胸部以及泌尿生殖系统。还包括癌前病症或病变和癌转移。在某些实施例中,癌症选自由以下项组成的组:肾细胞癌、皮肤癌、肺癌、结直肠癌、乳腺癌、脑癌、头颈癌。本领域的技术人员易于认识到,在许多情况下,本发明的双特异性抗原结合分子或抗体可能无法治愈,但可能提供益处。在一些方面,具有某些益处的生理变化也被视为具有治疗益处。因此,在一些方面,提供生理变化的本发明的双特异性抗原结合分子或抗体的量被视为“有效量”或“治疗有效量”。
为了预防或治疗疾病,本发明的双特异性抗原结合分子的适当剂量(当单独使用或与一种或多种附加治疗剂联合使用时)将取决于待治疗疾病的类型、施用途径、患者体重、具体分子、疾病的严重程度和进程、本发明的双特异性抗原结合分子是出于预防还是治疗目的施用、先前或同时进行的治疗性干预、患者的临床病史和对双特异性抗原结合分子的应答以及主治医师的酌处权。在任何情况下,负责施用的执业者将针对个体受试者来确定组合物中活性成分的浓度和适当剂量。本文考虑了各种投配时间安排,包括但不限于在各个时间点处的单次或多次施用、推注施用,以及脉冲输注。
本发明的双特异性抗原结合分子适于在一次或在一系列治疗中施用于患者。取决于疾病的类型和严重性,约1μg/kg至15mg/kg(例如0.1mg/kg-10mg/kg)的双特异性抗原结合分子可以是例如通过一次或多次单独施用或通过连续输注而施用于患者的初始候选剂量。取决于上述因素,一种典型的日剂量的范围可以为约1μg/kg至100mg/kg或更多。对于数天或更长时间的重复施用,取决于病症,治疗通常会持续直至发生所需的疾病症状抑制。本发明的双特异性抗原结合分子的一种示例性剂量的范围为约0.005mg/kg至约10mg/kg。在其他示例中,剂量还可包括每次施用约1μg/kg体重、约5μg/kg体重、约10μg/kg体重、约50μg/kg体重、约100μg/kg体重、约200μg/kg体重、约350μg/kg体重、约500μg/kg体重、约1mg/kg体重、约5mg/kg体重、约10mg/kg体重、约50mg/kg体重、约100mg/kg体重、约200mg/kg体重、约350mg/kg体重、约500mg/kg体重至约1000mg/kg体重或更多,以及其中可推导出的任何范围。在从本文所列数字可推导出的范围的示例中,可以基于上述数字施用约0.1mg/kg体重至约20mg/kg体重、约5μg/kg体重至约1mg/kg体重等的范围。因此,可以向患者施用约0.5mg/kg、2.0mg/kg、5.0mg/kg或10mg/kg(或它们的任何组合)的一种或多种剂量。此类剂量可以间歇地施用,例如每周或每三周施用(例如,使得患者接受约2至约20或例如约6个剂量的融合蛋白)。在一个特定方面,双特异性抗原结合分子将每三周施用一次。可施用初始较高负荷剂量,然后施用一种或多种较低剂量。然而,其他剂量方案可能有用。该疗法的进展通过常规技术和测定而容易地监测。
本发明的双特异性抗原结合分子将通常以有效实现预期目的量使用。为用于治疗或预防病症,将本发明的双特异性抗原结合分子或其药物组合物以治疗有效量施用或施加。治疗有效量的确定完全在本领域技术人员的能力范围内,特别是根据本文提供的详细公开内容。对于全身施用,最初可以根据体外测定(诸如细胞培养测定)来估计治疗有效剂量。可以随后在动物模型中配制剂量,以实现包括如在细胞培养中测定的IC
剂量和间隔可以单独地调节,以提供足以维持疗效的本发明的双特异性抗原结合分子的血浆水平。通过注射施用的常用患者剂量的范围是约0.1至50mg/kg/天,通常约0.1至1mg/kg/天。通过每天施用多个剂量可以实现治疗有效的血浆水平。可以例如通过HPLC来测量血浆中的水平。在局部施用或选择性摄入的情况下,本发明的双特异性抗原结合分子或抗体的有效局部浓度可能与血浆浓度无关。本领域的技术人员将能够在无需过多实验的情况下优化治疗有效的局部剂量。
本文所述的治疗有效剂量的本发明的双特异性抗原结合分子通常将提供治疗益处而不会引起显著的毒性。融合蛋白的毒性和治疗功效可通过细胞培养或实验动物中的标准药学程序来测定。细胞培养测定和动物研究可以用于测定LD
用本发明的融合蛋白治疗的患者的主治医师将知道如何以及何时由于毒性、器官功能障碍等终止、中断或调节施用。相反地,如果临床应答不充分(排除毒性),则主治医师也会知道将治疗调节到更高水平。在感兴趣的病症的管理中施用的剂量的大小将随着待治疗病症的严重程度、施用途径等而变化。例如,可以部分地通过标准预后评估方法来评估病症的严重性。此外,剂量和可能的剂量频率也将根据个体患者的年龄、体重和应答而变化。
其他药剂和治疗
本发明的双特异性抗原结合分子可在治疗中与一种或多种其他药剂联合施用。例如,本发明的双特异性抗原结合分子或抗体可与至少一种附加治疗剂联合施用。术语“治疗剂”包括可以施用用于治疗需要此类治疗的个体的症状或疾病的任何药剂。此类附加治疗剂可包含适合于所治疗的具体适应症的任何活性成分,优选地是具有不会彼此不利地影响的互补活性的活性成分。在某些实施例中,另外的治疗剂是另一种抗癌剂,例如微管破坏剂、抗代谢物、拓扑异构酶抑制剂、DNA嵌入剂、烷化剂、激素疗法、激酶抑制剂、受体拮抗剂、肿瘤细胞凋亡活化剂,或抗血管生成剂。在某些方面,附加治疗剂为免疫调节剂、细胞生长抑制剂、细胞粘附抑制剂、细胞毒性剂或细胞抑制剂、细胞凋亡活化剂或增加细胞对细胞凋亡诱导剂的敏感性的药剂。
因此,提供了本发明的双特异性抗原结合分子或包含它们的药物组合物,其用于治疗癌症,其中双特异性抗原结合分子与化学治疗剂、放射和/或用于癌症免疫疗法的其他药剂联合施用。
此类其他药剂适当地以对预期目的有效的量组合存在。此类其他药剂的有效量取决于所用融合蛋白的量、病症或治疗的类型,以及上面讨论的其他因素。本发明的双特异性抗原结合分子或抗体通常以与本文所述相同的剂量和施用途径,或本文所述剂量的约1%至99%,或以任何剂量且通过经验/临床上确定为合适的途径使用。
上述此类联合疗法涵盖联合施用(其中两种或更多种治疗剂被包含在同一组合物或单独的组合物中),以及单独施用,在分开施用的情况下,本发明的双特异性抗原结合分子或抗体的施用可以在施用附加治疗剂和/或佐剂之前、同时和/或之后发生。
在进一步方面,提供了如上文所述的双特异性抗原结合分子,其用于治疗癌症,其中双特异性抗原结合分子与另一种免疫调节剂联合施用。
术语“免疫调节剂”是指任何影响免疫系统的物质,包括单克隆抗体。本发明的分子可以被视为免疫调节剂。免疫调节剂可用作抗肿瘤剂治疗癌症。在一个方面,免疫调节剂包括但不限于抗CTLA4抗体(例如伊匹木单抗)、抗PD1抗体(例如纳武单抗或派姆单抗)、PD-L1抗体(例如阿特珠单抗、阿维单抗或德瓦鲁单抗)、OX-40抗体、4-1BB抗体和GITR抗体。在进一步方面,提供了如上文所述的双特异性抗原结合分子,其用于治疗癌症,其中双特异性抗原结合分子与阻断PD-L1/PD-1相互作用的药剂联合施用。在一个方面,阻断PD-L1/PD-1相互作用的药剂为抗PD-L1抗体或抗PD1抗体。更特别地,阻断PD-L1/PD-1相互作用的药剂为抗PD-L1抗体,特别是选自由阿特珠单抗、德瓦鲁单抗、派姆单抗和纳武单抗组成的组中的抗PD-L1抗体。在一个具体方面,阻断PD-L1/PD-1相互作用的药剂为阿特珠单抗(MPDL3280A,RG7446)。在另一方面,阻断PD-L1/PD-1相互作用的药剂为抗PD-L1抗体,该抗体包含SEQ ID NO:107的重链可变结构域VH(PDL-1)和SEQ ID NO:108的轻链可变结构域VL(PDL-1)。在另一方面,阻断PD-L1/PD-1相互作用的药剂为抗PD-L1抗体,该抗体包含SEQ IDNO:109的重链可变结构域VH(PDL-1)和SEQ ID NO:110的轻链可变结构域VL(PDL-1)。在另一方面,阻断PD-L1/PD-1相互作用的药剂为抗PD1抗体,特别是选自派姆单抗或纳武单抗的抗PD1抗体。此类其他药剂适当地以对预期目的有效的量组合存在。此类其他药剂的有效量取决于所用双特异性抗原结合分子的量、病症或治疗的类型,以及上面讨论的其他因素。如上文所述的双特异性抗原结合分子通常以与本文所述相同的剂量和施用途径,或本文所述剂量的约1%至99%,或以任何剂量且通过经验/临床上确定为合适的途径使用。
上述此类联合疗法涵盖联合施用(其中两种或更多种治疗剂被包含在同一组合物或单独的组合物中),以及单独施用,在分开施用的情况下,本发明的双特异性抗原结合分子的施用可以在施用附加治疗剂和/或佐剂之前、同时和/或之后发生。
制品
在本发明的另一个方面中,提供了一种制品,其含有可用于治疗、预防和/或诊断上述病症的物质。该制品包括容器和在所述容器上或与所述容器相关的标签或包装插页(package insert)。合适的容器包括例如瓶子、小瓶、注射器、IV溶液袋等。所述容器可以由诸如玻璃或塑料等多种材料形成。容器容纳组合物,该组合物单独或与另一种有效用于治疗、预防和/或诊断所述病症的组合物组合,并且容器可具有无菌入口(例如容器可以是静脉注射溶液袋或具有可用皮下注射针刺穿的塞子的小瓶)。组合物中的至少一种活性剂是本发明的双特异性抗原结合分子。
标签或包装插页指示该组合物用于治疗所选择的病症。此外,制品可包括(a)第一容器,所述第一容器中含有包含本发明的双特异性抗原结合分子的组合物;以及(b)第二容器,所述第二容器中含有包含进一步的细胞毒性剂或其他治疗剂组合物。本发明该实施例中的制品还可包含包装插页,该包装插页指示组合物可用于治疗特定病症。
另选地或另外地,制品还可包括第二(或第三)容器,该二(或第三)容器包括药学上可接受的缓冲液,诸如抑菌性注射用水(BWFI)、磷酸盐缓冲盐水、林格氏溶液和葡萄糖溶液。制品还可以包括从商业和用户角度所需的其他物质,包括其他缓冲剂、稀释剂、过滤器、针头和注射器。
表B(序列):
以下编号的段落描述了本发明的各方面:
1.一种双特异性抗原结合分子,包含:
(a)至少一个能够与CD40特异性结合的抗原结合结构域;以及
(b)至少一个能够与成纤维细胞活化蛋白(FAP)特异性结合的抗原结合结构域,其包含:重链可变区(V
(i)CDR-H1,其包含SEQ ID NO:3的氨基酸序列,(ii)CDR-H2,其包含选自由SEQ IDNO:4、SEQ ID NO:11和SEQ ID NO:12组成的组的氨基酸序列,和(iii)CDR-H3,其包含SEQID NO:5的氨基酸序列;以及轻链可变区(V
2.根据段落1所述的双特异性抗原结合分子,另外包含(c)Fc区,其由能够稳定缔合的第一亚基和第二亚基组成。
3.根据段落1或2所述的双特异性抗原结合分子,其中能够与FAP特异性结合的抗原结合结构域包含:重链可变区(V
4.根据段落1至3中任一项所述的双特异性抗原结合分子,其中能够与FAP特异性结合的抗原结合结构域包含:
重链可变区(V
5.根据段落1至4中任一项所述的双特异性抗原结合分子,其中能够与FAP特异性结合的抗原结合结构域包含:
(a)重链可变区(V
(b)重链可变区(V
(c)重链可变区(V
(d)重链可变区(V
6.根据段落1至5中任一项所述的双特异性抗原结合分子,其中能够与CD40特异性结合的抗原结合结构域包含:重链可变区(V
7.根据段落1至6中任一项所述的双特异性抗原结合分子,其中能够与CD40特异性结合的抗原结合结构域包含:
(i)重链可变区(V
(ii)轻链可变区(V
8.根据段落1至6中任一项所述的双特异性抗原结合分子,其中能够与CD40特异性结合的抗原结合结构域包含:
(i)重链可变区(V
(ii)轻链可变区(V
9.根据段落1至5或权利要求7中任一项所述的双特异性抗原结合分子,其中能够与CD40特异性结合的抗原结合结构域包含:
(a)VH,其包含SEQ ID NO:37的氨基酸序列,和VL,其包含SEQ ID NO:41的氨基酸序列,或者
(b)VH,其包含SEQ ID NO:37的氨基酸序列,和VL,其包含SEQ ID NO:42的氨基酸序列,或者
(c)VH,其包含SEQ ID NO:37的氨基酸序列,和VL,其包含SEQ ID NO:43的氨基酸序列,或者
(d)VH,其包含SEQ ID NO:37的氨基酸序列,和VL,其包含SEQ ID NO:44的氨基酸序列,或者
(e)VH,其包含SEQ ID NO:38的氨基酸序列,和VL,其包含SEQ ID NO:41的氨基酸序列,或者
(f)VH,其包含SEQ ID NO:38的氨基酸序列,和VL,其包含SEQ ID NO:42的氨基酸序列,或者
(g)VH,其包含SEQ ID NO:38的氨基酸序列,和VL,其包含SEQ ID NO:43的氨基酸序列,或者
(h)VH,其包含SEQ ID NO:38的氨基酸序列,和VL,其包含SEQ ID NO:44的氨基酸序列,或者
(i)VH,其包含SEQ ID NO:39的氨基酸序列,和VL,其包含SEQ ID NO:41的氨基酸序列,或者
(j)VH,其包含SEQ ID NO:39的氨基酸序列,和VL,其包含SEQ ID NO:42的氨基酸序列,或者
(k)VH,其包含SEQ ID NO:39的氨基酸序列,和VL,其包含SEQ ID NO:43的氨基酸序列,或者
(l)VH,其包含SEQ ID NO:39的氨基酸序列,和VL,其包含SEQ ID NO:44的氨基酸序列,或者
(m)VH,其包含SEQ ID NO:40的氨基酸序列,和VL,其包含SEQ ID NO:41的氨基酸序列,或者
(n)VH,其包含SEQ ID NO:40的氨基酸序列,和VL,其包含SEQ ID NO:42的氨基酸序列,或者
(o)VH,其包含SEQ ID NO:40的氨基酸序列,和VL,其包含SEQ ID NO:43的氨基酸序列,或者
(p)VH,其包含SEQ ID NO:40的氨基酸序列,和VL,其包含SEQ ID NO:44的氨基酸序列。
10.根据段落1至5或权利要求7或9中任一项所述的双特异性抗原结合分子,其中能够与CD40特异性结合的抗原结合结构域包含:VH,其包含SEQ ID NO:37的氨基酸序列,和VL,其包含SEQ ID NO:41的氨基酸序列。
11.根据段落1至5或权利要求8中任一项所述的双特异性抗原结合分子,其中能够与CD40特异性结合的抗原结合结构域包含:
(a)VH,其包含SEQ ID NO:45的氨基酸序列,和VL,其包含SEQ ID NO:51的氨基酸序列,或者
(b)VH,其包含SEQ ID NO:46的氨基酸序列,和VL,其包含SEQ ID NO:51的氨基酸序列,或者
(c)VH,其包含SEQ ID NO:47的氨基酸序列,和VL,其包含SEQ ID NO:51的氨基酸序列,或者
(d)VH,其包含SEQ ID NO:48的氨基酸序列,和VL,其包含SEQ ID NO:51的氨基酸序列,或者
(e)VH,其包含SEQ ID NO:45的氨基酸序列,和VL,其包含SEQ ID NO:52的氨基酸序列,或者
(f)VH,其包含SEQ ID NO:46的氨基酸序列,和VL,其包含SEQ ID NO:52的氨基酸序列,或者
(g)VH,其包含SEQ ID NO:47的氨基酸序列,和VL,其包含SEQ ID NO:52的氨基酸序列,或者
(h)VH,其包含SEQ ID NO:48的氨基酸序列,和VL,其包含SEQ ID NO:52的氨基酸序列,或者
(i)VH,其包含SEQ ID NO:49的氨基酸序列,和VL,其包含SEQ ID NO:53的氨基酸序列,或者
(j)VH,其包含SEQ ID NO:50的氨基酸序列,和VL,其包含SEQ ID NO:53的氨基酸序列,或者
(k)VH,其包含SEQ ID NO:49的氨基酸序列,和VL,其包含SEQ ID NO:54的氨基酸序列,或者
(l)VH,其包含SEQ ID NO:50的氨基酸序列,和VL,其包含SEQ ID NO:54的氨基酸序列。
12.根据段落1至5或段落8或11中任一项所述的双特异性抗原结合分子,其中能够与CD40特异性结合的抗原结合结构域包含:VH,其包含SEQ ID NO:45的氨基酸序列,和VL,其包含SEQ ID NO:51的氨基酸序列,或者其中能够与CD40特异性结合的抗原结合结构域包含:VH,其包含SEQ ID NO:48的氨基酸序列,和VL,其包含SEQ ID NO:51的氨基酸序列。
13.根据段落1至7中任一项所述的双特异性抗原结合分子,其包含:
(i)至少一个能够与CD40特异性结合的抗原结合结构域,其包含:重链可变区(V
(ii)至少一个能够与FAP特异性结合的抗原结合结构域,其包含:重链可变区(V
14.根据段落2至13中任一项所述的双特异性抗原结合分子,其中Fc区为IgG区,特别地为IgG1 Fc区或IgG4 Fc区,并且其中Fc区包含一个或多个氨基酸取代,其降低抗体与Fc受体的结合亲和力和/或效应子功能。
15.根据段落2至14中任一项所述的双特异性抗原结合分子,其中Fc区为人IgG1亚类,具有氨基酸突变L234A、L235A和P329G(根据Kabat EU索引编号)。
16.根据段落1至15中任一项所述的双特异性抗原结合分子,其中双特异性抗原结合分子包含
(a)至少两个能够与CD40特异性结合的Fab片段,其连接至Fc区,
以及
(b)一个能够与FAP特异性结合的抗原结合结构域,其连接至所述Fc区的C末端。
17.根据段落1至16中任一项所述的双特异性抗原结合分子,其中双特异性抗原结合分子包含
(a)至少两个能够与CD40特异性结合的Fab片段,其融合至Fc区,
以及
(b)能够与FAP特异性结合的交叉fab片段,其融合至所述Fc区的C末端。
18.根据段落17所述的双特异性抗原结合分子,其中能够与FAP特异性结合的交叉fab片段的VH-Cκ链融合至所述Fc区的所述C末端。
19.根据段落1至18中任一项所述的双特异性抗原结合分子,其中双特异性抗原结合分子包含四个能够与CD40特异性结合的Fab片段。
20.一种与FAP特异性结合的抗体,其中所述抗体包含:重链可变区(V
21.根据段落20所述的抗体,其中所述抗体包含
(a)重链可变区(V
(b)重链可变区(V
(c)重链可变区(V
(d)重链可变区(V
22.分离的核酸,其编码根据段落1至19中任一项所述的双特异性抗原结合分子或根据段落20或21所述的抗体。
23.一种表达载体,其包含根据段落22所述的分离的核酸。
24.一种宿主细胞,其包含根据段落22所述的分离的核酸或根据段落23所述的表达载体。
25.一种生产双特异性抗原结合分子的方法,其包含在适合于表达所述双特异性抗原结合分子的条件下培养根据段落24所述的宿主细胞,并且分离所述双特异性抗原结合分子。
26.一种药物组合物,其包含根据段落1至19中任一项所述的双特异性抗原结合分子或根据段落20或21所述的抗体以及药用运载体。
25.根据段落24所述的药物组合物,进一步包含附加治疗剂。
26.根据段落1至19中任一项所述的双特异性抗原结合分子或根据段落24所述的药物组合物,其用作药物。
27.根据段落1至19中任一项所述的双特异性抗原结合分子或根据段落24所述的药物组合物,其用于以下各项:
(i)通过表达CD40的抗原呈递细胞(APC)诱导免疫刺激,
(ii)刺激肿瘤特异性T细胞应答,
(iii)引起肿瘤细胞凋亡,
(iv)治疗癌症,
(v)延缓癌症发展,
(vi)延长癌症患者的存活期,
(vii)治疗感染。
28.根据段落1至19中任一项所述的双特异性抗原结合分子或根据段落24所述的药物组合物,其用于治疗癌症。
29.根据段落1至19中任一项所述的双特异性抗原结合分子或根据段落24所述的药物组合物在制造用于治疗癌症的药物中的用途。
30.一种治疗患有癌症的个体的方法,其包括向所述个体施用有效量的根据段落1至19中任一项所述的双特异性抗原结合分子或根据段落24所述的药物组合物。
31.根据段落1至19中任一项所述的双特异性抗原结合分子或根据段落24所述的药物组合物,其用于治疗癌症,其中所述双特异性抗原结合分子与化学治疗剂、放射和/或用于癌症免疫疗法的其他药剂联合施用。
实例
以下是本发明的方法和组合物的实例。应当理解,在给出以上提供的一般描述的情况下,可以实践各种其他实施例。
重组DNA技术
使用标准方法来操纵DNA,如在Sambrook等人,Molecular cloning:A laboratorymanual;Cold Spring Harbor Laboratory Press,Cold Spring Harbor,New York,1989中所述。根据制造商的说明来使用分子生物学试剂。关于人免疫球蛋白轻链和重链的核苷酸序列的一般信息在以下参考文献中给出:Kabat,E.A.等人,(1991)Sequences of Proteinsof Immunological Interest,第五版,NIH Publication No 91-3242。
DNA测序
通过双链测序确定DNA序列。
基因合成
通过使用适当模板进行PCR生成所需的基因区段,或由Geneart AG(Regensburg,Germany)通过自动化基因合成从合成寡核苷酸和PCR产物合成所需的基因区段。在确切的基因序列不可用的情况下,基于最接近的同源物的序列设计寡核苷酸引物,并且通过RT-PCR从来源于合适的组织的RNA中分离出基因。将侧接有单个限制性内切酶切割位点的基因区段克隆到标准克隆/测序载体中。从转化的细菌中纯化出质粒DNA,并通过紫外光谱法确定浓度。通过DNA测序来确认亚克隆基因片段的DNA序列。设计具有合适限制性位点的基因区段,以允许亚克隆到相应的表达载体中。所有构建体均设计有5’端DNA序列,该序列编码前导肽,该前导肽靶向真核细胞分泌的蛋白。
蛋白质纯化
参照标准方案从过滤的细胞培养物上清液中纯化蛋白质。简言之,将抗体施加至蛋白质A琼脂糖柱(GE healthcare)并用PBS洗涤。在pH 2.8实现抗体的洗脱,之后立即中和样品。通过尺寸排阻色谱法(Superdex 200,GE Healthcare)在PBS中或在20mM组氨酸、150mM NaCl(pH 6.0)中将聚集蛋白质与单体抗体分离。将单体抗体级分合并,使用例如MILLIPORE Amicon Ultra(30MWCO)离心浓缩器浓缩(若需要),冷冻并储存在-20℃或-80℃。提供样品的部分以例如通过SDS-PAGE、尺寸排阻色谱(SEC)或质谱法来进行后续的蛋白质分析和分析表征。
SDS-PAGE
根据制造商的说明使用
CE-SDS
通过CE-SDS使用微流控Labchip技术分析双特异性和对照抗体的纯度、抗体完整性和分子量(Caliper Life Science,USA)。使用HT Protein Express试剂盒根据制造商的说明制备5μl蛋白溶液用于CE-SDS分析,并使用HT Protein Express芯片在LabChip GXII系统上进行分析。使用LabChip GX软件3.0.618.0版分析数据。
分析型尺寸排阻色谱
用于测定抗体的聚集和低聚物状态的尺寸排阻色谱法(SEC)通过HPLC色谱法进行。简而言之,将蛋白A纯化的抗体施加至Agilent HPLC1100系统上的300mM NaCl、50mMKH
质谱
本节描述了对具有VH/VL或CH/CL交换(CrossMab)的多特异性抗体的表征,重点在于所述多特异性抗体的正确装配。通过对脱糖基化的完整CrossMab和脱糖基化/FabALACTICA或者另选地脱糖基化/GingisKHAN消化的CrossMab进行电喷雾电离质谱法(ESI-MS),来分析预期的一级结构。
在37℃以1mg/ml的蛋白质浓度将CrossMab用磷酸盐或Tris缓冲液中的N-糖苷酶F脱糖基化至多17h。FabALACTICA或GingisKHAN(Genovis Ab;瑞典)消化是在供应商供应的缓冲液中进行,该缓冲液中含有100μg去糖基化的CrossMab。在质谱分析之前,将样品经由HPLC在Sephadex G25柱(GE Healthcare)上脱盐。在配备有TriVersa NanoMate源(Advion)的maXis 4G UHR-QTOF MS系统(Bruker Daltonik)上经由ESI-MS 来测定总质量。
实例1
产生抗成纤维细胞活化蛋白(FAP)的新抗体
1.1小鼠的免疫接种
Balb/c和NMRI小鼠用于免疫接种。根据附录A“动物的住宿和照料指南”将动物圈养在AAALACi认可的动物设施中。所有动物免疫接种方案和实验均已获得上巴伐利亚行政区政府批准(许可号55.2-1-54-2531-19-10),并根据德国动物福利法和欧洲议会和理事会指令2010/63进行。6-8周龄的Balb/c和NMRI小鼠(n=5)接受了共价连接至His标签(SEQ IDNO:78)的人成纤维细胞活化蛋白α(氨基酸27-759;登录号NP_004451)的细胞外结构域的四轮免疫接种。在每次免疫接种之前,用氧气和异氟烷的混合气体麻醉小鼠。第一次免疫接种时,将溶于pH 7.4的PBS中的30μg蛋白与等体积的CFA(BD Difco,#263810)混合并腹膜内(i.p.)施用。在第6周皮下(s.c.)施用另外10μg在Abisco佐剂中乳化的蛋白。在第10周,腹膜内施用第三剂不含佐剂的5μg蛋白。最后,在使用杂交瘤技术制备用于抗体开发的脾细胞之前三天,用50μg蛋白对小鼠进行静脉内(i.v.)加强免疫接种。通过ELISA测试血清的抗原特异性总IgG抗体产生。最后一次免疫接种后三天,对小鼠实施安乐死,并无菌分离脾脏并准备用于杂交瘤产生。分离小鼠淋巴细胞并使用基于PEG的标准方案与小鼠骨髓瘤细胞系融合以产生杂交瘤。将得到的杂交瘤细胞以约10
杂交瘤的培养:在37℃和5%CO
1.2抗huFAP抗体与FAP克隆4B9和28H1的竞争性细胞结合
与FAP克隆4B9相比,测试了所得克隆的结合行为。FAP克隆4B9和28H1的产生和制备描述于WO 2012/020006 A2中,所述专利以引用方式并入本文。为了确定鼠FAP克隆是否将不同的表位识别为克隆4B9和28H1,进行了与在转染的HEK细胞上表达的人FAP的竞争结合。
简而言之,用细胞解离缓冲液收获靶细胞,用FACS缓冲液(PBS+2%FCS+5mM EDTA+0.25%钠酸)洗涤,并接种到96-U底板中(1x10
如图2A和图2B中可以看出,鉴定了10种杂交瘤衍生的鼠抗体(命名为克隆209、210、211、212、213、214、215、216、217和218),它们没有竞争与抗FAP抗体4B9或28H1结合。
1.3抗huFAP鼠抗体的靶标结合特异性
成纤维细胞活化蛋白(FAP,FAP-α,seprase)是II型跨膜丝氨酸蛋白酶,其属于脯氨酰寡肽酶家族。该家族包含丝氨酸蛋白酶,其在脯氨酸残基之后优先切割肽。在人蛋白组中表达的该家族的其他重要成员为脯氨酰寡肽酶(PREP)和二肽基肽酶(DPP)。DPP-IV为FAP的最接近同源物。与FAP相反,DPP-IV普遍表达,并在各种生物学过程(诸如T细胞共刺激、趋化因子生物学、葡萄糖代谢和肿瘤发生)中起作用,因此所需的抗人FAP抗体不应与人DPP-IV结合。
使用人FAP或人DPPIV转染的HEK细胞通过流式细胞术确定与人FAP和人DPP-IV的结合。简而言之,用细胞解离缓冲液收获靶细胞,用FACS缓冲液(PBS+2%FCS+5mM EDTA+0.25%钠酸)洗涤,并接种到96-U底板中(1x10
1.4huIgG1_LALA_PG形式的抗huFAP抗体的产生
用标准测序方法确定了新抗huFAP抗体的DNA序列。基于VH和VL结构域,根据WO2012/130831 A1中所述的方法,将新抗FAP抗体表达为具有效应子沉默Fc(P329G;L234,L235A)的huIgG1抗体,以消除与Fc受体的结合。详细地,通过与编码不同肽链的表达载体悬浮生长的HEK293-F细胞的瞬时转染来表达抗体。根据细胞供应商的说明,使用抗体载体的Maxiprep(Qiagen,Germany)制剂,基于F17的培养基(Invitrogen,USA),PEIpro(Polyscience Europe GmbH)和初始细胞密度为1-2百万活细胞/ml,在不含血清的FreeStyle 293表达培养基(Invitrogen)中,进行至HEK293-F细胞(Invitrogen,USA)中的转染。在摇瓶或搅拌的发酵罐中培养7天后,通过以14000g离心30分钟收集细胞培养上清液,并且通过0.22μm过滤器过滤。
使用MabSelectSure-Sepharose
1.5抗huFAP抗体的细胞结合
具有人IgG1 P329G LALA Fc的抗FAP抗体与人FAP的结合是通过流式细胞术使用人FAP转染的HEK细胞确定的。简而言之,用细胞解离缓冲液收获靶细胞,用FACS缓冲液(PBS+2%FCS+5mM EDTA+0.25%钠酸)洗涤,并接种到96-U底板中(1x10
如先前所见,所有抗FAP抗体显示出与人FAP的相似结合。所选结合物的EC
表1:抗FAP抗体与huFAP表达细胞的细胞结合
1.6抗huFAP抗体的细胞内化
使用人FAP转染的HEK细胞作为靶标,确定FAP结合物的内化。简而言之,用细胞解离缓冲液收获靶细胞,用冷FACS缓冲液(PBS+2%FCS+5mM EDTA+0.25%钠酸)洗涤,并以1.5x 10
由标记的次级抗体引起的信号随时间保持几乎恒定,这意味着随着时间的推移未观察到抗体的丢失,所有测试的抗hu FAP抗体均未被内化。
1.7抗huFAP抗体的结合动力学
为了评估人FAP结合动力学,根据制造商的说明,将生物素化的人FAP固定在S系列Biacore CAPture芯片(GE Healthcare 28-9202-34)上,产生大约20个共振单位(RU)的表面密度。使用HBS-P+(10mM HEPES,150mM NaCl pH 7.4,0.05%表面活性剂P20)作为运行和稀释缓冲液。依次注射一系列稀释的抗huFAP Fab(3.7–300nM,1:3稀释),每次120s,以30μl/分钟的流速(单循环动力学)监测解离1800s。通过注射6M盐酸胍、0.25M NaOH 120s使表面再生。通过减去空白注射并通过减去从没有捕获人FAP的对照流通池获得的应答来校正大折射率偏差。使用Biacore评估软件中的1:1Langmuir结合模型进行曲线拟合。亲和力数据示出于下表2中。
表2:如通过Biacore测量的抗FAP Fab对人FAP的亲和力
1.8抗huFAP克隆的形式依赖性结合
为了确定抗FAP克隆在C末端融合至Fc结构域时是否不会丧失其结合特性,构建体包含Fc突起链和Fc孔链,其中VH结构域融合至Fc突起链的C末端并且VL结构域融合至Fc孔链的C末端(图3A,C末端VH/VL融合),以及构建体包含Fc突起链和Fc孔链,其中整个Fab以VH结构域融合至Fc突起链的C末端(图3B,C末端Fab融合)。Fc突起链具有SEQ ID NO:97的氨基酸序列,并且Fc孔链具有SEQ ID NO:98的氨基酸序列。
与抗体相比,构建体对生物素化的重组人FAP和生物素化的重组食蟹猴FAP的亲和力示出于下表3中。
表3:如通过Biacore测量的对人FAP和食蟹猴FAP的亲和力
如上文所述,还已经确定了构建体与FAP转染的HEK细胞的细胞结合。EC
表4:与表达huFAP的细胞的细胞结合
1.9如通过Biacore确定的抗人FAP克隆的竞争性结合
使用基于Biacore T200仪器的表面等离子体共振(SPR)试验进行表位分级。FAP抗原被固定化的抗His抗体捕获。在第一步中,注射FAP结合物直至饱和。随后注射第二FAP结合物。该试验设计示意性示出于图3C中。加入第二抗体后结合信号的增加表明其与第一抗体不同的表位结合。没有另外的结合表明第一抗体和第二抗体识别相同的表位区。
通过胺偶联(GE Healthcare试剂盒BR-1000-50)将浓度为20μg/ml的抗His抗体(GE Healthcare试剂盒28-9950-56)固定在CM5传感器芯片(GE Healthcare BR-1005-30)的表面上。注射时间为600秒,以10μl/分钟的流速在两个流通池上产生12000个应答单位(RU),一个用作参考,并且一个用作活性流通池。运行缓冲液为HBS-N(GE Healthcare BR-1006-70)。为了测量,使用PBS-P+(GE Healthcare 28-9950-84)作为运行缓冲液和稀释缓冲液。流通池温度设置为25℃,样品室设置为12℃。整个运行的流速设置为10μL/min。
在活性流通池中以20μg/ml的浓度捕获His标记的FAP抗原180秒。在两个流通池中以10μg/ml的浓度依次注射第一抗体和第二抗体(FAP结合物),各自持续120秒。每个循环后,用10mM甘氨酸(pH 1.5)持续60秒使表面再生(GE Healthcare BR-1003-54)。
结果示出于下表5中:
表5:抗FAP抗体与4B9的竞争性结合
因此,鉴定了三个表位仓。按照要求,没有一种抗FAP抗体与抗体4B9(表位仓1)竞争结合。抗体210、211、212和214彼此竞争结合并因此形成一个组(表位仓3),而抗体209与任何其他抗体(表位仓2)不竞争结合。
1.9抗FAP抗体的热稳定性评估
在20mM组氨酸/组氨酸氯化物、140mM NaCl、pH 6.0中,以1mg/mL的浓度制备样品,通过经由0.4μm滤板离心而转移到384孔光学板中,并用石蜡油覆盖。在DynaPro读板仪(Wyatt)上通过动态光散射重复测量流体动力学半径,同时将样品以0.05℃/min的速度从25℃加热到80℃。替代性地,将样品转移到10μL微量比色皿阵列中,并用Optim1000仪器(Avacta Inc.)记录了静态光散射数据和266nm激光激发后的荧光数据,同时将它们以0.1℃/min的速度从25℃加热到90℃。聚集起始温度(T
表6:抗FAP抗体的聚集起始温度
选择抗FAP克隆212用于人源化,因为它以与抗体4B9相当高的亲和力与人FAP结合,并显示出有利于开发的特性。经由电脑模拟,对其序列的分析表明只有一个预测的降解热点(401位处的Trp)。鼠克隆212的序列示出于表7中。
表7:鼠抗FAP克隆212的可变结构域的氨基酸序列
1.10抗FAP克隆212的人源化
1.10.1方法
通过查询人V区和J区序列的BLASTp数据库来鉴定鼠输入序列(裁剪至可变部分)的合适的人受体框架。选择人受体框架的选择标准是序列同源性、相同或相似的CDR长度和人种系的估计频率,而且还有VH-VL结构域界面上某些氨基酸的保守性。在种系鉴定步骤之后,将鼠输入序列的CDR移植到人受体框架区上。这些初始CDR移植物和亲本抗体之间的每个氨基酸差异被评估对相应可变区的结构完整性的可能影响,并且在认为适当时引入对亲本序列的“回复突变”。结构评估基于亲代抗体和人源化变体两者的Fv区同源性模型,该模型通过内部抗体结构同源性建模方案创建得到,其使用Biovia Discovery StudioEnvironment版本17R2实现。在一些人源化变体中,包括“正向突变”,即,将亲本结合物的给定CDR位置上发生的原始氨基酸改变为人受体种系的等效位置上发现的氨基酸的氨基酸交换。目的是增加人源化变体的总体人特征(在框架区之外),以进一步降低免疫原性风险。
内部开发的经由电脑模拟的工具用于预测配对的VH和VL人源化变体的VH-VL结构域方向(参见WO 2016/062734)。将结果与亲本结合物的预测VH-VL结构域取向进行比较,以选择在几何结构上与原始抗体接近的构架组合。基本原理是检测VH-VL界面区中可能的氨基酸交换,这些交换可能导致两个结构域的配对发生破坏性变化,进而对结合特性产生不利作用。
1.10.2受体框架的选择及其适应
选择了以下受体框架:
表8:受体框架
后CDR3框架区改编自人IGHJ种系IGHJ6*01/02(YYYYYGMDVWGQGTTVTVSS)和人IGKJ种系IGKJ4*01/02(LTFGGGTKVEIK)。与受体框架相关的部分以粗体显示。
基于结构上的考虑,在位置H43(Q>K)、H44(G>S)、H48(M>I)、H71(R>I)、H73(T>K)、H93(A>T)[VH1]、H49(S>G)、H71(R>I)、H73(N>K)、H78(L>A)、H93(A>T)、H94(K>R)[VH2]、L36(Y>F)、L43(A>P)、L87(Y>F)[VL1]和L36(Y>F)、L42(K>Q)、L43(A>P)、L85(T>M)、L87(Y>F)[VL2]处引入了从人受体框架到亲本结合物中氨基酸的回复突变。
此外,位置H60(N>A)、H64(K>Q)[VH1]、H60(N>A)、H61(Q>D)、H62(K>S)、H63(F>V)[VH2]、L33(I>L)、L34(N>A)[VL1]和L27b(V>I)、L33(I>L)[VL2]被认定为正向突变的有希望的候选者。所有位置均在Kabat EU编号方案中给出。
表9:变体列表
注:回复突变冠以前缀b,正向突变冠以前缀f,例如bM48I是指在48位(Kabat)上从蛋氨酸到异亮氨酸的回复突变(人种系氨基酸到亲本抗体氨基酸)。
所得基于受体框架的人源化FAP抗体的VH和VL结构域可参见下表10。
表10:人源化FAP抗体的VH和VL结构域的氨基酸序列
1.10.3新人源化抗FAP Fab
基于VH和VL的新人源化变体,表达了新的抗FAP Fab。
表11:表达为Fab的VH/VL组合的命名
与抗FAP抗体4B9相比,分析了基于克隆212的新人源化抗FAP变体的亲和力。此外,计算人源化变体的人化程度,并且测量其聚集起始温度。
表12:如通过Biacore测量的克隆212人源化变体的亲和力
1.11FcRn/肝素结合和经由电脑模拟的电荷分布
在经由电脑模拟模型中,计算抗体4B9和P1AE1689在PBS(pH 7.4)中的电荷分布。根据该模型,4B9具有大阳性斑块,这有时与肝素结合的增加有关。另一方面,P1AE1689显示出大负电荷斑块,这可能表明肝素的相互作用弱。
通过使用FcRn亲和柱和pH梯度以及肝素亲和柱和pH梯度对两种抗体进行色谱分析,证实了这些预测。WO 2015/140126公开了一种用于基于FcRn亲和色谱柱上确定的保留时间来预测抗体的体内半衰期的方法,而肝素结合与细胞表面结构的非特异性相互作用有关。
实例2
抗CD40抗体S2C6的人源化变体的产生和生产
2.1抗CD40抗体S2C6的人源化变体的产生
2.2.1方法
在对抗CD40结合物S2C6的人源化期间,为了鉴定合适的人受体框架,使用了两种方法的组合。一方面,通过寻找具有高序列同源性的受体框架,将CDR嫁接到该框架上,并评估可以设想哪些回复突变来采用经典方法。更明确地说,判断鉴定出的构架与亲代抗体的每个氨基酸差异对结合剂的结构完整性的影响,并且在适当时引入朝向亲代序列的回复突变。结构评估基于亲代抗体及其人源化版本的Fv区同源性模型,该模型通过内部抗体结构同源性建模工具创建得到,该工具使用Biovia Discovery Studio Environment 4.5版实现。
另一方面,使用内部开发的经由电脑模拟的工具来预测人源化形式的VH和VL结构域彼此之间的取向(参见WO 2016062734,通过引用并入本文)。将结果与预测的亲本结合物的VH-VL结构域取向进行比较,以选择在几何结构上与起始抗体接近的构架组合。基本原理是检测VH-VL界面区中可能的氨基酸交换,该交换可能导致两个结构域的配对发生破坏性变化。
2.2.2受体框架的选择及其适应
如下表16和表18中所述选择两个不同的受体框架。
表13:受体框架1:“IGHV1-IGKV2D”
后CDR3框架区改编自人IGHJ种系IGHJ6*01/02(YYYYYGMDVWGQGTTVTVSS)和人IGKJ种系IGKJ4*01/02(LTFGGGTKVEIK)。与受体框架相关的部分以粗体显示。
基于结构上的考虑,在VH区的位置H43(Q>K)、H44(G>S)、H69(M>L)、H71(R>V)、H73(T>K)、H88(V>A)和H105(Q>H)以及在VL区的位置L2(I>V)、L4(M>V)、L87(Y>F)和L104(V>L)处引入了从人受体框架到亲本结合物中氨基酸的回复突变。在一个变体中,突变T70S(VH)被纳入以研究在该位置稍微更多亲水性残基的作用。
所有变体均包括N54A突变(VH),以解决推定的可开发性热点(天冬酰胺脱酰胺基作用)。所有位置均在Kabat EU编号方案中给出。
在下表14中,显示了人源化变体VH-VL配对矩阵:
突变N54A适用于所有VH变体,并且没有明确提及。回复突变冠以前缀b,正向突变冠以前缀f,并且其他突变冠以前缀x。
表15:受体框架2:“IGHV3-IGKV1”
后CDR3框架区改编自人IGHJ种系IGHJ6*01/02(YYYYYGMDVWGQGTTVTVSS)和人IGKJ种系IGKJ4*01/02(LTFGGGTKVEIK)。与受体框架相关的部分以粗体显示。
基于结构上的考虑,在VH区的位置H44(G>S)、H49(S>G)、H71(R>V)、H78(L>A)、H94(K>R)和H105(Q>H)以及VL区的位置L42(K>Q)、L43(A>S)和L87(Y>F)处引入了从人受体框架到亲本结合物中氨基酸的回复突变。此外,CDR-H2中的四个位置被认定为正向突变的有希望的候选者,即,从亲代结合物到人受体种系的氨基酸交换,为了提高总体人特征,即H60(N>G)、H61(Q>D)、H62(K>S)和H63(F>V)。
所有变体均包括N54A突变(VH),以解决推定的可开发性热点(天冬酰胺脱酰胺基作用)。所有位置均在Kabat EU编号方案中给出。
在下表16中,显示了人源化变体VH-VL配对矩阵:
回复突变冠以前缀b,正向突变冠以前缀f。
2.2.3所得人源化CD40抗体的VH和VL结构域
所得基于受体框架1的人源化CD40抗体的VH和VL结构域可参见下表17,并且所得基于受体框架2的人源化CD40抗体的VH和VL结构域列于下表18。
表17:基于受体框架1的人源化CD40抗体的VH和VL结构域的氨基酸序列
表18:基于受体框架2的人源化CD40抗体的VH和VL结构域的氨基酸序列
2.2.4huIgG1_LALA_PG形式的新人源化CD40抗体
基于VH和VL的新人源化变体,根据WO 2012/130831 A1中所述的方法,将新CD40抗体表达为具有效应子沉默Fc(P329G;L234,L235A)的huIgG1抗体,以消除与Fc受体的结合。
表19:表达为huIgG1_LALA_PG抗体的VH/VL组合的命名
作为人IgG1_LALAPG抗体的人源化CD40抗体的示例性全长序列可参见表20。
表20:人源化CD40 IgG1_LALAPG抗体的氨基酸序列
2.2.5huIgG1_LALA_PG形式的新人源化CD40抗体的产生
通过与编码不同肽链的表达载体悬浮生长的HEK293-F细胞的瞬时转染来表达抗体。根据细胞供应商的说明,使用抗体载体的Maxiprep(Qiagen,Germany)制剂,基于F17的培养基(Invitrogen,USA),PEIpro(Polyscience Europe GmbH)和初始细胞密度为1-2百万活细胞/ml,在不含血清的FreeStyle 293表达培养基(Invitrogen)中,进行至HEK293-F细胞(Invitrogen,USA)中的转染。在摇瓶或搅拌的发酵罐中培养7天后,通过以14000g离心30分钟收集细胞培养上清液,并且通过0.22μm过滤器过滤。
使用MabSelectSure-Sepharose
不同人源化CD40抗体的产量作为滴定值示出于表21中,滴定值是由使用MabSelectSure-Sepharose
在存在和不存在还原剂的情况下,通过CE-SDS分析来分析最终纯化步骤之后的分子的纯度和分子量。根据制造商的说明使用Caliper LabChip GXII系统(CaliperLifescience)。
使用TSKgel G3000 SW XL分析尺寸排阻柱(Tosoh),在25mM磷酸钾、125mM氯化钠、200mM左旋精氨酸单氢氯化物、0.02%(w/v)NaN
为了直接比较所有抗体,通过静态光散射(SLS)并通过测量应答于施加的温度应力的内在蛋白荧光来监测热稳定性。将30μg蛋白浓度为1mg/ml的过滤蛋白样品一式两份加到Optim 2仪器(Avacta Analytical Ltd)上。温度以0.1℃/分钟从25℃渐升至85℃,并收集了半径和总散射强度。为了测定内在蛋白荧光,将样品在266nm处激发,并在275nm至460nm之间收集发射。对于所有抗体,聚集温度(Tagg)在64℃至69℃之间,并在下表21或表22中提供。
具有不同框架的人源化CD40抗体的产量示出于下表21或表22中。表21:基于受体框架2的人源化CD40抗体的生产滴度、人化程度和聚集温度
表22:基于受体框架1的人源化CD40抗体的生产滴度、人化程度和聚集温度
2.2.6重组人和食蟹猴CD40细胞外结构域蛋白的产生
将以下构建体克隆并在HEK293细胞中通过瞬时表达来表达:
1)具有C末端His-AviTag
2)具有C末端His-AviTag
通过基因合成(Eurofins Genomics GmbH service,Germany)生成用于结合分析的CD40细胞外结构域抗原,使用标准克隆程序经由独特的限制性位点将其克隆到Roche内部表达载体中。通过测序验证所有构建体的克隆。所有抗原均在CMV启动子的控制下表达。为了瞬时表达CD40细胞外结构域构建体,将适应悬浮的HEK293-F细胞(LifeTechnologies,USA)用各自的质粒转染:通常,根据制造商的说明,用通过PEIpro转染试剂(Polysciences Europe GmbH,Germany)复合的500μg质粒DNA转染1L约2x10
CD40细胞外结构域的生物素化:
根据制造商的说明,通过使用BirA生物素蛋白连接酶试剂盒(Avidity LLC,USA)对含有C末端AviTag
2.2.7人/食蟹猴CD40结合表面等离子体共振光谱测定
通过使用GE Healthcare提供的胺偶联试剂盒,在CM5芯片(GE Healthcare BR-1005-30)上在pH 5.0偶联了捕获系统(10μg/mL山羊抗人F(ab)'
2.2.8用于CD40特异性人源化抗体的表征的细胞结合测定
使用胰蛋白酶将CD40阳性细胞(Raji细胞)从培养瓶中分离出,并使用Casy细胞计数器进行计数。在4℃造粒后,将细胞重悬于FACS缓冲液(2.5%FCS在PBS中),调节至2.0E+06细胞/mL,并分配至96孔PP V型底板(25μL/孔=5.0E+04Zellen/孔)。
在FACS缓冲液中将CD40特异性抗体调整为20μg/mL,从而导致终浓度为10μg/mL。将20μL加入至25μL细胞悬液中,并在4℃孵育1h。然后将细胞在FACS缓冲液中洗涤两次。在洗涤后,将细胞重悬于50μL含次级抗体的FACS缓冲液(
在表23中,显示了人源化的CD40抗体的亲和力(通过Biacore测量的)和与表达CD40的细胞(Raji细胞)的细胞结合。
表23:人源化CD40抗体与表达CD40的细胞的亲和力和细胞结合
2.2.9通过UHR-ESI-QTOF质谱法的抗体表征
使用具有2%甲酸(v/v)的40%乙腈在Sephadex G25 5x250 mm柱(AmershamBiosciences,Freiburg,Germany)上通过HPLC将样品脱盐。在配备有TriVersa NanoMate源(Advion,Ithaca,NY)的maXis 4G UHR-QTOF MS系统(Bruker Daltonik,Bremen,Germany)上通过UHR-ESI-QTOF MS测定总质量。以900-4000m/z(ISCID:0.0eV)进行数据采集。使用内部开发的软件工具评估原始质谱并将其转换为单个相对摩尔质量。
2.2.10抗体的热稳定性评估
在20mM组氨酸/组氨酸氯化物、140mM NaCl、pH 6.0中,以1mg/mL的浓度制备样品,通过经由0.4μm滤板离心而转移到384孔光学板中,并用石蜡油覆盖。在DynaPro读板仪(Wyatt)上通过动态光散射重复测量流体动力学半径,同时将样品以0.05℃/min的速度从25℃加热到80℃。替代性地,将样品转移到10μL微量比色皿阵列中,并用Optim1000仪器(Avacta Inc.)记录了静态光散射数据和266nm激光激发后的荧光数据,同时将它们以0.1℃/min的速度从25℃加热到90℃。聚集起始温度定义为处于流体动力学半径(DLS)时的温度或散射光强度(Optim1000)开始增大时的温度。解链温度定义为显示荧光强度相对于波长的曲线图中的拐点。
实例3
生成和生产包含新型抗FAP克隆212及其人源化变体的双特异性构建体3.1靶向成纤维细胞活化蛋白(FAP)和CD40的双特异性抗原结合分子的产生
以如下形式制备双特异性CD40-FAP 2+1形式:所述抗体由与在Fc的C末端与一个FAP结合部分组合的两个CD40结合部分组成(图1A至1c),或4+1形式:所述抗体由与在Fc的C末端与一个FAP结合部分组合的四个CD40结合部分组成(图1D至1F)。双特异性CD40-FAP抗体包括新型抗FAP克隆212(图1A和1D),但是为了比较,制备了包含FAP克隆4B9(图1B和1E)和28H1(图1C和1F)的相应的分子。FAP结合物28H1和4B9的产生和制备已描述于WO 2012/020006 A2中,所述专利以引用方式并入本文。为了产生4+1和2+1分子杵臼结构技术已实现异源二聚化。将S354C/T366W突变引入第一重链HC1(Fc突起重链),并且将Y349C/T366S/L368A/Y407V突变引入第二重链HC2(Fc孔重链)。独立于双特异性形式,在所有情况下,根据WO 2012/130831 A1中所述的方法使用效应子沉默Fc(P329G;L234,234A)消除与Fc受体的结合。双特异性分子的序列示出于表24。
所有基因在嵌合MPSV启动子的控制下瞬时表达,所述启动子由与CMV启动子增强子片段组合的MPSV核心启动子组成。表达盒在cDNA的3'末端还含有合成的polyA信号。表达载体还含有用于在含有EBNA(爱泼斯坦巴尔病毒核抗原)的宿主细胞中游离复制的oriP区域。
表24:双特异性抗原结合分子的氨基酸序列
3.2靶向FAP和CD40的双特异性抗原结合分子的产生
靶向成纤维细胞活化蛋白(FAP)和CD40的双特异性抗原结合分子是通过瞬时转染HEK细胞表达,该HEK细胞是与编码4条不同肽链的表达载体悬浮培养的。根据细胞供应商的说明,使用抗体载体的Maxiprep(Qiagen)制备,F17培养基(Invitrogen,USA),Peipro(Polyscience Europe GmbH)和初始细胞密度为1-2百万活细胞/ml,在不含血清的FreeStyle 293表达培养基(Invitrogen)中,进行到HEK293-F细胞(Invitrogen)中的转染。在摇瓶或搅拌的发酵罐中培养7天后,通过以14000g离心30分钟收集细胞培养上清液,并且通过0.22μm过滤器过滤。
使用MabSelectSure-Sepharose
表25:双特异性CD40-FAP抗原结合分子的产量
P1AE2734是一种双特异性抗体,其中crossfab的VH-Cκ链融合至Fc结构域重链中的一个的C末端。该双特异性抗体比双特异性抗体P1AE2423更难以生产和纯化,其中crossfab的VL-CH1链融合至Fc结构域重链中的一个的C末端。必须应用进一步纯化步骤,即离子交换色谱法和另外的CE-SDS,得到1mg物质。
3.3双特异性抗huFAP抗体的结合动力学
为评估双特异性FAP靶向抗体的人FAP结合动力学,按照实例1.7所述的方法测量结合动力学,但是使用带有HIS标记的人FAP胞外域作为配体。下表25A显示了具有不同融合的crossfab的双特异性2+1 CD40 x FAP抗体的亲和力数据。
表25A:通过SPR测量2+1 CD40 x FAP双特异性抗体与人FAP的亲和力
双特异性抗体在C末端Fab融合时的FAP结合亲和力在两种交叉变体之间处于相似的范围内(通过SPR测得)。
实例4
靶向CD40和FAP的双特异性构建体的表征
4.1与表达人FAP的鼠成纤维细胞的结合
使用表达人成纤维细胞活化蛋白(huFAP)的NIH/3T3-hFAP克隆19来测试与细胞表面FAP的结合。NIH/3T3-huFAP克隆19是通过用表达载体pETR4921转染小鼠胚胎成纤维细胞NIH/3T3细胞系(ATCC CRL-1658)以在1.5μg/mL嘌呤霉素选择下表达huFAP而产生。
将NIH/3T3-huFAP细胞与补充10%胎牛血清(FBS)(life technologies,Cat.No.16140,Lot No.1797306A)的1x Dulbecco's Modified Eagle培养基(DMEM)(gibco,Cat.No.42430-025)一起培养。将1.5μg/mL的嘌呤霉素(gibco,Cat.No.A11138-03)加入至培养基中以选择表达FAP的细胞。通过使用无酶细胞解离缓冲液(gibco,Cat.No.13151014)从培养瓶中取出NIH/3T3-huFAP细胞。将0.3x10
如图4所示,FAP的单价双特异性抗体结合至表达人FAP的靶细胞。因此,靶向FAP的抗CD40抗原结合分子表现出直接的肿瘤靶向特性。包含C末端FAP(212(P1AE1689))或FAP(4B9)结合结构域的FAP构建体的结合强度高于包含C末端FAP(28H1)结合结构域的FAP构建体,这可以通过FAP(212)和FAP(4B9)结合物对人FAP的结合亲和力高于FAP(28H1)结合物来解释。对于具有FAP(4B9)结合部分(P1AE2487)的2+1构建体,观察到最强的FAP结合。没有检测到同种型对照抗体DP47(人种系对照)与NIH/3T3-hFAP细胞的结合。不同双特异性抗体测量的EC
表26:不同双特异性抗体形式的212(P1AE1689)、4B9和28H1的人FAP结合表征
4.2与表达人CD40的原代B细胞的结合
使用从外周血单核细胞(PBMC)分离的人原代B细胞来测试与细胞表面CD40的结合。为了分离PBMC,从Stiftung Zürcher Blutspendedienst SRK获得了血沉棕黄层。将50mL血沉棕黄层稀释在相同体积的PBS(gibco,Cat.No.10010023)中。在50mL聚丙烯离心管(TPP,Cat.No.91050)中提供15mL Lymphoprep
如图5所示,所有描绘的克隆均与CD40结合,但其与CD40阳性B细胞的结合强度(EC
表27:不同双特异性抗体形式的CD40抗体的人CD40结合表征
实例5
靶向FAP的抗人CD40结合分子的功能特性
5.1通过靶向FAP的抗人CD40结合分子进行B细胞的CD40介导的活化
CD40的连接诱导B细胞和树突细胞(DC)成熟以及活化,并促进这些细胞类型的存活。在CD40信号传导后,在B细胞和DC的表面上细胞因子产生和共刺激分子表达增加(S.Quezada等人,Annu Rev Immunol.2004,22,307-328;S.Danese等人,Gut.2004,53,1035–1043;G.Bishop等人,Adv Exp Med Biol.2007,597,131–151)。
为了测试不同的FAP依赖性抗CD40抗体的激动特性和FAP特异性,将从人血沉棕黄层获得的Daudi细胞或原代B细胞与FAP依赖性激动性抗人CD40抗体在存在FAP包被的珠粒的情况下一起孵育,并且通过FACS测量B细胞活化。
5.1.1.通过使用FAP包被的
将1x10
与激动性抗CD40抗体孵育2天后分析的Daudi细胞显示,所有描绘的抗体的CD70表达均增加(参见图6A和图6B)。在不同靶向FAP的抗体的情况下,该活化标志物的上调取决于FAP。在较低的抗体浓度下,与交联的CD40抗体(P1AD4470)所诱导的表达水平的增加相比,这些依赖于FAP的抗体所诱导的表达水平的增加幅度更高。另外,与具有FAP(28H1)结合结构域的2+1形式的双特异性FAP-CD40抗体、具有FAP(212)、FAP(4B9)或FAP(28H1)结合结构域的4+1形式的双特异性FAP-CD40抗体或与FAP无关的阳性对照抗体所诱导的上调相比,具有FAP(212)或FAP(4B9)结合结构域的2+1形式的双特异性FAP-CD40抗体所诱导的CD70上调幅度更高。在不存在FAP(未经包被的珠粒)的情况下,用描绘的CD40的双特异性抗体未观察到CD70的增加,而四价CD40结合分子诱导CD70的上调,但程度要比存在FAP的情况下更小,指示在Daudi细胞中四价CD40结合物的FAP依赖性CD40活化程度低但可检测。
5.1.2通过使用FAP包被的
如实例4.2中所述,将B细胞从血沉棕黄层中分离出来,并且将1×10
与激动性抗CD40抗体孵育2天后分析的B细胞显示,所有描绘的抗体的CD86表达均增加(参见图7A和图7B)。在靶向FAP的不同抗体的情况下,CD86的上调依赖于FAP,并且这些依赖于FAP的抗体所诱导的表达水平的增加与交联的抗CD40抗体P1AD4470所诱导的增加幅度相当或略低。包含一个FAP(212)结合结构域的单价结合FAP的双特异性抗体诱导的活化标志物表达水平的上调与包含一个FAP(4B9)结合部分的分子相似。在较低抗体浓度下,与包含FAP(212)或FAP(4B9)结合部分的2+1形式相比,4+1形式诱导了更高的B细胞活化,无论其FAP结合部分如何。
5.2通过靶向FAP的抗CD40结合分子,CD40介导的DC活化和随后的T细胞的引发
为了证明由FAP依赖性抗人CD40抗体活化的DC有效地引发T细胞的能力,建立了体外T细胞引发测定。对于这些测定,分离来自表达人CD40受体(huCD40tg小鼠;具有相似的人和鼠CD40受体表达模式的小鼠;C57BL/6背景;由Taconic产生)的转基因小鼠脾脏的DC,用SIINFEKL肽或卵清蛋白(OVA;DEC-205受体介导的抗原摄取)脉冲,并与不同的激动性抗人CD40抗体一起孵育。FAP是经由FAP包被的
5.2.1经由通过靶向FAP的抗CD40结合分子活化的OVA脉冲的DC的T细胞引发
将DC从huCD40tg小鼠的脾脏中分离出。为了分离脾脏DC,将来自huCD40tg小鼠的脾脏放入6孔板的一个孔中,该板含有含钙
然后将DC与1ng/mL SIINFEKL(Ovalbumin residues 257-264,Eurogentec,Cat.No.AS-60193-5,批号1360618)混合作为阳性对照,这不需要通过DC摄取和处理,或与OVA蛋白一起上样作为抗原。为了以Toll样受体(TLR)刺激独立方式促进OVA摄取(另外的TLR刺激可能会导致DC的高整体活化,使得不可能由于用激动性抗CD40抗体刺激来检测不同的活化状态),根据制造商的方案,将OVA抗原递送试剂(Miltenyi,Cat.No.130-094-663)与生物素化的抗小鼠DEC205抗体(Miltenyi,克隆NLDC-145,Cat.No.130-101-854)联合使用。简而言之,将DC与结合DEC205受体的生物素化抗体一起孵育,所述抗体在CD8阳性交叉呈递DC上高度表达(M.Lahoud等人,Int Immunol.2000,12(5),731-735)。之后,将OVA递送试剂(一种与FITC和OVA偶联的抗生物素抗体)加入至细胞中,从而导致DEC205受体介导的OVA摄取。为了提供阴性对照,仅用抗DEC205抗体标记DC,而不添加OVA。另外,如实例5.1.1中所述,将人FAP包被的或未经包被的
在第二天,将脾脏CD8阳性细胞从OT1小鼠中分离出。为了这样做,将OT1小鼠的脾脏粉碎通过具有端部3mL注射器塞的40μm滤器滤到50mL管中。将滤器用R10洗涤,并将脾细胞在室温以1500rpm离心5分钟。将1mL的1x细胞裂解缓冲液(用蒸馏水以1:10稀释)加入至细胞中,并在室温孵育四分钟后,加入20mL R10。将管在室温以1500rpm离心5分钟,并弃去上清液。将脾细胞重悬于30mL R10中,并用自动EVE细胞计数器确定细胞计数和生存力。根据制造商的说明,使用小鼠CD8a
图8A和图8B显示了用OVA递送试剂孵育并用靶向人CD40和FAP的双特异性抗原结合分子刺激的DC大幅增强了CD8阳性OT1 T细胞增殖。这些作用是依赖FAP的。与由交联的CD40抗体(P1AD4470)诱导的增加相比,由描绘的FAP依赖性抗体诱导的T细胞增殖的增加略微更低。通过具有一个FAP(212)或FAP(4B9)结合部分的2+1或4+1双特异性抗CD40抗体刺激的DC诱导的增殖水平是相当的。
5.3针对FAP结合和CD40信号转导的两项细胞系桥接试验
双特异性抗体的作用方式反映在两项细胞系桥接试验中:抗体与表达FAP的细胞系上的FAP的结合以及与报告基因细胞系上的CD40受体的结合。在CD40受体簇聚的情况下,信号转导诱导NFκB依赖性产生分泌型胚胎碱性磷酸酶(SEAP)。
简而言之:用Accutase(Life Technologies)分离报告细胞系HEK-Blue
将在表面上表达FAP的天然靶细胞(WM-266-4,ATCC)用作随时可用的冷冻细胞。每个小瓶包含在2mL DMEM(含10%FBS和5%DMSO)中的2×10
利用HEK-Blue
将抗体稀释液在HEK-Blue
将两种细胞系(报告细胞系和靶细胞系)在室温以180×g离心4分钟。除去上清液,将各种细胞系重悬于17mL HEK-Blue
表28:在两项细胞系桥接试验中测得的EC
包含两个结合CD40的Fab片段和结合FAP的crossfab片段的2+1分子P1AE2423与P1AE2302的EC
5.4与FAP无关的CD40特异性测定
评估了CD40途径在不通过FAP簇聚的情况下的潜在活化。在CD40受体活化的情况下,信号转导诱导NFκB依赖性产生分泌型胚胎碱性磷酸酶(SEAP)。
简而言之:用Accutase(Life Technologies)分离报告细胞系HEK-Blue
将报告细胞系在室温以180×g离心4分钟。除去上清液,将细胞重悬于17mL HEK-Blue
利用FAP依赖性测定中所用的最高浓度,将CD40特异性报告细胞与分子一起孵育。在不通过FAP结合簇聚的情况下,CD40受体的活化和随后的信号转导非常低。对于P1AF0873(1+1crossmab)和P1AA9663(2+2crossmab),CD40受体的活化和随后的信号转导甚至低于2+1分子P1AE2423和P1AE2302(误差条重叠)。
实例6
FAP靶向抗人CD40结合分子与PD-L1联用的抗肿瘤功效评估
设计了一项体内研究(图11),以便确定激动性抗FAP x CD40抗体在免疫活性小鼠中表达FAP的肿瘤细胞的作用。特别地,对包含与鼠FAP结合的28H1 FAP结合结构域的4+1形式的双特异性FAP-CD40抗体、包含28H1 FAP结合结构域的2+1形式的双特异性FAP-CD40抗体以及经过或未经aPD-L1处理的与FAP无关的CD40对照抗体之间的差异进行了评估。
根据规定指南(GV-Solas;Felasa;TierschG),将最初从Taconic获得的总共97+9(再激发实验所需)只huCD40 tg HO雌性小鼠(在实验开始时为6-9周龄,购自法国CharlesRivers)维持在不含特定病原体的条件下,日循环为12h光照/12h黑暗。实验研究方案经过地方政府的审查和批准(ZH225-17)。到达之后,将动物维持一周以适应新环境并进行观察。然后,在小鼠背部右侧皮下植入应答器以进行识别,并保持一周以上以使其恢复。定期进行连续健康状态监测。每天控制动物的临床症状并且检测不良反应。
MC38-muFAP invipa细胞(CRC)得自在Roche Glycart AG进行的体内传代,其扩增后沉积到Glycart内部细胞库中。将MC38-muFAP invipa细胞在包含10%FCS(PAALaboratories,Austria)、1mM丙酮酸盐、1x NEAA和6μg/mL嘌呤霉素的DMEM中培养。在5%CO
对于肿瘤细胞接种,将小鼠麻醉并且皮下注射到左胁腹,用29G针(BD)注射100μL含2×10
第22天,当肿瘤平均体积达到200mm
在随机分组和治疗注射后,每周通过卡尺测量肿瘤大小3次。每天进行监测以确保遵守动物福利要求。在许可标准(ZH225-17)中,如果肿瘤溃烂、肿瘤体积超过3000mm
表29:根据第22天的肿瘤体积将动物随机分组
表30:所用抗原结合分子
表31:治疗研究组
从排除的动物中,选择12只动物并且用A、C、D和E组中的疗法进行治疗(每种疗法3只动物),以便将其作为侦察动物(scouts)处死,分析第1次治疗注射后第4天的免疫药效学效应。
6.1靶向FAP的抗CD40结合分子诱导肿瘤完全缓解和有效的抗肿瘤免疫记忆反应
通过卡尺测量每周监测3次肿瘤大小。利用JMP12软件对肿瘤生长数据进行统计分析。为检验多次比较的组平均值的显著差异,使用Tukey-Kramer方法进行标准方差分析(ANOVA)。Tukey-Kramer对平均值之间的所有成对差异进行检验;如果样本量相同,则是精密的α水平检验。
在第1次治疗后第93天,在表现出完全肿瘤消退的小鼠的相对胁腹中皮下注射MC38-FAP肿瘤细胞,以便确定针对该肿瘤细胞系的免疫细胞记忆的形成。作为对照,根据兽医许可,9只天然C57bl/6-huCD40 tg小鼠接受相同的MC38-FAP肿瘤细胞注射,然后每周通过卡尺测量2-3次肿瘤生长。
如图12所示,在经过或不经PD-L1治疗的情况下,两种FAP-CD40双特异性分子(2+1和4+1)均诱导肿瘤完全缓解。与媒介物组相比,接受非靶向CD40、PD-L1或两者联合治疗的小鼠的肿瘤生长无延迟。另外,再激发后,之前接受FAP-CD40抗体治疗的组中无MC38-FAP肿瘤生长,而100%的天然小鼠中发生肿瘤生长。这些结果表明,与非靶向CD40治疗相比,靶向CD40治疗具有强效MC38-FAP抗肿瘤功效和肿瘤生长消退作用。由于接受抗FAP-CD40抗体治疗的所有小鼠均表现出100%的肿瘤缓解,因此在这些小鼠中未观察到明显的抗PD-L1附加效应。另外,在肿瘤生长抑制方面,2+1形式与4-1形式之间不存在明显差异。再激发数据表明,在接受FAP-CD40抗体治疗的所有小鼠中,均形成了针对MC38-FAP肿瘤细胞的有效的免疫记忆反应。
统计分析表明,与媒介物相比,2+1FAP-CD40抗体治疗组从第28天(治疗后第6天)开始出现显著差异(图13A,统计分析比较了所有组在第28天的数据),并且4+1FAP-CD40抗体治疗组从第31天(治疗后第9天)开始出现显著差异(图13B,统计分析比较了所有组在第31天的数据)。
6.2与2+1靶向FAP的抗CD40结合分子相比,4+1靶向FAP的抗CD40结合分子具有更高的血清清除率
采集每组3-4只小鼠的血样,以便跟踪腹腔注射后药物的药代动力学特征。在第一种抗体治疗后6h、24h、48h、72h、96h、8天和15天采集血清样品,以分析血液中的抗CD40水平。每个时间点最多采集100μL血液。通过加热至39℃的盒子使小鼠体温略微升高,以确保良好的血管扩张。用22G针对尾静脉进行穿刺。将血液收集到1.1mL Z-Gel微量管(SARSTEDT)中,并通过离心后收集上清液来提取血清。样品储存于-20℃,然后使用相应的校准抗体分析抗CD40水平。
根据方案(Roche-Glycart,Switzerland),使用CD40/Fc嵌合体(R&D,1493-CD-050)ELISA检测血清中的抗CD40浓度,以定量分析抗CD40huIgG。使用以下试剂捕获蛋白:人IgG-Fc生物素(Abcam,ab98561)和CD40/Fc嵌合体(R&D,1493-CD-050),检测抗体:抗小鼠IgG(HRP)(Abcam,ab97040)或抗huCH1-DIG(Roche-Penzberg,Germany)和抗DIG-POD(Roche,11633716001)。根据注射的剂量(mg/kg)和抗CD40注射后的采集时间(6h,24h,48h,96h),对连续稀释系列中的血清样品进行分析。使用405nm的测量波长和490nm的参比波长(SpectraMax i3酶标仪,Molecular Devices)测量吸光度。
对于PK分析,使用Phoenix 64、WinNonlin 6.4软件计算曲线下面积以及Cmax。由于仅监测最长96小时的药代动力学,因此未显示PK参数诸如清除率和半衰期。
如图14和表32所示,观察到4+1形式的清除率高于2+1形式。这一较高的清除率可以通过四价CD40结合物(4+1形式)的CD40特异性靶标介导的药物处置(TMDD)比二价CD40结合物(2+1形式)更突出来解释。
另外,与两种FAP-CD40分子相比,非靶向CD40表现出更高的清除率。这一清除率无法通过TMDD假设来解释,而可能是由于CD40治疗后形成了抗药物抗体(ADA)。
表32:PK参数
6.3治疗后96小时,靶向FAP的抗CD40结合分子引起CD40介导的DC、B细胞和T细胞活化
在第1次治疗后96h,将每组3只小鼠处死,并采集以下器官在PBS中用于流式细胞术分析:肿瘤、脾脏、腹股沟引流淋巴结以及腹股沟非引流淋巴结、轴非引流淋巴结和肱非引流淋巴结。
在流式细胞术分析中,按照实例5.2.1所述制备所有收集的器官的单细胞悬液,并且用荧光标记的抗体染色。为此,将制备的单细胞悬液转移至96孔平底板中,用PBS进行洗涤,然后与50μL的3μg/mL Fc受体阻断小鼠IgG同种型对照(ThermoFisher Scientific,Cat.No.10400C)在PBS中一起孵育。在4℃孵育15分钟后,将细胞用PBS洗涤,并且将50μL的在PBS中的荧光标记抗体混合物加入至细胞中。使用以下抗体:抗小鼠CD3 Pacific Blue
如图15所示,两种靶向FAP的抗CD40抗体形式(2+1和4+1)均在治疗后四天诱导肿瘤引流淋巴结、非引流淋巴结和脾脏中的DC活化(CD86表达)显著增加,但是增加幅度低于非靶向抗CD40抗体。相比之下,与媒介物组相比,仅2+1形式的靶向FAP的抗CD40抗体诱导肿瘤中发生显著的DC活化。如图16所示,观察到相同的T细胞活化模式(Ki-67表达)。与媒介物组相比,仅在接受非靶向抗CD40抗体治疗的小鼠中观察到所有分析的组织样品中发生显著的B细胞活化(CD69表达)(图17)。
6.4小鼠中与非靶向CD40抗体相比,靶向FAP的抗CD40结合分子诱导的副作用更少
6.4.1与接受非靶向抗CD40抗体治疗的小鼠相比,接受靶向FAP的抗CD40抗体治疗的小鼠未表现出体重减轻
在注射治疗后,每天测量每只小鼠的体重,并且持续15天。将体重减轻达到其初始体重(第0天)的20%以上的小鼠处死(此处无需处死小鼠)。
如图18所示,接受非靶向抗CD40抗体治疗的小鼠在注射抗体后体重明显减轻,而在联合注射抗CD40和抗PD-L1抗体的情况下,体重减轻的持续时间更长。在4+1或2+1 FAP-CD40单药、PD-L1单药注射组中以及FAP-CD40(4+1或2+1)+PD-L1联合治疗组中,均未观察到体重减轻。该结果显示了靶向FAP抗CD40结合分子相比于非靶向抗CD40抗体的安全性。
6.4.1与接受非靶向抗CD40抗体治疗的小鼠相比,接受靶向FAP的抗CD40抗体表现出较低的抗PD-L1介导的不良事件
在第3次注射抗PD-L1抗体后观察到一些不良事件。无需处死小鼠,但是将症状描述为在注射治疗后5分钟内,小鼠的活动能力下降、弓背和皮毛脏乱。表33列出了第3次治疗后表现出这些症状的小鼠的百分比。
表33:在第3次抗PD-L1治疗后表现出不良事件的动物百分比。
结果表明,与抗PD-L1单药治疗组相比,PD-L1+CD40联合治疗组的不良事件明显增加,而抗PD-L1+FAP-CD40组(4+1和2+1)中观察到的不良事件百分比仅略有增加。
观察到的不良事件可能是由于针对抗PD-L1的ADA形成了强烈的免疫反应(细胞因子风暴)的结果,这种情况在存在非靶向抗CD40抗体的情况下进一步加剧。该观察结果表明,当两种分子与抗PD-L1抗体疗法联合使用时,与非靶向抗CD40抗体相比,靶向FAP-CD40分子具有更出色的安全性。
总之,包含FAP依赖性活化CD40的4+1和2+1形式的FAP靶向抗CD40分子诱导荷瘤小鼠产生有效的抗肿瘤免疫应答,其全身毒性低于非靶向抗CD40亲本抗体。
***
序列表
<110> 豪夫迈·罗氏有限公司(F. Hoffmann-La Roche AG)
<120> 包含抗 FAP 克隆 212 的双特异性抗原结合分子
<130> P35043-WO
<150> EP18197866.9
<151> 2018-10-01
<160> 110
<170> PatentIn 3.5 版
<210> 1
<211> 277
<212> PRT
<213> 智人
<400> 1
Met Val Arg Leu Pro Leu Gln Cys Val Leu Trp Gly Cys Leu Leu Thr
1 5 10 15
Ala Val His Pro Glu Pro Pro Thr Ala Cys Arg Glu Lys Gln Tyr Leu
20 25 30
Ile Asn Ser Gln Cys Cys Ser Leu Cys Gln Pro Gly Gln Lys Leu Val
35 40 45
Ser Asp Cys Thr Glu Phe Thr Glu Thr Glu Cys Leu Pro Cys Gly Glu
50 55 60
Ser Glu Phe Leu Asp Thr Trp Asn Arg Glu Thr His Cys His Gln His
65 70 75 80
Lys Tyr Cys Asp Pro Asn Leu Gly Leu Arg Val Gln Gln Lys Gly Thr
85 90 95
Ser Glu Thr Asp Thr Ile Cys Thr Cys Glu Glu Gly Trp His Cys Thr
100 105 110
Ser Glu Ala Cys Glu Ser Cys Val Leu His Arg Ser Cys Ser Pro Gly
115 120 125
Phe Gly Val Lys Gln Ile Ala Thr Gly Val Ser Asp Thr Ile Cys Glu
130 135 140
Pro Cys Pro Val Gly Phe Phe Ser Asn Val Ser Ser Ala Phe Glu Lys
145 150 155 160
Cys His Pro Trp Thr Ser Cys Glu Thr Lys Asp Leu Val Val Gln Gln
165 170 175
Ala Gly Thr Asn Lys Thr Asp Val Val Cys Gly Pro Gln Asp Arg Leu
180 185 190
Arg Ala Leu Val Val Ile Pro Ile Ile Phe Gly Ile Leu Phe Ala Ile
195 200 205
Leu Leu Val Leu Val Phe Ile Lys Lys Val Ala Lys Lys Pro Thr Asn
210 215 220
Lys Ala Pro His Pro Lys Gln Glu Pro Gln Glu Ile Asn Phe Pro Asp
225 230 235 240
Asp Leu Pro Gly Ser Asn Thr Ala Ala Pro Val Gln Glu Thr Leu His
245 250 255
Gly Cys Gln Pro Val Thr Gln Glu Asp Gly Lys Glu Ser Arg Ile Ser
260 265 270
Val Gln Glu Arg Gln
275
<210> 2
<211> 760
<212> PRT
<213> 智人
<400> 2
Met Lys Thr Trp Val Lys Ile Val Phe Gly Val Ala Thr Ser Ala Val
1 5 10 15
Leu Ala Leu Leu Val Met Cys Ile Val Leu Arg Pro Ser Arg Val His
20 25 30
Asn Ser Glu Glu Asn Thr Met Arg Ala Leu Thr Leu Lys Asp Ile Leu
35 40 45
Asn Gly Thr Phe Ser Tyr Lys Thr Phe Phe Pro Asn Trp Ile Ser Gly
50 55 60
Gln Glu Tyr Leu His Gln Ser Ala Asp Asn Asn Ile Val Leu Tyr Asn
65 70 75 80
Ile Glu Thr Gly Gln Ser Tyr Thr Ile Leu Ser Asn Arg Thr Met Lys
85 90 95
Ser Val Asn Ala Ser Asn Tyr Gly Leu Ser Pro Asp Arg Gln Phe Val
100 105 110
Tyr Leu Glu Ser Asp Tyr Ser Lys Leu Trp Arg Tyr Ser Tyr Thr Ala
115 120 125
Thr Tyr Tyr Ile Tyr Asp Leu Ser Asn Gly Glu Phe Val Arg Gly Asn
130 135 140
Glu Leu Pro Arg Pro Ile Gln Tyr Leu Cys Trp Ser Pro Val Gly Ser
145 150 155 160
Lys Leu Ala Tyr Val Tyr Gln Asn Asn Ile Tyr Leu Lys Gln Arg Pro
165 170 175
Gly Asp Pro Pro Phe Gln Ile Thr Phe Asn Gly Arg Glu Asn Lys Ile
180 185 190
Phe Asn Gly Ile Pro Asp Trp Val Tyr Glu Glu Glu Met Leu Ala Thr
195 200 205
Lys Tyr Ala Leu Trp Trp Ser Pro Asn Gly Lys Phe Leu Ala Tyr Ala
210 215 220
Glu Phe Asn Asp Thr Asp Ile Pro Val Ile Ala Tyr Ser Tyr Tyr Gly
225 230 235 240
Asp Glu Gln Tyr Pro Arg Thr Ile Asn Ile Pro Tyr Pro Lys Ala Gly
245 250 255
Ala Lys Asn Pro Val Val Arg Ile Phe Ile Ile Asp Thr Thr Tyr Pro
260 265 270
Ala Tyr Val Gly Pro Gln Glu Val Pro Val Pro Ala Met Ile Ala Ser
275 280 285
Ser Asp Tyr Tyr Phe Ser Trp Leu Thr Trp Val Thr Asp Glu Arg Val
290 295 300
Cys Leu Gln Trp Leu Lys Arg Val Gln Asn Val Ser Val Leu Ser Ile
305 310 315 320
Cys Asp Phe Arg Glu Asp Trp Gln Thr Trp Asp Cys Pro Lys Thr Gln
325 330 335
Glu His Ile Glu Glu Ser Arg Thr Gly Trp Ala Gly Gly Phe Phe Val
340 345 350
Ser Thr Pro Val Phe Ser Tyr Asp Ala Ile Ser Tyr Tyr Lys Ile Phe
355 360 365
Ser Asp Lys Asp Gly Tyr Lys His Ile His Tyr Ile Lys Asp Thr Val
370 375 380
Glu Asn Ala Ile Gln Ile Thr Ser Gly Lys Trp Glu Ala Ile Asn Ile
385 390 395 400
Phe Arg Val Thr Gln Asp Ser Leu Phe Tyr Ser Ser Asn Glu Phe Glu
405 410 415
Glu Tyr Pro Gly Arg Arg Asn Ile Tyr Arg Ile Ser Ile Gly Ser Tyr
420 425 430
Pro Pro Ser Lys Lys Cys Val Thr Cys His Leu Arg Lys Glu Arg Cys
435 440 445
Gln Tyr Tyr Thr Ala Ser Phe Ser Asp Tyr Ala Lys Tyr Tyr Ala Leu
450 455 460
Val Cys Tyr Gly Pro Gly Ile Pro Ile Ser Thr Leu His Asp Gly Arg
465 470 475 480
Thr Asp Gln Glu Ile Lys Ile Leu Glu Glu Asn Lys Glu Leu Glu Asn
485 490 495
Ala Leu Lys Asn Ile Gln Leu Pro Lys Glu Glu Ile Lys Lys Leu Glu
500 505 510
Val Asp Glu Ile Thr Leu Trp Tyr Lys Met Ile Leu Pro Pro Gln Phe
515 520 525
Asp Arg Ser Lys Lys Tyr Pro Leu Leu Ile Gln Val Tyr Gly Gly Pro
530 535 540
Cys Ser Gln Ser Val Arg Ser Val Phe Ala Val Asn Trp Ile Ser Tyr
545 550 555 560
Leu Ala Ser Lys Glu Gly Met Val Ile Ala Leu Val Asp Gly Arg Gly
565 570 575
Thr Ala Phe Gln Gly Asp Lys Leu Leu Tyr Ala Val Tyr Arg Lys Leu
580 585 590
Gly Val Tyr Glu Val Glu Asp Gln Ile Thr Ala Val Arg Lys Phe Ile
595 600 605
Glu Met Gly Phe Ile Asp Glu Lys Arg Ile Ala Ile Trp Gly Trp Ser
610 615 620
Tyr Gly Gly Tyr Val Ser Ser Leu Ala Leu Ala Ser Gly Thr Gly Leu
625 630 635 640
Phe Lys Cys Gly Ile Ala Val Ala Pro Val Ser Ser Trp Glu Tyr Tyr
645 650 655
Ala Ser Val Tyr Thr Glu Arg Phe Met Gly Leu Pro Thr Lys Asp Asp
660 665 670
Asn Leu Glu His Tyr Lys Asn Ser Thr Val Met Ala Arg Ala Glu Tyr
675 680 685
Phe Arg Asn Val Asp Tyr Leu Leu Ile His Gly Thr Ala Asp Asp Asn
690 695 700
Val His Phe Gln Asn Ser Ala Gln Ile Ala Lys Ala Leu Val Asn Ala
705 710 715 720
Gln Val Asp Phe Gln Ala Met Trp Tyr Ser Asp Gln Asn His Gly Leu
725 730 735
Ser Gly Leu Ser Thr Asn His Leu Tyr Thr His Met Thr His Phe Leu
740 745 750
Lys Gln Cys Phe Ser Leu Ser Asp
755 760
<210> 3
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> FAP (212) CDR-H1
<400> 3
Asp Tyr Asn Met Asp
1 5
<210> 4
<211> 17
<212> PRT
<213> 人工序列
<220>
<223> FAP (212) CDR-H2
<400> 4
Asp Ile Tyr Pro Asn Thr Gly Gly Thr Ile Tyr Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 5
<211> 10
<212> PRT
<213> 人工序列
<220>
<223> FAP (212) CDR-H3
<400> 5
Phe Arg Gly Ile His Tyr Ala Met Asp Tyr
1 5 10
<210> 6
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> FAP (212) CDR-L1
<400> 6
Arg Ala Ser Glu Ser Val Asp Asn Tyr Gly Leu Ser Phe Ile Asn
1 5 10 15
<210> 7
<211> 7
<212> PRT
<213> 人工序列
<220>
<223> FAP (212) CDR-L2
<400> 7
Gly Thr Ser Asn Arg Gly Ser
1 5
<210> 8
<211> 9
<212> PRT
<213> 人工序列
<220>
<223> FAP (212) CDR-L3
<400> 8
Gln Gln Ser Asn Glu Val Pro Tyr Thr
1 5
<210> 9
<211> 119
<212> PRT
<213> 人工序列
<220>
<223> FAP (212) VH
<400> 9
Glu Val Leu Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ala Cys Lys Ala Ser Gly Tyr Thr Leu Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Arg Gln Ser His Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Asp Ile Tyr Pro Asn Thr Gly Gly Thr Ile Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ile Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Asp Leu Arg Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Arg Gly Ile His Tyr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Ser Val Thr Val Ser Ser
115
<210> 10
<211> 111
<212> PRT
<213> 人工序列
<220>
<223> FAP (212) VL
<400> 10
Asp Ile Val Leu Thr Gln Ser Pro Val Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Leu Ser Phe Ile Asn Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Gly Thr Ser Asn Arg Gly Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Ser Leu Asn Ile His
65 70 75 80
Pro Met Glu Glu Asp Asp Thr Ala Met Tyr Phe Cys Gln Gln Ser Asn
85 90 95
Glu Val Pro Tyr Thr Phe Gly Gly Gly Thr Asn Leu Glu Ile Lys
100 105 110
<210> 11
<211> 17
<212> PRT
<213> 人工序列
<220>
<223> FAP (VH1G3a) CDR-H2
<400> 11
Asp Ile Tyr Pro Asn Thr Gly Gly Thr Ile Tyr Ala Gln Lys Phe Gln
1 5 10 15
Gly
<210> 12
<211> 17
<212> PRT
<213> 人工序列
<220>
<223> FAP (VH2G3a) CDR-H2
<400> 12
Asp Ile Tyr Pro Asn Thr Gly Gly Thr Ile Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 13
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> FAP (VL1G3a) CDR-L1
<400> 13
Arg Ala Ser Glu Ser Val Asp Asn Tyr Gly Leu Ser Phe Leu Ala
1 5 10 15
<210> 14
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> FAP (VL2G3a) CDR-L1
<400> 14
Arg Ala Ser Glu Ser Ile Asp Asn Tyr Gly Leu Ser Phe Leu Asn
1 5 10 15
<210> 15
<211> 119
<212> PRT
<213> 人工序列
<220>
<223> FAP (VH1G1a)
<400> 15
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Leu Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Asp Ile Tyr Pro Asn Thr Gly Gly Thr Ile Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Met Thr Ile Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Arg Gly Ile His Tyr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
<210> 16
<211> 119
<212> PRT
<213> 人工序列
<220>
<223> FAP (VH1G2a)
<400> 16
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Leu Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Asp Ile Tyr Pro Asn Thr Gly Gly Thr Ile Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Met Thr Ile Asp Lys Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Arg Gly Ile His Tyr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
<210> 17
<211> 119
<212> PRT
<213> 人工序列
<220>
<223> FAP (VH1G3a)
<400> 17
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Leu Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Asp Ile Tyr Pro Asn Thr Gly Gly Thr Ile Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Met Thr Ile Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Arg Gly Ile His Tyr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
<210> 18
<211> 119
<212> PRT
<213> 人工序列
<220>
<223> FAP (VH2G1a)
<400> 18
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Leu Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Asp Ile Tyr Pro Asn Thr Gly Gly Thr Ile Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Arg Gly Ile His Tyr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
<210> 19
<211> 119
<212> PRT
<213> 人工序列
<220>
<223> FAP (VH2G2a)
<400> 19
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Leu Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Asp Ile Tyr Pro Asn Thr Gly Gly Thr Ile Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ile Asp Lys Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Arg Gly Ile His Tyr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
<210> 20
<211> 119
<212> PRT
<213> 人工序列
<220>
<223> FAP (VH2G3a)
<400> 20
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Leu Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Asp Ile Tyr Pro Asn Thr Gly Gly Thr Ile Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Arg Gly Ile His Tyr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser
115
<210> 21
<211> 111
<212> PRT
<213> 人工序列
<220>
<223> FAP (VL1G1a)
<400> 21
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Leu Ser Phe Ile Asn Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro
35 40 45
Arg Leu Leu Ile Tyr Gly Thr Ser Asn Arg Gly Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Phe Cys Gln Gln Ser Asn
85 90 95
Glu Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 22
<211> 111
<212> PRT
<213> 人工序列
<220>
<223> FAP (VL1G2a)
<400> 22
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Leu Ser Phe Ile Asn Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Arg Leu Leu Ile Tyr Gly Thr Ser Asn Arg Gly Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Phe Cys Gln Gln Ser Asn
85 90 95
Glu Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 23
<211> 111
<212> PRT
<213> 人工序列
<220>
<223> FAP (VL1G3a)
<400> 23
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Leu Ser Phe Leu Ala Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro
35 40 45
Arg Leu Leu Ile Tyr Gly Thr Ser Asn Arg Gly Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Phe Cys Gln Gln Ser Asn
85 90 95
Glu Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 24
<211> 111
<212> PRT
<213> 人工序列
<220>
<223> FAP (VL2G1a)
<400> 24
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Leu Ser Phe Ile Asn Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Gly Thr Ser Asn Arg Gly Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ser Asn
85 90 95
Glu Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 25
<211> 111
<212> PRT
<213> 人工序列
<220>
<223> FAP (VL2G2a)
<400> 25
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Leu Ser Phe Ile Asn Trp Phe Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Gly Thr Ser Asn Arg Gly Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Met Tyr Phe Cys Gln Gln Ser Asn
85 90 95
Glu Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 26
<211> 111
<212> PRT
<213> 人工序列
<220>
<223> FAP (VL2G3a)
<400> 26
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Ile Asp Asn Tyr
20 25 30
Gly Leu Ser Phe Leu Asn Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Gly Thr Ser Asn Arg Gly Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ser Asn
85 90 95
Glu Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 27
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> hu CD40 CDR-H1
<400> 27
Gly Tyr Tyr Ile His
1 5
<210> 28
<211> 17
<212> PRT
<213> 人工序列
<220>
<223> hu CD40 CDR-H2
<400> 28
Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe Lys
1 5 10 15
Gly
<210> 29
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> hu CD40 CDR-H3
<400> 29
Glu Gly Ile Tyr Trp
1 5
<210> 30
<211> 16
<212> PRT
<213> 人工序列
<220>
<223> hu CD40 CDR-L1
<400> 30
Arg Ser Ser Gln Ser Leu Val His Ser Asn Gly Asn Thr Phe Leu His
1 5 10 15
<210> 31
<211> 7
<212> PRT
<213> 人工序列
<220>
<223> hu CD40 CDR-L2
<400> 31
Thr Val Ser Asn Arg Phe Ser
1 5
<210> 32
<211> 9
<212> PRT
<213> 人工序列
<220>
<223> hu CD40 CDR-L3
<400> 32
Ser Gln Thr Thr His Val Pro Trp Thr
1 5
<210> 33
<211> 114
<212> PRT
<213> 人工序列
<220>
<223> hu CD40 VH
<400> 33
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Leu Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Leu Val Thr Val
100 105 110
Ser Ser
<210> 34
<211> 112
<212> PRT
<213> 人工序列
<220>
<223> hu CD40 VL
<400> 34
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
35 40 45
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 35
<211> 114
<212> PRT
<213> 人工序列
<220>
<223> CD40 (S2C6) VH
<400> 35
Glu Val Gln Leu Gln Gln Ser Gly Pro Asp Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Lys Gln Ser His Gly Lys Ser Leu Glu Trp Ile
35 40 45
Gly Arg Val Ile Pro Asn Asn Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Ile Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Arg Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly His Gly Thr Thr Leu Thr Val
100 105 110
Ser Ser
<210> 36
<211> 112
<212> PRT
<213> 人工序列
<220>
<223> CD40 (S2C6) VL
<400> 36
Asp Val Val Val Thr Gln Thr Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Ala Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Val Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Gln
100 105 110
<210> 37
<211> 114
<212> PRT
<213> 人工序列
<220>
<223> VH1a (CD40)
<400> 37
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 38
<211> 114
<212> PRT
<213> 人工序列
<220>
<223> VH1b (CD40)
<400> 38
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 39
<211> 114
<212> PRT
<213> 人工序列
<220>
<223> VH1c (CD40)
<400> 39
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly His Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 40
<211> 114
<212> PRT
<213> 人工序列
<220>
<223> VH1d (CD40)
<400> 40
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Ser Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 41
<211> 112
<212> PRT
<213> 人工序列
<220>
<223> VL1a (CD40)
<400> 41
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 42
<211> 112
<212> PRT
<213> 人工序列
<220>
<223> VL1b (CD40)
<400> 42
Asp Ile Val Val Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 43
<211> 112
<212> PRT
<213> 人工序列
<220>
<223> VL1c (CD40)
<400> 43
Asp Val Val Val Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 44
<211> 112
<212> PRT
<213> 人工序列
<220>
<223> VL1d (CD40)
<400> 44
Asp Val Val Val Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 45
<211> 114
<212> PRT
<213> 人工序列
<220>
<223> VH2a (CD40)
<400> 45
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 46
<211> 114
<212> PRT
<213> 人工序列
<220>
<223> VH2b (CD40)
<400> 46
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Ser Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 47
<211> 114
<212> PRT
<213> 人工序列
<220>
<223> VH2c (CD40)
<400> 47
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly His Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 48
<211> 114
<212> PRT
<213> 人工序列
<220>
<223> VH2d (CD40)
<400> 48
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Gly Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 49
<211> 114
<212> PRT
<213> 人工序列
<220>
<223> VH2ab (CD40)
<400> 49
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 50
<211> 114
<212> PRT
<213> 人工序列
<220>
<223> VH2ac (CD40)
<400> 50
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser
<210> 51
<211> 112
<212> PRT
<213> 人工序列
<220>
<223> VL2a (CD40)
<400> 51
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
35 40 45
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 52
<211> 112
<212> PRT
<213> 人工序列
<220>
<223> VL2b (CD40)
<400> 52
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 53
<211> 112
<212> PRT
<213> 人工序列
<220>
<223> VL2ab (CD40)
<400> 53
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
35 40 45
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 54
<211> 112
<212> PRT
<213> 人工序列
<220>
<223> VL2ac (CD40)
<400> 54
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Ile Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
35 40 45
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 55
<211> 443
<212> PRT
<213> 人工序列
<220>
<223> P1AE0400 重链
<400> 55
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440
<210> 56
<211> 219
<212> PRT
<213> 人工序列
<220>
<223> P1AE0400 轻链
<400> 56
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
35 40 45
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 57
<211> 443
<212> PRT
<213> 人工序列
<220>
<223> P1AE0403 重链
<400> 57
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Gly Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440
<210> 58
<211> 219
<212> PRT
<213> 人工序列
<220>
<223> P1AE0403 轻链
<400> 58
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
35 40 45
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 59
<211> 443
<212> PRT
<213> 人工序列
<220>
<223> P1AE0817 重链
<400> 59
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440
<210> 60
<211> 219
<212> PRT
<213> 人工序列
<220>
<223> P1AE0817 轻链
<400> 60
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 61
<211> 226
<212> PRT
<213> 人工序列
<220>
<223> (P1AE1689) 轻链交叉 VH-Cκ
<400> 61
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Leu Thr Asp Tyr
20 25 30
Asn Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Asp Ile Tyr Pro Asn Thr Gly Gly Thr Ile Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Met Thr Ile Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Phe Arg Gly Ile His Tyr Ala Met Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Val Thr Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe
115 120 125
Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val
130 135 140
Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp
145 150 155 160
Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr
165 170 175
Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr
180 185 190
Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val
195 200 205
Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly
210 215 220
Glu Cys
225
<210> 62
<211> 219
<212> PRT
<213> 人工序列
<220>
<223> VL1a (CD40) 轻链(带电荷的)
<400> 62
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Arg
115 120 125
Lys Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 63
<211> 679
<212> PRT
<213> 人工序列
<220>
<223> VH1a (CD40)(VHCH1 带电荷的)Fc 突起_PGLALA_(P1AE1689) (VL-CH1)
<400> 63
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Glu
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Glu Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly Gly Gly Gly Ser
435 440 445
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu
450 455 460
Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly Glu
465 470 475 480
Arg Ala Thr Leu Ser Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr Gly
485 490 495
Leu Ser Phe Ile Asn Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro Arg
500 505 510
Leu Leu Ile Tyr Gly Thr Ser Asn Arg Gly Ser Gly Ile Pro Ala Arg
515 520 525
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
530 535 540
Leu Glu Pro Glu Asp Phe Ala Val Tyr Phe Cys Gln Gln Ser Asn Glu
545 550 555 560
Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Ser Ser
565 570 575
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
580 585 590
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
595 600 605
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
610 615 620
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
625 630 635 640
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
645 650 655
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
660 665 670
Lys Val Glu Pro Lys Ser Cys
675
<210> 64
<211> 443
<212> PRT
<213> 人工序列
<220>
<223> VH1a (CD40)(VHCH1 带电荷的)Fc 孔_PGLALA
<400> 64
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Glu
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Glu Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440
<210> 65
<211> 216
<212> PRT
<213> 人工序列
<220>
<223> (P1AE1689) 轻链交叉 VL-CH1
<400> 65
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Glu Ser Val Asp Asn Tyr
20 25 30
Gly Leu Ser Phe Ile Asn Trp Phe Gln Gln Lys Pro Gly Gln Ala Pro
35 40 45
Arg Leu Leu Ile Tyr Gly Thr Ser Asn Arg Gly Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Phe Cys Gln Gln Ser Asn
85 90 95
Glu Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Ser
100 105 110
Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser
115 120 125
Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp
130 135 140
Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr
145 150 155 160
Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr
165 170 175
Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln
180 185 190
Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp
195 200 205
Lys Lys Val Glu Pro Lys Ser Cys
210 215
<210> 66
<211> 219
<212> PRT
<213> 人工序列
<220>
<223> VL1a (CD40) 轻链
<400> 66
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 67
<211> 689
<212> PRT
<213> 人工序列
<220>
<223> VH1a (CD40) (VHCH1) Fc 突起_PGLALA_(P1AE1689) (VH-Cκ)
<400> 67
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly Gly Gly Gly Ser
435 440 445
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln
450 455 460
Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala Ser
465 470 475 480
Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Leu Thr Asp Tyr Asn
485 490 495
Met Asp Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile Gly
500 505 510
Asp Ile Tyr Pro Asn Thr Gly Gly Thr Ile Tyr Asn Gln Lys Phe Lys
515 520 525
Gly Arg Val Thr Met Thr Ile Asp Thr Ser Thr Ser Thr Val Tyr Met
530 535 540
Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys Thr
545 550 555 560
Arg Phe Arg Gly Ile His Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr
565 570 575
Thr Val Thr Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile
580 585 590
Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val
595 600 605
Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys
610 615 620
Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu
625 630 635 640
Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu
645 650 655
Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr
660 665 670
His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu
675 680 685
Cys
<210> 68
<211> 443
<212> PRT
<213> 人工序列
<220>
<223> VH1a (CD40) (VHCH1) Fc 孔_PGLALA
<400> 68
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440
<210> 69
<211> 907
<212> PRT
<213> 人工序列
<220>
<223> VH1a (CD40)(VHCH1带电荷的)_VH1a (CD40)(VHCH1 带电荷的)-Fc
突起_PGLALA_(P1AE1689) (VL-CH1)
<400> 69
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Glu
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Glu Lys Val Glu Pro Lys Ser Cys Asp Gly Gly Gly Gly Ser Gly
210 215 220
Gly Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys
225 230 235 240
Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser
245 250 255
Phe Thr Gly Tyr Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser
260 265 270
Leu Glu Trp Met Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr
275 280 285
Asn Gln Lys Phe Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile
290 295 300
Ser Thr Ala Tyr Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala
305 310 315 320
Val Tyr Tyr Cys Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr
325 330 335
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
340 345 350
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
355 360 365
Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
370 375 380
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
385 390 395 400
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
405 410 415
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
420 425 430
Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
435 440 445
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
450 455 460
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
465 470 475 480
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
485 490 495
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
500 505 510
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
515 520 525
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
530 535 540
Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr
545 550 555 560
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
565 570 575
Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys
580 585 590
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
595 600 605
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
610 615 620
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
625 630 635 640
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
645 650 655
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly
660 665 670
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
675 680 685
Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu
690 695 700
Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Glu Ser Val
705 710 715 720
Asp Asn Tyr Gly Leu Ser Phe Ile Asn Trp Phe Gln Gln Lys Pro Gly
725 730 735
Gln Ala Pro Arg Leu Leu Ile Tyr Gly Thr Ser Asn Arg Gly Ser Gly
740 745 750
Ile Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu
755 760 765
Thr Ile Ser Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Phe Cys Gln
770 775 780
Gln Ser Asn Glu Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu
785 790 795 800
Ile Lys Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala
805 810 815
Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu
820 825 830
Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly
835 840 845
Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser
850 855 860
Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu
865 870 875 880
Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr
885 890 895
Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
900 905
<210> 70
<211> 671
<212> PRT
<213> 人工序列
<220>
<223> VH1a (CD40)(VHCH1 带电荷的)_VH1a (CD40)(VHCH1 带电荷的)-Fc
孔_PGLALA
<400> 70
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Glu
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Glu Lys Val Glu Pro Lys Ser Cys Asp Gly Gly Gly Gly Ser Gly
210 215 220
Gly Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys
225 230 235 240
Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser
245 250 255
Phe Thr Gly Tyr Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser
260 265 270
Leu Glu Trp Met Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr
275 280 285
Asn Gln Lys Phe Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile
290 295 300
Ser Thr Ala Tyr Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala
305 310 315 320
Val Tyr Tyr Cys Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr
325 330 335
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
340 345 350
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
355 360 365
Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
370 375 380
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
385 390 395 400
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
405 410 415
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
420 425 430
Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
435 440 445
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
450 455 460
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
465 470 475 480
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
485 490 495
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
500 505 510
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
515 520 525
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
530 535 540
Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr
545 550 555 560
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu
565 570 575
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys
580 585 590
Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
595 600 605
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
610 615 620
Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser
625 630 635 640
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
645 650 655
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
660 665 670
<210> 71
<211> 213
<212> PRT
<213> 人工序列
<220>
<223> 4B9 轻链交叉 VL-CH1
<400> 71
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Thr Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Asn Val Gly Ser Arg Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Ile Met Leu Pro
85 90 95
Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Ser Ser Ala Ser
100 105 110
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
115 120 125
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
130 135 140
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
145 150 155 160
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
165 170 175
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
180 185 190
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
195 200 205
Glu Pro Lys Ser Cys
210
<210> 72
<211> 687
<212> PRT
<213> 人工序列
<220>
<223> VH1a (CD40) (VHCH1) Fc 突起_PGLALA_4B9 (VH-Cκ)
<400> 72
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly Gly Gly Gly Ser
435 440 445
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu
450 455 460
Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser
465 470 475 480
Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr Ala
485 490 495
Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
500 505 510
Ala Ile Ile Gly Ser Gly Ala Ser Thr Tyr Tyr Ala Asp Ser Val Lys
515 520 525
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
530 535 540
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
545 550 555 560
Lys Gly Trp Phe Gly Gly Phe Asn Tyr Trp Gly Gln Gly Thr Leu Val
565 570 575
Thr Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile Phe Pro
580 585 590
Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu
595 600 605
Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp
610 615 620
Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp
625 630 635 640
Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys
645 650 655
Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln
660 665 670
Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
675 680 685
<210> 73
<211> 915
<212> PRT
<213> 人工序列
<220>
<223> VH1a (CD40) (VHCH1) _VH1a (CD40) (VHCH1)-Fc 突起_PGLALA_(4B9)
(VH-Cκ)
<400> 73
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Gly Gly Gly Gly Ser Gly
210 215 220
Gly Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys
225 230 235 240
Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser
245 250 255
Phe Thr Gly Tyr Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser
260 265 270
Leu Glu Trp Met Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr
275 280 285
Asn Gln Lys Phe Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile
290 295 300
Ser Thr Ala Tyr Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala
305 310 315 320
Val Tyr Tyr Cys Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr
325 330 335
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
340 345 350
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
355 360 365
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
370 375 380
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
385 390 395 400
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
405 410 415
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
420 425 430
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
435 440 445
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
450 455 460
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
465 470 475 480
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
485 490 495
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
500 505 510
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
515 520 525
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
530 535 540
Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr
545 550 555 560
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
565 570 575
Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys
580 585 590
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
595 600 605
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
610 615 620
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
625 630 635 640
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
645 650 655
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly
660 665 670
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
675 680 685
Gly Gly Ser Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln
690 695 700
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
705 710 715 720
Ser Ser Tyr Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
725 730 735
Glu Trp Val Ser Ala Ile Ile Gly Ser Gly Ala Ser Thr Tyr Tyr Ala
740 745 750
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn
755 760 765
Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
770 775 780
Tyr Tyr Cys Ala Lys Gly Trp Phe Gly Gly Phe Asn Tyr Trp Gly Gln
785 790 795 800
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val
805 810 815
Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser
820 825 830
Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln
835 840 845
Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val
850 855 860
Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu
865 870 875 880
Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu
885 890 895
Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg
900 905 910
Gly Glu Cys
915
<210> 74
<211> 671
<212> PRT
<213> 人工序列
<220>
<223> VH1a (CD40) (VHCH1)_VH1a (CD40) (VHCH1)-Fc 孔_PGLALA
<400> 74
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Gly Gly Gly Gly Ser Gly
210 215 220
Gly Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys
225 230 235 240
Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser
245 250 255
Phe Thr Gly Tyr Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser
260 265 270
Leu Glu Trp Met Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr
275 280 285
Asn Gln Lys Phe Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile
290 295 300
Ser Thr Ala Tyr Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala
305 310 315 320
Val Tyr Tyr Cys Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr
325 330 335
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
340 345 350
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
355 360 365
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
370 375 380
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
385 390 395 400
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
405 410 415
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
420 425 430
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
435 440 445
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
450 455 460
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
465 470 475 480
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
485 490 495
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
500 505 510
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
515 520 525
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
530 535 540
Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr
545 550 555 560
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Cys Thr Leu
565 570 575
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Ser Cys
580 585 590
Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
595 600 605
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
610 615 620
Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val Asp Lys Ser
625 630 635 640
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
645 650 655
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
660 665 670
<210> 75
<211> 223
<212> PRT
<213> 人工序列
<220>
<223> 28H1 轻链交叉 VH-Cκ
<400> 75
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser His
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ala Ile Trp Ala Ser Gly Glu Gln Tyr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Lys Gly Trp Leu Gly Asn Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Ala Ser Val Ala Ala Pro Ser Val Phe Ile Phe Pro
115 120 125
Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu
130 135 140
Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp
145 150 155 160
Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp
165 170 175
Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys
180 185 190
Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln
195 200 205
Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215 220
<210> 76
<211> 676
<212> PRT
<213> 人工序列
<220>
<223> VH1a (CD40)(VHCH1 带电荷的)Fc 突起_PGLALA_28H1 (VL-CH1)
<400> 76
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Glu
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Glu Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly Gly Gly Gly Ser
435 440 445
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu
450 455 460
Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu
465 470 475 480
Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Arg Ser Tyr
485 490 495
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
500 505 510
Ile Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser Gly
515 520 525
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro
530 535 540
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Gln Val Ile Pro Pro
545 550 555 560
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Ser Ser Ala Ser Thr
565 570 575
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
580 585 590
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
595 600 605
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
610 615 620
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
625 630 635 640
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
645 650 655
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
660 665 670
Pro Lys Ser Cys
675
<210> 77
<211> 904
<212> PRT
<213> 人工序列
<220>
<223> VH1a (CD40)(VHCH1 带电荷的)_VH1a (CD40)(VHCH1 带电荷的)Fc
突起_PGLALA_28H1 (VL-CH1)
<400> 77
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser Leu Glu Trp Met
35 40 45
Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Thr Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Glu
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Glu Lys Val Glu Pro Lys Ser Cys Asp Gly Gly Gly Gly Ser Gly
210 215 220
Gly Gly Gly Ser Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys
225 230 235 240
Lys Pro Gly Ala Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser
245 250 255
Phe Thr Gly Tyr Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Ser
260 265 270
Leu Glu Trp Met Gly Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr
275 280 285
Asn Gln Lys Phe Lys Gly Arg Val Thr Leu Thr Val Asp Lys Ser Ile
290 295 300
Ser Thr Ala Tyr Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala
305 310 315 320
Val Tyr Tyr Cys Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr
325 330 335
Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
340 345 350
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
355 360 365
Cys Leu Val Glu Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
370 375 380
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
385 390 395 400
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
405 410 415
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
420 425 430
Asn Thr Lys Val Asp Glu Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
435 440 445
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
450 455 460
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
465 470 475 480
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
485 490 495
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
500 505 510
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
515 520 525
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
530 535 540
Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr
545 550 555 560
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
565 570 575
Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Trp Cys
580 585 590
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
595 600 605
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
610 615 620
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
625 630 635 640
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
645 650 655
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly
660 665 670
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
675 680 685
Gly Gly Ser Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu
690 695 700
Ser Pro Gly Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val
705 710 715 720
Ser Arg Ser Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
725 730 735
Arg Leu Leu Ile Ile Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Asp
740 745 750
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
755 760 765
Arg Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Gln
770 775 780
Val Ile Pro Pro Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Ser
785 790 795 800
Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser
805 810 815
Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp
820 825 830
Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr
835 840 845
Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr
850 855 860
Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln
865 870 875 880
Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp
885 890 895
Lys Lys Val Glu Pro Lys Ser Cys
900
<210> 78
<211> 748
<212> PRT
<213> 人工序列
<220>
<223> hu FAP 胞外域+poly-lys-标签+his6-标签
<400> 78
Arg Pro Ser Arg Val His Asn Ser Glu Glu Asn Thr Met Arg Ala Leu
1 5 10 15
Thr Leu Lys Asp Ile Leu Asn Gly Thr Phe Ser Tyr Lys Thr Phe Phe
20 25 30
Pro Asn Trp Ile Ser Gly Gln Glu Tyr Leu His Gln Ser Ala Asp Asn
35 40 45
Asn Ile Val Leu Tyr Asn Ile Glu Thr Gly Gln Ser Tyr Thr Ile Leu
50 55 60
Ser Asn Arg Thr Met Lys Ser Val Asn Ala Ser Asn Tyr Gly Leu Ser
65 70 75 80
Pro Asp Arg Gln Phe Val Tyr Leu Glu Ser Asp Tyr Ser Lys Leu Trp
85 90 95
Arg Tyr Ser Tyr Thr Ala Thr Tyr Tyr Ile Tyr Asp Leu Ser Asn Gly
100 105 110
Glu Phe Val Arg Gly Asn Glu Leu Pro Arg Pro Ile Gln Tyr Leu Cys
115 120 125
Trp Ser Pro Val Gly Ser Lys Leu Ala Tyr Val Tyr Gln Asn Asn Ile
130 135 140
Tyr Leu Lys Gln Arg Pro Gly Asp Pro Pro Phe Gln Ile Thr Phe Asn
145 150 155 160
Gly Arg Glu Asn Lys Ile Phe Asn Gly Ile Pro Asp Trp Val Tyr Glu
165 170 175
Glu Glu Met Leu Ala Thr Lys Tyr Ala Leu Trp Trp Ser Pro Asn Gly
180 185 190
Lys Phe Leu Ala Tyr Ala Glu Phe Asn Asp Thr Asp Ile Pro Val Ile
195 200 205
Ala Tyr Ser Tyr Tyr Gly Asp Glu Gln Tyr Pro Arg Thr Ile Asn Ile
210 215 220
Pro Tyr Pro Lys Ala Gly Ala Lys Asn Pro Val Val Arg Ile Phe Ile
225 230 235 240
Ile Asp Thr Thr Tyr Pro Ala Tyr Val Gly Pro Gln Glu Val Pro Val
245 250 255
Pro Ala Met Ile Ala Ser Ser Asp Tyr Tyr Phe Ser Trp Leu Thr Trp
260 265 270
Val Thr Asp Glu Arg Val Cys Leu Gln Trp Leu Lys Arg Val Gln Asn
275 280 285
Val Ser Val Leu Ser Ile Cys Asp Phe Arg Glu Asp Trp Gln Thr Trp
290 295 300
Asp Cys Pro Lys Thr Gln Glu His Ile Glu Glu Ser Arg Thr Gly Trp
305 310 315 320
Ala Gly Gly Phe Phe Val Ser Thr Pro Val Phe Ser Tyr Asp Ala Ile
325 330 335
Ser Tyr Tyr Lys Ile Phe Ser Asp Lys Asp Gly Tyr Lys His Ile His
340 345 350
Tyr Ile Lys Asp Thr Val Glu Asn Ala Ile Gln Ile Thr Ser Gly Lys
355 360 365
Trp Glu Ala Ile Asn Ile Phe Arg Val Thr Gln Asp Ser Leu Phe Tyr
370 375 380
Ser Ser Asn Glu Phe Glu Glu Tyr Pro Gly Arg Arg Asn Ile Tyr Arg
385 390 395 400
Ile Ser Ile Gly Ser Tyr Pro Pro Ser Lys Lys Cys Val Thr Cys His
405 410 415
Leu Arg Lys Glu Arg Cys Gln Tyr Tyr Thr Ala Ser Phe Ser Asp Tyr
420 425 430
Ala Lys Tyr Tyr Ala Leu Val Cys Tyr Gly Pro Gly Ile Pro Ile Ser
435 440 445
Thr Leu His Asp Gly Arg Thr Asp Gln Glu Ile Lys Ile Leu Glu Glu
450 455 460
Asn Lys Glu Leu Glu Asn Ala Leu Lys Asn Ile Gln Leu Pro Lys Glu
465 470 475 480
Glu Ile Lys Lys Leu Glu Val Asp Glu Ile Thr Leu Trp Tyr Lys Met
485 490 495
Ile Leu Pro Pro Gln Phe Asp Arg Ser Lys Lys Tyr Pro Leu Leu Ile
500 505 510
Gln Val Tyr Gly Gly Pro Cys Ser Gln Ser Val Arg Ser Val Phe Ala
515 520 525
Val Asn Trp Ile Ser Tyr Leu Ala Ser Lys Glu Gly Met Val Ile Ala
530 535 540
Leu Val Asp Gly Arg Gly Thr Ala Phe Gln Gly Asp Lys Leu Leu Tyr
545 550 555 560
Ala Val Tyr Arg Lys Leu Gly Val Tyr Glu Val Glu Asp Gln Ile Thr
565 570 575
Ala Val Arg Lys Phe Ile Glu Met Gly Phe Ile Asp Glu Lys Arg Ile
580 585 590
Ala Ile Trp Gly Trp Ser Tyr Gly Gly Tyr Val Ser Ser Leu Ala Leu
595 600 605
Ala Ser Gly Thr Gly Leu Phe Lys Cys Gly Ile Ala Val Ala Pro Val
610 615 620
Ser Ser Trp Glu Tyr Tyr Ala Ser Val Tyr Thr Glu Arg Phe Met Gly
625 630 635 640
Leu Pro Thr Lys Asp Asp Asn Leu Glu His Tyr Lys Asn Ser Thr Val
645 650 655
Met Ala Arg Ala Glu Tyr Phe Arg Asn Val Asp Tyr Leu Leu Ile His
660 665 670
Gly Thr Ala Asp Asp Asn Val His Phe Gln Asn Ser Ala Gln Ile Ala
675 680 685
Lys Ala Leu Val Asn Ala Gln Val Asp Phe Gln Ala Met Trp Tyr Ser
690 695 700
Asp Gln Asn His Gly Leu Ser Gly Leu Ser Thr Asn His Leu Tyr Thr
705 710 715 720
His Met Thr His Phe Leu Lys Gln Cys Phe Ser Leu Ser Asp Gly Lys
725 730 735
Lys Lys Lys Lys Lys Gly His His His His His His
740 745
<210> 79
<211> 761
<212> PRT
<213> 鼠
<400> 79
Met Lys Thr Trp Leu Lys Thr Val Phe Gly Val Thr Thr Leu Ala Ala
1 5 10 15
Leu Ala Leu Val Val Ile Cys Ile Val Leu Arg Pro Ser Arg Val Tyr
20 25 30
Lys Pro Glu Gly Asn Thr Lys Arg Ala Leu Thr Leu Lys Asp Ile Leu
35 40 45
Asn Gly Thr Phe Ser Tyr Lys Thr Tyr Phe Pro Asn Trp Ile Ser Glu
50 55 60
Gln Glu Tyr Leu His Gln Ser Glu Asp Asp Asn Ile Val Phe Tyr Asn
65 70 75 80
Ile Glu Thr Arg Glu Ser Tyr Ile Ile Leu Ser Asn Ser Thr Met Lys
85 90 95
Ser Val Asn Ala Thr Asp Tyr Gly Leu Ser Pro Asp Arg Gln Phe Val
100 105 110
Tyr Leu Glu Ser Asp Tyr Ser Lys Leu Trp Arg Tyr Ser Tyr Thr Ala
115 120 125
Thr Tyr Tyr Ile Tyr Asp Leu Gln Asn Gly Glu Phe Val Arg Gly Tyr
130 135 140
Glu Leu Pro Arg Pro Ile Gln Tyr Leu Cys Trp Ser Pro Val Gly Ser
145 150 155 160
Lys Leu Ala Tyr Val Tyr Gln Asn Asn Ile Tyr Leu Lys Gln Arg Pro
165 170 175
Gly Asp Pro Pro Phe Gln Ile Thr Tyr Thr Gly Arg Glu Asn Arg Ile
180 185 190
Phe Asn Gly Ile Pro Asp Trp Val Tyr Glu Glu Glu Met Leu Ala Thr
195 200 205
Lys Tyr Ala Leu Trp Trp Ser Pro Asp Gly Lys Phe Leu Ala Tyr Val
210 215 220
Glu Phe Asn Asp Ser Asp Ile Pro Ile Ile Ala Tyr Ser Tyr Tyr Gly
225 230 235 240
Asp Gly Gln Tyr Pro Arg Thr Ile Asn Ile Pro Tyr Pro Lys Ala Gly
245 250 255
Ala Lys Asn Pro Val Val Arg Val Phe Ile Val Asp Thr Thr Tyr Pro
260 265 270
His His Val Gly Pro Met Glu Val Pro Val Pro Glu Met Ile Ala Ser
275 280 285
Ser Asp Tyr Tyr Phe Ser Trp Leu Thr Trp Val Ser Ser Glu Arg Val
290 295 300
Cys Leu Gln Trp Leu Lys Arg Val Gln Asn Val Ser Val Leu Ser Ile
305 310 315 320
Cys Asp Phe Arg Glu Asp Trp His Ala Trp Glu Cys Pro Lys Asn Gln
325 330 335
Glu His Val Glu Glu Ser Arg Thr Gly Trp Ala Gly Gly Phe Phe Val
340 345 350
Ser Thr Pro Ala Phe Ser Gln Asp Ala Thr Ser Tyr Tyr Lys Ile Phe
355 360 365
Ser Asp Lys Asp Gly Tyr Lys His Ile His Tyr Ile Lys Asp Thr Val
370 375 380
Glu Asn Ala Ile Gln Ile Thr Ser Gly Lys Trp Glu Ala Ile Tyr Ile
385 390 395 400
Phe Arg Val Thr Gln Asp Ser Leu Phe Tyr Ser Ser Asn Glu Phe Glu
405 410 415
Gly Tyr Pro Gly Arg Arg Asn Ile Tyr Arg Ile Ser Ile Gly Asn Ser
420 425 430
Pro Pro Ser Lys Lys Cys Val Thr Cys His Leu Arg Lys Glu Arg Cys
435 440 445
Gln Tyr Tyr Thr Ala Ser Phe Ser Tyr Lys Ala Lys Tyr Tyr Ala Leu
450 455 460
Val Cys Tyr Gly Pro Gly Leu Pro Ile Ser Thr Leu His Asp Gly Arg
465 470 475 480
Thr Asp Gln Glu Ile Gln Val Leu Glu Glu Asn Lys Glu Leu Glu Asn
485 490 495
Ser Leu Arg Asn Ile Gln Leu Pro Lys Val Glu Ile Lys Lys Leu Lys
500 505 510
Asp Gly Gly Leu Thr Phe Trp Tyr Lys Met Ile Leu Pro Pro Gln Phe
515 520 525
Asp Arg Ser Lys Lys Tyr Pro Leu Leu Ile Gln Val Tyr Gly Gly Pro
530 535 540
Cys Ser Gln Ser Val Lys Ser Val Phe Ala Val Asn Trp Ile Thr Tyr
545 550 555 560
Leu Ala Ser Lys Glu Gly Ile Val Ile Ala Leu Val Asp Gly Arg Gly
565 570 575
Thr Ala Phe Gln Gly Asp Lys Phe Leu His Ala Val Tyr Arg Lys Leu
580 585 590
Gly Val Tyr Glu Val Glu Asp Gln Leu Thr Ala Val Arg Lys Phe Ile
595 600 605
Glu Met Gly Phe Ile Asp Glu Glu Arg Ile Ala Ile Trp Gly Trp Ser
610 615 620
Tyr Gly Gly Tyr Val Ser Ser Leu Ala Leu Ala Ser Gly Thr Gly Leu
625 630 635 640
Phe Lys Cys Gly Ile Ala Val Ala Pro Val Ser Ser Trp Glu Tyr Tyr
645 650 655
Ala Ser Ile Tyr Ser Glu Arg Phe Met Gly Leu Pro Thr Lys Asp Asp
660 665 670
Asn Leu Glu His Tyr Lys Asn Ser Thr Val Met Ala Arg Ala Glu Tyr
675 680 685
Phe Arg Asn Val Asp Tyr Leu Leu Ile His Gly Thr Ala Asp Asp Asn
690 695 700
Val His Phe Gln Asn Ser Ala Gln Ile Ala Lys Ala Leu Val Asn Ala
705 710 715 720
Gln Val Asp Phe Gln Ala Met Trp Tyr Ser Asp Gln Asn His Gly Ile
725 730 735
Ser Ser Gly Arg Ser Gln Asn His Leu Tyr Thr His Met Thr His Phe
740 745 750
Leu Lys Gln Cys Phe Ser Leu Ser Asp
755 760
<210> 80
<211> 749
<212> PRT
<213> 人工序列
<220>
<223> 鼠 FAP 胞外域+poly-lys-标签+his6-标签
<400> 80
Arg Pro Ser Arg Val Tyr Lys Pro Glu Gly Asn Thr Lys Arg Ala Leu
1 5 10 15
Thr Leu Lys Asp Ile Leu Asn Gly Thr Phe Ser Tyr Lys Thr Tyr Phe
20 25 30
Pro Asn Trp Ile Ser Glu Gln Glu Tyr Leu His Gln Ser Glu Asp Asp
35 40 45
Asn Ile Val Phe Tyr Asn Ile Glu Thr Arg Glu Ser Tyr Ile Ile Leu
50 55 60
Ser Asn Ser Thr Met Lys Ser Val Asn Ala Thr Asp Tyr Gly Leu Ser
65 70 75 80
Pro Asp Arg Gln Phe Val Tyr Leu Glu Ser Asp Tyr Ser Lys Leu Trp
85 90 95
Arg Tyr Ser Tyr Thr Ala Thr Tyr Tyr Ile Tyr Asp Leu Gln Asn Gly
100 105 110
Glu Phe Val Arg Gly Tyr Glu Leu Pro Arg Pro Ile Gln Tyr Leu Cys
115 120 125
Trp Ser Pro Val Gly Ser Lys Leu Ala Tyr Val Tyr Gln Asn Asn Ile
130 135 140
Tyr Leu Lys Gln Arg Pro Gly Asp Pro Pro Phe Gln Ile Thr Tyr Thr
145 150 155 160
Gly Arg Glu Asn Arg Ile Phe Asn Gly Ile Pro Asp Trp Val Tyr Glu
165 170 175
Glu Glu Met Leu Ala Thr Lys Tyr Ala Leu Trp Trp Ser Pro Asp Gly
180 185 190
Lys Phe Leu Ala Tyr Val Glu Phe Asn Asp Ser Asp Ile Pro Ile Ile
195 200 205
Ala Tyr Ser Tyr Tyr Gly Asp Gly Gln Tyr Pro Arg Thr Ile Asn Ile
210 215 220
Pro Tyr Pro Lys Ala Gly Ala Lys Asn Pro Val Val Arg Val Phe Ile
225 230 235 240
Val Asp Thr Thr Tyr Pro His His Val Gly Pro Met Glu Val Pro Val
245 250 255
Pro Glu Met Ile Ala Ser Ser Asp Tyr Tyr Phe Ser Trp Leu Thr Trp
260 265 270
Val Ser Ser Glu Arg Val Cys Leu Gln Trp Leu Lys Arg Val Gln Asn
275 280 285
Val Ser Val Leu Ser Ile Cys Asp Phe Arg Glu Asp Trp His Ala Trp
290 295 300
Glu Cys Pro Lys Asn Gln Glu His Val Glu Glu Ser Arg Thr Gly Trp
305 310 315 320
Ala Gly Gly Phe Phe Val Ser Thr Pro Ala Phe Ser Gln Asp Ala Thr
325 330 335
Ser Tyr Tyr Lys Ile Phe Ser Asp Lys Asp Gly Tyr Lys His Ile His
340 345 350
Tyr Ile Lys Asp Thr Val Glu Asn Ala Ile Gln Ile Thr Ser Gly Lys
355 360 365
Trp Glu Ala Ile Tyr Ile Phe Arg Val Thr Gln Asp Ser Leu Phe Tyr
370 375 380
Ser Ser Asn Glu Phe Glu Gly Tyr Pro Gly Arg Arg Asn Ile Tyr Arg
385 390 395 400
Ile Ser Ile Gly Asn Ser Pro Pro Ser Lys Lys Cys Val Thr Cys His
405 410 415
Leu Arg Lys Glu Arg Cys Gln Tyr Tyr Thr Ala Ser Phe Ser Tyr Lys
420 425 430
Ala Lys Tyr Tyr Ala Leu Val Cys Tyr Gly Pro Gly Leu Pro Ile Ser
435 440 445
Thr Leu His Asp Gly Arg Thr Asp Gln Glu Ile Gln Val Leu Glu Glu
450 455 460
Asn Lys Glu Leu Glu Asn Ser Leu Arg Asn Ile Gln Leu Pro Lys Val
465 470 475 480
Glu Ile Lys Lys Leu Lys Asp Gly Gly Leu Thr Phe Trp Tyr Lys Met
485 490 495
Ile Leu Pro Pro Gln Phe Asp Arg Ser Lys Lys Tyr Pro Leu Leu Ile
500 505 510
Gln Val Tyr Gly Gly Pro Cys Ser Gln Ser Val Lys Ser Val Phe Ala
515 520 525
Val Asn Trp Ile Thr Tyr Leu Ala Ser Lys Glu Gly Ile Val Ile Ala
530 535 540
Leu Val Asp Gly Arg Gly Thr Ala Phe Gln Gly Asp Lys Phe Leu His
545 550 555 560
Ala Val Tyr Arg Lys Leu Gly Val Tyr Glu Val Glu Asp Gln Leu Thr
565 570 575
Ala Val Arg Lys Phe Ile Glu Met Gly Phe Ile Asp Glu Glu Arg Ile
580 585 590
Ala Ile Trp Gly Trp Ser Tyr Gly Gly Tyr Val Ser Ser Leu Ala Leu
595 600 605
Ala Ser Gly Thr Gly Leu Phe Lys Cys Gly Ile Ala Val Ala Pro Val
610 615 620
Ser Ser Trp Glu Tyr Tyr Ala Ser Ile Tyr Ser Glu Arg Phe Met Gly
625 630 635 640
Leu Pro Thr Lys Asp Asp Asn Leu Glu His Tyr Lys Asn Ser Thr Val
645 650 655
Met Ala Arg Ala Glu Tyr Phe Arg Asn Val Asp Tyr Leu Leu Ile His
660 665 670
Gly Thr Ala Asp Asp Asn Val His Phe Gln Asn Ser Ala Gln Ile Ala
675 680 685
Lys Ala Leu Val Asn Ala Gln Val Asp Phe Gln Ala Met Trp Tyr Ser
690 695 700
Asp Gln Asn His Gly Ile Leu Ser Gly Arg Ser Gln Asn His Leu Tyr
705 710 715 720
Thr His Met Thr His Phe Leu Lys Gln Cys Phe Ser Leu Ser Asp Gly
725 730 735
Lys Lys Lys Lys Lys Lys Gly His His His His His His
740 745
<210> 81
<211> 748
<212> PRT
<213> 人工序列
<220>
<223> 食蟹猴 FAP 胞外域+poly-lys-标签+his6-标签
<400> 81
Arg Pro Pro Arg Val His Asn Ser Glu Glu Asn Thr Met Arg Ala Leu
1 5 10 15
Thr Leu Lys Asp Ile Leu Asn Gly Thr Phe Ser Tyr Lys Thr Phe Phe
20 25 30
Pro Asn Trp Ile Ser Gly Gln Glu Tyr Leu His Gln Ser Ala Asp Asn
35 40 45
Asn Ile Val Leu Tyr Asn Ile Glu Thr Gly Gln Ser Tyr Thr Ile Leu
50 55 60
Ser Asn Arg Thr Met Lys Ser Val Asn Ala Ser Asn Tyr Gly Leu Ser
65 70 75 80
Pro Asp Arg Gln Phe Val Tyr Leu Glu Ser Asp Tyr Ser Lys Leu Trp
85 90 95
Arg Tyr Ser Tyr Thr Ala Thr Tyr Tyr Ile Tyr Asp Leu Ser Asn Gly
100 105 110
Glu Phe Val Arg Gly Asn Glu Leu Pro Arg Pro Ile Gln Tyr Leu Cys
115 120 125
Trp Ser Pro Val Gly Ser Lys Leu Ala Tyr Val Tyr Gln Asn Asn Ile
130 135 140
Tyr Leu Lys Gln Arg Pro Gly Asp Pro Pro Phe Gln Ile Thr Phe Asn
145 150 155 160
Gly Arg Glu Asn Lys Ile Phe Asn Gly Ile Pro Asp Trp Val Tyr Glu
165 170 175
Glu Glu Met Leu Ala Thr Lys Tyr Ala Leu Trp Trp Ser Pro Asn Gly
180 185 190
Lys Phe Leu Ala Tyr Ala Glu Phe Asn Asp Thr Asp Ile Pro Val Ile
195 200 205
Ala Tyr Ser Tyr Tyr Gly Asp Glu Gln Tyr Pro Arg Thr Ile Asn Ile
210 215 220
Pro Tyr Pro Lys Ala Gly Ala Lys Asn Pro Phe Val Arg Ile Phe Ile
225 230 235 240
Ile Asp Thr Thr Tyr Pro Ala Tyr Val Gly Pro Gln Glu Val Pro Val
245 250 255
Pro Ala Met Ile Ala Ser Ser Asp Tyr Tyr Phe Ser Trp Leu Thr Trp
260 265 270
Val Thr Asp Glu Arg Val Cys Leu Gln Trp Leu Lys Arg Val Gln Asn
275 280 285
Val Ser Val Leu Ser Ile Cys Asp Phe Arg Glu Asp Trp Gln Thr Trp
290 295 300
Asp Cys Pro Lys Thr Gln Glu His Ile Glu Glu Ser Arg Thr Gly Trp
305 310 315 320
Ala Gly Gly Phe Phe Val Ser Thr Pro Val Phe Ser Tyr Asp Ala Ile
325 330 335
Ser Tyr Tyr Lys Ile Phe Ser Asp Lys Asp Gly Tyr Lys His Ile His
340 345 350
Tyr Ile Lys Asp Thr Val Glu Asn Ala Ile Gln Ile Thr Ser Gly Lys
355 360 365
Trp Glu Ala Ile Asn Ile Phe Arg Val Thr Gln Asp Ser Leu Phe Tyr
370 375 380
Ser Ser Asn Glu Phe Glu Asp Tyr Pro Gly Arg Arg Asn Ile Tyr Arg
385 390 395 400
Ile Ser Ile Gly Ser Tyr Pro Pro Ser Lys Lys Cys Val Thr Cys His
405 410 415
Leu Arg Lys Glu Arg Cys Gln Tyr Tyr Thr Ala Ser Phe Ser Asp Tyr
420 425 430
Ala Lys Tyr Tyr Ala Leu Val Cys Tyr Gly Pro Gly Ile Pro Ile Ser
435 440 445
Thr Leu His Asp Gly Arg Thr Asp Gln Glu Ile Lys Ile Leu Glu Glu
450 455 460
Asn Lys Glu Leu Glu Asn Ala Leu Lys Asn Ile Gln Leu Pro Lys Glu
465 470 475 480
Glu Ile Lys Lys Leu Glu Val Asp Glu Ile Thr Leu Trp Tyr Lys Met
485 490 495
Ile Leu Pro Pro Gln Phe Asp Arg Ser Lys Lys Tyr Pro Leu Leu Ile
500 505 510
Gln Val Tyr Gly Gly Pro Cys Ser Gln Ser Val Arg Ser Val Phe Ala
515 520 525
Val Asn Trp Ile Ser Tyr Leu Ala Ser Lys Glu Gly Met Val Ile Ala
530 535 540
Leu Val Asp Gly Arg Gly Thr Ala Phe Gln Gly Asp Lys Leu Leu Tyr
545 550 555 560
Ala Val Tyr Arg Lys Leu Gly Val Tyr Glu Val Glu Asp Gln Ile Thr
565 570 575
Ala Val Arg Lys Phe Ile Glu Met Gly Phe Ile Asp Glu Lys Arg Ile
580 585 590
Ala Ile Trp Gly Trp Ser Tyr Gly Gly Tyr Val Ser Ser Leu Ala Leu
595 600 605
Ala Ser Gly Thr Gly Leu Phe Lys Cys Gly Ile Ala Val Ala Pro Val
610 615 620
Ser Ser Trp Glu Tyr Tyr Ala Ser Val Tyr Thr Glu Arg Phe Met Gly
625 630 635 640
Leu Pro Thr Lys Asp Asp Asn Leu Glu His Tyr Lys Asn Ser Thr Val
645 650 655
Met Ala Arg Ala Glu Tyr Phe Arg Asn Val Asp Tyr Leu Leu Ile His
660 665 670
Gly Thr Ala Asp Asp Asn Val His Phe Gln Asn Ser Ala Gln Ile Ala
675 680 685
Lys Ala Leu Val Asn Ala Gln Val Asp Phe Gln Ala Met Trp Tyr Ser
690 695 700
Asp Gln Asn His Gly Leu Ser Gly Leu Ser Thr Asn His Leu Tyr Thr
705 710 715 720
His Met Thr His Phe Leu Lys Gln Cys Phe Ser Leu Ser Asp Gly Lys
725 730 735
Lys Lys Lys Lys Lys Gly His His His His His His
740 745
<210> 82
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 肽连接基 G4S
<400> 82
Gly Gly Gly Gly Ser
1 5
<210> 83
<211> 10
<212> PRT
<213> 人工序列
<220>
<223> 肽连接基 (G4S)2
<400> 83
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 84
<211> 10
<212> PRT
<213> 人工序列
<220>
<223> 肽连接基 (SG4)2
<400> 84
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
1 5 10
<210> 85
<211> 14
<212> PRT
<213> 人工序列
<220>
<223> 肽连接基 G4(SG4)2
<400> 85
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
1 5 10
<210> 86
<211> 10
<212> PRT
<213> 人工序列
<220>
<223> 肽连接基
<400> 86
Gly Ser Pro Gly Ser Ser Ser Ser Gly Ser
1 5 10
<210> 87
<211> 15
<212> PRT
<213> 人工序列
<220>
<223> 肽连接基 (G4S)3
<400> 87
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 88
<211> 20
<212> PRT
<213> 人工序列
<220>
<223> 肽连接基 (G4S)4
<400> 88
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser
20
<210> 89
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> 肽连接基
<400> 89
Gly Ser Gly Ser Gly Ser Gly Ser
1 5
<210> 90
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> 肽连接基
<400> 90
Gly Ser Gly Ser Gly Asn Gly Ser
1 5
<210> 91
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> 肽连接基
<400> 91
Gly Gly Ser Gly Ser Gly Ser Gly
1 5
<210> 92
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 肽连接基
<400> 92
Gly Gly Ser Gly Ser Gly
1 5
<210> 93
<211> 4
<212> PRT
<213> 人工序列
<220>
<223> 肽连接基
<400> 93
Gly Gly Ser Gly
1
<210> 94
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> 肽连接基
<400> 94
Gly Gly Ser Gly Asn Gly Ser Gly
1 5
<210> 95
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> 肽连接基
<400> 95
Gly Gly Asn Gly Ser Gly Ser Gly
1 5
<210> 96
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 肽连接基
<400> 96
Gly Gly Asn Gly Ser Gly
1 5
<210> 97
<211> 227
<212> PRT
<213> 人工序列
<220>
<223> Fc 突起链
<400> 97
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<210> 98
<211> 227
<212> PRT
<213> 人工序列
<220>
<223> Fc 孔链
<400> 98
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<210> 99
<211> 20
<212> PRT
<213> 智人
<400> 99
Tyr Tyr Tyr Tyr Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Thr Val
1 5 10 15
Thr Val Ser Ser
20
<210> 100
<211> 12
<212> PRT
<213> 智人
<400> 100
Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
1 5 10
<210> 101
<211> 20
<212> PRT
<213> 智人
<400> 101
Tyr Tyr Tyr Tyr Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Thr Val
1 5 10 15
Thr Val Ser Ser
20
<210> 102
<211> 12
<212> PRT
<213> 智人
<400> 102
Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
1 5 10
<210> 103
<211> 20
<212> PRT
<213> 智人
<400> 103
Tyr Tyr Tyr Tyr Tyr Gly Met Asp Val Trp Gly Gln Gly Thr Thr Val
1 5 10 15
Thr Val Ser Ser
20
<210> 104
<211> 12
<212> PRT
<213> 智人
<400> 104
Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
1 5 10
<210> 105
<211> 219
<212> PRT
<213> 人工序列
<220>
<223> huCD40 轻链(带电荷的)
<400> 105
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ser Ser Gln Ser Leu Val His Ser
20 25 30
Asn Gly Asn Thr Phe Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala
35 40 45
Pro Lys Leu Leu Ile Tyr Thr Val Ser Asn Arg Phe Ser Gly Val Pro
50 55 60
Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
65 70 75 80
Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Ser Gln Thr
85 90 95
Thr His Val Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Arg
115 120 125
Lys Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 106
<211> 676
<212> PRT
<213> 人工序列
<220>
<223> huCD40(VHCH1 带电荷的)Fc PGLALA FAP (VL-CH1)
<400> 106
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Val Ile Pro Asn Ala Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Arg Phe Thr Leu Ser Val Asp Asn Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ile Tyr Trp Trp Gly Gln Gly Thr Leu Val Thr Val
100 105 110
Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser
115 120 125
Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Glu
130 135 140
Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu
145 150 155 160
Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu
165 170 175
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
180 185 190
Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val
195 200 205
Asp Glu Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro
210 215 220
Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe
225 230 235 240
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
245 250 255
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
260 265 270
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
275 280 285
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
290 295 300
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
305 310 315 320
Ser Asn Lys Ala Leu Gly Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
325 330 335
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
340 345 350
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
355 360 365
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
370 375 380
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
385 390 395 400
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
405 410 415
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
420 425 430
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly Gly Gly Gly Ser
435 440 445
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu
450 455 460
Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly Glu
465 470 475 480
Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Arg Ser Tyr
485 490 495
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
500 505 510
Ile Gly Ala Ser Thr Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser Gly
515 520 525
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu Pro
530 535 540
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Gly Gln Val Ile Pro Pro
545 550 555 560
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Ser Ser Ala Ser Thr
565 570 575
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
580 585 590
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
595 600 605
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
610 615 620
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
625 630 635 640
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
645 650 655
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
660 665 670
Pro Lys Ser Cys
675
<210> 107
<211> 118
<212> PRT
<213> 人工序列
<220>
<223> VH (PD-L1)
<400> 107
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Ser
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Trp Ile Ser Pro Tyr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg His Trp Pro Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 108
<211> 107
<212> PRT
<213> 人工序列
<220>
<223> VL (PD-L1)
<400> 108
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Ser Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Leu Tyr His Pro Ala
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 109
<211> 121
<212> PRT
<213> 人工序列
<220>
<223> VH (PD-L1)
<400> 109
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Tyr
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asn Ile Lys Gln Asp Gly Ser Glu Lys Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Gly Trp Phe Gly Glu Leu Ala Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 110
<211> 108
<212> PRT
<213> 人工序列
<220>
<223> VL (PD-L1)
<400> 110
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Arg Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Asp Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Leu Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
- 包含抗FAP克隆212的双特异性抗原结合分子
- 包含抗4-1BB克隆20H4.9的双特异性抗原结合分子