一种创建高棕榈酸和多叶片转基因植株的方法及应用
文献发布时间:2023-06-19 10:33:45
技术领域
本发明涉及一种应用脂酰-酰基载体蛋白硫酯酶AhFATB2基因创建高棕榈酸和多叶片转基因植株的方法及应用,属于植物基因工程技术领域。
背景技术
在脂肪酸合成过程中,脂酰-酰基载体蛋白硫酯酶(Fat)通过催化硫酯键的水解决定了脂肪酸链长度。根据序列和底物的特异性Fat分为两类,分别是FatA和FatB,这两类Fat构成了TE14硫酯活性酶基因家族。FatA在不饱和长链脂酰-酰基载体蛋白合成中发挥特异性的作用,尤其在油酰辅酶A(oleoyl-ACP)的合成过程,而在饱和脂酰-酰基载体蛋白合成过程中活性较低。FatB主要以8-18碳链为底物生成的饱和脂酰-酰基载体蛋白。FatBs又可以分为FatB1和FatB2两个亚类,FatB1存在于所有植物中,主要以长链饱和酰基-ACP为底物,尤其是16:0-ACP;FatB2仅存在于积累8-14碳链脂肪酸的植物中,主要以短链及中链饱和脂肪酸为底物。Fat底物特异性对脂肪酸组分调控至关重要,因此Fat编码基因决定了脂肪组分。
目前已经克隆了多种植物中以8-18碳链为底物的Fat基因,并且很多长链特异性的FatB通过转基因验证了功能。过表达GarmFatA1、MlFatB和AtFATA2导致硬脂酸(18:0)含量增加,过表达AtFatB1、BnFatB(2),烟草中过表达CpFatB2a使得叶片中的十四烷酸含量(14:0)增加了46%,而过表达ClFatB1和GmFATA1同时增加了棕榈酸(16:0)和硬脂酸(18:0)的含量。JtFatB1、AtFatB主要增加了棕榈酸(16:0)的含量。油菜中过表达UcFatB1导致月桂酸(12:0)含量从忽略不计增加到接近60%,烟草中过表达CpFatB2a使得叶片中的十四烷酸含量(14:0)增加了46%,而过表达ClFatB1和GmFATA1同时增加了棕榈酸(16:0)和硬脂酸(18:0)的含量。
棕榈酸(Palmitic acid),是一种饱和高级脂肪酸,以甘油酯的形式普遍存在于动植物油脂中,主要来自于棕榈油和棕榈仁油。棕榈酸不含C=C双键,属于饱和脂肪酸非必需脂肪酸,且与儿童生长发育和成长健康有关,更有降血脂、防治冠心病等治疗作用,且与智力发育、记忆等生理功能有一定关系。棕榈酸的酯化反应可以产生生物柴油,证明可作为生物柴油的原料油,棕榈酸酯是一种典型的脂肪酸酯,可用作增塑剂、表面活性剂、精细化工产品等的原料或中间体,广泛用于农业、食品、医药和精细化工产品的制造。东南亚国家利用优越的气候条件大量种植棕榈酸来获得棕榈酸,用在生物柴油的原料。因此棕榈酸在食品加工和工业生产中具有广泛的应用价值。
发明内容
针对现有技术的不足,本发明的目的是提供一种应用脂酰-酰基载体蛋白硫酯酶AhFATB2基因创建高棕榈酸和多叶片转基因植株的方法及应用。本发明方法以野生型植物为转基因受体,通过基因工程的手段,应用脂酰-酰基载体蛋白硫酯酶(AhFATB2)基因,通过过表达AhFATB2基因,人工创建转基因植物。该转基因植物可以提高植物种子中棕榈酸含量及其叶片生物量,具有一定的开发价值。
为了实现上述目的,本发明所采用的技术方案是:
一种脂酰-酰基载体蛋白硫酯酶AhFATB2基因,所述AhFATB2基因的序列如SEQ IDNO.3所示。
一种应用脂酰-酰基载体蛋白硫酯酶AhFATB2基因创建转基因植株的方法,以野生型植物为转基因受体,通过基因工程手段,应用脂酰-酰基载体蛋白硫酯酶AhFATB2基因,通过过表达AhFATB2基因,人工创建转基因植物;所述AhFATB2基因的序列如SEQ ID NO.3所示。
所述的创建转基因植株的方法,包括以下步骤:
(1)花生脂酰-酰基载体蛋白硫酯酶AhFATB2基因的克隆;
(2)AhFATB2基因的植物过表达载体pBI-AhFATB2的构建;
(3)pBI-AhFATB2在拟南芥中的遗传转化和阳性苗筛选,获得转基因植株。
所述AhFATB2基因的克隆方法为:利用Trizol进行花生种子RNA的提取,以RNA逆转录的cDNA为模板进行AhFATB2基因的扩增,引物为AhFATB2S和AhFATB2A;
AhFATB2S:5′-CGCGGATCCATGGTTGCTACTGCTGCTACG-3′;
AhFATB2A:5′-GACGAGCTCTCAGTTTTCTGCTGGAAAAACC-3′。
所述AhFATB2基因扩增的反应体系为25μL内含:1×PCRbuffer,MgCI 1.5mmol/L,dNTP0.2mmol/L,引物浓度均为0.5mol/L,Pfu酶1.5单位,模板100ng;
PCR程序为:94℃,5min;94℃,45s,57℃,45s,72℃,2min,33个循环;72℃,10min。
所述过表达载体pBI-AhFATB2的构建方法为:用步骤(1)克隆得到的AhFATB2基因片段替换质粒pBI121上的GUS基因片段。
所述过表达载体pBI-AhFATB2构建的具体方法为:用BamHI和SacI双酶切克隆载体pBI121的GUS片段,同时用BamHI和SacI酶切步骤(1)克隆得到的AhFATB2基因片段;将AhFATB2基因片段的酶切产物和克隆载体pBI121上切下的片段混合,用DNA凝胶回收试剂盒回收,混合后加入T4 DNA连接酶5单位,10×反应缓冲液,用无菌水补充体积至20μL,16℃连接过夜,转化后,即得重组质粒pBI-AhFATB2。
所述步骤(3)的具体方法为:制备含有重组质粒pBI-AhFATB2的根癌农杆菌GV3101菌液,转化植株,筛选得到重组质粒pBI-AhFATB2成功转入植株的转基因阳性苗,将阳性苗自交3代以上,获得纯合植株HF-AhFATB2。
所述阳性苗的筛选方法为:当叶片长到3-4叶期时,取绿苗叶片进行PCR阳性检测,引物为:
AhFATB2S:5′-CGCGGATCCATGGTTGCTACTGCTGCTACG-3′;
AhFATB2A:5′-GACGAGCTCTCAGTTTTCTGCTGGAAAAACC-3′;
PCR反应体系为:基因组DNA模板1μL、10×Taq酶反应缓冲液2uL、25mMMgCL
反应程序为:94℃变性5min;94℃,45s;55℃,45s;72℃,2min;32循环,72℃延伸5min。
所述的脂酰-酰基载体蛋白硫酯酶AhFATB2基因在提高油料作物种子棕榈酸含量和提高植物叶片生物量中的应用。
本发明有益效果:
本发明提供成功克隆了脂酰-酰基载体蛋白硫酯酶AhFATB2基因,通过应用证明该基因可以应用于该转基因植物,可提高植物种子中棕榈酸含量及其叶片生物量,具有一定开发价值。
本发明以野生型植物为转基因受体,通过过表达AhFATB2基因,人工创建了转基因植物。其中通过转基因技术过表达AhFATB2基因,研究该基因在植物发育及种子棕榈酸合成中的作用,明确了该基因具有提高植物棕榈酸含量及增加植物叶片生物量的功能。与此同时,该基因的过表达不会引起其他可见植物表型的变化,不会引起植物表型缺陷。
本发明花生中的AhFATB2基因,可应用在植物基因工程和油料作物种子生产以及蔬菜、烟草等作物的叶片生成中,用于提高植物种子中棕榈酸含量及其叶片生物量。可以通过人工构建AhFATB2过表达载体,或其它的基因工程、分子生物学手段,增加或强化AhFATB2基因的表达,人工创建高棕榈酸及高叶片生物量的植物,具备一定的开发价值。如利用该基因人工创建具有某些优异农艺性状的植物材料,开发具有高棕榈酸含量的油料作物,如创制高棕榈酸的油菜、花生、大豆等油料作物,以满足各类商业需求;也可以利用该基因人工创制多叶片植物材料,如开发以收获叶片为目的的作物如烟草、饲草或蔬菜,以满足不同的需求。
附图说明
图1为一种花生AhFATB2基因克隆的电泳图;
其中:泳道1为以花生种子的DNA为模板的PCR扩增结果;泳道2为以花生种子的cDNA为模板的PCR扩增结果;泳道M为核酸Marker;
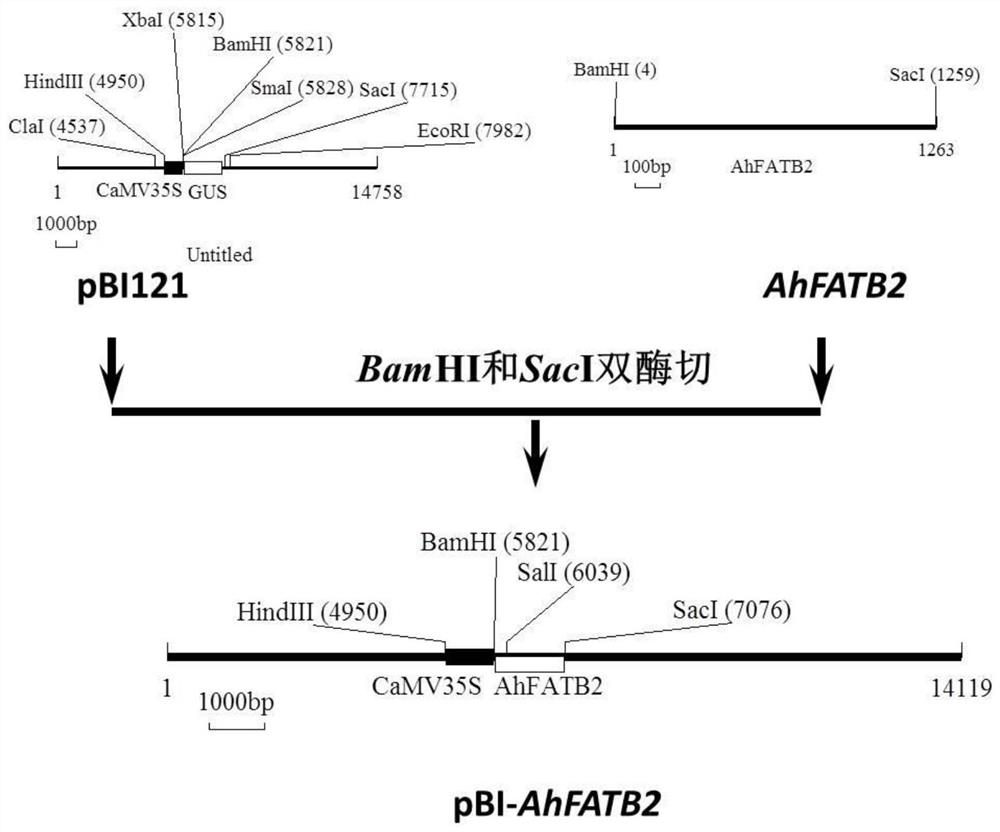
图2为一种构建的pBI-AhFATB2载体示意图;
图3为一种转化pBI-AhFATB2的转基因植株PCR鉴定图;
其中:M,DNAmarker DL2000;Lane 1、2、3、4、5、6、7、8,阳性株;Lane CK,野生型拟南芥;P,质粒对照;
图4为一种过表达AhFATB2基因植株后代多叶片表型;
其中:A,转基因植株1叶片;B,转基因植株2叶片;C,野生型拟南芥叶片;
图5为一种过表达AhFATB2基因植株后代种子脂肪酸组分分析;
其中:A,转基因植株1种子油组分分析;B,转基因植株2种子油组分分析;C,野生型拟南芥种子油组分分析。
具体实施方式
以下结合实施例对本发明的具体实施方式作进一步详细说明。
实施例1:
花生脂酰-酰基载体蛋白硫酯酶(AhFATB2)基因的克隆:
取花生开花后40天至70天之间任意阶段的种子中的子叶0.05-0.1g,在液氮中研磨至粉状,根据Trizol提取试剂盒(购自Invitrogen公司)的要求进行RNA的提取。提取的总RNA溶于60uL的无RNase的双蒸水中。用DNaseⅠ去除可能残留的DNA。用蛋白检测仪(DU650BECKMAN,USA)分别检测RNA在260纳米和280纳米下的光吸收值,结合1%(质量体积比)琼脂糖凝胶电泳鉴定RNA的纯度及浓度。
以上述获得的RNA为模板,按下列方案进行逆转录:2μg RNA中加1μL Oligo(dT),70℃温育5min,立即置于冰上5min,短暂离心,加入5×M-MLVBuffer 4μL,dNTP(10mmol/L)1μL,RNase Inhibitor 20单位,M-MLV逆转录酶(购自Promega公司)200单位,用DEPC(焦碳酸二乙酯)处理过的无菌水补至总体积20μL,混匀,42℃温育1h,70℃水浴15min,得到的cDNA分装后于-20℃保存,备用。
以花生种子cDNA为模板,进行AhFATB2基因的全长克隆。根据花生AhFATB2基因序列设计引物,引物为:
AhFATB2S:5′-CGC
AhFATB2A:5′-GAC
引物序列的5′末端分别引入BamHI和SacI限制性内切酶。
反应体系为25μL内含:1×PCRbuffer,MgCI 1.5mmol/L,dNTP 0.2mmol/L,引物浓度均为0.5mol/L,Pfu酶1.5单位,模板100ng。
PCR程序为:94℃,5min;94℃,45s,57℃,45s,72℃,2min,33个循环;72℃,10min。
PCR反应产物在1%(质量体积比)的低熔点琼脂糖上电泳,结果如图1所示。
把扩增产物条带从胶上切下放入1.5ml Eppendorf离心管中,65℃水浴15min,加入等体积苯酚(PH7.9),颠倒摇匀5min,13000转/分钟离心8分钟,取上清,加入等体积氯仿和异戊醇的混合物(氯仿:异戊醇的体积比为24:1)颠倒摇匀5min,13000转/分钟离心8分钟,取上清,加入1/10体积的3mol/L醋酸钠(PH5.2)溶液,2倍体积预冷的95%乙醇,混匀后置-20℃冰箱中20min以上,13000转/分钟离心15分钟,倒掉95%乙醇后再用75%乙醇浸洗沉淀,自然风干,DNA沉淀溶于20μl无菌去离子水。回收纯化目的片段,获得AhFATB2基因。该基因序列如SEQ ID NO.3所示。
实施例2:
AhFATB2基因的植物过表达载体pBI-AhFATB2的构建和根癌农杆菌菌株GV3101的转化:
构建的重组载体pBI-AhFATB2是将质粒pBI121上的GUS基因用前期克隆得到的AhFATB2基因片段替换而来,载体图谱如图2所示。
首先,用BamHI和SacI双酶切克隆载体pBI121的GUS片段,同时用BamHI和SacI酶切AhFATB2基因片段。酶切反应在37度培养箱中进行,约4-6个小时之后,用1%琼脂糖胶电泳检测。
将AhFATB2基因片段的酶切产物和克隆载体pBI121上切下的大片段用DNA凝胶回收试剂盒回收。将AhFATB2基因片段的酶切产物与pBI121载体按照3:1的比例混合样品(其中AhFATB2基因片段的酶切产物150ng,pBI121载体片段50ng),加入T4 DNA连接酶5单位,10×反应缓冲液,无菌水补充体积至20μL,16℃连接过夜。
转化后,利用冻融法将pBI-AhFATB2转入根癌农杆菌GV3101(购于上海沪尚生物科技有限公司),在含有卡那霉素(50μg/mL)和利福平(50μg/mL)的固体LB平板上进行双抗性平板筛选,挑斑做菌落PCR,BamHI和SacI双切酶切验证,将验证正确的重组质粒命名为pBI-AhFATB2。
实施例3:
pBI-AhFATB2在拟南芥中的遗传转化及转基因植株筛选:
制备含有构建好载体pBI-AhFATB2的根癌农杆菌GV3101菌液,在转化前一天转入含有卡那霉素50μg/ml、利福平50μg/ml的LB液体培养基中,28℃过夜培养。第二天,用紫外分光光度计(SPEKOL 1300)于276nm波长下检测吸光值,当菌液的吸光值达到1.6-2.0之间时取出。室温(20-25℃)下以4000g离心力离心10min,弃上清,沉淀悬浮于等体积的5%蔗糖(质量体积比)中。将浑浊的蔗糖溶液倒入一大培养皿中,转化前加入终浓度为0.02%(体积比)的Silwet l-77(购自北京五洲元业科贸中心)。
混匀后把待转化的拟南芥整个花序轻轻的浸没在蔗糖中15s,取出植株。转化后的植株用一个黑色塑料袋包好,放在生长箱培养。第二天将塑料袋揭开,放在光强的地方培养。隔一周再做一次转化。培养一个月左右收获种子,种子在培养箱或者日光下干燥3-5天。
将转化收获的T0代种子用70%(体积比)酒精和0.01%(体积比)的升汞表面消毒10分钟后用蒸馏水洗涤数次(5~7次),然后均匀地吹打到MS固体筛选培养基表面。(MS大量元素母液100ml;MS微量元素母液10ml;MS有机母液10ml;MS铁盐10ml;肌醇10ml;蔗糖30g;用1M NaOH调PH至5.8,12g琼脂粉,定容至1L,高压121度灭菌后备用。加入8g琼脂粉可配置成固体培养基。各种母液配方见表1)。
表1 MS培养基母液配方
4℃春化4-6天,放入恒温培养箱培养。根据表达载体上所特有的卡那霉素抗性筛选阳性苗。叶片长到足够大时(3-4叶期),取少许绿苗叶片进行PCR阳性检测,引物为:
AhFATB2S:5′-CGCGGATCCATGGTTGCTACTGCTGCTACG-3′;
AhFATB2A:5′-GACGAGCTCTCAGTTTTCTGCTGGAAAAACC-3′;
PCR反应体系如下:基因组DNA模板1μL(约50ng)、10×Taq酶反应缓冲液2ul、25mMMgCL
反应程序为:94℃变性5min,94℃45s、55℃45s、72℃2min 32循环,72℃延伸5min。
用1%琼脂糖凝胶电泳检测PCR反应产物,结果表明该AhFATB2基因表达载体pBI-AhFATB2已经成功转入拟南芥获得了转基因阳性苗(图3)。将阳性苗自交3代以上,获得纯合植株HF-AhFATB2。
实施例4:
HF-AhFATB2莲座叶数目与棕榈酸含量测定结果:
把获得的纯合植株HF-AhFATB2种子(转化子1-10)播种于蛭石中,用塑料布蒙上,4℃春化后在生长箱中萌发。长出2-3片真叶后间苗,使得行距及株距均为3厘米,蛭石每两周浇一次营养液。同时播种野生型植株1-3作为对比。
约20天左右,拟南芥植株出现花序后,统计莲座叶数目,HF-AhFATB2莲座叶数目在19-21片之间,野生型拟南芥莲座叶数目在12-13片之间,叶片大小没有明显变化,其生物量明显增加(图4,表2)。
拟南芥种植成熟后,收集并进行脂肪酸组分分析,种子中的棕榈酸含量由野生型的7.8%增加到19.2-28.8%,增加了146-269%(表3)。从脂肪酸组分分析可以看出(图5):转基因植株种子的脂肪酸气相色谱中代表棕榈酸含量的峰(对应数值分别是12.882,12.893,12.881)的面积明显变大,峰面积从野生型的7.85%(12.881)变为转基因植株1种子的22.49%(12.882)、转基因植株2种子的28.65%(12.893)。结果表明,通过该方法获得的转基因植株的叶片数目和棕榈酸含量明显增加。
表2 HF-AhFATB2转化子莲座叶数目统计表
表3 HF-AhFATB2转化子棕榈酸含量统计表
因此,花生的AhFATB2基因可以应用在植物基因工程和油料作物种子生产以及蔬菜、烟草等作物的叶片生成中,具备一定应用价值。通过人工构建AhFATB2过表达载体,或其它基因工程、分子生物学手段,增加或强化AhFATB2基因的表达,人工创建高棕榈酸及高叶片生物量的植物用于商业生产中。如在油菜、花生、大豆等油料作物中过表达AhFATB2,创制高棕榈酸的油菜、花生、大豆等油料作物,用于商业生成棕榈酸;在烟草、白菜、菠菜等以收获叶片为目的的作物中过表达AhFATB2创制多叶材料等。
序列表
<110> 河南省农业科学院
<120> 一种创建高棕榈酸和多叶片转基因植株的方法及应用
<130> 植物基因工程
<160> 3
<170> SIPOSequenceListing 1.0
<210> 1
<211> 30
<212> DNA
<213> 人工序列()
<400> 1
cgcggatcca tggttgctac tgctgctacg 30
<210> 2
<211> 31
<212> DNA
<213> 人工序列()
<400> 2
gacgagctct cagttttctg ctggaaaaac c 31
<210> 3
<211> 1433
<212> DNA
<213> 人工序列()
<400> 3
aggtgaagaa ttcaattatg gttgctactg ctgctacgtc gtcgtttttc cctgtgacgt 60
cacgaaccgg tggaggagga ggaggaggaa tccctgccag cctcggcgga gggctcaaac 120
aaaatcacag gtcttcaagt gttaaggcca atgcgcatgc tccttcaaag atcaacggaa 180
ccgccacaaa ggttccaaaa tccatggaga gcatgaagct ggaatcctcg tcgacgaccg 240
gggctaatgc gccgaggact ttcattaacc agattccgga ttggagcatg ctgctggccg 300
ccatcacgac agccttcctt gcggcggaga agcagtggat gatgatcgat tggaagccga 360
agcgatccga tgtgctatct gatccatttg gtattgggag gattgtgcag gatgggcttg 420
ctttcaggca aaatttctcc attcgatctt acgagatggg cgccgataag accgcgtcta 480
tagagacgct aatgaatcat ttgcaggaaa ctgcacttaa tcatgttaag actgctgggc 540
ttcttggtga tggctttggt tcgacgccag aaatgtgtaa gaagaacctg atatgggttg 600
tgactcggat gcaagttgaa gttgatcgtt acccaacatg gggagatgta gttcaagttg 660
acacttgggt ttctgcatca gggaaaaatg gtatgcgtcg tgattggatc atacgtgacg 720
ccaatacggg tgaaatcttg acaagagcct ccagtatttg ggtcatgatg aataaagtga 780
caaggagact atccaaaatt ccagaagaag tcaggcaaga gattgcgtca tattttgtgg 840
attctcctcc agttgtcgaa gaggataaca gaaaactgtc aaaacttgat gatactgcag 900
atcatattcg tcgtggtcta agtcctagat ggagtgatct agatgttaat cagcatgtta 960
acaatgtgaa gtacattggc tggcttctgg agagtgctcc acaggcaatc ttggagagtc 1020
atgagtacag gagggagtgt ggcaaggaca gtgtgctgga ttccctaacc gatgtctctg 1080
gtgctgatat cgggaactta gctggcggcg gatctctcga gtgcaaacac ttgcttaggc 1140
ttgaagatgg tggtgagatt gtgaggggta ggactgaatg gaggcccaag cctgtgaaca 1200
actttggtgc tatgaatcag gtttttccag cagaaaactg aagcattgag tgcctaaaat 1260
gaaattgctg ggaatgtcaa tctccattat actttacaaa ttggatctga ccaatggaga 1320
gttttgcttg ttgttatcac cgccaccatc tccaccacca ccaccaccat catcatgtct 1380
acctatctta tataaatata tgcatatcta attcgtgtta atatggtggg gtt 1433
- 一种创建高棕榈酸和多叶片转基因植株的方法及应用
- 一种农杆菌介导的叶片转化培育柑橘转基因植株的方法