高效合成ω-氨基脂肪酸和α,ω-二胺的方法
文献发布时间:2024-04-18 20:01:55
技术领域
本发明属于生物工程技术领域,更具体地,本发明涉及高效合成ω-氨基脂肪酸和α,ω-二胺的方法。
背景技术
ω-氨基脂肪酸是尼龙的重要单体,6-氨基己酸(6-ACA)是尼龙6的单体。目前,6-ACA的工业化生产主要依赖于以苯(通过环己烷)和己内酰胺为原料的化学催化。同样,α,ω-二胺作为尼龙单体,应用广泛,如己二胺作为尼龙66的两个单体之一。目前工业上生产己二胺主要是杜邦公司开发的以丁二烯为起始原料的能量密集型多阶段化学反应,虽然规模大,但是该过程使用了剧毒氰化氢。为了响应“绿色生物制造”号召,近些年来越来越多的专家尝试利用可再生的二元酸为底物来合成ω-氨基脂肪酸和α,ω-二胺。
2020年,加拿大Yakunin团队首次报道利用羧酸还原酶和转氨酶催化羧酸还原胺化由己二酸合成6-氨基己酸,随后继续催化6-氨基己酸的羧酸还原胺化合成己二胺,主要催化剂形式为纯酶,但是最终产物6-氨基己酸的滴度只达到9.5mM,己二胺滴度也只有3mM(J.Am.Chem.Soc.,2020,142,1038-1048)。2023年,江南大学邓禹团队在大肠杆菌中构建了用于生物催化己二酸合成己二胺的重定向羧酸还原酶级联,虽然相对于初始菌株,己二胺的滴度提高567倍,但是只达到2.1mM(ACS Sustainable Chem.Eng.,2023,11(15):6011-6020)。与此同时,我们实验室团队通过虚拟筛选的方法成功筛选到多个高底物特异性且催化效率高的羧酸还原酶(CAR),随后和转氨酶级联催化己二酸合成6-氨基己酸,转化率接近90%,但是底物上载量只有5mM,己二胺的产率只有14%(Biotechnol Bioeng,2023,120,1773–1783)。为了进一步提高底物上载量和产率,亟需改造CAR使其对己二酸的催化效率进一步提高,同时降低其对产物6-氨基己酸的酶活力,以保证6-氨基己酸的高产率,随后将底物谱拓展至其它链长的α,ω-二元羧酸(C3-C12)合成ω-氨基脂肪酸。2020年,美国University of Nebraska的Levi Kramer等为提高CAR对AA的催化效率进行了一些改造工作,最优突变体的催化效率提高了17倍,但是最终产物是己二醇,并没有考虑对6-氨基己酸的催化效率及底物特异性(ACS Synth.Biol.,2020,9,1632-1637)。同时,为了高效合成α,ω-二胺,需要进一步的改造CAR提高其对ω-氨基脂肪酸的催化活力,目前只有Yakunin等通过解析来源于Mycobacterium abscessus的CAR晶体结构,使其对6-ACA的催化效率提高了9.1倍,但是最终底物上载量也只有10mM(J.Am.Chem.Soc.,2020,142,1038-1048)。
国内外也有多篇专利报道利用生物酶法催化合成多种不同的ω-氨基脂肪酸,如5-氨基戊酸(CN106011191A),6-氨基己酸等(CN113430119A,US20070254341A1,TW1627153B,US20190271014A1),但是这些路线的底物(赖氨酸,6-氨基-2-羟基己酸,α-酮庚二酸,6-羟基己酸)都是非生物基来源的,且产物单一,元件酶广谱性差。生物法合成不同链长的α,ω-二胺亦有报道,如1,4-丁二胺(CN116355824A),戊二胺(CN116376789A),己二胺(US20190271014A1,US9,580,732B2),1,8-辛二胺和1,10-癸二胺(US9,580,732B2),这些路线的底物主要是L-谷氨酸、赖氨酸、6-羟基己酸和α,ω-二醇等。2021年,重庆大学王丹等将重组质粒组合物转入大肠杆菌BL21中得到两株基因工程菌,将赖氨酸(5g/L)作为发酵底物与所述基因工程菌LRAD培养,得到2.18~6.65g/L己二胺(CN113755419A)。
也有专利报道通过设计级联路线同时合成6-氨基己酸和己二胺等尼龙单体。2015年,瑞士英威达技术有限责任公司博特斯等描述了用于从2,6-二氨基庚二酸生产2-氨基庚二酸的生物化学途径,并用于将2-氨基庚二酸转化为6-氨基己酸、己二胺等,但是具体产物滴度未加详述(CN106574283A)。德国赢创德固赛有限公司沙费尔等提出从羧酸或二羧酸或其单酯生产胺和二胺,但是具体实施例中只有氨基十一烷酸甲酯(CN104937104A)。随后,多伦多大学Yakunin团队以20mM己二酸为底物合成6-氨基己酸和己二胺,但是6-氨基己酸的最大转化效率只达到21.3±0.8%,而己二胺产率在专利中没有明确指出(CN116064690A)。
因此,本领域亟需解决多酶级联催化α,ω-二元羧酸(C3-C12)合成ω-氨基脂肪酸和α,ω-二胺体系中限速酶羧酸还原酶(CAR)对模式底物己二酸(AA)和6-氨基己酸(6-ACA)催化活性低、底物特异性差等问题。
发明内容
本发明的目的在于提供一种高效合成α,ω-二胺的方法,其包括如下步骤:(1)以α,ω-二元羧酸为底物,使用含有第一羧酸还原酶突变体的催化剂,催化合成ω-氨基脂肪酸;(2)以α,ω-二元羧酸和ω-氨基脂肪酸为底物,使用含有第二羧酸还原酶突变体的催化剂,催化合成α,ω-二胺;其中所述第一羧酸还原酶突变体对底物α,ω-二元羧酸的催化活性增强,对ω-氨基脂肪酸的催化活性降低;和/或,其中所述第二羧酸还原酶突变体对底物α,ω-二元羧酸和ω-氨基脂肪酸的催化活性均增强。本发明也提供了一种高效合成ω-氨基脂肪酸的方法。同时,本发明还揭示了一系列羧酸还原酶突变体,及其用于合成ω-氨基脂肪酸和α,ω-二胺的应用。
在本发明的第一方面,提供一种高效合成α,ω-二胺的方法,其包括如下步骤:
(1)以α,ω-二元羧酸为底物,使用含有第一羧酸还原酶突变体的催化剂,催化合成ω-氨基脂肪酸;
(2)以α,ω-二元羧酸和ω-氨基脂肪酸为底物,使用含有第二羧酸还原酶突变体的催化剂,催化合成α,ω-二胺;
其中所述第一羧酸还原酶突变体对底物α,ω-二元羧酸的催化活性增强,对ω-氨基脂肪酸的催化活性降低;和/或,
其中所述第二羧酸还原酶突变体对底物α,ω-二元羧酸和ω-氨基脂肪酸的催化活性均增强。
在一种或多种实施方式中,所述第一羧酸还原酶突变体是:
(a1)氨基酸序列对应于SEQ ID No.2,第303位、第306位、第342位、第344位、第395位、第418位、第426位中的一个或多个氨基酸发生突变的蛋白;
(b1)将(a1)蛋白的氨基酸序列经过一个或多个氨基酸残基的取代、缺失或添加而形成的,且具有(a1)蛋白功能的由(a1)衍生的蛋白,但对应于SEQ ID No.2的第303位、第306位、第342位、第344位、第395位、第418位、第426位中的一个或多个氨基酸与(a1)蛋白相应位置突变后的氨基酸相同;
(c1)与(a1)蛋白的氨基酸序列有80%以上同源性(例如85%以上、88%以上、90%以上、92%以上、95%以上、98%以上或99%以上同源性)且具有(a1)蛋白功能的由(a1)衍生的蛋白,但对应于SEQ ID No.2的第303位、第306位、第342位、第344位、第395位、第418位、第426位中的一个或多个氨基酸与(a)蛋白相应位置突变后的氨基酸相同;
(d1)(a1)蛋白的活性片段,其包含羧酸还原酶空间结构中与羧酸底物相互作用的结构,且在对应于SEQ ID No.2的第303位、第306位、第342位、第344位、第395位、第418位、第426位中的一个或多个氨基酸与(a1)蛋白相应位置突变后的氨基酸相同。
在一种或多种实施方式中,所述第一羧酸还原酶突变体选自如下的一组:
(1)第303位丙氨酸(Ala)突变为赖氨酸(Lys);
(2)第306位亮氨酸(Leu)突变为赖氨酸(Lys)或精氨酸(Arg);
(3)第342位亮氨酸(Leu)突变为赖氨酸(Lys)或精氨酸(Arg);
(4)第344位缬氨酸(Val)突变为赖氨酸(Lys);
(5)第395位甘氨酸(Gly)突变为赖氨酸(Lys);
(6)第418位甘氨酸(Gly)突变为组氨酸(His)、赖氨酸(Lys)、天冬酰胺(Asn)或丝氨酸(Ser);
(7)第426位甘氨酸(Gly)突变为组氨酸(His)。
在一种或多种实施方式中,所述第二羧酸还原酶突变体是:
(a2)氨基酸序列对应于SEQ ID No.2,第284位、第303位、第306位、第342位、第393位、第395位、第418位、第426位中的一个或多个氨基酸发生突变的蛋白;
(b2)将(a2)蛋白的氨基酸序列经过一个或多个氨基酸残基的取代、缺失或添加而形成的,且具有(a2)蛋白功能的由(a2)衍生的蛋白,但对应于SEQ ID No.2的第284位、第303位、第306位、第342位、第393位、第395位、第418位、第426位中的一个或多个氨基酸与(a2)蛋白相应位置突变后的氨基酸相同;
(c2)与(a2)蛋白的氨基酸序列有80%以上同源性(例如85%以上、88%以上、90%以上、92%以上、95%以上、98%以上或99%以上同源性)且具有(a2)蛋白功能的由(a2)衍生的蛋白,但对应于SEQ ID No.2的第284位、第303位、第306位、第342位、第393位、第395位、第418位、第426位中的一个或多个氨基酸与(a)蛋白相应位置突变后的氨基酸相同;
(d2)(a2)蛋白的活性片段,其包含羧酸还原酶空间结构中与羧酸底物相互作用的结构,且在对应于SEQ ID No.2的第284位、第303位、第306位、第342位、第393位、第395位、第418位、第426位中的一个或多个氨基酸与(a2)蛋白相应位置突变后的氨基酸相同。
在一种或多种实施方式中,所述第二羧酸还原酶突变体选自如下的一组,或为如下的两组或两组以上的组合:
(8)第284位亮氨酸(Leu)突变为天冬氨酸(Asp)、谷氨酸(Glu)、甘氨酸(Gly)、天冬酰胺(Asn)、苏氨酸(Thr)、缬氨酸(Val)或色氨酸(Trp);
(9)第303位丙氨酸(Ala)突变为谷氨酸(Glu)、甲硫氨酸(Met)或缬氨酸(Val);
(10)第306位亮氨酸(Leu)突变为甲硫氨酸(Met)或缬氨酸(Val);
(11)第342位亮氨酸(Leu)突变为天冬氨酸(Asp)、谷氨酸(Glu)、赖氨酸(Lys)、甲硫氨酸(Met)、天冬酰胺(Asn)、谷氨酰胺(Gln)、缬氨酸(Val)或色氨酸(Trp);
(12)第393位甘氨酸(Gly)突变为天冬氨酸(Asp)或谷氨酸(Glu);
(13)第395位甘氨酸(Gly)突变为谷氨酸(Glu);
(14)第418位甘氨酸(Gly)突变为天冬氨酸(Asp)或谷氨酸(Glu);
(15)第426位甘氨酸(Gly)突变为丝氨酸(Ser)。
在一种或多种实施方式中,所述第二羧酸还原酶突变体选自如下的一组:
(16)第284位亮氨酸(Leu)突变为天冬氨酸(Asp),第303位丙氨酸(Ala)突变为甲硫氨酸(Met),第306位亮氨酸(Leu)突变为异亮氨酸(Ile),第342位亮氨酸(Leu)突变为谷氨酸(Glu),和第393位甘氨酸(Gly)突变为丙氨酸(Ala);
(17)第284位亮氨酸(Leu)突变为天冬氨酸(Asp),第306位亮氨酸(Leu)突变为甲硫氨酸(Met),和第342位亮氨酸(Leu)突变为谷氨酸(Glu);
(18)第284位亮氨酸(Leu)突变为天冬氨酸(Asp),第303位丙氨酸(Ala)突变为异亮氨酸(Ile),第306位亮氨酸(Leu)突变为异亮氨酸(Ile),第342位亮氨酸(Leu)突变为谷氨酸(Glu),和第418位甘氨酸(Gly)突变为天冬氨酸(Asp);
(19)第284位亮氨酸(Leu)突变为异亮氨酸(Ile),第306位亮氨酸(Leu)突变为甲硫氨酸(Met),第342位亮氨酸(Leu)突变为谷氨酸(Glu),和第393位甘氨酸(Gly)突变为谷氨酸(Glu);
(20)第284位亮氨酸(Leu)突变为天冬氨酸(Asp),第306位亮氨酸(Leu)突变为甲硫氨酸(Met),第342位亮氨酸(Leu)突变为谷氨酸(Glu),第393位甘氨酸(Gly)突变为丙氨酸(Ala),和第418位甘氨酸(Gly)突变为谷氨酸(Glu);
(21)第284位亮氨酸(Leu)突变为天冬氨酸(Asp),第306位亮氨酸(Leu)突变为甲硫氨酸(Met),第342位亮氨酸(Leu)突变为谷氨酸(Glu),和第393位甘氨酸(Gly)突变为谷氨酸(Glu);
(22)第284位亮氨酸(Leu)突变为谷氨酸(Glu),第306位亮氨酸(Leu)突变为甲硫氨酸(Met),第342位亮氨酸(Leu)突变为谷氨酸(Glu),第393位甘氨酸(Gly)突变为丙氨酸(Ala),和第426位甘氨酸(Gly)突变为丝氨酸(Ser);
(23)第284位亮氨酸(Leu)突变为苯丙氨酸(Phe),第306位亮氨酸(Leu)突变为异亮氨酸(Ile),第342位亮氨酸(Leu)突变为天冬氨酸(Asp),和第393位甘氨酸(Gly)突变为谷氨酸(Glu);
(24)第284位亮氨酸(Leu)突变为苯丙氨酸Phe,第306位亮氨酸(Leu)突变为甲硫氨酸(Met),第342位亮氨酸(Leu)突变为天冬氨酸(Asp),和第393位甘氨酸(Gly)突变为谷氨酸(Glu);
(25)第303位丙氨酸(Ala)突变为甲硫氨酸(Met),第306位亮氨酸(Leu)突变为异亮氨酸(Ile),第342位亮氨酸(Leu)突变为天冬氨酸(Asp),和第393位甘氨酸(Gly)突变为谷氨酸(Glu);
(26)第303位丙氨酸(Ala)突变为甲硫氨酸(Met),第306位亮氨酸(Leu)突变为甲硫氨酸(Met),第342位亮氨酸(Leu)突变为天冬氨酸(Asp),和第393位甘氨酸(Gly)突变为谷氨酰胺(Gln);
(27)第284位亮氨酸(Leu)突变为天冬氨酸(Asp),第306位亮氨酸(Leu)突变为甲硫氨酸(Met),第342位亮氨酸(Leu)突变为天冬氨酸(Asp),和第393位甘氨酸(Gly)突变为谷氨酸(Glu);
(28)第306位亮氨酸(Leu)突变为异亮氨酸(Ile),第342位亮氨酸(Leu)突变为天冬氨酸(Asp),和第418位甘氨酸(Gly)突变为天冬氨酸(Asp);
(29)第284位亮氨酸(Leu)突变为谷氨酸(Glu),第303位丙氨酸(Ala)突变为异亮氨酸(Ile),第306位亮氨酸(Leu)突变为异亮氨酸(Ile),第342位亮氨酸(Leu)突变为谷氨酸(Glu),和第393位甘氨酸(Gly)突变为谷氨酸(Glu);
(30)第284位亮氨酸(Leu)突变为天冬氨酸(Asp),第303位丙氨酸(Ala)突变为甲硫氨酸(Met),第306位亮氨酸(Leu)突变为异亮氨酸(Ile),第342位亮氨酸(Leu)突变为谷氨酸(Glu),第393位甘氨酸(Gly)突变为丙氨酸(Ala),和第418位甘氨酸(Gly)突变为天冬氨酸(Asp);
(31)第284位亮氨酸(Leu)突变为谷氨酰胺(Gln),第306位亮氨酸(Leu)突变为甲硫氨酸(Met),第342位亮氨酸(Leu)突变为谷氨酸(Glu),第418位甘氨酸(Gly)突变为谷氨酸(Glu),和第426位甘氨酸(Gly)突变为丝氨酸(Ser);
(32)第284位亮氨酸(Leu)突变为天冬氨酸(Asp),第303位丙氨酸(Ala)突变为异亮氨酸(Ile),第306位亮氨酸(Leu)突变为异亮氨酸(Ile),第342位亮氨酸(Leu)突变为天冬氨酸(Asp),第393位甘氨酸(Gly)突变为谷氨酸(Glu),和第418位甘氨酸(Gly)突变为谷氨酸(Glu);
(33)第284位亮氨酸(Leu)突变为苯丙氨酸(Phe),第306位亮氨酸(Leu)突变为异亮氨酸(Ile),第342位亮氨酸(Leu)突变为谷氨酸(Glu),和第393位甘氨酸(Gly)突变为谷氨酰胺(Gln)。
在一种或多种实施方式中,所述的α,ω-二元羧酸包括:丙二酸(C3)、丁二酸(C4)、戊二酸(C5)、己二酸(C6)、庚二酸(C7)、辛二酸(C8)、壬二酸(C9)、癸二酸(C10)、十一烷二酸(C11)和十二烷二酸(C12)。
在一种或多种实施方式中,所述ω-氨基脂肪酸是3-氨基丙酸(C3)、4-氨基丁酸(C4)、5-氨基戊酸(C5)、6-氨基己酸(C6)、7-氨基庚酸(C7)、8-氨基辛酸(C8)、9-氨基壬酸(C9)、10-氨基癸酸(C10)、11-氨基十一酸(C11)、12-氨基十二酸(C12)或其衍生物。
在一种或多种实施方式中,所述α,ω-二胺是短链α,ω-二胺,中链α,ω-二胺,中长链α,ω-二胺或长链α,ω-二胺。
在一种或多种实施方式中,所述短链是指5C以下(例如3C-5C)长度的碳链,所述中链是指6C-9C长度(6C作为尼龙单体,占比最大)的碳链,所述中长链是指10C-12C长度的碳链,所述长链是指13C以上长度的碳链。
在一种或多种实施方式中,所述“以上”或“以下”是指包含本数。
在一种或多种实施方式中,所述的3C-5C、6C-9C、10C-12C,均指包含端点值。
在一种或多种实施方式中,所述α,ω-二胺包括:1,3-丙二胺(C3)、1,4-丁二胺(C4)、1,5-戊二胺(C5)、1,6-己二胺(C6)、1,7-庚二胺(C7)、1,8-辛二胺(C8)、1,9-壬二胺(C9)、1,10-癸二胺(C10)、1,11-十一烷二胺(C11)、1,12-十二烷二胺(C12)或其衍生物。
在一种或多种实施方式中,所述催化剂还包括:转氨酶、葡萄糖脱氢酶和多磷酸激酶中的一种或多种。
在一种或多种实施方式中,所述转氨酶以有机胺作为氨基供体。
在一种或多种实施方式中,所述氨基供体包括:脂肪族胺供体(例如异丙胺、丙氨酸、谷氨酸等)、芳香族氨基供体(例如苄胺等)。
在一种或多种实施方式中,所述葡萄糖脱氢酶以葡萄糖和NADP
在一种或多种实施方式中,所述葡萄糖脱氢酶为葡萄糖脱氢酶BmGDH。
在一种或多种实施方式中,所述多磷酸激酶以六偏磷酸钠为底物,催化AMP生成ATP。
在一种或多种实施方式中,所述多磷酸激酶为多磷酸激酶PPK12。
在本发明的第二方面,提供一种高效合成ω-氨基脂肪酸的方法,其包括:以α,ω-二元羧酸为底物,使用第一羧酸还原酶突变体作为催化剂,催化合成ω-氨基脂肪酸;其中所述第一羧酸还原酶突变体对底物α,ω-二元羧酸的催化活性增强,对ω-氨基脂肪酸的催化活性降低。
在一种或多种实施方式中,所述第一羧酸还原酶突变体是:
(a1)氨基酸序列对应于SEQ ID No.2,第303位、第306位、第342位、第344位、第395位、第418位、第426位中的一个或多个氨基酸发生突变的蛋白;
(b1)将(a1)蛋白的氨基酸序列经过一个或多个氨基酸残基的取代、缺失或添加而形成的,且具有(a1)蛋白功能的由(a1)衍生的蛋白,但对应于SEQ ID No.2的第303位、第306位、第342位、第344位、第395位、第418位、第426位中的一个或多个氨基酸与(a1)蛋白相应位置突变后的氨基酸相同;
(c1)与(a1)蛋白的氨基酸序列有80%以上同源性(例如85%以上、88%以上、90%以上、92%以上、95%以上、98%以上或99%以上同源性)且具有(a1)蛋白功能的由(a1)衍生的蛋白,但对应于SEQ ID No.2的第303位、第306位、第342位、第344位、第395位、第418位、第426位中的一个或多个氨基酸与(a)蛋白相应位置突变后的氨基酸相同;
(d1)(a1)蛋白的活性片段,其包含羧酸还原酶空间结构中与羧酸底物相互作用的结构,且在对应于SEQ ID No.2的第303位、第306位、第342位、第344位、第395位、第418位、第426位中的一个或多个氨基酸与(a1)蛋白相应位置突变后的氨基酸相同。
在一种或多种实施方式中,其中所述第一羧酸还原酶突变体选自如下的一组:
(1)第303位丙氨酸(Ala)突变为赖氨酸(Lys);
(2)第306位亮氨酸(Leu)突变为赖氨酸(Lys)或精氨酸(Arg);
(3)第342位亮氨酸(Leu)突变为赖氨酸(Lys)或精氨酸(Arg);
(4)第344位缬氨酸(Val)突变为赖氨酸(Lys);
(5)第395位甘氨酸(Gly)突变为赖氨酸(Lys);
(6)第418位甘氨酸(Gly)突变为组氨酸(His)、赖氨酸(Lys)、天冬酰胺(Asn)或丝氨酸(Ser);
(7)第426位甘氨酸(Gly)突变为组氨酸(His)。
在本发明的第三方面,提供一种羧酸还原酶突变体,所述突变体相对于野生型的羧酸还原酶,其对底物的催化活性改变。
在一种或多种实施方式中,当所述底物是α,ω-二元羧酸时,所述羧酸还原酶突变体对底物α,ω-二元羧酸的催化活性增强,对ω-氨基脂肪酸的催化活性降低;较佳地,所述羧酸还原酶突变体为本发明任一实施方案所定义的第一羧酸还原酶突变体。
在一种或多种实施方式中,当所述底物是α,ω-二元羧酸和ω-氨基脂肪酸时,所述羧酸还原酶突变体对底物α,ω-二元羧酸和ω-氨基脂肪酸的催化活性均增强;较佳地,所述羧酸还原酶突变体为本发明任一实施方案所定义的第二羧酸还原酶突变体。
在本发明的第四方面,提供一种分离的多核苷酸,所述的核酸是编码本发明所述的羧酸还原酶突变体,或所述的核酸编码本发明任一实施方案所定义的第一羧酸还原酶突变体或第二羧酸还原酶突变体。
在本发明的第五方面,提供一种载体,其含有本发明所述的多核苷酸;较佳地,所述载体是表达载体或表达转化体;更佳地,所述表达转化体包含表达载体。
在本发明的第六方面,提供一种遗传工程化的宿主细胞,其含有本发明所述的载体,或基因组中整合有本发明所述的多核苷酸。
在本发明的第七方面。提供选自如下的应用:
(a)本发明所述的羧酸还原酶突变体,或本发明任一实施方案所定义的第一羧酸还原酶突变体或第二羧酸还原酶突变体在催化合成α,ω-二胺中的应用;
(b)本发明所述的第一羧酸还原酶突变体,或本发明任一实施方案所定义的第一羧酸还原酶突变体在催化合成ω-氨基脂肪酸中的应用;
(c)本发明所述的羧酸还原酶突变体,或如本发明任一实施方案所定义的第一羧酸还原酶突变体或第二羧酸还原酶突变体在制备用于催化合成ω-氨基脂肪酸和/或α,ω-二胺的催化剂中的应用。
在一种或多种实施方式中,本发明所述的第一羧酸还原酶突变体,或本发明任一实施方案所定义的第一羧酸还原酶突变体可用于制备催化合成ω-氨基脂肪酸的催化剂,也可以用于制备催化合成α,ω-二胺的催化剂。
在一种或多种实施方式中,本发明所述的第二羧酸还原酶突变体,或本发明任一实施方案所定义的第二羧酸还原酶突变体可用于制备催化合成α,ω-二胺的催化剂。
本发明的其它方面由于本文的公开内容,对本领域的技术人员而言是显而易见的。
附图说明
图1、合成BOC保护的ω-氨基脂肪酸的反应方案。
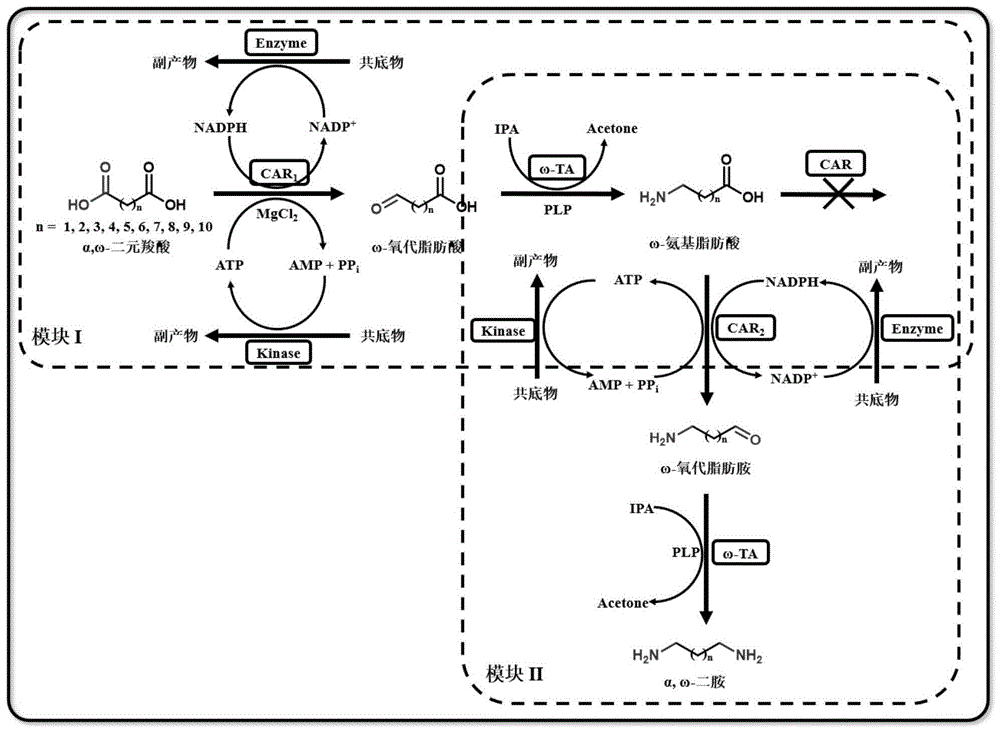
图2、多酶级联催化α,ω-二元羧酸合成不同链长ω-氨基脂肪酸和α,ω-二胺示意图。
具体实施方式
针对多酶级联催化α,ω-二元羧酸(C3-C12)合成ω-氨基脂肪酸和α,ω-二胺体系中限速酶羧酸还原酶(CAR)对模式底物己二酸(AA)和6-氨基己酸(6-ACA)催化活性低、底物特异性差等问题,本发明人经过深入的研究,提供了一种高效合成ω-氨基脂肪酸和α,ω-二胺的方法,通过定点突变羧酸还原酶,精准调控羧酸还原酶对AA和6-ACA的催化效率以高效合成ω-氨基脂肪酸和α,ω-二胺。
当高效合成ω-氨基脂肪酸时,与野生型羧酸还原酶相比,本发明提供的羧酸还原酶突变体对α,ω-二元羧酸具备更高的催化效率,同时对产物ω-氨基脂肪酸的催化效率降低,从而使得底物上载量进一步提高,ω-氨基脂肪酸的产率更高。当高效合成α,ω-二胺时,与野生型羧酸还原酶相比,本发明提供的羧酸还原酶突变体对α,ω-二元羧酸和ω-氨基脂肪酸的催化效率均提高,尤其是对ω-氨基脂肪酸的催化效率k
如本文所用,除非另外说明,所述的“羧酸还原酶突变体”、“重组羧酸还原酶突变体”可互换使用,是指对应于野生型羧酸还原酶在相应于其底物结合口袋附近发生突变后构成的多肽或催化活性发生改变的多肽,较佳地相应于其序列第284位、第303位、第306位、第342位、第344位、第393位、第395位、第418位、第426位中的一个或多个位点发生突变后构成的多肽。
如本文所用,“第一”和“第二”仅仅是为了区分在不同步骤中应用的羧酸还原酶突变体,并不代表次序或先后。
若需要表示野生型的羧酸还原酶,其可以为氨基酸序列如SEQ ID No.2的蛋白,或者也可以是该蛋白的同功能变体或活性片段。较佳地,所述的野生型羧酸还原酶来源于Mycobacteroides abscessus;但是应理解,本发明中也涵盖来源于其它的与之具有同源性且功能相同的羧酸还原酶同源物。若需要表示野生型羧酸还原酶的多核苷酸,其可以为包括编码此蛋白的多核苷酸,例如核苷酸序列如SEQ ID No.1所示的多核苷酸,或者也可以是包括附加编码和/或非编码序列的多核苷酸。
如本文所用,“分离的羧酸还原酶突变体”是指羧酸还原酶突变体基本上不含天然与其相关的其它蛋白、脂类、糖类或其它物质。本领域的技术人员能用标准的蛋白质纯化技术纯化羧酸还原酶突变体。基本上纯的蛋白在非还原聚丙烯酰胺凝胶上能产生单一的主带。
如本文所用,“底物结合口袋”是指羧酸还原酶突变体的空间结构中与底物发生相互作用(结合)的位置。
本发明的蛋白可以是重组蛋白、天然蛋白、合成蛋白,优选重组蛋白。本发明的蛋白可以是天然纯化的产物,或是化学合成的产物,或使用重组技术从原核或真核宿主(例如,细菌、酵母、高等植物、昆虫和哺乳动物细胞)中产生。
本发明还包括所述羧酸还原酶突变体的片段、衍生物和类似物。如本文所用,术语“片段”、“衍生物”和“类似物”是指基本上保持本发明的天然羧酸还原酶突变体相同的生物学功能或活性的蛋白。本发明的蛋白片段、衍生物或类似物可以是(i)有一个或多个保守或非保守性氨基酸残基(优选保守性氨基酸残基)被取代的蛋白,而这样的取代的氨基酸残基可以是也可以不是由遗传密码编码的,或(ii)在一个或多个氨基酸残基中具有取代基团的蛋白,或(iii)附加的氨基酸序列融合到此蛋白序列而形成的蛋白(如前导序列或分泌序列或用来纯化此蛋白的序列或蛋白原序列,或融合蛋白)。根据本文的定义,这些片段、衍生物和类似物属于本领域熟练技术人员公知的范围。然而,所述的羧酸还原酶突变体及其片段、衍生物和类似物的氨基酸序列中,肯定存在本发明上面所述的突变;较佳地,该突变为对应于SEQ ID NO:2的第284位、第303位、第306位、第342位、第344位、第393位、第395位、第418位、第426位中的一个或多个氨基酸的突变。
在本发明中,术语“羧酸还原酶突变体”还包括(但并不限于):若干个(通常为1-20个,更佳地1-10个,还更佳如1-8个、1-5个、1-3个或1-2个)氨基酸的缺失、插入、替换和/或取代,以及在C末端和/或N末端添加或缺失一个或数个(通常为20个以内,较佳地为10个以内,更佳地为5个以内)氨基酸。例如,在本领域中,用性能相近或相似的氨基酸进行取代时,通常不会改变蛋白质的功能。又比如,在C末端和/或N末端添加或缺失一个或数个氨基酸通常也不会改变蛋白质的功能。该术语还包括羧酸还原酶突变体的活性片段和活性衍生物。但是在这些变异形式中,肯定存在本发明上面所述的突变;较佳地,该突变为对应于SEQ IDNO:2的第284位、第303位、第306位、第342位、第344位、第393位、第395位、第418位、第426位中的一个或多个氨基酸的突变。
在本发明中,术语“羧酸还原酶突变体”还包括(但并不限于):与所述的羧酸还原酶突变体的氨基酸序列具有80%以上,较佳地85%以上,更佳地90%以上,进一步更佳地95%以上,如98%以上、99%以上序列相同性的保留其蛋白活性的衍生的蛋白。同样地,这些衍生的蛋白中,肯定存在本发明所述的突变;较佳地,该突变为对应于SEQ ID NO:2的第284位、第303位、第306位、第342位、第344位、第393位、第395位、第418位、第426位中的一个或多个氨基酸的突变。
如本文所用,“己二酸”和“AA”可互换使用,“α,ω-二元羧酸”包括“己二酸”。在本文中,通常,“α,ω-二元羧酸”指代“己二酸”。
在本文中,“ω-氨基脂肪酸”和“6-氨基脂肪酸”可互换使用,“ω-氨基脂肪酸”包括“6-氨基己酸”。在本文中,通常,“ω-氨基脂肪酸”指代“6-氨基己酸”。所述“6-氨基己酸”、“6-ACA”和“ACA”可互换使用。
如本文所用,“1,6-己二胺”和“己二胺”可互换使用,“α,ω-二胺”包括“1,6-己二胺”。在本文中,通常,“α,ω-二胺”指代“1,6-己二胺”。
如本文所用,“催化反应”、“酶联反应”和“酶促反应”可互换使用,均指在酶的催化下进行的反应。在本文中,所述“多酶联反应”、“多酶级联反应”和“多级酶联反应”可互换使用,包括至少两酶级联反应。在具体的实施例中,合成α,ω-二胺的多酶级联反应包括:使用第一羧酸还原酶突变体作为催化剂催化合成ω-氨基脂肪酸的多酶级联反应(即模块I),以及使用第二羧酸还原酶突变体作为催化剂催化合成α,ω-二胺的多酶级联反应(即模块II)。多酶级联反应的条件是本领域技术人员已知的,例如可以在常温(例如20-40℃)常压以及pH 6.0-7.5的条件下进行。
如本文所用,“催化效率”和“催化活性”可以互换使用。
高效合成ω-氨基脂肪酸的方法
本发明提供了一种高效合成ω-氨基脂肪酸的方法,包括使用第一羧酸还原酶突变体作为催化剂。由于所述第一羧酸还原酶突变体对α,ω-二元羧酸催化效率高、对ω-氨基脂肪酸催化效率低,从而使得底物上载量进一步提高,ω-氨基脂肪酸的产率更高。
如本发明所用,所述“催化效率”通常是指酶对底物的催化活性。在一些具体的实施方式中,以比活力数值的高低表示酶的催化效率高低。
在一种或多种实施方式中,所述“催化效率”提高是指相比于“野生型”或“对照”而言,催化效率提高5%、10%、15%、20%、30%或更高。
在一种或多种实施方式中,所述“催化效率”降低是指相比于“野生型”或“对照”而言,催化效率降低5%、10%、15%、20%、30%或更低。如本发明所用,“第一羧酸还原酶突变体”对应于野生型羧酸还原酶,对α,ω-二元羧酸和ω-氨基脂肪酸催化效率发生改变。例如,对应于野生型羧酸还原酶,其对α,ω-二元羧酸的催化效率提高,对ω-氨基脂肪酸催化效率降低。由于α,ω-二元羧酸为底物,ω-氨基脂肪酸为产物,也可以说,“第一羧酸还原酶突变体”的底物专一性和/或底物特异性发生变化,即对底物α,ω-二元羧酸的专一性和/或特异性增强。在一种或多种实施方式中,对应于野生型羧酸还原酶,第一羧酸还原酶突变体在相应于SEQ ID No.2所示氨基酸序列的第303位、第306位、第342位、第344位、第395位、第418位、第426位中的一个或多个氨基酸发生突变。
在一种或多种实施方式中,所述第一羧酸还原酶突变体在相应于SEQ ID No.2所示氨基酸序列的第303位丙氨酸(Ala)、第306位亮氨酸(Leu)、第342位亮氨酸(Leu)、第344位缬氨酸(Val)、第395位甘氨酸(Gly)、第418位甘氨酸(Gly)、第426位甘氨酸(Gly)中的一个或多个氨基酸发生突变。在一种或多种优选的实施方式中,对应于SEQ ID No.2所示氨基酸序列,所述第一羧酸还原酶突变体在如下的一个或多个氨基酸位点处发生突变:
(1)第303位丙氨酸(Ala)突变为赖氨酸(Lys);
(2)第306位亮氨酸(Leu)突变为赖氨酸(Lys)或精氨酸(Arg);
(3)第342位亮氨酸(Leu)突变为赖氨酸(Lys)或精氨酸(Arg);
(4)第344位缬氨酸(Val)突变为赖氨酸(Lys);
(5)第395位甘氨酸(Gly)突变为赖氨酸(Lys);
(6)第418位甘氨酸(Gly)突变为组氨酸(His)、赖氨酸(Lys)、天冬酰胺(Asn)或丝氨酸(Ser);
(7)第426位甘氨酸(Gly)突变为组氨酸(His)。
在一种或多种实施方式中,所述高效合成ω-氨基脂肪酸的方法包括:利用本发明所述的第一羧酸还原酶突变体,以α,ω-二元羧酸为底物,在提供氨基供体的条件下,通过多酶级联反应催化合成ω-氨基脂肪酸。
高效合成α,ω-二胺的方法
本发明提供了一种高效合成α,ω-二胺的方法,包括使用第二羧酸还原酶突变体作为催化剂,催化合成α,ω-二胺。由于所述第二羧酸还原酶突变体对α,ω-二元羧酸和ω-氨基脂肪酸的催化效率都高,从而高效合成α,ω-二胺。
在一种或多种实施方式中,所述高效合成α,ω-二胺的方法中,除了使用第二羧酸还原酶突变体作为催化剂,催化合成α,ω-二胺的步骤(模块II)外,还包括使用第一羧酸还原酶突变体作为催化剂高效合成ω-氨基脂肪酸的步骤(模块I)。
如本发明所用,所述“催化效率”通常是指酶对底物的催化活性。在一些具体的实施方式中,以比活力数值的高低表示酶的催化效率高低。
在一种或多种实施方式中,所述“催化效率”提高是指相比于“野生型”或“对照”而言,催化效率提高5%、10%、15%、20%、30%或更高。如本发明所用,“第二羧酸还原酶突变体”对应于野生型羧酸还原酶,对α,ω-二元羧酸和ω-氨基脂肪酸催化效率发生改变。例如,对应于野生型羧酸还原酶,其对α,ω-二元羧酸的催化效率提高,对ω-氨基脂肪酸催化效率也提高。由于在合成α,ω-二胺的过程中,α,ω-二元羧酸为底物,ω-氨基脂肪酸为中间产物,也可以说,“第二羧酸还原酶突变体”的底物和中间产物的专一性和/或特异性发生变化,即对底物α,ω-二元羧酸和中间产物ω-氨基脂肪酸的专一性和/或特异性增强。在一种或多种实施方式中,对应于野生型羧酸还原酶,第二羧酸还原酶突变体在相应于SEQ IDNo.2所示氨基酸序列的第284位、第303位、第306位、第342位、第393位、第395位、第418位、第426位中的一个或多个氨基酸发生突变。
在一种或多种实施方式中,所述第二羧酸还原酶突变体在相应于SEQ ID No.2所示氨基酸序列的第284位亮氨酸(Leu)、第303位丙氨酸(Ala)、第306位亮氨酸(Leu)、第342位亮氨酸(Leu)、第393位甘氨酸(Gly)、第395位甘氨酸(Gly)、第418位甘氨酸(Gly)、第426位甘氨酸(Gly)中的一个或多个氨基酸发生突变。
在一种或多种优选的实施方式中,对应于SEQ ID No.2所示氨基酸序列,所述第二羧酸还原酶突变体在如下的一个或多个氨基酸位点处发生突变:
(8)第284位亮氨酸(Leu)突变为天冬氨酸(Asp)、谷氨酸(Glu)、甘氨酸(Gly)、天冬酰胺(Asn)、苏氨酸(Thr)、缬氨酸(Val)或色氨酸(Trp);
(9)第303位丙氨酸(Ala)突变为谷氨酸(Glu)、甲硫氨酸(Met)或缬氨酸(Val);
(10)第306位亮氨酸(Leu)突变为甲硫氨酸(Met)或缬氨酸(Val);
(11)第342位亮氨酸(Leu)突变为天冬氨酸(Asp)、谷氨酸(Glu)、赖氨酸(Lys)、甲硫氨酸(Met)、天冬酰胺(Asn)、谷氨酰胺(Gln)、缬氨酸(Val)或色氨酸(Trp);
(12)第393位甘氨酸(Gly)突变为天冬氨酸(Asp)或谷氨酸(Glu);
(13)第395位甘氨酸(Gly)突变为谷氨酸(Glu);
(14)第418位甘氨酸(Gly)突变为天冬氨酸(Asp)或谷氨酸(Glu);(15)第426位甘氨酸(Gly)突变为丝氨酸(Ser)。
在一种或多种实施方式中,所述第二羧酸还原酶突变体在相应于SEQ ID No.2所示氨基酸序列中的多个氨基酸发生突变,所述发生突变的氨基酸选自如下的任意一组:
第284位、第303位、第306位、第342位和第393位;或
第284位、第306位和第342位;或
第284位、第303位、第306位、第342位和第418位;或
第284位、第306位、第342位和第393位;或
第284位、第306位、第342位、第393位和第418位;或
第284位、第306位;第342位和第393位;或
第284位、第306位、第342位、第393位和第426位;或
第284位、第306位、第342位和第393位;或
第284位、第306位、第342位和第393位;或
第303位、第306位、第342位和第393位;或
第303位、第306位、第342位和第393位;或
第284位、第306位、第342位和第393位;或
第306位、第342位和第418位;或
第284位、第303位、第306位、第342位和第393位;或
第284位、第303位、第306位、第342位、第393位和第418位;或
第284位、第306位、第342位、第418位和第426位;或
第284位、第303位、第306位、第342位、第393位和第418位;或第284位、第306位、第342位和第393位。
在一种或多种实施方式中,所述第二羧酸还原酶突变体在相应于SEQ ID No.2所示氨基酸序列中的多个氨基酸发生突变,所述发生突变的氨基酸选自如下的任意一组:
第284位亮氨酸(Leu)、第303位丙氨酸(Ala)、第306位亮氨酸(Leu)、第342位亮氨酸(Leu)和第393位甘氨酸(Gly);或
第284位亮氨酸(Leu)、第306位亮氨酸(Leu)和第342位亮氨酸(Leu);或
第284位亮氨酸(Leu)、第303位丙氨酸(Ala)、第306位亮氨酸(Leu)、第342位亮氨酸(Leu)和第418位甘氨酸(Gly);或
第284位亮氨酸(Leu)、第306位亮氨酸(Leu)、第342位亮氨酸(Leu)和第393位甘氨酸(Gly);或
第284位亮氨酸(Leu)、第306位亮氨酸(Leu)、第342位亮氨酸(Leu)、第393位甘氨酸(Gly)和第418位甘氨酸(Gly);或
第284位亮氨酸(Leu)、第306位亮氨酸(Leu)、第342位亮氨酸(Leu)和第393位甘氨酸(Gly);或
第284位亮氨酸(Leu)、第306位亮氨酸(Leu)、第342位亮氨酸(Leu)、第393位甘氨酸(Gly)和第426位甘氨酸(Gly);或
第284位亮氨酸(Leu)、第306位亮氨酸(Leu)、第342位亮氨酸(Leu)和第393位甘氨酸(Gly);或
第284位亮氨酸(Leu)、第306位亮氨酸(Leu)、第342位亮氨酸(Leu)和第393位甘氨酸(Gly);或
第303位丙氨酸(Ala)、第306位亮氨酸(Leu)、第342位亮氨酸(Leu)和第393位甘氨酸(Gly);或
第303位丙氨酸(Ala)、第306位亮氨酸(Leu)、第342位亮氨酸(Leu)和第393位甘氨酸(Gly);或
第284位亮氨酸(Leu)、第306位亮氨酸(Leu)、第342位亮氨酸(Leu)和第393位甘氨酸(Gly);或
第306位亮氨酸(Leu)、第342位亮氨酸(Leu)和第418位甘氨酸(Gly);或
第284位亮氨酸(Leu)、第303位丙氨酸(Ala)、第306位亮氨酸(Leu)、第342位亮氨酸(Leu)和第393位甘氨酸(Gly);或
第284位亮氨酸(Leu)、第303位丙氨酸(Ala)、第306位亮氨酸(Leu)、第342位亮氨酸(Leu)、第393位甘氨酸(Gly)和第418位甘氨酸(Gly);或
第284位亮氨酸(Leu)、第306位亮氨酸(Leu)、第342位亮氨酸(Leu)、第418位甘氨酸(Gly)和第426位甘氨酸(Gly);或
第284位亮氨酸(Leu)、第303位丙氨酸(Ala)、第306位亮氨酸(Leu)、第342位亮氨酸(Leu)、第393位甘氨酸(Gly)和第418位甘氨酸(Gly);或
第284位亮氨酸(Leu)、第306位亮氨酸(Leu)、第342位亮氨酸(Leu)和第393位甘氨酸(Gly)。
在一种或多种优选的实施方式中,对应于SEQ ID No.2所示氨基酸序列,所述第二羧酸还原酶突变体选自如下的其中一组:
(16)第284位亮氨酸(Leu)突变为天冬氨酸(Asp),第303位丙氨酸(Ala)突变为甲硫氨酸(Met),第306位亮氨酸(Leu)突变为异亮氨酸(Ile),第342位亮氨酸(Leu)突变为谷氨酸(Glu),和第393位甘氨酸(Gly)突变为丙氨酸(Ala);
(17)第284位亮氨酸(Leu)突变为天冬氨酸(Asp),第306位亮氨酸(Leu)突变为甲硫氨酸(Met),和第342位亮氨酸(Leu)突变为谷氨酸(Glu);
(18)第284位亮氨酸(Leu)突变为天冬氨酸(Asp),第303位丙氨酸(Ala)突变为异亮氨酸(Ile),第306位亮氨酸(Leu)突变为异亮氨酸(Ile),第342位亮氨酸(Leu)突变为谷氨酸(Glu),和第418位甘氨酸(Gly)突变为天冬氨酸(Asp);
(19)第284位亮氨酸(Leu)突变为异亮氨酸(Ile),第306位亮氨酸(Leu)突变为甲硫氨酸(Met),第342位亮氨酸(Leu)突变为谷氨酸(Glu),和第393位甘氨酸(Gly)突变为谷氨酸(Glu);
(20)第284位亮氨酸(Leu)突变为天冬氨酸(Asp),第306位亮氨酸(Leu)突变为甲硫氨酸(Met),第342位亮氨酸(Leu)突变为谷氨酸(Glu),第393位甘氨酸(Gly)突变为丙氨酸(Ala),和第418位甘氨酸(Gly)突变为谷氨酸(Glu);
(21)第284位亮氨酸(Leu)突变为天冬氨酸(Asp),第306位亮氨酸(Leu)突变为甲硫氨酸(Met),第342位亮氨酸(Leu)突变为谷氨酸(Glu),和第393位甘氨酸(Gly)突变为谷氨酸(Glu);
(22)第284位亮氨酸(Leu)突变为谷氨酸(Glu),第306位亮氨酸(Leu)突变为甲硫氨酸(Met),第342位亮氨酸(Leu)突变为谷氨酸(Glu),第393位甘氨酸(Gly)突变为丙氨酸(Ala),和第426位甘氨酸(Gly)突变为丝氨酸(Ser);
(23)第284位亮氨酸(Leu)突变为苯丙氨酸(Phe),第306位亮氨酸(Leu)突变为异亮氨酸(Ile),第342位亮氨酸(Leu)突变为天冬氨酸(Asp),和第393位甘氨酸(Gly)突变为谷氨酸(Glu);
(24)第284位亮氨酸(Leu)突变为苯丙氨酸Phe,第306位亮氨酸(Leu)突变为甲硫氨酸(Met),第342位亮氨酸(Leu)突变为天冬氨酸(Asp),和第393位甘氨酸(Gly)突变为谷氨酸(Glu);
(25)第303位丙氨酸(Ala)突变为甲硫氨酸(Met),第306位亮氨酸(Leu)突变为异亮氨酸(Ile),第342位亮氨酸(Leu)突变为天冬氨酸(Asp),和第393位甘氨酸(Gly)突变为谷氨酸(Glu);
(26)第303位丙氨酸(Ala)突变为甲硫氨酸(Met),第306位亮氨酸(Leu)突变为甲硫氨酸(Met),第342位亮氨酸(Leu)突变为天冬氨酸(Asp),和第393位甘氨酸(Gly)突变为谷氨酰胺(Gln);
(27)第284位亮氨酸(Leu)突变为天冬氨酸(Asp),第306位亮氨酸(Leu)突变为甲硫氨酸(Met),第342位亮氨酸(Leu)突变为天冬氨酸(Asp),和第393位甘氨酸(Gly)突变为谷氨酸(Glu);
(28)第306位亮氨酸(Leu)突变为异亮氨酸(Ile),第342位亮氨酸(Leu)突变为天冬氨酸(Asp),和第418位甘氨酸(Gly)突变为天冬氨酸(Asp);
(29)第284位亮氨酸(Leu)突变为谷氨酸(Glu),第303位丙氨酸(Ala)突变为异亮氨酸(Ile),第306位亮氨酸(Leu)突变为异亮氨酸(Ile),第342位亮氨酸(Leu)突变为谷氨酸(Glu),和第393位甘氨酸(Gly)突变为谷氨酸(Glu);
(30)第284位亮氨酸(Leu)突变为天冬氨酸(Asp),第303位丙氨酸(Ala)突变为甲硫氨酸(Met),第306位亮氨酸(Leu)突变为异亮氨酸(Ile),第342位亮氨酸(Leu)突变为谷氨酸(Glu),第393位甘氨酸(Gly)突变为丙氨酸(Ala),和第418位甘氨酸(Gly)突变为天冬氨酸(Asp);
(31)第284位亮氨酸(Leu)突变为谷氨酰胺(Gln),第306位亮氨酸(Leu)突变为甲硫氨酸(Met),第342位亮氨酸(Leu)突变为谷氨酸(Glu),第418位甘氨酸(Gly)突变为谷氨酸(Glu),和第426位甘氨酸(Gly)突变为丝氨酸(Ser);
(32)第284位亮氨酸(Leu)突变为天冬氨酸(Asp),第303位丙氨酸(Ala)突变为异亮氨酸(Ile),第306位亮氨酸(Leu)突变为异亮氨酸(Ile),第342位亮氨酸(Leu)突变为天冬氨酸(Asp),第393位甘氨酸(Gly)突变为谷氨酸(Glu),和第418位甘氨酸(Gly)突变为谷氨酸(Glu);
(33)第284位亮氨酸(Leu)突变为苯丙氨酸(Phe),第306位亮氨酸(Leu)突变为异亮氨酸(Ile),第342位亮氨酸(Leu)突变为谷氨酸(Glu),和第393位甘氨酸(Gly)突变为谷氨酰胺(Gln)。
在一种或多种实施方式中,所述高效合成α,ω-二胺的方法以α,ω-二元羧酸为底物,所述α,ω-二元羧酸包括但不限于:丙二酸(C3)、丁二酸(C4)、戊二酸(C5)、己二酸(C6)、庚二酸(C7)、辛二酸(C8)、壬二酸(C9)、癸二酸(C10)、十一烷二酸(C11)和十二烷二酸(C12)。
在一种或多种实施方式中,所述ω-氨基脂肪酸是3-氨基丙酸(C3)、4-氨基丁酸(C4)、5-氨基戊酸(C5)、6-氨基己酸(C6)、7-氨基庚酸(C7)、8-氨基辛酸(C8)、9-氨基壬酸(C9)、10-氨基癸酸(C10)、11-氨基十一酸(C11)、12-氨基十二酸(C12)或其衍生物。
在一种或多种实施方式中,所述高效合成α,ω-二胺的方法中以有机氨作为氨基供体,所述氨基供体包括:异丙胺、丙氨酸、谷氨酸等脂肪族胺供体或苄胺等芳香族氨基供体。本领域技术人员知晓,所有能够用于合成ω-氨基脂肪酸和/或α,ω-二胺的氨基供体均可应用于本发明中。
在一种或多种实施方式中,所述催化剂除了羧酸还原酶突变体以外,还包括(但不限于):转氨酶、葡萄糖脱氢酶和多磷酸激酶。
在一种或多种实施方式中,所述催化剂的催化形式为全细胞、粗细胞提取物(粗酶液)或纯酶中的任意一种。所述粗细胞提取物可以是对转入了本发明所述转化体的宿主细胞进行破碎,分离获得的含有所述羧酸还原酶突变体的粗酶液,也可以是对所述的粗酶液进行冻干得到的冻干酶粉。所述纯酶可以是对所述粗酶液进行纯化得到的纯酶。
在一些具体的实施方式中,本发明所述的羧酸还原酶突变体,与其他催化剂联合使用时,还可以提高产物浓度。例如,当使用本发明所述羧酸还原酶突变体,同时以异丙胺作为氨基供体时,由于本发明所述羧酸还原酶突变体的催化效率高,可以高效的进一步催化中间产物ω-氨基脂肪酸,有利于上一步转氨酶的平衡移动,从而获得更高浓度的α,ω-二胺。
在一种或多种实施方式中,所述葡萄糖脱氢酶以葡萄糖和NADP
在一种或多种实施方式中,所述多磷酸激酶以六偏磷酸钠为底物,催化AMP生成ATP。在一种或多种具体的实施方式中,所述多磷酸激酶为来源于Erysipelotrichaceaebacterium的多磷酸激酶PPK12(NCBI Reference Sequence:HCY06753.1),可用于ATP再生。
在一种或多种实施方式中,所述的α,ω-二胺可以是短链α,ω-二胺,中链α,ω-二胺,中长链α,ω-二胺或长链α,ω-二胺。
在一种或多种实施方式中,所述短链是指5C以下长度的碳链,例如3C-5C长度的碳链。所述中链是指6C-9C长度的碳链。所述中长链是指10C-12C长度的碳链。所述长链是指13C以上长度的碳链。
在一种或多种实施方式中,所述α,ω-二胺是1,3-丙二胺(C3)、1,4-丁二胺(C4)、1,5-戊二胺(C5)、1,6-己二胺(C6)、1,7-庚二胺(C7)、1,8-辛二胺(C8)、1,9-壬二胺(C9)、1,10-癸二胺(C10)、1,11-十一烷二胺(C11)、1,12-十二烷二胺(C12)或其衍生物。
本领域技术人员知晓,在合成不同长度的α,ω-二胺时,需要根据希望获得的α,ω-二胺产物的链的长度,选择合适长度的α,ω-二元羧酸和/或ω-氨基脂肪酸作为底物。
采用多酶级联反应合成α,ω-二胺的方法是本领域已知的,所述方法中使用的酶促反应体系、反应温度、反应时间等条件均是本领域技术人员已知的或通过常规实验可以获得的,例如可参考Biotechnol Bioeng,2023,120,1773–1783。应理解,在得知本发明的羧酸还原酶突变体后,所有能够利用本发明所述的羧酸还原酶突变体合成ω-氨基脂肪酸和/或α,ω-二胺的方法均包含在本发明的保护范围之内。
ω-氨基脂肪酸和/或α,ω-二胺的测定方法也是本领域已知的,包括但不限于:高效液相色谱法、气相色谱法、液相色谱-质谱联用技术、气相色谱-质谱联用技术。
核酸、重组表达载体、重组表达转化体
本发明还提供了编码本发明羧酸还原酶突变体或其保守性变异蛋白的多核苷酸序列。
本发明的多核苷酸可以是DNA形式或RNA形式。DNA形式包括cDNA、基因组DNA或人工合成的DNA。DNA可以是单链的或是双链的。DNA可以是编码链或非编码链。
编码所述突变体的成熟蛋白的多核苷酸包括:只编码成熟蛋白的编码序列;成熟蛋白的编码序列和各种附加编码序列;成熟蛋白的编码序列(和任选的附加编码序列)以及非编码序列。
“编码蛋白的多核苷酸”可以是包括编码此蛋白的多核苷酸,也可以是还包括附加编码和/或非编码序列的多核苷酸。
本发明也涉及包含本发明的多核苷酸的载体,以及用本发明的载体或羧酸还原酶突变体编码序列经基因工程产生的宿主细胞,以及经重组技术产生本发明所述蛋白的方法。
通过常规的重组DNA技术,可利用本发明的多核苷酸序列来表达或生产重组的羧酸还原酶突变体。一般来说有以下步骤:
(1).用本发明的编码羧酸还原酶突变体的多核苷酸(或变异体),或用含有该多核苷酸的重组表达载体转化或转导合适的宿主细胞;
(2).在合适的培养基中培养的宿主细胞;
(3).从培养基或细胞中分离、纯化蛋白质。
本发明中,羧酸还原酶突变体多核苷酸序列可插入到重组表达载体中。术语“重组表达载体”指本领域熟知的细菌质粒、噬菌体、酵母质粒、植物细胞病毒、哺乳动物细胞病毒或其他载体。总之,只要能在宿主体内复制和稳定,任何质粒和载体都可以用。表达载体的一个重要特征是通常含有复制起点、启动子、标记基因和翻译控制元件。
在一个或多个实施方案中,可以将除了羧酸还原酶突变体之外的其他多核苷酸序列(例如编码其他催化剂的多核苷酸序列)也插入到重组表达载体中。其他多核苷酸序列可以与所述羧酸还原酶突变体多核苷酸序列插入在同一个重组表达载体中,也可以插入在不同的重组表达载体中。
因此,本发明也提供了一种重组表达转化体,其包括本发明所述的插入了羧酸还原酶突变体多核苷酸序列的重组表达载体。所述重组表达转化体可以通过本领域常规方法,将本发明所述的两种或两种以上的重组表达载体(例如插入羧酸还原酶突变体多核苷酸序列的重组表达载体与插入其他催化剂多核苷酸序列的重组表达载体)同时转化导入宿主细胞来制备重组表达转化体。
在一个或多个具体的实施方案中,通过本领域常规方法,可以将本发明所述的羧酸还原酶突变体的编码核酸序列连接于pET30b质粒,所述的转氨酶的编码核酸序列连接于pETDuet-1质粒(例如第二个多克隆位点MCS II上),所述的葡萄糖脱氢酶的编码核酸序列连接于pET-28a质粒,所述的多磷酸激酶的编码核酸序列连接于pET-28a质粒。在优选的实施方案中,为了使羧酸还原酶突变体成功表达,并表现出催化活性,可以将来源于Bacilussubtilis的磷酸泛酰乙炔基修饰酶Bssfp(NCBI Reference Sequence:WP_003234549.1)的编码核酸序列连接于pCDFDuet质粒(例如第一个多克隆位点MCS I上)。
本领域的技术人员熟知的方法能用于构建含羧酸还原酶突变体编码DNA序列和合适的转录/翻译控制信号的表达载体。这些方法包括体外重组DNA技术、DNA合成技术、体内重组技术等。所述的DNA序列可有效连接到表达载体中的适当启动子上,以指导mRNA合成。表达载体还包括翻译起始用的核糖体结合位点和转录终止子。表达载体优选地包含一个或多个选择性标记基因,以提供用于选择转化的宿主细胞的表型性状。
包含上述的适当DNA序列以及适当启动子或者控制序列的载体,可以用于转化适当的宿主细胞,以使其能够表达蛋白质。
本发明中,所述的宿主细胞可以是原核细胞,如细菌细胞;或是低等真核细胞,如酵母细胞;或是高等真核细胞,如植物细胞。宿主细胞代表性例子有:大肠杆菌(E.coli)、枯草杆菌(Bacillus subtilis)、谷氨酸棒杆菌(Corynebacterium glutamicum)、酿酒酵母(Saccharomyces cerevisiae)、链霉菌、农杆菌等。在本发明的具体实施例中,以大肠杆菌E.coli BL21(DE3)作为宿主细胞。
本领域一般技术人员都清楚如何选择适当的载体、启动子、增强子和宿主细胞。
与现有技术相比,本发明具有优异的技术效果:
1、与其他合成ω-氨基脂肪酸和α,ω-二胺的酶催化剂相比,本发明提供的羧酸还原酶突变体具有催化活性高、底物特异性强和催化底物范围广等优点,底物上载量和时空产率均达到目前报道生物法合成最高水平,为生物法工业合成尼龙单体提供了新的借鉴和思路。
2、由于羧酸还原酶催化过程需要昂贵的辅因子NADPH和ATP,随着底物上载量的提高和反应体系的扩大,所需辅因子的成本也进一步提高。本发明提供了一种来源于Bacillus megaterium IWG3的葡萄糖脱氢酶BmGDH(NCBI Reference Sequence:1GCO_A)用于NADPH再生,以及一种来源于Erysipelotrichaceae bacterium的多磷酸激酶PPK12(NCBIReference Sequence:HCY06753.1)用于ATP再生。这两种酶能够高效再生辅因子NADPH和ATP,有利于节约羧酸还原酶催化过程中辅因子的成本,使工业化扩大生产成为可能。
下面结合具体实施例,进一步阐述本发明。应理解,这些实施例仅用于说明本发明而不用于限制本发明的范围。下列实施例中未注明具体条件的实验方法,通常按照常规条件,或按照制造厂商所建议的条件。
材料和方法
1、多酶级联催化α,ω-二元羧酸合成ω-氨基脂肪酸和α,ω-二胺
多酶级联催化α,ω-二元羧酸合成中不同链长ω-氨基脂肪酸和α,ω-二胺示意图如图2所示。
酶促反应在pH 6.5的Hepes-Na缓冲液中,30℃条件下进行,合成ω-氨基脂肪酸模块1反应体系包括终浓度为100mM的α,ω-二元羧酸(C3-C12)、1mM NADP(H)、10mM ATP、120mM MgCl
合成α,ω-二胺的模块1和模块2反应体系包括终浓度为100mM的α,ω-二元羧酸(C3-C12)、2mM NADP(H)、10mM ATP、240mM MgCl
反应在震荡反应器(1000rpm)的EP管中进行,或在利用pH Stat流加2M NaOH的反应瓶(8mL或200mL)中反应,用于控制反应液pH值恒定,反应时间以底物完全转化或产物浓度不再继续上升为准。反应过程中取样100μL反应液,并加5μL 6M HCl终止反应。
2、利用高效液相色谱法(HPLC)检测产物ω-氨基脂肪酸
样品处理方法如下:取20μL样品加入20μL乙腈,混匀置于-20℃冰箱冷冻半个小时以上,随后10000rpm离心1min,取20μL上清,加入20μL 28mM Marfey和100μL DMSO,混匀后,40℃静置衍生化1h,最后加入40μL 1M HCl猝灭,混匀后用有机尼龙膜过滤。
具体分析条件如下:
色谱柱为伊利特C18,流动相为甲醇:水(0.1%三氟乙酸)=50:50(v/v),流速为1mL/min;柱温为35℃,紫外检测波长为340nm。
3、利用高效液相色谱法(HPLC)检测产物α,ω-二胺
样品处理方法如下:取42μL样品加入250μL丹磺酰氯(5mg/mL)和208μL Hepes-Na缓冲液(0.1M,pH 6.5),混匀静置10min,最后加入500μL甲醇,混匀后用有机尼龙膜过滤。
具体分析条件如下:
色谱柱为伊利特C18,流动相为甲醇:水(0.1%三氟乙酸)=83:17(v/v),流速为1mL/min;柱温为35℃,紫外检测波长为254nm。
4、利用气相色谱(GC)检测底物α,ω-二元羧酸
取30μL样品加入300μL乙酸乙酯(含内标),充分震荡5-10min,随后12000rpm离心1min,取150μL有机相,加入无水硫酸钠震荡干燥1h以上,12000rpm离心1min,取50μL有机相加入内存管,加入10μL三甲基硅烷化重氮甲烷和50μL甲醇-乙醚(v/v=1:1)。
具体分析条件如下:
色谱柱为SH-Rxi-5Sil MS column(30m×0.25mm×0.25μm),使用氮气作为载气,流速为3mL/min。进样器和检测器的温度分别为250℃和280℃,具体GC检测程序为85℃保持3分钟,以5℃/分钟速率升温至100℃,再以10℃/分钟速率升温到160℃,最后以20℃/分钟的速率升温到280℃,于280℃保持2分钟。
5、ω-氨基脂肪酸的制备方法
采用1000mL的三口烧瓶作为反应器,按照上述的反应体系进行200mL规模反应。反应结束后,反应液于12000rpm离心40min去除细胞,加入6M HCl溶液将反应液酸化至pH3.0,通过离心再一次去除沉淀的蛋白质。将反应液进行浓缩,减少反应体积并提高产物浓度,用乙酸乙酯(3×20mL)萃取,去除未反应的底物或中间产物等,剩余的水相即主要包含产物以及缓冲液中的盐。小心添加6M NaOH中和水相后加入等体积的叔丁醇、1.1equiv.NaOH以及2.0equiv.二碳酸二叔丁酯(Boc-酸酐),将反应混合物在室温中搅拌16h(反应式见图1)。反应结束后,在冰水浴中缓慢添加2M HCl中和,再用乙酸乙酯(1×50mL+2×20mL)萃取,合并乙酸乙酯相,用饱和NaCl溶液洗涤一下,无水硫酸钠进行干燥后,通过旋蒸仪进行真空浓缩,获得产物粗品。通过硅胶柱层析进一步过柱纯化,流动相采用乙酸乙酯:石油醚:乙酸=75:200:1,收集较纯的流动相,通过旋蒸仪进行真空浓缩,获得较纯的产物,使用GC-MS或H
实施例1:羧酸还原酶MabCAR3突变体的制备
本实施例首先通过基因挖掘获得一种来源于Mycobacteroides abscessus的羧酸还原酶(NCBI Reference Sequence:WP_052537360.1)编码基因,含有该基因的重组表达载体pET30b和重组表达转化体E.coli BL21(DE3)。对如SEQ ID No.2所示氨基酸序列中的第284位Leu、第303位Ala、第306位Leu、第342位Leu、第344位Val、第393位Gly、第395位Gly、第418位Gly、第426位Gly进行定点突变或多点组合突变。定点突变采用IISite-DirectedMutegenesis Kit(Stratagenem Catalog#200502)所述方案执行操作。首先设计含有突变点的突变引物,本发明所需含有突变点的突变引物为本领域常用的引物,只需通过一般基因工程技术将SEQ ID No.2中的氨基酸残基突变为本发明所需氨基酸残基即可,其核酸具体的突变密码不受限定。所示序列中第284位、第303位、第306位、第342位、第344位、第393位、第395位、第418位、第426位引入突变均为定点突变引物。
PCR反应体系(10μL):模板20~50ng,一对突变引物各0.5μL(10μM),5μLPrimeStarmix,灭菌双蒸水补足体系至10μL。其中所述的模板为羧酸还原酶(MabCAR3,WP_198151610.1,SEQ ID No.2)。
PCR反应程序:(1)95℃变性3min;(2)98℃变性10s;(3)55℃退火5s;(4)72℃延伸9min;步骤(2)~(4)共延伸25个循环,最后72℃延伸10min,4℃保藏产物。
对于多点组合突变,为了进一步提高突变效率,设计一段包含多个组合突变位点的短基因片段,通过一步克隆的方式替换野生型羧酸还原酶对应的基因片段。
得到的PCR产物经限制性内切酶Dpn I于37℃消化0.5-2h后转化至大肠杆菌E.coli BL21(DE3)感受态细胞,并均匀涂布于含有50μg/mL卡那霉素的LB培养基(蛋白胨:10g/L,酵母提取物:5g/L,氯化钠:10g/L,琼脂粉:20g/L)琼脂平板。37℃培养10-16h后,挑选单克隆,即得到含有突变体表达质粒的E.coli BL21(DE3)菌株,送上海擎科生物科技有限公司进行测序分析。测序结果用SnapGene软件与野生型羧酸还原酶基因序列进行比对,确认突变前后基因序列及相应的氨基酸序列的差异。
本发明多酶级联反应中所需的其它生物酶催化剂,如转氨酶、葡萄糖脱氢酶和多磷酸激酶等的基因序列基于大肠杆菌表达系统对密码子优化后,由上海擎科生物科技有限公司进行合成,并导入对应的载体质粒,最后转化至大肠杆菌E.coli BL21(DE3)中。
实施例2:羧酸还原酶MabCAR3突变体基因和磷酸泛酰巯基乙胺基转移酶sfp基因共表达
用Qiagen小量质粒抽提试剂盒从包含突变体质粒的E.coli BL21(DE3)菌株中提取质粒,同时从含有sfp的转化体中大批量提取表达sfp的质粒pCDFDuet。所得双质粒同时化学转化至E.coli BL21(DE3)感受态细胞中,并均匀涂布于含有50μg/mL卡那霉素和链霉素双抗性的LB培养基琼脂平板(蛋白胨:10g/L,酵母提取物:5g/L,氯化钠:10g/L,琼脂粉:20g/L),37℃培养12-18h,挑取单克隆体,即得到表达不同羧酸还原酶突变体的表达菌株,该菌株同时共表达sfp,其作用是对羧酸还原酶的末端进行修饰以保证其催化效率。
实施例3:MabCAR3及其突变体、转氨酶、葡萄糖脱氢酶/甲酸脱氢酶和多磷酸激酶生物酶催化剂的制备
粗酶的制备:将实施例1和实施例2中所得的单克隆菌落挑至4mL LB培养基(含链霉素和卡那霉素)于37℃、180rpm摇床振荡培养12-16h,随后按1%(v/v)的接种量接入100mL LB培养基(含链霉素,卡那霉素或氨苄霉素)于37℃、180rpm摇床振荡培养。当培养液的OD达到0.6-0.8时,加入终浓度为0.2mM的异丙基-β-D-硫代半乳糖苷(IPTG)作为诱导剂在16℃诱导16~24h。将培养液10000rpm离心3min,用生理盐水洗涤沉淀两次,获得重组表达转化体细胞。再将收获的重组细胞悬浮于10mL的Buffer A缓冲液中,超声破碎,离心收集上清液,即可获得所述重组羧酸还原酶突变体、转氨酶、葡萄糖脱氢酶和多磷酸激酶粗酶液,所获得的粗酶液也可以进一步利用100kDa或10kDa超滤管浓缩10-20倍,随后复溶置于-80℃冰箱冷冻保藏。
纯酶的制备:获得的重组羧酸还原酶突变体、转氨酶、葡萄糖脱氢酶和多磷酸激酶粗酶液进一步纯化。以下为蛋白纯化缓冲液配方:Buffer A:25mM Hepes-Na,pH 7.5,500mMNaCl,20mM咪唑;Buffer B:25mM Hepes-Na,pH 7.5,500mM NaCl,500mM咪唑;Buffer C:25mM Hepes-Na,pH 7.5,150mM NaCl,1mM DTT,5%甘油。将破碎获得的羧酸还原酶的粗酶液上样至用A液平衡后的镍柱上,上样完毕后,用10% B液洗脱柱内的杂蛋白,随后用50%B液洗脱目标蛋白,利用SDS-PAGE验证是否含有目标蛋白,然后收集含有目标蛋白的洗脱液,利用超滤管(10kDa或30kDa)超滤浓缩,然后用C液置换两次,以除去蛋白溶液内的咪唑,液氮速冻后于-80℃保存备用。
冻干酶粉的制备:离心收集上述的湿菌体之后,再用磷酸钠缓冲液(100mM,pH7.5)重悬菌体(150-200g/L),随后过筛,并通过高压匀浆机(700-800MPa)进行细胞破碎三次,破碎液在4℃,8000rpm条件下离心40min收集上清,于-80℃冷冻过夜。采用冷冻干燥机对冻好的酶液进行冻干,收集获得的冻干酶粉,置于4℃密封保存。
实施例4:MabCAR3及其突变体对模式底物己二酸和6-氨基己酸的比活力测定
由于MabCAR3催化模式底物己二酸的反应过程中需要消耗辅因子NADPH,而NADPH在340nm处有较高的吸收值,对应的产物NADP
MabCAR3及其突变体对己二酸和6-氨基己酸的比活力测定体系为:总反应体系为1mL,所用缓冲液为100mM的Hepes-Na缓冲液(pH 7.5)中,在30℃条件下进行反应,反应体系中包括终浓度为10mM的己二酸或6-氨基己酸、0.15mM NADPH、1mM ATP和10mM MgCl
首先我们基于Rosetta能量评分和NAC频率对一些关键氨基酸位点进行定点饱和突变,共计约8000多个突变体进行虚拟筛选,随后选择小部分突变体用于湿实验验证,再选择实验验证后的优势单点突变体进行组合突变的虚拟筛选计算,最后选择虚拟筛选后的优势多点组合突变体进行湿实验验证。
当合成6-氨基己酸时,我们最理想的羧酸还原酶突变体是对底物己二酸比活力提高,同时对产物6-氨基己酸的比活力降低甚至没有。结果表明,野生型的MabCAR3纯酶对底物己二酸比活力为0.20U/mg,同时,也获得了一系列比活力提高的单点或多点优势突变体。这些突变体的突变序列以及这些突变体对己二酸的催化活性变化情况列于表1中。在表1的列表中,序列标号分别指表1后面相对应的一系列序列;突变体比活力提高倍数中,一个加号“+”表示突变体蛋白质对己二酸的比活力比由SEQ ID No.2所示氨基酸序列组成的蛋白质提高了0.1-1.5倍;两个加号“++”表示突变体蛋白质对己二酸的比活力比由SEQ ID No.2所示氨基酸序列组成的蛋白质提高了1.6-3.0倍;三个加号“+++”表示突变体蛋白质对己二酸的比活力比由SEQ ID No.2所示氨基酸序列组成的蛋白质提高了3.1-5.0倍。四个加号“++++”表示突变体蛋白质对己二酸的比活力比由SEQ ID No.2所示氨基酸序列组成的蛋白质提高了5.0倍以上。
野生型的MabCAR3纯酶对产物6-氨基己酸比活力为0.017U/mg,同时,也获得了一系列比活力降低的单点优势突变体。这些突变体的突变序列以及这些突变体对6-氨基己酸的催化活性变化情况列于表1中。在表1的列表中,序列标号分别指表1后面相对应的一系列序列;突变体比活力降低倍数中,一个加号“+”表示突变体蛋白质对6-氨基己酸的比活力比由SEQ ID No.2所示氨基酸序列组成的蛋白质降低为原来的40%以上;两个加号“++”表示突变体蛋白质对6-氨基己酸的比活力比由SEQ ID No.2所示氨基酸序列组成的蛋白质降低为原来的20%-40%;三个加号“+++”表示突变体蛋白质对6-氨基己酸的比活力比由SEQ IDNo.2所示氨基酸序列组成的蛋白质降低为原来的10%-20%。四个加号“++++”表示突变体蛋白质对6-氨基己酸的比活力比由SEQ ID No.2所示氨基酸序列组成的蛋白质降低为原来的10%以下。
表1MaCAR3及其突变体对底物己二酸(AA)比活力提高和产物6-氨基己酸(6-ACA)的比活力降低比例
为了进一步高效合成己二胺,我们需要筛选对6-氨基己酸催化效率提高的优势突变体,首先尝试测定虚拟筛选后的单点突变体比活力。结果表明,野生型的MabCAR3纯酶对6-氨基己酸比活力为0.017U/mg。这些突变体的突变序列以及这些突变体对6-氨基己酸的催化活性提高倍数情况列于表2中。在表2的列表中,序列标号分别指表2后面相对应的一系列序列;突变体比活力提高倍数中,一个加号“+”表示突变体蛋白质对6-氨基己酸的比活力比由SEQ ID No.2所示氨基酸序列组成的蛋白质提高了0.1-1.5倍;两个加号“++”表示突变体蛋白质对6-氨基己酸的比活力比由SEQ ID No.2所示氨基酸序列组成的蛋白质提高了1.6-3.0倍;三个加号“+++”表示突变体蛋白质对6-氨基己酸的比活力比由SEQ ID No.2所示氨基酸序列组成的蛋白质提高了3.1-5.0倍。四个加号“++++”表示突变体蛋白质对6-氨基己酸的比活力比由SEQ ID No.2所示氨基酸序列组成的蛋白质提高了5.0倍以上。
表2MaCAR3及其突变体对6-氨基己酸(6-ACA)的比活力提高倍数
/>
单点突变取得了较好的结果,成功获得对6-ACA比活力有明显提高的突变体,所以继续对6个优势突变点L284/L306/L342/G393/G418/A303进行组合计算,累计8000多个突变体,最后选择如表3所示的18个组合突变体测定其对己二酸和6-氨基己酸的比活力,表3中突变体提高倍数表示方法如表1所述,与表1不同之处在于表3用“+”表示对6-ACA活力提高倍数,而不是降低倍数。如表3所示,多个多点组合突变体对己二酸和6-氨基己酸的比活力均有明显的提高,能同时提高5倍以上,对6-氨基己酸最高更是提高了65倍以上,组合突变的优势突变体可用于后续实施例中高效合成α,ω-二胺。
表3 MaCAR3及其突变体对底物己二酸(AA)和产物6-氨基己酸(6-ACA)的比活力提高倍数
实施例6:野生型羧酸还原酶MabCAR3
本发明不局限于α,ω-二元羧酸到ω-氨基脂肪酸和α,ω-二胺的转化,也同样适用于其他类似α,ω-二元羧酸到ω-氨基脂肪酸和α,ω-二胺的转化。
本实施例利用葡萄糖脱氢酶GDH再生NADPH,多磷酸激酶PPK再生ATP,CAR和TA级联催化己二酸合成6-氨基己酸,反应体系8mL,在pH 6.5的Hepes-Na缓冲液中,30℃(通过水循环控温)、800rpm条件下进行,由于级联反应过程产生大量的酸,所以通过pH Stat流加2MNaOH控制反应液的pH恒定。反应体系中包括终浓度为100mM AA、1mM NADP(H)、10mM ATP、120mM MgCl
通过HPLC检测产物6-氨基己酸的生成量,GC检测底物己二酸的消耗量和可能副产物6-羟基己酸和己二醇等,6-氨基己酸的产率达到24.8%(滴度:24.8mM),未检测到任何副产物6-羟基己酸和己二醇等。同时,检测到了少量的己二胺,说明野生型羧酸还原酶MabCAR3
实施例7:羧酸还原酶突变体MabCAR3
本实施例利用葡萄糖脱氢酶GDH再生NADPH,多磷酸激酶PPK再生ATP,CAR和TA级联催化己二酸合成6-氨基己酸,反应体系8mL,在pH 6.5的Hepes-Na缓冲液中,30℃(通过水循环控温)、800rpm条件下进行,由于级联反应过程产生大量的酸,所以通过pH Stat流加2MNaOH控制反应液的pH恒定。反应体系中包括终浓度为100mM AA、1mM NADP(H)、10mM ATP、120mM MgCl
通过HPLC检测产物6-氨基己酸的生成量,GC检测底物己二酸的消耗量和可能副产物6-羟基己酸和己二醇等,6-氨基己酸的产率达到65.1%(滴度:65.1mM),未检测到任何副产物6-羟基己酸和己二醇等。同时,未检测到己二胺,表明羧酸还原酶突变体MabCAR3
实施例8:羧酸还原酶突变体MabCAR3
本实施例利用甲酸脱氢酶FDH再生NADPH,多磷酸激酶PPK再生ATP,CAR和TA级联催化己二酸合成6-氨基己酸,反应体系8mL,在pH 6.5的Hepes-Na缓冲液中,30℃(通过水循环控温)、800rpm条件下进行,由于级联反应过程产生大量的酸,所以通过pH Stat流加2MNaOH控制反应液的pH恒定。反应体系中包括终浓度为100mM AA、1mM NADP(H)、10mM ATP、120mM MgCl
通过HPLC检测产物6-氨基己酸的生成量,GC检测底物己二酸的消耗量和可能副产物6-羟基己酸和己二醇等,6-氨基己酸的产率达到62.4%(滴度:62.4mM),未检测到任何副产物6-羟基己酸和己二醇等,该实施例中的6-氨基己酸产物滴度和实施例7中利用葡萄糖脱氢酶GDH再生NADPH时几乎一致。
实施例9:羧酸还原酶MabCAR3突变体在合成短链ω-氨基脂肪酸(C3-C5)中的应用
本实施例利用葡萄糖脱氢酶GDH再生NADPH,多磷酸激酶PPK再生ATP,CAR和TA级联催化α,ω-二元羧酸(C3-C5)合成ω-氨基脂肪酸(C3-C5),其它应用于该路线再生NADPH和ATP的生物酶催化剂也同样受此专利保护。反应体系8mL,在pH 6.5的Hepes-Na缓冲液中,30℃(通过水循环控温)、800rpm条件下进行,由于级联反应过程产生大量的酸,所以通过pHStat流加2M NaOH控制反应液的pH恒定。反应体系中包括终浓度为50mM丙二酸/丁二酸/戊二酸、0.5mM NADP(H)、5mM ATP、60mM MgCl
通过HPLC检测产物3-氨基丙酸/4-氨基丁酸/5-氨基戊酸的生成量,GC检测底物丙二酸/丁二酸/戊二酸的消耗量和可能副产物3-羟基丙酸/4-羟基丁酸/5-羟基戊酸和丙二醇/丁二醇/戊二醇等。结果如表4所示,5-氨基戊酸的产物滴度达到20.3mM,相比于6-氨基己酸,产物滴度有明显的降低,主要原因在于碳原子数降低至短链碳,CAR比活力明显降低,而且随着碳原子数的降低,产物滴度也呈现出逐渐降低的趋势,3-氨基丙酸和4-氨基丁酸的产物滴度分别为11.6mM和15.8mM。
表4 MabCAR3突变体和转氨酶级联催化短链α,ω-二元羧酸(C3-C5)合成ω-氨基脂肪酸
实施例10:羧酸还原酶MabCAR3突变体在合成中链ω-氨基脂肪酸(C7-C9)中的应用
本实施例利用葡萄糖脱氢酶GDH再生NADPH,多磷酸激酶PPK再生ATP,CAR和TA级联催化α,ω-二元羧酸(C7-C9)合成ω-氨基脂肪酸(C7-C9),其它应用于该路线再生NADPH和ATP的生物酶催化剂也同样受此专利保护。反应体系8mL,在pH 6.5的Hepes-Na缓冲液中,30℃(通过水循环控温)、1000rpm条件下进行,由于级联反应过程产生大量的酸,所以通过pHStat流加2M NaOH控制反应液的pH恒定。反应体系中包括终浓度为100mM庚二酸/辛二酸/壬二酸、1mM NADP(H)、10mM ATP、120mM MgCl
通过HPLC检测产物7-氨基庚酸/8-氨基辛酸/9-氨基壬酸的生成量,GC检测底物庚二酸/辛二酸/壬二酸的消耗量和可能副产物7-羟基庚酸/8-羟基辛酸/9-羟基壬酸和庚二醇/辛二醇/壬二醇等。如表5所示,相比于6-氨基己酸产率,7-氨基庚酸的产率更高,达到74.3%(74.3mM滴度),8-氨基辛酸和9-氨基壬酸滴度进一步提高,分别为82.5mM和90.3mM,未检测到任何副产物。
表5 MabCAR3突变体和转氨酶级联催化中链α,ω-二元羧酸(C7-C9)合成ω-氨基脂肪酸
实施例11:羧酸还原酶MabCAR3突变体在合成中长链ω-氨基脂肪酸(C10-C12)中应用
本实施例利用葡萄糖脱氢酶GDH再生NADPH,多磷酸激酶PPK再生ATP,CAR和TA级联催化α,ω-二元羧酸(C10-C12)合成ω-氨基脂肪酸(C10-C12),其它应用于该路线再生NADPH和ATP的生物酶催化剂也同样受此专利保护。反应体系8mL,在pH 6.5的Hepes-Na缓冲液中,30℃(通过水循环控温)、1000rpm条件下进行,由于级联反应过程产生大量的酸,所以通过pH Stat流加3M NaOH控制反应液的pH恒定。反应体系中包括终浓度为200mM癸二酸/十一烷二酸/十二烷二酸、1mM NADP(H)、15mM ATP、240mM MgCl
通过HPLC检测产物10-氨基癸酸/11-氨基十一酸/12-氨基十二酸的生成量,GC检测底物癸二酸/十一烷二酸/十二烷二酸的消耗量和可能副产物10-羟基癸酸/11-羟基十一烷酸/12-羟基十二烷酸和癸二醇/十一烷二醇/十二烷二醇等。随着碳链的增加,CAR比活力有增加的趋势,所以进一步的将底物上载量提高至200mM,结果如表6所示,10-氨基癸酸的滴度为110.4mM,产物11-氨基十一酸(122.3mM滴度)和12-氨基十二酸(103.9mM滴度)滴度和10-氨基癸酸相当,未检测到任何副产物。
表6 MabCAR3突变体和转氨酶级联催化中长链α,ω-二元羧酸(C10-C12)合成ω-氨基脂肪酸
实施例12:羧酸还原酶突变体MabCAR3
6-氨基己酸作为尼龙6的单体,本实施例中将反应体系进一步放大到300mL体系,利用葡萄糖脱氢酶GDH再生NADPH,多磷酸激酶PPK再生ATP,CAR和TA级联催化己二酸合成6-氨基己酸,在pH 6.5的Hepes-Na缓冲液中,30℃(通过水循环控温)、800rpm条件下进行,由于级联反应过程产生大量的酸,所以通过pH Stat流加2M NaOH控制反应液的pH恒定。反应体系中包括终浓度为100mM AA、1mM NADP
通过HPLC检测产物6-氨基己酸的生成量,GC检测底物己二酸的消耗量和可能副产物6-羟基己酸和己二醇等,6-氨基己酸的产率达到55.1%(55.1mM),未检测到任何副产物6-羟基己酸和己二醇等。最后,按照ω-氨基脂肪酸制备方法,从反应混合液中分离制备6-氨基己酸,并通过GC-MS进行验证,分离得率达到42.1%,成功实现尼龙单体的克级制备。
实施例13:野生型羧酸还原酶MabCAR3
本实施例利用葡萄糖脱氢酶GDH再生NADPH,多磷酸激酶PPK再生ATP,CAR和TA级联催化己二酸先合成6-氨基己酸,然后CAR和TA级联催化6-氨基己酸合成1,6-己二胺。也可以将其它再生NADPH和ATP的生物酶催化剂应用于该路线。反应体系8mL,在pH 6的Hepes-Na缓冲液中,30℃(通过水循环控温)、800rpm条件下进行,由于级联反应过程产生大量的酸,所以通过pH Stat流加2M NaOH控制反应液的pH恒定。反应体系中包括终浓度为100mM AA、2mMNADP(H)、10mM ATP、240mM MgCl
通过HPLC检测产物1,6-己二胺和中间产物6-氨基己酸的生成浓度,GC检测底物己二酸的消耗量和可能副产物6-羟基己酸和己二醇等,产物1,6-己二胺浓度为1.4mM,绝大部分积累在中间产物6-氨基己酸,未检测到任何副产物6-羟基己酸和己二醇等。这个结果表明野生型羧酸还原酶MabCAR3
实施例14:羧酸还原酶突变体MabCAR3
本实施例利用葡萄糖脱氢酶GDH再生NADPH,多磷酸激酶PPK再生ATP,MabCAR3
通过HPLC检测产物1,6-己二胺和中间产物6-氨基己酸的生成浓度,GC检测底物己二酸的消耗量和可能副产物6-羟基己酸和己二醇等,产物1,6-己二胺浓度提高到49.5mM,其余一些积累在中间产物6-氨基己酸(25.1mM)以及底物己二酸(15.3mM)少量未消耗完全,未检测到任何副产物6-羟基己酸和己二醇等。结果表明:相比于野生型羧酸还原酶MabCAR3
实施例15:羧酸还原酶MabCAR3突变体在合成短链α,ω-二胺(C3-C5)中应用
本实施例利用葡萄糖脱氢酶GDH再生NADPH,多磷酸激酶PPK再生ATP,CAR和TA级联连续羧酸还原胺化催化α,ω-二元羧酸(C3-C5)合成α,ω-二胺(C3-C5),其它应用于该路线再生NADPH和ATP的生物酶催化剂也同样受此专利保护。应体系8mL,在pH 6.5的Hepes-Na缓冲液中,25℃(通过水循环控温)、800rpm条件下进行,由于级联反应过程产生大量的酸,所以通过pH Stat流加2M NaOH控制反应液的pH恒定。反应体系中包括终浓度为50mM丙二酸/丁二酸/戊二酸、0.5mM NADP(H)、5mM ATP、120mM MgCl
通过HPLC检测产物1,3-丙二胺/1,4-丁二胺/1,5-戊二胺和中间产物3-氨基丙酸/4-氨基丁酸/5-氨基戊酸的生成浓度,GC检测底物丙二酸/丁二酸/戊二酸的消耗量和可能副产物ω-羟基脂肪酸和α,ω-二醇等。结果如表7所示,1,3-丙二胺、1,4-丁二胺和1,5-戊二胺的产物浓度分别为16.2mM,26.1mM和31.3mM,未检测到任何副产物,均有少量中间产物ω-氨基脂肪酸积累。
表7MabCAR3突变体和转氨酶级联催化短链α,ω-二元羧酸(C3-C5)合成α,ω-二胺
实施例16:羧酸还原酶MabCAR3突变体在合成中链α,ω-二胺(C7-C9)中的应用
本实施例利用葡萄糖脱氢酶GDH再生NADPH,多磷酸激酶PPK再生ATP,CAR和TA级联连续羧酸还原胺化α,ω-二元羧酸(C7-C9)合成α,ω-二胺(C7-C9),其它应用于该路线再生NADPH和ATP的生物酶催化剂也同样受此专利保护。反应体系8mL,在pH 6.5的Hepes-Na缓冲液中,30℃(通过水循环控温)、1000rpm条件下进行,由于级联反应过程产生大量的酸,所以通过pH Stat流加2M NaOH控制反应液的pH恒定。反应体系中包括终浓度为100mM庚二酸/辛二酸/壬二酸、1mM NADP(H)、10mM ATP、240mM MgCl
通过HPLC检测产物1,7-庚二胺/1,8-辛二胺/1,9-壬二胺和中间产物7-氨基庚酸/8-氨基辛酸/9-氨基壬酸的生成浓度,GC检测底物庚二酸/辛二酸/壬二酸的消耗量和可能副产物7-羟基庚酸/8-羟基辛酸/9-羟基壬酸和庚二醇/辛二醇/壬二醇等。结果如表8所示,1,7-庚二胺的产率达到60.2%(60.2mM),未检测到任何副产物。其产率高于实施例14中的1,6-己二胺浓度,且随着碳链长度的增加,α,ω-二胺产物滴度有小幅度提高,1,8-辛二胺和1,9-壬二胺滴度分别为71.8mM和80.2mM。
表8 MabCAR3突变体和转氨酶级联催化中链α,ω-二元羧酸(C7-C9)合成α,ω-二胺
实施例17:羧酸还原酶MabCAR3突变体在合成中长链α,ω-二胺(C10-C12)中的应用
本实施例利用葡萄糖脱氢酶GDH再生NADPH,多磷酸激酶PPK再生ATP,CAR和TA级联连续羧酸还原胺化α,ω-二元羧酸(C10-C12)合成α,ω-二胺(C10-C12),其它应用于该路线再生NADPH和ATP的生物酶催化剂也同样受此专利保护。反应体系8mL,在pH 6.5的Hepes-Na缓冲液中,25℃(通过水循环控温)、1000rpm条件下进行,由于级联反应过程产生大量的酸,所以通过pH Stat流加3M NaOH控制反应液的pH恒定。反应体系中包括终浓度为200mM癸二酸/十一烷二酸/十二烷二酸、2mM NADP(H)、15mM ATP、480mM MgCl
通过HPLC检测产物1,10-癸二胺/1,11-十一烷二胺/1,12-十二烷二胺和中间产物10-氨基癸酸/11-氨基十一酸/12-氨基十二酸的生成浓度,GC检测底物癸二酸/十一烷二酸/十二烷二酸的消耗量和可能副产物10-羟基癸酸/11-羟基十一酸/12-羟基十二酸和癸二醇/十一烷二醇/十二烷二醇等。由于链长进一步增加,CAR比活力有增加趋势,底物上载量也提高至200mM,结果如表9所示,1,10-癸二胺的产物滴度为105.3mM,未检测到任何副产物。本实施例中以生物基α,ω-二元羧酸作为底物,α,ω-二胺产物滴度是目前所有文献和专利报道中利用生物法合成最高的,尤其是针对中长链底物(C10-C12)时,底物位阻一般会更大,这也进一步的体现了本实施例中羧酸还原酶突变体的创新性和先进性。
表9MabCAR3突变体和转氨酶级联催化中长链α,ω-二元羧酸(C10-C12)合成α,ω-二胺
实施例18:羧酸还原酶MabCAR3突变体合成1,6-己二胺体系放大及产物制备
1,6-己二胺作为尼龙66的两个单体之一,本实施例中将反应体系进一步放大到300mL体系,利用葡萄糖脱氢酶GDH再生NADPH,多磷酸激酶PPK再生ATP,CAR和TA级联催化己二酸合成1,6-己二胺,在pH 6.5的Hepes-Na缓冲液中,30℃(通过水循环控温)、1000rpm条件下进行,由于级联反应过程产生大量的酸,所以通过pH Stat流加3M NaOH控制反应液的pH恒定。反应体系中包括终浓度为100mM AA、1mM NADP
通过HPLC检测产物1,6-己二胺的生成量以及中间产物6-氨基己酸的浓度,GC检测底物己二酸的消耗量和可能副产物6-羟基己酸和己二醇等,1,6-己二胺的产率达到42.1%,未检测到任何副产物6-羟基己酸和己二醇等。最后,可利用正丁醇从反应混合液中萃取出1,6-己二胺,然后通过旋蒸仪旋蒸干燥得到粉末,并通过GC-MS进行验证,分离得率达到31.2%,成功实现1,6-己二胺的克级制备。
以上所述实施例仅表达了本发明的几种实施方式,其描述较为具体和详细,但并不能因此而理解为对本发明专利范围的限制。应当指出的是,对于本领域的普通技术人员来说,在不脱离本发明构思的前提下,还可以做出若干变形和改进,这些都属于本发明的保护范围。因此,本发明专利的保护范围应以所附权利要求为准。同时,在本发明提及的所有文献都在本申请中引用作为参考,就如同每一篇文献被单独引用作为参考那样。
序列表
SEQ ID No.1
/>
SEQ ID No.2
SEQ ID No.3
atgcatcatcatcatcatcatgcaaccattaccaaccacatgccgaccgcagaactgcaggcactggatgcggcgcaccacctgcacccgttcagcgcaaacaacgc
cctgggtgaagaaggcacccgcgttattacccgtgcacgcggcgtctggctgaatgacagtgaaggtgaagaaattctggatgcaatggcaggtctgtggtgtgttaat
attgggtatggtcgtgatgaactggcagaagttgcagcgcgtcagatgcgtgagctgccgtactataatacgttttttaaaacaacccacgtgccggcaattgcactggct
cagaaactggcagaactggcaccgggagatctgaatcatgttttttttgcaggaggtggtagcgaagcgaacgacaccaatattcgtatggttcgcacctattggcaga
ataaaggtcaaccggaaaaaaccgttattattagccgtaaaaatgcatatcatggtagcaccgttgcaagcagcgcactgggtggtatggcaggtatgcatgcacagag
cggtctgattccggatgttcatcatattaatcagccgaattggtgggcagaaggtggtgatatggaccctgaagaatttggtctggcacgtgcacgtgaactggaagaag
caattctggaactgggtgaaaatcgtgttgcagcatttattgcagaaccggttcagggtgcaggtggtgttattgttgcaccggatagctattggccggaaattcagcgtat
ttgtgataaatatgatattctgctgattgcagatgaagttatttgtggttttggtcgtaccggtaattggtttggtacacagacaatgggtattcgtccgcatattatgaccattg
caaaaggtctgagcagcggttatgcaccgattggtggtagcattgtttgtgatgaagttgcacatgttattggtaaagatgaatttaatcatggttatacctatagcggtcatc
cggttgcagcagcagttgcactggaaaatctgcgtattctggaagaagaaaatattctggatcatgttcgtaatgttgcagcaccgtatctgaaagaaaaatgggaagca
ctgaccgatcatccgctggttggtgaagcaaaaattgttggtatgatggcaagcattgcactgaccccgaataaagcaagccgtgcaaaatttgcaagcgaaccgggta
caattggttatatttgtcgtgaacgttgttttgcaaataatctgattatgcgtcatgttggtgatcgtatgattattagcccgccgctggttattaccccggcagaaattgatgaaatgtttgttcgtattcgtaaaagcctggatgaagcacaggcagaaattgaaaaacagggtctgatgaaaagcgcataa*
SEQ ID No.4
atgtataaagatttagaaggaaaagtagtggtcataacaggttcatctacaggtttgggaaaatcaatggcgattcgttttgcgacagaaaaagctaaagtagttgtgaatt
atcgttctaaggaagacgaagctaacagcgttttagaagaaattaaaagagttggcggagaggctattgccgttaaaggtgacgtaacagttgagtctgatgtaatcaatt
tagttcaatctgcaattaaagaatttggaaagctagacgttatgattaacaacgcaggactagaaaatccggtttcatctcatgaaatgtctttaagcgattggaataaagta
attgatacgaacttaacgggagctttcttaggtagtcgtgaagcgattaaatattttgttgaaaatgatattaagggaacagttattaacatgtcgagtgttcacgagaaaatt
ccttggccattatttgttcattatgcagcaagtaaaggcggtatgaagcttatgactgaaacactggcattagaatacgctccaaaaggtattcgtgtaaataacattggacc
gggagcgattaatacaccgattaacgctgagaaatttgctgatcctgagcagcgtgcagatgtagaaagcatgattccaatgggatacatcggagagccggaagaaat
tgcagcagttgctgcatggctagcttcttcagaggcgagttatgtaacaggaattacgctctttgctgacggcggtatgacactgtacccatcattccaagcaggacgcggataa*
SEQ ID No.5
atgatcaacatctataaaatcgataaactgaacaactttaacctgaacaaccataaaaccgatgattatagcctgtgtaaagataaagataccgcactggaact
gacccagaaaaacatccagaaaatttatgattatcagcagaaactgtatgccgaaaaaaaagaaggtctgattattgcatttcaggcaatggatgcagcaggtaaagatg
gtaccattcgtgaagttctgaaagcactggcaccgcagggtgttcatgaaaaaccgtttaaaagcccgagcagcaccgaactggcacatgattatctgtggcgtgttcat
aatgcagttccggaaaaaggtgaaattaccatttttaatcgtagccattatgaagatgttctgattggtaaagttaaagaactgtataaatttcagaataaagcagatcgtatt
gatgaaaataccgttgttgataatcgttatgaagatattcgtaattttgaaaaatatctgtataataatagcgttcgtattattaaaatttttctgaatgttagcaaaaaagaacag
gcagaacgttttctgagccgtattgaagaaccggaaaaaaattggaaatttagcgatagcgattttgaagaacgtgtttattgggataaatatcagcaggcatttgaagatg
caattaatgcaaccagcaccaaagattgtccgtggtatgttgttccggcagatcgtaaatggtatatgcgttatgttgttagcgaaattgttgttaaaaccctggaagaaatg
aatccgaaatatccgaccgttaccaaagaaaccctggaacgttttgaaggttatcgtaccaaactgctggaagaatataattatgatctggataccattcgtccgattgaaaaa*
- 一种胸外科手术麻醉呼吸器
- 一种胸外科手术麻醉呼吸器