一种兔抗小鼠Ccdc189多克隆抗体的制备方法和应用
文献发布时间:2023-06-19 19:28:50
技术领域
本发明属于生物医药技术领域,具体涉及一种兔抗小鼠Ccdc189多克隆抗体的制备方法和应用。
背景技术
据了解,CCDC(coiled-coil domain containing,CCDC)蛋白家族是一种由 2-7个α螺旋互相缠绕形成的蛋白质超二级结构。CCDC蛋白在组织中广泛表达,其细胞定位也呈多样化,因而参与了机体多种生理过程,有研究显示,CCDC 基因表达异常可以影响细胞生长因子的水平,以及配子发生、胚胎发育、造血、血管生成和纤毛发育等多种进程。Ccdc189蛋白是CCDC蛋白家族中的一员,是一种精子尾部相关蛋白,主要定位于成熟精子的主段和顶体区域。已有研究表明,Ccdc189基因突变与北欧红奶牛弱精子症相关,Ccdc189基因突变会导致公牛睾丸线粒体功能障碍,精子鞭毛运动丧失,产生雄性不育表型。Ccdc189 蛋白在睾丸组织中特异性高表达,其基因敲除雄性小鼠不育,但具体作用及分子机制尚未阐明。然而,Ccdc189在人类及小鼠中是否发挥重要作用目前尚未可知,其在雄性生殖系统中作用阶段及调控的分子机制至今也未见任何报道。因此,深入探索Ccdc189的作用及发生机制,有助于为男性不育症的诊断及治疗提供新的理论基础和实践参考价值。
为探索Ccdc189的功能,需对其进行功能分析及作用机制进行研究。经检索,目前未发现有商品化的抗小鼠Ccdc189抗体出售,同时经文献检索,也未发现抗小鼠Ccdc189的多克隆及单克隆抗体制备的文献报道。
为研究Ccdc189的功能,研究其在精子发生过程中的具体作用及分子机制,亟需一种Ccdc189多克隆抗体。
发明内容
本发明针对所要解决的技术问题,克服现有技术的不足而提供一种兔抗小鼠Ccdc189多克隆抗体的制备方法和应用。
本发明的目的之一在于,提供一种兔抗小鼠Ccdc189多克隆抗体的制备方法,该方法包括以下步骤:
S1、Ccdc189原核蛋白表达重组质粒的构建
设计一对Ccdc189基因开放阅读框全长序列特异引物,以Ccdc189真核表达质粒为模板,利用Ccdc189基因开放阅读框全长序列特异引物进行PCR扩增,将PCR扩增产物经胶回收后与pGEM-T-Easy载体连接,连接产物转化入大肠杆菌感受态细胞中,经培养获得pGEM-T-Easy-Ccdc189质粒,将 pGEM-T-Easy-Ccdc189质粒、pET-28a(+)质粒分别酶切后胶回收,获得Ccdc189 目的片段、pET-28a表达载体,Ccdc189目的片段与pET-28a表达载体连接后转化到大肠杆菌感受态细胞中,培养得到pET-28a-Ccdc189重组质粒;
S2、Ccdc189原核蛋白的诱导表达与纯化
将pET-28a-Ccdc189重组质粒转入E.coli BL21感受态细胞,诱导培养,离心收集菌体,菌体经重悬、裂解、离心洗涤,即得Ccdc189原核蛋白,通过Ni-NTA 亲和层析柱对Ccdc189原核蛋白进行纯化,并采用唑咪洗脱液洗脱,得到 Ccdc189纯化蛋白;
S3、Ccdc189多克隆抗体的制备
以Ccdc189纯化蛋白为免疫原,对兔进行免疫,收集兔抗血清,纯化得到兔抗鼠Ccdc189多克隆抗体。
本发明首先运用分子克隆方法构建了pET28a-Ccdc189重组质粒,在大肠杆菌中诱导表达Ccdc189蛋白,将纯化后Ccdc189原核蛋白免疫新西兰大白兔从而获取Ccdc189多克隆抗体。应用制备的Ccdc189多克隆抗体,对小鼠睾丸组织及人睾丸组织进行WesternBlot、免疫荧光分析,结果发现,该抗体能够特异性识别小鼠睾丸组织及人睾丸组织中的Ccdc189蛋白。本发明为研究Ccdc189 的功能提供了有利工具及技术支持,同时也有可能为临床男性不育症的诊断和治疗提供新的靶标。
本发明进一步优化的技术方案如下:
本发明优先采用Ccdc189变异剪接体1(NM_001033040)基因开放阅读框全长序列设计引物,采用特异性酶切位点EcoRI、HindIII,应用Gene Runner 软件设计Ccdc189开放阅读框全长序列特异引物,其中前向引物: 5’-GGGAATTCATGATTACCCCCAGTTCCAG-3’,反向引物: 5’-GGAAGCTTGCTTGGTCTTGGTCTTGCC-3’。
Ccdc189变异剪接体1基因开放阅读框的核苷酸序列如SEQ ID NO.3所示。所表达的原核蛋白为Ccdc189变异剪接体1编码蛋白全长(NP_001028212),其氨基酸序列如SEQ IDNO.4所示。
所述步骤S1中,PCR反应条件为:95℃,5min;95℃ 30s、60℃ 45s、72℃ 62s,共35个循环;72℃ 10min,4℃保存PCR扩增产物;
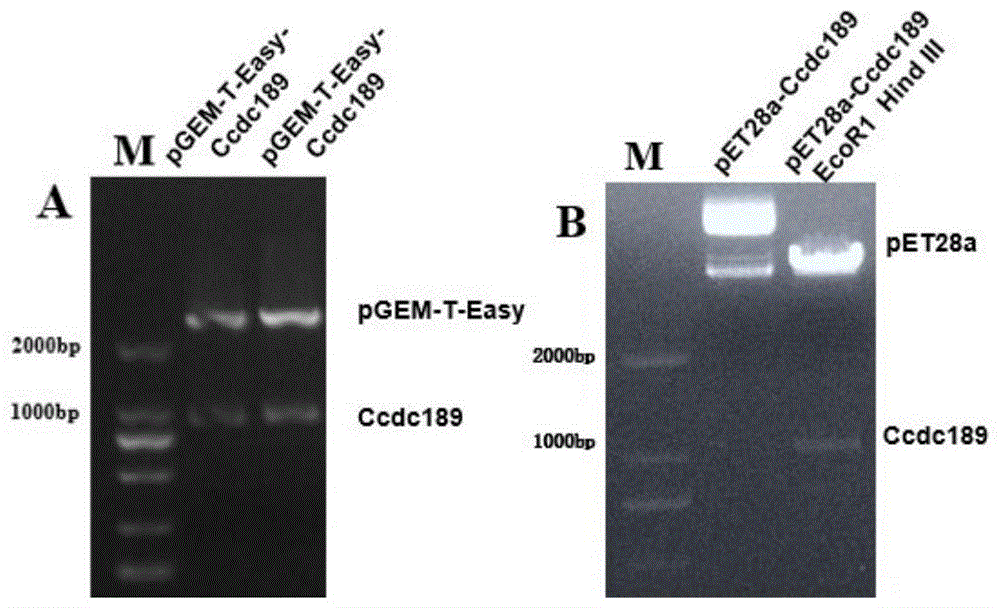
Ccdc189原核蛋白表达重组质粒的具体构建方法为:应用琼脂糖凝胶电泳分离PCR扩增产物并切胶纯化,利用T4-DNA连接酶将纯化的PCR产物与 pGEM-T-Easy载体相连接,16℃反应过夜,连接产物转化大肠杆菌JM109感受态细胞后,与IPTG、X-gal涂布于氨苄青霉素抗性的LB固体培养板上;挑选白色单克隆菌落,摇菌提取质粒,使用限制性内切酶EcoRI水浴酶切后,琼脂糖凝胶电泳检测酶切产物,将连接成功的pGEM-T-Easy-Ccdc189重组质粒进行测序鉴定;使用限制性内切酶EcoRI和Hind III将测序正确的 pGEM-T-Easy-Ccdc189质粒与pET-28a(+)质粒进行酶切,用琼脂糖凝胶电泳分离并回收酶切产物,将回收的Ccdc189目的片段与pET-28a表达载体相连接,转化大肠杆菌JM109感受态细胞后,涂布于卡那霉素抗性的LB固体平板上培养;挑选单克隆菌落,经限制性内切酶EcoRI和Hind III双酶切鉴定,确定 pET-28a-Ccdc189重组质粒构建成功。
所述步骤S2中,Ccdc189原核蛋白诱导表达的具体方法为:
将pET-28a-Ccdc189重组质粒转化E.coli BL21感受态细胞,在卡那霉素抗性的LB固体培养基上培养,挑选阳性单克隆菌落置于卡那霉素抗性的LB液中,培养过夜,将过夜培养的菌液加入卡那霉素抗性的LB液中,培养一段时间后,取菌液于试管中,加入诱导剂IPTG进行原核诱导表达,离心收集菌体,加入重悬缓冲液充分重悬后,加入细菌裂解液裂解,裂解后的菌液超声离心收集沉淀,采用洗涤液洗涤后溶于尿素溶液中静置过夜,离心收集上清;
Ccdc189原核蛋白纯化的具体方法为:将上清液过Ni-NTA亲和层析柱厚采用咪唑溶液洗脱层析柱,然后将洗脱液装入处理好的透析袋中,置入7M、6M、 5M、4M、3M、2M尿素溶液中进行梯度复性,收集复性后的蛋白溶液。
所述步骤S3中,制备Ccdc189多克隆抗体的方法为:以复性后的Ccdc189 原核蛋白作为免疫原,免疫成年雄性新西兰大白兔,初次时免疫将复性后的 Ccdc189原核蛋白与福氏完全佐剂等体积混合,冰上乳化,背部皮下多点注射免疫成年新西兰大白兔,然后将复性后的Ccdc189原核蛋白与福氏不完全佐剂等体积混合,每隔2周免疫1次,免疫5次。
本发明的目的只为在于,提供一种兔抗小鼠Ccdc189多克隆抗体,其抗原的氨基酸序列如SEQ ID NO.4所示,为深入研究Ccdc189蛋白的功能及分子机制提供可能。
应用本发明的方法可以重复制备多克隆抗体,因实验动物个体差异可能导致不同实验组抗体效价出现波动,但其均可用于Western Blot、ELISA、免疫组织荧光分析等。建议同时免疫5只实验用兔制备抗体。
本发明的目的之三在于,提供兔抗小鼠Ccdc189多克隆抗体在研究Ccdc189 蛋白的功能及作用机制中的应用。
上述兔抗小鼠Ccdc189多克隆抗体在研究Ccdc蛋白的功能及作用机制中的应用,具体包括酶联免疫吸附剂测定(ELISA)、免疫印迹试验(Western Blot)、免疫荧光分析、免疫组织化学分析。
本发明所提供的兔抗小鼠Ccdc189多克隆抗体能特异性识别小鼠及人组织中的Ccdc189蛋白,可应用于小鼠及人多种组织ELISA、Western Blot、免疫荧光、免疫组织化学等分析,具有特异性高、适用范围广泛等特点。
本发明的有益效果如下:本发明的制备方法采用分子生物学和基因工程的方法构建了Ccdc189基因的重组表达载体,诱导表达,亲和层析获得纯化的原核表达蛋白,所制备的Ccdc189多克隆抗体具有很强的特异性,可用于酶联免疫吸附剂测定、免疫印迹试验、免疫荧光分析、免疫组织化学分析等技术,为深入研究Ccdc蛋白的功能提供了重要工具。
本发明应用Ccdc189基因全长可读框序列进行原核蛋白的表达,经纯化复性浓缩后获得高纯度的Ccdc189原核蛋白,应用该蛋白混合佐剂免疫成年雄性新西兰大白兔,经过5次免疫后,获得的抗小鼠Ccdc189多克隆抗体效价达到 1:10
本发明提供的抗小鼠Ccdc189多克隆抗体的应用范围较广,能有效识别小鼠和人睾丸组织中的Ccdc189蛋白,可用于Ccdc189蛋白的ELISA、Western Blot 及免疫荧光等技术分析,更多的应用领域还有待验证。
本发明提供的这种多克隆抗体将为研究小鼠Ccdc189的功能及作用机制提供有力的技术支持。
附图说明
图1为本发明中pET-28a-Ccdc189重组质粒成功构建后酶切鉴定分析的结果示意图。图中:M表示DNA marker;A表示pGEM-T-Easy-Ccdc189重组质粒EcoR1酶切鉴定结果;B表示pET-28a-Ccdc189重组质粒EcoR1、HindⅢ双酶切鉴定结果。
图2为本发明中SDS-PAGE分析Ccdc189原核蛋白成功诱导表达的结果示意图。图中:M表示蛋白质marker;IPTG-表示IPTG未诱导的大肠杆菌;IPTG+ 表示IPTG诱导后的大肠杆菌;IPTG-沉淀表示IPTG未诱导的大肠杆菌超声沉淀;IPTG-上清表示IPTG未诱导的大肠杆菌超声上清;IPTG+I沉淀表示IPTG 诱导的大肠杆菌第一次超声沉淀;IPTG+I上清表示IPTG诱导的大肠杆菌第一次超声上清;IPTG+II沉淀表示IPTG诱导的大肠杆菌第二次超声沉淀;IPTG+II 上清表示IPTG诱导的大肠杆菌第二次超声上清。
图3为本发明中SDS-PAGE分析Ccdc189原核蛋白富集纯的结果示意图。图中:M表示蛋白质marker;1表示Ccdc189原核蛋白溶液;2表示0mmol/L咪唑洗脱液;3表示25mmol/L咪唑洗脱液;4表示50mmol/L咪唑洗脱液;5表示100mmol/L咪唑洗脱液;6表示200mmol/L咪唑洗脱液。
图4为本发明中ELISA检测Ccdc189多克隆抗体的效价均达到1:10
图5为本发明中Ccdc189多克隆抗体特异性识别野生型小鼠睾丸组织中 Ccdc189蛋白的Western Blot分析示意图。图中:HOM表示Ccdc189基因敲除纯合子小鼠睾丸组织;WT表示成年野生型小鼠睾丸组织。
图6为本发明中Ccdc189多克隆抗体能特异性识别定位人及野生型小鼠睾丸组织中Ccdc189蛋白的细胞免疫荧光分析示意图。图中:A表示Ccdc189蛋白在野生型小鼠睾丸组织中定位,Bar=100um;B表示Ccdc189蛋白在野生型小鼠睾丸组织中定位,Bar=50um;C表示Ccdc189蛋白在人睾丸组织中定位, Bar=100um;D表示Ccdc189蛋白在人睾丸组织中定位,Bar=50um。
具体实施方式
下面结合实施例对本发明的技术方案做进一步的详细说明:本实施例在以本发明技术方案为前提下进行实施,给出了详细的实施方式和具体的操作过程,但本发明的保护权限不限于下述的实施例。
实施例所涉及试剂及材料的来源如下:E.coli JM109及E.coliBL21(DE3)感受态细胞均由扬州大学医学院妇产与生殖医学研究所制备并保存,可免费或有偿向大众发放;pET-28a(+)表达载体购自Clontech公司;AlexaFluo 555标记的驴抗兔IgG购自赛默飞世尔科技(中国)有限公司;pGEM-T-Easy试剂盒购自Progema公司;限制性内切酶EcoRI、HindIII、高保真Taq DNA聚合酶、T4 DNA连接酶购自TaKaRa公司;Freund完全佐剂、Freund不完全佐剂购自 Sigma公司;Ni-NTA亲和层析树脂购自上海锐享生物科技有限公司;辣根过氧化物酶(horseradish peroxidase,HRP)标记的山羊抗兔IgG购自北京中杉金桥生物技术有限公司;二硫苏糖醇(DL-dithiothreitol,DTT)、四甲基联苯胺(tetramethylbenzidine,TMB)显色液、牛血清白蛋白(bovine serum albumin,BSA)及4',6-二脒基-2-苯基吲哚(4',6-diamidino-2-phenylindole,DAPI)、异丙基-β-D-硫代吡喃半乳糖苷(Isopropylβ-D-1-thiogalactopyranoside,IPTG)5- 溴4-氯-3-吲哚-β-D-半乳糖苷(5-bromo-4-chloro-3-indolylβ -D-galactopyranoside,X-gal)、乙二胺四乙酸(EDTA)、盐酸三(羟甲基)氨基甲烷(Tris-HCL)、曲拉通Triton-X-100、咪唑、溶菌酶购自北京索莱宝科技有限公司;二辛可宁酸(bicinchoninincacid,BCA)蛋白定量试剂盒、DNaseI为脱氧核糖核酸酶I购自北京康为世纪生物科技有限公司;超敏ECL化学发光试剂盒购自上海新赛和生物科技有限公司。C57BL/6小鼠购自Jackson实验室, Ccdc189
除了上述特殊说明的生物材料及试剂外,本发明中所提及的其余材料及试剂为市购,公众能从国内外商业渠道购买到,此处不再一一说明。
本发明中的“%”为体积百分比。
实施例1
(1)pET-28a-Ccdc189原核蛋白表达重组质粒的构建
应用Gene Runner软件设计Ccdc189开放阅读框全长序列特异引物,其中 Ccdc189变异剪接体1基因开放阅读框的核苷酸序列如SEQ ID NO.3所示。前向引物的核苷酸序列如SEQ ID NO.1所示,具体序列如下:
5’-GGGAATTCATGATTACCCCCAGTTCCAG-3’,反向引物的核苷酸序列如SEQ ID NO.2所示,具体序列如下:
5’-5’-GGAAGCTTGCTTGGTCTTGGTCTTGCC-3’。以Ccdc189真核表达质粒为模板,利用Ccdc189特异性引物,进行PCR扩增。
PCR反应条件为:95℃,5min;95℃ 30s、60℃ 45s、72℃ 62s,共35个循环;72℃10min,4℃保存PCR扩增产物。应用12g/L琼脂糖凝胶电泳分离PCR扩增产物并切胶纯化,利用T4-DNA连接酶将纯化的PCR产物与 pGEM-T-Easy载体相连接,16℃反应过夜。将连接产物转化入大肠杆菌JM109 感受态细胞中,与IPTG(异丙基-β-D-硫代吡喃半乳糖苷)、X-gal(5-溴-4- 氯-3-吲哚-β-D-半乳糖苷)一并涂布于氨苄青霉素抗性的LB固体培养板(LB 固体培养板中氨苄青霉素的浓度为100μg/ml)上。利用蓝白斑筛选原理,挑选白色单克隆菌落,摇菌提取质粒,使用限制性内切酶EcoRI 37℃水浴酶切4小时,用12g/L琼脂糖凝胶电泳检测酶切产物。将连接成功的 pGEM-T-Easy-Ccdc189重组质粒进行测序鉴定。
使用限制性内切酶EcoRI和Hind III将测序正确的pGEM-T-Easy-Ccdc189 质粒与pET-28a(+)质粒进行酶切,应用12g/L琼脂糖凝胶电泳分离并回收酶切产物,将回收的Ccdc189目的片段、pET-28a表达载体相连接,转化到大肠杆菌JM109感受态细胞中,涂布于卡那霉素抗性的LB固体平板上培养(LB固体平板中卡那霉素的浓度为100μg/ml)。挑选单克隆菌落,经限制性内切酶 EcoRI和Hind III双酶切鉴定,确定pET-28a-Ccdc189重组质粒构建成功,结果见图1。
(2)Ccdc189原核蛋白的诱导表达
将pET-28a-Ccdc189重组质粒转化E.coli BL21感受态细胞中,在卡那霉素抗性的LB固体培养基上培养。挑选阳性单克隆菌落于5mL卡那霉素抗性的LB 液(LB液中卡那霉素的浓度为100μg/ml)中,37℃,225r/min,培养过夜。次日,将过夜菌液加入500mL卡那霉素抗性的LB液中,37℃,225r/min,培养3h。取1mL菌液于试管中,作为阴性对照,在其余菌液中加入诱导剂IPTG 至终浓度为1mmol/L,37℃,225r/min,诱导4h。SDS-PAGE检测阴性对照、诱导后菌液,判断原核蛋白是否表达,结果见图2。
将诱导后表达蛋白的大肠杆菌离心收集菌体,加入60mL重悬缓冲液 (50mmol/LTris-HCl、1mmol/L EDTA、250mL/L蔗糖、10mmol/L DTT,pH=8.0) 充分重悬。重悬后加入4.2mL细菌裂解液(1mg/mL溶菌酶、5mmol/L MgCl
上述描述中,DNaseI为脱氧核糖核酸酶I(购自赛默飞)、TritonX-100为聚乙二醇辛基苯基醚(购自赛默飞)。
(3)pET-28a-Ccdc189原核蛋白的纯化
由于重组蛋白带有His标签,因此先使用能够吸附His标签的亲和层析柱纯化含有Ccdc189重组蛋白的上清液,再利用咪唑与His标签具有相似的性质从而可以竞争性洗脱目的蛋白的原理,分别用不同浓度的咪唑溶液进行洗柱。将上清液过Ni-NTA亲和层析柱,然后用25、50、100及200mmol/L咪唑分别洗脱层析柱,SDS-PAGE结果显示50mmol/L、100mmol/L咪唑洗脱液中原核蛋白洗脱效率最高,纯度较高(见图3)。将纯化效率最高的洗脱液装入处理好的透析袋,分别置入7M、6M、5M、4M、3M及2M尿素溶液中进行梯度复性,收集复性后的蛋白溶液,采用BCA法测复性后蛋白溶液的浓度为2.77mg/ml。 Ccdc189编码蛋白的氨基酸序列如SEQ ID NO.4所示。
(4)Ccdc189多克隆抗体的制备
以复性后的Ccdc189原核蛋白作为免疫原,免疫5只成年雄性新西兰大白兔。初次免疫时将复性后的Ccdc189原核蛋白与等体积福氏完全佐剂混合,冰上乳化,背部皮下多点注射免疫成年新西兰大白兔(200μg/只)。此后,将福氏完全佐剂替换为不完全佐剂与已复性的Ccdc189原核蛋白等体积混合,每隔 2周免疫1次,共5次。末次免疫后,取兔耳缘静脉血血清检测抗体效价。
(5)ELISA检测Ccdc189多克隆抗体效价
将纯化后的Ccdc189原核蛋白稀释至8ug/mL,96孔酶标板每孔加入100μL 进行包被,放入湿盒中,4℃过夜。次日,去除板内液体,用PBS-T溶液(普诺赛)洗涤5min×3次。然后加入含1g/L脱脂奶粉的PBS-T溶液,37℃封闭1h。去除封闭液,扣干,用PBS-T溶液洗涤5min×3次。加入稀释比例为1:10~1: 1000000的抗Ccdc189血清和正常兔血清,37℃孵育90min。用PBS-T溶液洗涤5min×3次。每孔加入100μL HRP标记的山羊抗兔IgG(1:2000稀释),37℃孵育40min。去除二抗,用PBS-T溶液洗涤5min×3次。每孔加入100μL TMB 显色液,室温避光反应10min,每孔加入100μL终止液终止反应。酶标仪测定波长490nm处各孔吸光度值,结果见图4,5只免疫后的新西兰大白兔血清均有免疫性,且抗体效价均能达到1:10
(6)应用Ccdc189多克隆抗体进行睾丸组织Western Blot分析
取成年的Ccdc189基因敲除纯合子(HOM)小鼠及野生型(WT)小鼠的睾丸总蛋白分别与上样蛋白缓冲液混匀后,沸水浴10min,冰浴5min后,进行 SDS-PAGE。电泳后,将蛋白电转至PVDF膜上,用含50g/L脱脂奶粉的TBS 溶液(MCE中国)室温封闭2h。加Ccdc189抗血清(1∶2000稀释),4℃孵育过夜。次日,TBST溶液(赛默飞)洗膜10min×3次,加入HRP标记的山羊抗兔IgG(1:2000稀释),室温孵育1h;TBST洗膜10min×3次。用ECL显影试剂盒(北京奥德佳)在Tanon-2500化学发光成像系统中显影,结果见图5,所制备的兔抗小鼠Ccdc189多克隆抗体可特异性识别野生型小鼠睾丸中内源性 Ccdc189蛋白,Ccdc189基因敲除小鼠中未检测到Ccdc189蛋白。
(7)应用Ccdc189多克隆抗体进行睾丸组织及精子细胞免疫荧光分析
取人睾丸组织及成年小鼠睾丸组织石蜡切片,小鼠精子细胞涂片进行常规脱蜡水化,置于封闭液中(配方:63mL甲醇、7mL双氧水),37℃水浴10min,去除内源性过氧化物酶。PBS溶液(赛默飞)洗涤5min×3次,置于枸橼酸盐缓冲液(pH=6.0,赛默飞)中,微波炉高火2.5min,低火7.5min进行修复,室温自然冷却。PBS溶液洗涤5min×3次,10g/L BSA(赛默飞)室温封闭1h。加兔抗Ccdc189多克隆抗体(1:250),4℃孵育过夜。次日,切片置于37℃温箱复性30min。PBS溶液洗涤5min×3次。加入AlexaFluor555标记的驴抗兔IgG(1∶ 1000稀释),室温避光孵育1h。PBS溶液洗涤5min×3次。DAPI(4',6-二脒基-2- 苯基吲哚,赛默飞)室温避光染核10min,PBS溶液洗涤5min×3次。甘油封片,正置荧光显微镜下观察拍照,结果见图6,Ccdc189蛋白位于人及野生型小鼠睾丸生精小管各级生精细胞中,在Ccdc189基因敲除小鼠中未观察到荧光信号,同时在以正常兔血清为阴性对照组中也未观察到阳性信号,提示我们制备的兔抗小鼠Ccdc189多克隆抗体可以特异性识别睾丸组织中的内源性Ccdc189蛋白。
本发明通过构建pET28a-Ccdc189原核蛋白表达重组质粒,在大肠杆菌中诱导表达带有His标签的小鼠Ccdc189原核蛋白,利用Ni-NTA亲和层析柱纯化 Ccdc189原核蛋白并进行复性浓缩后免疫新西兰大白兔,收集免疫兔血清检测抗体效价并进行应用分析,同等条件下,我们公开的这种抗体制备方法优于应用小鼠Ccdc189羧基端多肽作为免疫原产生的多克隆抗体。
以上所述,仅为本发明中的具体实施方式,但本发明的保护范围并不局限于此,任何熟悉该技术的人在本发明所揭露的技术范围内,可理解想到的变换或替换,都应涵盖在本发明的包含范围之内,因此,本发明的保护范围应该以权利要求书的保护范围为准。
- 一种兔抗银环蛇毒多克隆抗体及其制备方法和应用
- 一种兔抗卵转铁蛋白过敏原多克隆抗体的制备方法及其免疫分析方法