一种厌氧氨氧化系统的废水处理方法
文献发布时间:2024-04-18 19:58:30
技术领域
本发明属于废水处理领域,具体涉及一种厌氧氨氧化系统的废水处理方法。
背景技术
在当今大背景下,厌氧氨氧化(anammox)工艺被认为是极具推广潜力能够推动城市污水厂朝向能源中性转型的技术。凭借其低能源需求、低温室气体排放和高处理效率等优势,厌氧氨氧化技术有望成为替换传统污水生物脱氮处理的完美选择(Liu et al.,2023;Wang et al.,2019)。
尽管厌氧氨氧化性能卓著,但其生长倍增缓慢、敏感环境耐受性差以及关键基质亚硝酸盐(NO
投加以颗粒活性炭(GAC)为主的载体材料被认为是可行的方案。颗粒活性炭作为多孔碳基材料,具有高比表面积以及疏水性表面,是选择性吸附被富含疏水蛋白的胞外聚合物(EPS)包裹着的厌氧氨氧化菌(AnAOB)的优良吸附剂(Jia et al.,2017;Liang etal.,2022;Wang et al.,2023)。在缩短厌氧氨氧化反应器启动时间、促进生物质定殖与富集以及优化厌氧氨氧化性能等方面颗粒活性炭的赫赫功绩已被广泛报道。例如,深度处理环节的类低基质环境下(~60mgN/L),中枢厌氧氨氧化代谢基因在颗粒活性炭生物膜中的转录活性较常规厌氧氨氧化强了2-3倍,同时厌氧氨氧化菌获得了高富集率和高活性(Yanget al.,2021)。颗粒活性炭生物膜中厌氧氨氧化菌属Candidatus Brocadia驱动的异化硝酸盐还原氨过程和异养细胞合成,构建了更为灵活的代谢途径,在不含厌氧氨氧化接种物的低基质合成废水(70mg N/L)中获得了最高26.9%的厌氧氨氧化菌富集效果以及高于0.21kg N/m
发明内容
颗粒活性碳是厌氧氨氧化生物量定殖的良好载体,但人们对颗粒活性炭对暴露于极端饥饿压力下的厌氧氨氧化的影响仍然知之甚少。
为此,本发明构建了极低氮强度的不利生境,以颗粒活性炭为对照,在长达60天的饥饿抑制后,梯度恢复基质供应。本发明研究的重点聚焦在颗粒活性炭介导的厌氧氨氧化系统在饥饿胁迫下的自我守卫以及解除胁迫后的快速恢复,以微生物群落的恢复及动态演替、关键代谢行为的强化为靶向。所揭示的颗粒活性炭介导的厌氧氨氧化系统抵抗极低基质的恶劣环境的潜在代谢机制,有望为厌氧氨氧化应对实际污水的饥饿工况提供原理性依据以及技术性方案。
为了解决上述存在的技术问题,本申请提供如下技术方案:
本发明提供一种厌氧氨氧化系统的废水处理方法,包括如下步骤:
S11:向完全混合式厌氧(CSTR)反应器中加入颗粒活性碳和接种的厌氧氨氧化污泥;所述接种的厌氧氨氧化污泥包含厌氧氨氧化菌;所述颗粒活性碳占完全混合式厌氧反应器体积的20-30%;
S12:向所述完全混合式厌氧反应器中通入废水后进行持续低基质阶段反应;所述持续低基质阶段反应时,氨氮(NH
S13:将所述完全混合式厌氧反应器进行解除饥饿胁迫阶段反应,得到处理废水;所述解除饥饿胁迫阶段反应时,氨氮与亚硝态氮的浓度均为78-82mg N/L。
优选的,所述步骤S11中,接种的厌氧氨氧化污泥加入前用水冲洗3次,去除杂质和悬浮物。
优选的,所述完全混合式厌氧反应器由聚甲基丙烯酸甲酯制备得到。
优选的,完全混合式厌氧反应器采用遮光塑料膜遮罩避光,温度为33-37℃。
优选的,所述步骤S12中,废水的pH为6.5-7.5,所述废水包含钾盐、钙盐、镁盐、微量元素I和微量元素II;所述微量元素I包含FeSO
优选的,所述步骤S12中,完全混合式厌氧反应器中混合液挥发性悬浮固体浓度(MLVSS)为20-20.3g/L。
优选的,所述步骤S12中,通入废水后进行搅拌,搅拌的速度为50-70r/min。
优选的,所述步骤S12中,通入废水后水力停留时间(HRT)保持10-14h。
优选的,所述步骤S12中,持续低基质阶段反应的时间为55-65天。
优选的,所述步骤S13中,解除饥饿胁迫阶段反应的时间为55-65天。
本发明建立了两个有/无颗粒活性炭的完全混合式饥饿厌氧反应器,以研究颗粒活性炭对厌氧氨氧化系统性能、微生物动态和代谢特征的长期影响。结果表明,颗粒活性炭对厌氧氨氧化的长期饥饿压力(氨氮和亚硝态氮均为10mg-N/L,持续60天)具有缓冲能力。与不使用颗粒活性炭的反应器(27天)相比,使用颗粒活性炭的反应器的性能恢复滞后期(6天)更短;同时,在运行120天后,颗粒活性炭介导的厌氧反应器的总脱氮效率(84.9%)明显高于不使用颗粒活性炭的反应器的43.2%。微生物群落分析表明,颗粒活性炭介导的反应器中优势厌氧菌Candidatus Brocadia的丰度(2.77%)在恢复期后比非颗粒活性炭反应器中的丰度(1.04%)增加得更明显。完全氨氧化(comammox)硝化螺旋菌(comammoxNitrospira)在非颗粒活性炭厌氧生物反应器中占主导地位,并在饥饿条件下表现出“规模效应”;这是厌氧系统恢复过程中的一个中途现象,饥饿的厌氧菌在与完全氨氧化的底物竞争中处于弱势。
在颗粒活性炭介导的厌氧系统中,由于颗粒活性炭加速了恢复过程,完全氨氧化的丰度大大降低。代谢功能分析显示,颗粒活性炭在饥饿过程中增强了氨基酸代谢和糖核苷酸生物合成相关基因的表达,从而产生了更多具有氧化还原特性的胞外高分子物质来包裹和保护厌氧氨氧化微生物群以抵御饥饿,这可能进一步加强了颗粒活性炭在厌氧氨氧化系统中对胞外电子传递的调控作用。同时颗粒活性炭还能促进微生物交叉进食,进一步增强细菌的新陈代谢,这表明颗粒活性炭在为厌氧氨氧化复合菌群触发更多样化的新陈代谢网络方面具有卓越的潜力。这项发明表现了颗粒活性炭介导的氨氧化系统在不利的饥饿生境中的微生物和代谢机制,有利于将氨氧化技术广泛应用于低底质废水的市政处理。
本发明的技术方案相比现有技术具有以下优点:
颗粒活性炭介导可以强化厌氧氨氧化在低基质环境中的适应性和恢复能力。持续暴露在极端条件下(10mgN/L的氨氮和亚硝态氮)厌氧氨氧化会引发减活、凋亡和代谢异常等状态,这些负面影响在含有颗粒活性炭的反应器中是减轻的。氮负荷率梯度上升期间,颗粒活性炭介导的厌氧氨氧化表现出更加稳健的复原势头,在第120天恢复脱氮效率至84.9%,Candidatus Brocadia成为占主导份额的厌氧氨氧化菌。颗粒活性炭介导的厌氧氨氧化菌群中的反硝化与异化硝酸盐还原氨也同步出现良性发展。意外的发现颗粒活性炭能够选择性促进完全氨氧化Nitrospira代谢同时抑制典型Nitrospira。表明颗粒活性炭似乎扩大了完全氨氧化菌在工程系统中的生态位,而不是通常假设的寡营养环境。厌氧氨氧化菌属在颗粒活性炭表面微环境上衍生出更错综的代谢网络与更激烈的代谢活动,为将颗粒活性炭引入厌氧氨氧化系统以应对实际污水底物匮乏的复杂生态的可行性提供了理论依据。
附图说明
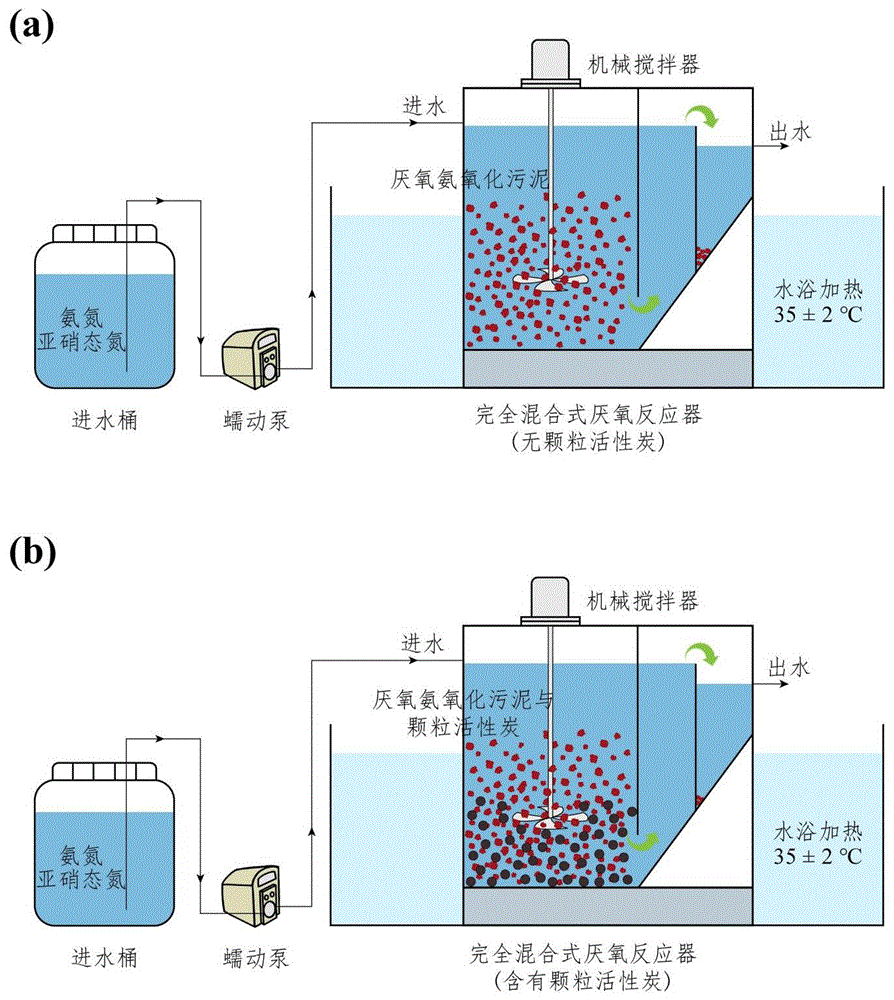
图1为R1(a)和R2(b)厌氧氨氧化生物反应器示意图。
图2为长期运行期间R1(a-c)和R2(d-f)的氮转化性能图。
图3为种子污泥、R1和R2污泥中的微生物群落结构和α多样性图。
图4为各阶段种子污泥、R1和R2污泥的代谢途径及相关基因的丰度图。
图5为各反应器在不同时期的异位试验监测比图。
图6为R1污泥样品在第0天(a、b)、第60天(c、d)和第120天(e、f)的扫描电子显微镜图像。
图7为R2污泥样品在第0天(a、b)、第60天(c、d)和第120天(e、f)的扫描电子显微镜图像。
图8为从R1(a-d)和R2(e-h)中提取的胞外聚合物的三位荧光激发-发射光谱图。
图9为不同pH下颗粒活性炭加入反应器前(a)和后(b)的Zeta电位图。
图10为颗粒活性炭投加进反应器之前(a)和之后(b)的傅立叶变换红外光谱图。
图11为R1、R2和种子污泥中微生物代谢概况图。
图12为通过实时荧光定量PCR对每个反应器中不同时期的羟胺氧化还原酶(hzo)基因进行定量图。
图13为颗粒活性炭介导厌氧氨氧化菌(包括厌氧氨氧化的恢复和多元代谢途径的构建)的拟议机制图。
具体实施方式
下面结合附图和具体实施例对本发明作进一步说明,以使本领域的技术人员可以更好地理解本发明并能予以实施,但所举实施例不作为对本发明的限定。
图2中,对应图片分别为:进水和出水中氨氮的浓度和去除率(a、d)。进水和出水中亚硝态氮的浓度和去除率,以及出水中硝态氮和总无机氮(TIN)的浓度(b、e)。总氮去除率(TNRE)、总氮负荷率(TNLR)、总氮去除率(TNRR)、亚硝态氮变化量与氨氮变化量比率以及硝态氮变化量与氨氮变化量比率(c、f)。
图3中,对应图片分别为:门分类微生物的相对丰度(a),属分类微生物的相对丰度(b),微生物的丰富度和多样性(c)。
图4中,对应图片分别为:厌氧氨氧化菌的基本代谢(包括TCA循环、Wood-Ljungdahl途径、氨基酸代谢和糖核苷酸生物合成)(a)。厌氧氨氧化菌群的氮代谢(b)。
图8中,对应图片分别为:第0天(a、e),第30天(b、f),第60天(c、g),第120天(d、h)。
图10中,3640cm
图11中,圆圈面积代表样本基因数与五组基因数最低值的差值。
实施例1反应器配置和操作条件
由有机玻璃制成的两个相同的实验室规模的完全混合式厌氧反应器,其中一个以颗粒活性炭为载体进行操作,另一个不添加载体作为对照。反应器有效容积为2.5L,其中反应区2.15L,沉淀区0.35L。根据载体条件不同,分别将两个反应器命名为R1(无颗粒活性炭)和R2(含有颗粒活性炭)(如图1)。以反应区容积的30%(v/v)的填充比向R2投放150g消过毒的颗粒活性炭。颗粒活性炭基本参数如表1所示。采用蠕动泵(BT101S,中国)由反应区顶部进水,控制系统水力停留时间(HRT)保持12h。反应区内置机械搅拌器(DJ1C-60,中国),搅拌速度保持在60r/min。两个反应器在恒温水浴缸内运行使系统内部温度保持35±2℃,并用遮光塑料膜遮罩避光,以抑制光养生物的生长,防止厌氧氨氧化活性下降。
表1.颗粒活性炭的基本特性参数
本试验持续120天,共分两个阶段:持续低基质阶段(阶段I,60天)和解除饥饿胁迫阶段(阶段II,60天)。在阶段I,通过保持进水低总氮负荷率(TNLR),持续营造饥饿环境,以对比研究饥饿胁迫对厌氧氨氧化与颗粒活性炭介导的厌氧氨氧化代谢活性的抑制程度。在阶段II,梯度恢复底物浓度至正常水平,研究解除饥饿胁迫后颗粒活性炭对厌氧氨氧化菌群及代谢性能恢复的影响。各阶段反应器运行条件详见表2。
表2.系统运行概述
在第二阶段,根据以下原则增加进水氮浓度:第60-90天,每10天梯度增加10mg N/L;第91-105天,每5天梯度增加12.5mg N/L;第106-120天,保持80mg N/L。
实施例2合成废水和接种污泥
实施例中使用的废水为人工合成废水。营养液和微量元素溶液I和II的组成如表3所示。使用前调节pH值至7.0±0.5。完全混合式厌氧反应器接种污泥收集自实验室规模的序批式(SBR)厌氧氨氧化反应器,该反应器已稳定运行1年以上(Zhang et al.,2022a)。其混合液挥发性悬浮固体浓度(MLVSS)为20.15±0.15g/L。系统运行前,用超纯水冲洗接种的厌氧氨氧化污泥3次,以去除杂质和悬浮物。然后,在每个反应器中加入0.35L的厌氧氨氧化污泥,以启动反应器。
表3合成废水的成分
实施例3化学分析
在进行化学分析之前,定期从完全混合式厌氧反应器中收集出水样品,并用定性滤纸(孔径:15-20μm)进行过滤。氨氮、亚硝态氮和硝态氮的浓度严格按照标准方法测定(Apha,1998)。总氮去除率(TNRE)、总氮负荷率(TNLR)和总氮去除率(TNRR)的计算公式见最近的研究(Chen et al.,2022;X.Zhang et al.,2022)。
实施例4异位活性检测
通过异位试验监测比厌氧氨氧化活性(SAA),以确定厌氧氨氧化菌的脱氮能力。实验在工作容积为120mL的150mL血清瓶中进行。取样后,污泥样品用矿物培养基洗涤三次,以去除泥浆中残留的氨氮、亚硝态氮和硝态氮。矿物介质的成分与用作完全混合式厌氧进水的合成废水相同,只是不提供氨氮和亚硝态氮。为了尽量减少颗粒活性炭对R2污泥比厌氧氨氧化活性可能产生的影响,在厌氧室(充满96%氮气-4%氢气混合气体)中用镊子夹取并筛分颗粒活性炭,人工去除R2污泥中的颗粒活性炭。R1污泥样品可省略这一步骤。然后,将大约10mL不含颗粒活性炭的污泥样品重新悬浮在110mL合成废水(接种物与进料比为1:11v/v)中,合成废水的成分与用于完全混合式厌氧反应器的进水完全相同。同时,另取10mL污泥样本用于测定挥发性悬浮固体(VSS)。加入0.1M HCl或NaOH将初始pH值调整为7.50(Xuet al.,2021)。所有血清瓶用丁基橡胶密封后,用>99.9%的氮气吹扫10分钟,以维持厌氧条件。培养在恒温振荡器中进行,温度和振荡速度控制在35±1℃和180rpm。所有试验均一式三份,以便进行统计分析。批量试验持续9小时,使用无菌注射器每1.5小时取样一次。收集液体样品用于氮化合物浓度测定。最后,监测MLVSS以计算特定反应速率,从而获得比厌氧氨氧化活性(mg-N/(g-VSS·d))。
实施例5生物质结构观测
使用扫描电子显微镜(SEM,日立S4300,日本)分析样品的表观生物质特性和颗粒活性炭结构特征。扫描电子显微镜样品的制备和检测按照之前的方法进行(Wu et al.,2010)。
实施例6胞外聚合物(EPS)和颗粒活性炭表征
采用阳离子交换树脂法(CER)从样品中提取胞外聚合物。提取胞外聚合物的三个组分,即S-EPS、LB-EPS和TB-EPS。提取S-EPS时,加入30mL的0.1M NaCl溶液,在4000g转速下离心10分钟。上清液经孔径为0.45μm的滤膜过滤,得到S-EPS。残留固体随后与相同浓度和数量的NaCl溶液一起在8000g下离心10分钟。再次使用相同的膜过滤上清液,得到L-EPS。在残留固体中加入阳离子交换树脂(Na
通过三维荧光激发-发射矩阵(3D-EEM)光谱鉴定了提取的胞外聚合物中的荧光化合物。使用FluoroMax-4分光光度计(Horiba,日本)测量3D-EEM荧光光谱。在200-400nm的激发波长范围内以5nm的采样间隔扫描8个未稀释的胞外聚合物样品,在220-550nm范围内每隔5nm采集发射光谱。激发和发射狭缝带宽均设定为5nm。三维荧光激发-发射矩阵图谱一式两份。
傅立叶变换红外光谱(FTIR)用于检测颗粒活性炭表面的主要官能团。从反应器中收集的颗粒活性炭样品首先用超纯水冲洗3次,以去除絮状物和残留基质。然后将冷冻的颗粒活性炭样品研磨并在真空干燥器中干燥48小时。然后在研磨机中将干燥的颗粒活性炭粉末与溴化钾按1:100的比例混合。将混合物压缩后,用傅立叶变换红外光谱仪(Nicolet5700,美国)测量4000-400cm
颗粒活性炭注入反应器前后的Zeta电位由Zetasize Nano ZS仪器(Malvern,美国)测量。从反应器中收集的颗粒活性炭样品首先用超纯水冲洗3次,以去除絮状物和残留基质。然后将颗粒活性炭样品研磨并分散在10mL的0.1M NaCl中。所有数据均通过计算六个平行样品的平均值得出(Hou et al.,2015)。未放入反应器中的颗粒活性炭也通过相同的程序进行测量。
实施例7微生物群落分析
(1)DNA提取、16S rRNA基因测序以及生物信息分析
分别在第0天、第60天和第120天从R1和R2采集了六个污泥样本,并冷冻干燥(Labconco Co.Free Zone2.0,美国)。种子污泥(即第0天的污泥)和R1污泥样品在4000rpm转速下离心10分钟,然后弃去上清液,从颗粒中提取DNA。R2污泥样品首先用镊子去除颗粒活性炭,然后进行上述DNA提取程序。根据制造商的说明,使用用于土壤的Fast DNA@SPIN试剂盒(MP Biomedicals LLC,美国)提取总DNA。提取DNA的质量由Nanodrop 3300(ThermoFisher Scientific,美国)进行评估。获得的DNA浓度由Qubit 4.0荧光仪(赛默飞世尔科技公司,美国)检测。
用通用引物338F(5′-ACTCCTRCGGAGGCAGCAG-3′)和806R(5′-GGACTACHVGGTWT-CTAAT-3′)对所有提取的V3-V4区DNA进行一式三份的原核生物16S rRNA基因聚合酶链反应(PCR)扩增,然后进行16S rRNA基因测序。PCR产物的测序在Illumina MiSeq PE300平台(Majorbio生物医药科技有限公司,中国上海)上进行。PCR扩增在以下热循环条件下进行:94℃初始变性10分钟,30个扩增循环(94℃、55℃和72℃各30秒),72℃最后延伸7分钟。对原始读数中的适配体和引物序列进行修剪。在去除具有模糊碱基调用、低质量分数或潜在嵌合体的原始读数后,使用DADA2 ver.1.18.0将合格序列聚类为扩增子序列变体(ampliconsequence variants,ASVs)。利用QIIME pipeline ver.1.9.0中的UCLUST工具,对照RDP数据库对鉴定出的扩增子序列变体进行分类(Choi et al.,2023)。根据特征扩增子序列变体在不同样本中的分布情况,评估了每个样本的α多样性指数(ACE、Chao1、Shannon和Simpson)。使用特征分类器插件中的贝叶斯分类器,针对Silva v132数据库对扩增子序列变体进行分类(Bokulich et al.,2018)。参考之前报道的研究,利用“未观察状态重建群落系统进化调查”(PICRUSt2)从扩增子序列变体数据和参考数据库(即Kyoto Encyclopediaof Genes and Genomes,KEGG)中预测功能基因(Douglas et al.,2020;Gao et al.,2023;Kanehisa et al.,2016)。通过访问美吉生物平台平台提供的“未观察状态重建群落系统进化调查”文件获得样本的KO和相应的基因丰度。使用Adobe Illustrator绘制代谢途径图,以比较各阶段样本的代谢能力。16S rRNA数据集已存入美国国家生物技术信息中心(NCBI)序列读取档案(生物项目登录号:PRJNA991303)。
实施例8实时定量聚合酶链反应(qPCR)测定
对代表厌氧氨氧化菌独特代谢转录过程的核心酶肼氧化还原酶(hzo)基因进行了定量分析。表4列出了目标片段的正反引物和反应方案。操作步骤详见之前的研究(Zhanget al.,2021)。表4用于qPCR分析的目标片段引物和协议
参考文献(Schmid et al.,2008)。
效果评价1脱氮性能比较
(1)无颗粒活性炭的厌氧反应器脱氮性能
在不投加颗粒活性炭的R1中,进水氨氮与亚硝态氮浓度分别为10.0(±2.6)mg/L与10.0(±1.9)mg/L,厌氧氨氧化系统临近奔溃,总氮去除率由最初的82.6%锐减至最低20.6%(如图2)。
低基质阶段结束时(第60天后),梯度提升进水底物浓度,逐步解除厌氧氨氧化菌饥饿胁迫。观察到R1的氨氮与亚硝态氮去除性能的缓和存在滞后期,去除率继续降低直至第88天的21.5%和24.6%;在此阶段(60-88天)进水氨氮与亚硝态氮浓度从10mg/L提升至40mg/L,出水氨氮与亚硝态氮浓度最高甚至达到32.0mg/L与31.6mg/L。性能好转开始于R1的第89天,观察到氨氮去除率在持续的31天内平稳上升。在此期间,进水氨氮与亚硝态氮浓度从40mg/L提升至80mg/L。在第120天运行结束时,出水浓度分别为氨氮33.3mg/L、亚硝态氮32.5mg/L和硝态氮10.8mg/L,相应的氨氮、亚硝态氮和总氮去除率分别为58.5%、59.9%和51.1%(如图2)。在整个低基质阶段,R1的总氮负荷率稳定在0.03(±0.01)kg-N/m
(2)颗粒活性炭介导的厌氧反应器脱氮性能
在投加颗粒活性炭的R2中,总氮去除率从初始的83.6%仅降至最低49.8%就开始回升。在低基质阶段R2出水中检测到的氨氮、亚硝态氮和硝态氮浓度与同期R1中的变化趋势相似(如图2)。解除饥饿胁迫后,R2滞后期仅从第60天维持到第66天,较R1缩短了12天;此时观察到氨氮、亚硝态氮去除率最低为52.1%和53.1%。从第67天起至第104天氨氮与亚硝态氮去除性能稳步回升,去除率提升至84.2±1.7%、85.5±1.3%。在此期间,总氮去除速率由0.03(±0.01)kgN/m
R2运行结束时,出水氨氮、亚硝态氮仅为6.82-8.04mg/L与3.45-7.06mg/L,氨氮、亚硝态氮和总氮去除率分别为91.2±1.0%、93.0±0.6%和83.6±1.3%,总氮负荷率为0.32(±0.01)kg N/m
效果评价2污泥性状
通过扫描电子显微镜观察了R1(常规厌氧氨氧化)and R2(颗粒活性炭介导的厌氧氨氧化)中的污泥结构与表面形态(如图6和图7)。第一阶段初期,R1中存在大量以球状、椭球状或杆状形式存在的细菌污泥形态(如图6)。相较而言,R2花椰菜状的菌落形态更加明显,表面存在错落有致的凸起,尺寸约为1.0μm;胞外聚合物被发现将厌氧氨氧化菌固定在颗粒活性炭上,整体附着状态初步形成(如图7)。第一阶段后期,由于进水基质浓度持续降低,可以看到R1中的菌体分泌大量胞外聚合物抵抗不适周境,然而持续的饥饿胁迫使胞外聚合物被菌体内源消耗殆尽,细菌数量大幅衰减,且剩余细菌呈现破损或裂解的状态(如图6)。由于颗粒活性炭的负载,R2中留存了少量胞外聚合物(如图8)维持着小部分厌氧氨氧化菌的基础代谢活性,但虚弱的厌氧氨氧化菌也表现出褶皱萎缩的形态趋势(如图7)。随着饥饿胁迫解除,在第二阶段后期,R1内的厌氧氨氧化菌外形重新变得饱满(如图6),胞外聚合物分泌仍处于较低水平,同时由于菌体粒径较小,花椰菜结构消失。R2中颗粒活性炭被厌氧氨氧化完全包裹着,在其表面形成菌膜状结构,同时厌氧氨氧化形态较为丰盈,从菌体饱满的外形可直观看出其活性在颗粒活性炭负载下得到了明显恢复(如图7)。
三维荧光激发-发射矩阵(3D-EEM)荧光光谱分析揭示了污泥样品胞外聚合物的成分变化(如图8)。初始阶段,在250-300/280-360、200-250/330-380的激发/发射波长(Ex/Em)中,从R1(无颗粒活性炭)和R2(含有颗粒活性炭)内识别出两个较为明显的特征荧光峰,分别对应于色氨酸蛋白(峰A)和酪氨酸蛋白(峰B)(Wang et al.,2020)。试验开始前,R1与R2种泥的基质条件适宜,较明显的峰A和峰B显示了较高含量的胞外聚合物能够反映厌氧氨氧化代谢活动正常(a,e)。当进水氨氮和亚硝态氮浓度由试验开始前的80mg/L降至第一阶段的10mg/L时,R1和R2中色氨酸和酪氨酸的荧光强度明显增强,尤其是保持底物缺乏环境的30天后,荧光强度达到最强水平(如图8)。随着低底物供给周期的持续延长,两个反应器的峰A和峰B的峰强显著跌落(如图8),R1中甚至无法清晰识别出直观的峰,R2仅能辨别出模糊的界限。第二阶段逐步解除饥饿胁迫,当进水氨氮和亚硝态氮梯度递增至80mg N/L时,R1峰强缓慢回升但幅度极小,而R2峰强恢复超过初始水平,特别是类色氨酸物质。
不同pH条件下颗粒活性炭投入反应器前后的Zeta电位结果显示(如图9)。颗粒活性炭投入反应器前的零电荷点(pH
此外,在500-4000cm
效果评价3微生物群落演替
(1)门水平上的微生物群落演替
在阶段I-II结束时通过16s rRNA基因扩增子测序揭示R1与R2在门水平的微生物群落结构(如图3)。归属于浮游霉菌门、变形杆菌门和绿弯菌门门的细菌在种泥中占据主导地位,相对丰度分别为13.3%、18.5%和24.9%。这些功能微生物对厌氧氨氧化菌的茁壮萌发以及厌氧氨氧化菌群氮转化性能的改良可能发挥着协同互补作用。
随着低氮冲击时间的推移,R1中厌氧氨氧化菌群逐渐以硝化螺旋菌门、绿弯菌门、变形杆菌门和拟杆菌门为主导,他们的富集率分别为22.5%、22.8%、11.3%和9.6%;R2中厌氧氨氧化菌群也表现出类似演变情况,四种优势菌门分别占比20.6%、11.4%、22.5%和8.2%。值得注意的是,硝化螺旋菌门在饥饿胁迫阶段成为两组反应器中丰度排序靠前的菌门。隶属于浮游霉菌门的厌氧氨氧化菌无法在竞争中成为有利者,在R1与R2中比例分别仅为5.4%和6.8%。
营养条件恢复后,R1中的浮游霉菌门因长期暴露于低氮冲击,丰度持续衰减至4.3%;相反,在R2中浮游霉菌门受到颗粒活性炭保护,丰度显著跃升至10.9%。整个运行阶段结束时,两个反应器中厌氧氨氧化菌群现差异化演替结果。在R1中,硝化螺旋菌门以35.0%的高丰度成为唯一优势菌门,变形杆菌门与拟杆菌门分别富集至14.3%和12.6%而紧随其后。而R2中优势菌门演替为变形杆菌门、绿弯菌门和浮游霉菌门,其中变形杆菌门、绿弯菌门分别占总门的20.9%和19.7%。颗粒活性炭可作为促进厌氧氨氧化菌群健康进化的营地,使得历经饥饿胁迫的厌氧氨氧化菌能以颗粒活性炭为骨料重获生机。
(2)属水平上的微生物群落演替
为了进一步阐明颗粒活性炭负载对厌氧氨氧化响应低氮饥饿条件下的微生物群落结构与演变的潜在影响,从属水平上讨论了常规厌氧氨氧化与颗粒活性炭介导的厌氧氨氧化由种泥开始的菌群演替。如图3显示了从各反应阶段末期检测到的以变形杆菌门、浮游霉菌门、硝化螺旋菌门、绿弯菌门和拟杆菌门为主的菌门的优势属。R1中检测到厌氧氨氧化功能菌Candidatus Brocadia与Candidatus Kuenenia在整个菌群中含量微不足道。R2厌氧氨氧化污泥种群基本恢复到与种泥相近的菌属水平;其中,变形杆菌门以Denitratisoma为主,Candidatus Brocadia占浮游霉菌门的比例最大,Nitrospira在硝化螺旋菌门中占优势,SBR1031是属于绿弯菌门的优势属,拟杆菌门中Ignavibacterium的富集程度最高。
具体而言,种泥中包含两种厌氧氨氧化微生物Candidatus Brocadia与Candidatus Kuenenia,其初始丰度分别为2.72%与1.20%。在R1与R2全周期运行后,Candidatus Kuenenia丰度随着氮负荷率的逐步回升分别由0.12%与0.39%浮升至0.26%与0.66%,而Candidatus Brocadia分别由0.33%与1.67%跃升为1.04%与2.77%。
在种泥、R1与R2污泥中检出了相同菌属不同含量的反硝化菌以及异化硝酸盐还原氨菌。其中,A4b在R2中由0.84%迅速恢复至1.45%;在R1中丰度由1.06%持续降低至0.91%,明显低于种泥中的1.46%。SBR1031在R1、R2以及种泥中均为主优势属或次优势属,分别占13.11%、6.23%和9.60%。OLB14在R1中由初始的2.98%持续降至末期的0.79%;在R2中由1.13%变为最终的1.66%。Gemmatimonadaceae在R2中丰度3.34%显著高于R1中的1.71%。此外,Anaerolineae、Ignavibacterium、Denitratisoma与Limnobacter在R2中富集分别占比1.87%、1.07%、4.25%和0.46%;在R1中丰度仅为0.87%、0.39%、1.25%和0.34%。
硝化菌属Nitrospira在R1中占比达34.84%;在R2中丰度降至9.72%。从代谢相关基因分析得出,系统中的Nitrospira应当是完全氨氧化Nitrospira。下一节从代谢基因角度详细展开讨论。
效果评价4颗粒活性炭介导与厌氧氨氧化微生物代谢之间的联系
(1)微生物代谢总览
代谢途径差异总览(如图11)表征了各阶段样品中主要代谢相关的基因数与五组基因数最低值之间的差值。对于R2,在颗粒活性炭改良之后,大多数代谢途径的基因计数都增加了。其中上调极其突出的基因主要对应菌群的基础代谢,包括糖核苷酸生物合成和蛋白质代谢。
(2)代谢相关的关键酶基因的差异表达
利用“未观察状态重建群落系统进化调查”对16srRNA测序数据进行功能预测分析,参考KEGG数据库进行功能基因丰度统计,得到R1与R2中厌氧氨氧化菌在各运行阶段的代谢相关基因丰度比对结果(如图4)。由于缺乏KEGG注释指南,催化厌氧氨氧化的关键酶联氨合成酶(hzs)、联氨脱氢酶(hdh)和联氨氧化酶(hzo)的基因未被鉴定。通过实时荧光定量PCR定量了厌氧氨氧化菌氮代谢末梢特有的酶联氨氧化酶(hzo)的基因丰度(如图12)。对比R1,R2中hzo基因定量始终更多。厌氧氨氧化菌氮代谢另一核心酶羟胺脱氢酶(hao)也具有同样的氮转化能力,其基因在不同反应器样品中被检出各异的丰度。对比R1,R2中hao基因相对丰度始终更多,这与较高的厌氧氨氧化菌相对丰度一致。样品中仅完全氨氧化Nitrospira独有的基因能够进一步证实系统中的并非典型Nitrospira(表5),即碱性磷酸酶(phoD)、铜稳态基因(copCD and copAB)以及NO反应调节酶(nnrS)。amoABC基因的表达也暗示了羟胺在本厌氧氨氧化系统中很大可能是中间体。
表5参与完全氨氧化(comammox)的特定基因的相对丰度
柠檬酸循环(三羧酸循环,TCAcycle)与伍德-永达尔(Wood-Ljungdahl)途径是碳水化合物代谢的重要组成部分,它们共同作用于厌氧氨氧化固定CO
以厌氧氨氧化合成UDP-N-乙酰甘露糖胺和CDP-葡萄糖为主的糖核苷酸生物合成途径相关基因也作为检测的一部分。R1与R2系统均编码了部分胞外多糖前体物的生物合成途径,包括果糖代谢产生的UDP-N-乙酰葡萄糖胺;但颗粒活性炭刺激了大多数基因在R2中高度表达,如葡萄糖-6-磷酸异构酶(pgi)、谷氨酰胺果糖-6-磷酸转氨酶2(glmS)、1-磷酸葡萄糖胞苷酸转移酶(rfbF)以及UDP-N-乙酰葡糖胺2-酰亚胺酶(wecB)。
此外,R2氨基酸代谢相关基因丰度也较R1更高。颗粒活性炭促升了大多数氨基酸合成基因的相对丰度,如2-异丙基丙二酸合成酶(leuA)、组胺醇磷酸转氨酶(hisC)、二羟基酸脱水酶(ilvD)、谷氨酰胺合成酶(glnA)和5-甲基四氢叶酸--高半胱氨酸甲基转移酶(metH),并在运行后期达到峰值,促进了亮氨酸、组氨酸、异亮氨酸、谷氨酰胺和蛋氨酸等氨基酸的产生,从而助力解除饥饿胁迫后胞外聚合物中的蛋白质合成。
效果评价5
实施例中构建了两个有/无颗粒活性炭的连续流饥饿厌氧氨氧化反应器,通过控制长期的极低基质条件(氨氮与亚硝态氮均为10mg N/L,60天),揭示了颗粒活性炭能够缓和恶劣饥饿胁迫对厌氧氨氧化的不利影响;并表明颗粒活性炭在解除饥饿胁迫(梯度恢复进水基质为80mg N/L)的60天内就能修复极低底物(10mg N/L)对厌氧氨氧化菌群造成的创伤,在相同时间内,无颗粒活性炭负载的厌氧氨氧化无法实现自行恢复效果。在菌群恢复以及代谢强化方面拟议了保护和增益机制(如图13),包括(i)颗粒活性炭活性吸附位点为厌氧氨氧化菌落提供利于黏附存活的港湾,以抵抗极端饥饿的残酷生境;(ii)颗粒活性炭刺激基础代谢以加强胞外聚合物合成分泌,形成保护层,并诱导厌氧氨氧化快速恢复;(iii)颗粒活性炭构建更多元的代谢环路,助力厌氧氨氧化、完全氨氧化、反硝化与异化硝酸盐还原氨共代谢与交叉饲喂;(iv)颗粒活性炭充当胞外不溶性电子受体介导基于胞外电子传递的厌氧氨氧化(EET-anammox)。
显然,上述实施例仅仅是为清楚地说明所作的举例,并非对实施方式的限定。对于所属领域的普通技术人员来说,在上述说明的基础上还可以做出其它不同形式变化或变动。这里无需也无法对所有的实施方式予以穷举。而由此所引申出的显而易见的变化或变动仍处于本发明创造的保护范围之中。