用于靶向遗传修饰的方法和组合物,以及这些组合物的使用方法
文献发布时间:2023-06-19 12:24:27
分案申请说明
本申请是申请日为2015年06月26日,申请号为201580046401.3,发明名称为“用于靶向遗传修饰的方法和组合物,以及这些组合物的使用方法”的发明专利申请的分案申请。
相关申请的交叉引用
本申请要求2014年6月26日提交的美国临时申请No.62/017,582和2014年6月26日提交的美国临时申请No.62/017,627的权益,这两份临时申请均据此全文以引用方式并入本文。
所述序列表的正式文本是经由EFS网络作为ASCII格式序列表以电子方式提交的,文件名为463545SEQLIST.TXT,创建于2015年6月25日,大小为14000字节,并且与本说明书同时提交。该ASCII格式文件中含有的序列表是本说明书的组成部分,其全文以引用方式并入本文。
技术领域
本发明涉及用于维持或培养多能和/或全能细胞的方法与组合物,以及用于产生细胞群体和转基因动物的方法与组合物。
背景技术
也许是由于Y染色体具有独特的结构特征,所以在小鼠胚胎干细胞中产生Y连锁基因突变的常规基因靶向策略已获得的成果有限。因此通常情况下,对鼠Y连锁基因功能的理解受限于从对携带自发缺失、随机基因捕获插入或常染色体转基因的小鼠的研究获得的见解。需要开发用于提高靶向Y染色体上的基因组座位的能力的方法。
Sry蛋白(性别决定区Y)是胎盘哺乳动物中决定雄性性别的关键调控因子。Sry基因(也称为睾丸决定因子(TDF))驻留在Y染色体上。Sry被认为是通过其高迁移率族(HMG)结构域结合DNA的转录因子。小鼠Sry基因的表达在胚胎发育第11天左右的狭窄时间窗内局限于生殖嵴;既检测了Sry mRNA,又检测了Sry蛋白。必须在该时间窗内制备足够多的Sry,以使双潜能生殖嵴朝着雄性睾丸形成程序转变,同时抑制雌性卵巢发育程序。在成体睾丸中,检测到了环状Sry转录物,但未检测到Sry蛋白。Sry基因中导致产生失活Sry蛋白或改变基因表达的定时和强度的突变可引起雄性到雌性的性逆转,从而产生具有X和Y染色体但在解剖学上为雌性的动物。所谓的XY雌性通常是不育的,或只具有很低的可育性。能够通过调控Sry来控制性别决定,在产生经遗传修饰动物的过程中将具有极大的价值。
发明内容
本发明提供了一种用于制备能够在F0代中产生可育XY雌性非人哺乳动物的XY胚胎干(ES)细胞系的方法。该方法包括:(a)修饰非人哺乳动物XY胚胎干(ES)细胞,以具有降低Sry蛋白的水平和/或活性的修饰;以及(b)在一定条件下培养所述经修饰的ES细胞系,所述条件允许制备能够在F0代中产生可育XY雌性非人哺乳动物的ES细胞系。
还提供了一种用于在F0代中产生可育XY雌性非人哺乳动物的方法。该方法包括:(a)将通过上述方法制备的具有降低Sry蛋白水平和/或活性的修饰的非人哺乳动物XY ES细胞引入宿主胚胎中;(b)孕育该宿主胚胎;以及(c)获得F0 XY雌性非人哺乳动物,其中在达到性成熟后,该F0 XY雌性非人哺乳动物是可育的。在一个实施例中,所述雌性XY F0非人哺乳动物在与野生型小鼠杂交时是可育的。在具体实施例中,所述野生型小鼠为C57BL/6。
在一个实施例中,所述非人哺乳动物XY ES细胞来自啮齿动物。在一个具体实施例中,所述啮齿动物为小鼠。在一个实施例中,小鼠XY ES细胞来源于129品系。在一个实施例中,小鼠XY ES细胞为VGF1小鼠ES细胞。在一个实施例中,小鼠XY ES细胞包含来源于129品系的Y染色体。在一个实施例中,小鼠XY ES细胞来自C57BL/6品系。在另一个实施例中,啮齿动物为大鼠或仓鼠。
在一些实施例中,Sry蛋白的水平和/或活性降低由Sry基因中的遗传修饰引起。在一些此类方法中,Sry基因中的遗传修饰包括一个或多个核苷酸的插入、一个或多个核苷酸的缺失、一个或多个核苷酸的置换、敲除、敲入,用同源、异源或直系同源核酸序列替换内源核酸序列,或它们的组合。
在本文提供的方法中,靶向遗传修饰可包括插入、缺失、敲除、敲入、点突变或它们的组合。在另一个实施例中,靶向遗传修饰位于常染色体上。
在一些实施例中,Sry基因的修饰包括可选标记和/或报告基因的插入,所述可选标记和/或报告基因有效连接至所述非人哺乳动物ES细胞中有活性的启动子。在一些实施例中,Sry基因的修饰包括报告基因的插入,所述报告基因有效连接至内源Sry启动子。在一个具体实施例中,报告基因编码报告蛋白LacZ。
在一个实施例中,培养步骤包括在适用于在培养中维持非人哺乳动物ES细胞、包含基础培养基和补充物的培养基中,培养非人哺乳动物XY ES细胞,其中所述培养基为低渗透压培养基。在一个实施例中,所述低渗透压培养基表现出约200mOsm/kg至小于约329mOsm/kg的渗透压。在其他实施例中,所述低渗透压培养基表现出以下一种或多种特征:约11mS/cm至约13mS/cm的电导率;约50mM至约110mM的碱金属卤化物盐浓度;约17mM至约30mM的碳酸盐浓度;约85mM至约130mM的碱金属卤化物盐和碳酸盐总浓度;和/或这些特征之中任何两种或更多种的组合。
在一些实施例中,在将所述非人哺乳动物XY ES细胞引入宿主胚胎中并接着孕育该宿主胚胎之后,至少80%、至少85%、至少90%或至少95%的F0非人哺乳动物为XY雌性,在达到性成熟后,所述F0 XY雌性非人哺乳动物是可育的。
在一个实施例中,所述非人哺乳动物XY ES细胞在Y染色体上包含靶基因组座位,所述靶基因组座位包含核酸酶试剂的识别位点,并且其中所述核酸酶试剂在所述识别位点处诱导切口或双链断裂。这种方法还可包括在存在包含插入多核苷酸的靶向载体的情况下使所述ES细胞暴露于所述核酸酶试剂,其中在暴露于所述核酸酶试剂和所述靶向载体后,所述ES细胞被修饰成包含所述插入多核苷酸。在一个实施例中,所述核酸酶试剂为编码核酸酶的mRNA。在具体实施例中,所述核酸酶试剂为(a)锌指核酸酶(ZFN);(b)转录激活因子样效应物核酸酶(TALEN);或(c)大范围核酸酶。在其他实施例中,所述核酸酶试剂包括成簇规律间隔短回文重复序列(CRISPR)相关(Cas)蛋白和向导RNA(gRNA)。在此类方法中,所述向导RNA(gRNA)包括(a)靶向第一识别位点的成簇规律间隔短回文重复序列(CRISPR)RNA(crRNA);和(b)反式激活CRISPR RNA(tracrRNA)。在一些情况下,所述识别位点紧邻地侧接前间区序列邻近基序(PAM)序列。在一个实施例中,Cas蛋白为Cas9。
还提供了一种体外培养物,其包含根据本文所提供的任何方法的非人哺乳动物XYES细胞系。
提供了一种体外培养物,并且该体外培养物包含(a)具有降低Sry蛋白水平和/或活性的修饰的非人哺乳动物XY胚胎干(ES)细胞;以及(b)适用于在培养中维持非人哺乳动物ES细胞、包含基础培养基和补充物的培养基。在一个实施例中,所述基础培养基表现出约200mOsm/kg至小于约329mOsm/kg的渗透压。在其他实施例中,所述基础培养基表现出以下一种或多种特征:约11mS/cm至约13mS/cm的电导率;约50mM至约110mM的碱金属卤化物盐浓度;约17mM至约30mM的碳酸盐浓度;约85mM至约130mM的碱金属卤化物盐和碳酸盐总浓度;和/或这些特征之中任何两种或更多种的组合。在一个实施例中,所述非人哺乳动物XY ES细胞来自啮齿动物。在一个实施例中,啮齿动物为小鼠或大鼠。在一个实施例中,小鼠XY ES细胞为VGF1小鼠ES细胞。在一个实施例中,啮齿动物为大鼠或仓鼠。在一个实施例中,Sry蛋白的水平和/或活性降低源自Sry基因中的遗传修饰。在一个实施例中,Sry基因中的遗传修饰包括一个或多个核苷酸的插入、一个或多个核苷酸的缺失、一个或多个核苷酸的置换、敲除、敲入、用异源核酸序列替换内源核酸序列,或它们的组合。在一个实施例中,所述非人哺乳动物ES细胞包含一种、两种、三种或更多种靶向遗传修饰。在一个实施例中,靶向遗传修饰包括插入、缺失、敲除、敲入、点突变或它们的组合。在一个实施例中,靶向遗传修饰包括异源多核苷酸在所述XY ES细胞的基因组中的至少一个插入。在一个实施例中,靶向遗传修饰位于常染色体上。在一个实施例中,所述基础培养基表现出50±5mM NaCl、26±5mM碳酸盐以及218±22mOsm/kg。在一个实施例中,所述基础培养基表现出约3mg/mL NaCl、2.2mg/mL碳酸氢钠以及218mOsm/kg。在一个实施例中,所述基础培养基表现出87±5mM NaCl、18±5mM碳酸盐以及261±26mOsm/kg。在一个实施例中,所述基础培养基表现出约5.1mg/mLNaCl、1.5mg/mL碳酸氢钠以及261mOsm/kg。在一个实施例中,所述基础培养基表现出110±5mM NaCl、18±5mM碳酸盐以及294±29mOsm/kg。在一个实施例中,所述基础培养基表现出约6.4mg/mL NaCl、1.5mg/mL碳酸氢钠以及294mOsm/kg。在一个实施例中,所述基础培养基表现出87±5mM NaCl、26±5mM碳酸盐以及270±27mOsm/kg。在一个实施例中,所述基础培养基表现出约5.1mg/mL NaCl、2.2mg/mL碳酸氢钠以及270mOsm/kg。在一个实施例中,所述基础培养基表现出87±5mM NaCl、26±5mM碳酸盐、86±5mM葡萄糖以及322±32mOsm/kg。在一个实施例中,所述基础培养基表现出约5.1mg/mL NaCl、2.2mg/mL碳酸氢钠、15.5mg/mL葡萄糖以及322mOsm/kg。在一个实施例中,在将所述非人哺乳动物XY ES细胞引入宿主胚胎中并接着孕育该宿主胚胎之后,至少80%的所述F0非人哺乳动物为XY雌性,在达到性成熟后,所述F0 XY雌性非人哺乳动物是可育的。
还提供了一种用于在F0代中产生可育雌性XY非人哺乳动物的方法,包括:(a)在适用于在培养中维持非人哺乳动物ES细胞、包含基础培养基和补充物的培养基中培养具有降低Sry蛋白水平和/或活性的修饰的供体非人哺乳动物XY胚胎干(ES)细胞,(b)将所述供体XY非人哺乳动物ES细胞引入宿主胚胎中;(c)孕育该宿主胚胎;以及(d)获得F0 XY雌性非人哺乳动物,其中在达到性成熟后,所述F0 XY雌性非人哺乳动物是可育的。在一个实施例中,所述培养基表现出约200mOsm/kg至小于约329mOsm/kg的渗透压。在其他实施例中,所述培养基表现出包括以下一者或多者的特征:约11mS/cm至约13mS/cm的电导率;约50mM至约110mM的碱金属卤化物盐浓度;约17mM至约30mM的碳酸盐浓度;约85mM至约130mM的碱金属卤化物盐和碳酸盐总浓度;和/或这些特征之中任何两种或更多种的组合。在一个实施例中,所述非人哺乳动物XY ES细胞来自啮齿动物。
在一个实施例中,啮齿动物为小鼠或大鼠。在一个实施例中,小鼠XY ES细胞为VGF1小鼠ES细胞。在一个实施例中,啮齿动物为大鼠或仓鼠。在一个实施例中,Sry蛋白的水平和/或活性降低源自Sry基因中的遗传修饰。
在一个实施例中,Sry基因中的遗传修饰包括一个或多个核苷酸的插入、一个或多个核苷酸的缺失、一个或多个核苷酸的置换、敲除、敲入、用异源核酸序列替换内源核酸序列,或它们的组合。在一个实施例中,所述非人哺乳动物ES细胞包含一种、两种、三种或更多种靶向遗传修饰。在一个实施例中,靶向遗传修饰包括插入、缺失、敲除、敲入、点突变或它们的组合。在一个实施例中,靶向遗传修饰包括异源多核苷酸在所述XY ES细胞的基因组中的至少一个插入。在一个实施例中,靶向遗传修饰位于常染色体上。在一个实施例中,所述基础培养基表现出50±5mM NaCl、26±5mM碳酸盐以及218±22mOsm/kg。在一个实施例中,所述基础培养基表现出约3mg/mL NaCl、2.2mg/mL碳酸氢钠以及218mOsm/kg。在一个实施例中,所述基础培养基表现出87±5mM NaCl、18±5mM碳酸盐以及261±26mOsm/kg。在一个实施例中,所述基础培养基表现出约5.1mg/mL NaCl、1.5mg/mL碳酸氢钠以及261mOsm/kg。在一个实施例中,所述基础培养基表现出110±5mM NaCl、18±5mM碳酸盐以及294±29mOsm/kg。在一个实施例中,所述基础培养基表现出约6.4mg/mL NaCl、1.5mg/mL碳酸氢钠以及294mOsm/kg。在一个实施例中,所述基础培养基表现出87±5mM NaCl、26±5mM碳酸盐以及270±27mOsm/kg。在一个实施例中,所述基础培养基表现出约5.1mg/mL NaCl、2.2mg/mL碳酸氢钠以及270mOsm/kg。在一个实施例中,其中所述基础培养基表现出87±5mM NaCl、26±5mM碳酸盐、86±5mM葡萄糖以及322±32mOsm/kg。在一个实施例中,其中所述基础培养基表现出约5.1mg/mL NaCl、2.2mg/mL碳酸氢钠、15.5mg/mL葡萄糖以及322mOsm/kg。
还提供了在F1代中产生对于靶向遗传突变为纯合的转基因非人哺乳动物的方法,包括:(a)使所述Sry蛋白的水平和/或活性降低的F0 XY可育雌性与衍生自相同ES细胞克隆的同龄群无性系同胞F0 XY雄性非人哺乳动物杂交,其中所述F0 XY可育雌性非人哺乳动物和所述F0 XY雄性非人哺乳动物各自对于所述遗传突变均为杂合的;以及(b)获得对于所述遗传修饰为纯合的F1子代小鼠。
还提供了一种用于修饰细胞中Y染色体上的靶基因组座位的方法,包括:(a)提供在Y染色体上包含靶基因组座位的细胞,所述靶基因组座位包含核酸酶试剂的识别位点,(b)向该细胞中引入(i)核酸酶试剂,其中所述核酸酶试剂在第一识别位点处诱导切口或双链断裂;和(ii)包含第一插入多核苷酸的第一靶向载体,所述第一插入多核苷酸侧接第一同源臂和第二同源臂,所述第一同源臂和第二同源臂与位置足够接近第一识别位点的第一靶位点和第二靶位点相对应;以及(c)鉴定在其基因组中包含在所述靶基因组座位处整合的第一插入多核苷酸的至少一个细胞。在一个实施例中,第一同源臂和第二同源臂的总和为至少4kb但少于150kb。在一个实施例中,第一同源臂和/或第二同源臂的长度为至少400bp但少于1000bp。在另一个实施例中,第一同源臂和/或第二同源臂的长度为约700bp至约800bp。
还提供了一种用于修饰细胞中Y染色体上的靶基因组座位的方法,包括:(a)提供在Y染色体上包含靶基因组座位的细胞,所述靶基因组座位包含核酸酶试剂的识别位点;(b)向该细胞中引入包含第一插入多核苷酸的第一靶向载体,所述第一插入多核苷酸侧接与第一靶位点和第二靶位点对应的第一同源臂和第二同源臂;以及(c)鉴定在其基因组中包含在所述靶基因组座位处整合的第一插入多核苷酸的至少一个细胞。在一个实施例中,第一同源臂和/或第二同源臂的长度为至少400bp但少于1000bp。在另一个实施例中,第一同源臂和/或第二同源臂的长度为约700bp至约800bp。在一个实施例中,所述细胞为哺乳动物细胞。在一个实施例中,所述哺乳动物细胞为非人细胞。在一个实施例中,所述哺乳动物细胞来自啮齿动物。在一个实施例中,啮齿动物为大鼠、小鼠或仓鼠。在一个实施例中,所述细胞为多能细胞。在一个实施例中,所述哺乳动物细胞为诱导性多能干(iPS)细胞。在一个实施例中,所述多能细胞为非人胚胎干(ES)细胞。在一个实施例中,所述多能细胞为啮齿动物胚胎干(ES)细胞、小鼠胚胎干(ES)细胞或大鼠胚胎干(ES)细胞。在一个实施例中,所述核酸酶试剂为编码核酸酶的mRNA。在一个实施例中,所述核酸酶试剂为锌指核酸酶(ZFN)。在一个实施例中,所述核酸酶试剂为转录激活因子样效应物核酸酶(TALEN)。在一个实施例中,所述核酸酶试剂为大范围核酸酶。在一些实施例中,所述核酸酶试剂包括成簇规律间隔短回文重复序列(CRISPR)相关(Cas)蛋白和向导RNA(gRNA)。在这种方法中,所述向导RNA(gRNA)可包括(a)靶向第一识别位点的成簇规律间隔短回文重复序列(CRISPR)RNA(crRNA);和(b)反式激活CRISPR RNA(tracrRNA)。在一个实施例中,第一识别位点或第二识别位点紧邻地侧接前间区序列邻近基序(PAM)序列。在一些实施例中,Cas蛋白为Cas9。
在一些实施例中,所述修饰包括内源核酸序列的缺失。在一些实施例中,所述缺失在约5kb至约10kb、约10kb至约20kb、约20kb至约40kb、约40kb至约60kb、约60kb至约80kb、约80kb至约100kb、约100kb至约150kb、或约150kb至约200kb、约200kb至约300kb、约300kb至约400kb、约400kb至约500kb、约500kb至约1Mb、约1Mb至约1.5Mb、约1.5Mb至约2Mb、约2Mb至约2.5Mb、或约2.5Mb至约3Mb的范围内。在一个具体实施例中,所述缺失为至少500kb。在一个实施例中,所述细胞为哺乳动物细胞。在一个实施例中,所述哺乳动物细胞为非人细胞。在一个实施例中,所述哺乳动物细胞来自啮齿动物。在一个实施例中,啮齿动物为大鼠、小鼠或仓鼠。在一个实施例中,所述细胞为多能细胞。在一个实施例中,所述哺乳动物细胞为诱导性多能干(iPS)细胞。在一个实施例中,所述多能细胞为非人胚胎干(ES)细胞。在一个实施例中,所述多能细胞为啮齿动物胚胎干(ES)细胞、小鼠胚胎干(ES)细胞或大鼠胚胎干(ES)细胞。在一些实施例中,所述核酸酶试剂包括成簇规律间隔短回文重复序列(CRISPR)相关(Cas)蛋白和向导RNA(gRNA)。在这种方法中,所述向导RNA(gRNA)可包括(a)靶向第一识别位点的成簇规律间隔短回文重复序列(CRISPR)RNA(crRNA);和(b)反式激活CRISPR RNA(tracrRNA)。在一个实施例中,第一识别位点或第二识别位点紧邻地侧接前间区序列邻近基序(PAM)序列。在一些实施例中,Cas蛋白为Cas9。在一个实施例中,所述核酸酶试剂为锌指核酸酶(ZFN)。在一个实施例中,所述核酸酶试剂为转录激活因子样效应物核酸酶(TALEN)。在一个实施例中,所述核酸酶试剂为大范围核酸酶。
用于修饰Y染色体的方法包括在存在包含至少10kb核酸序列的大靶向载体(LTVEC)的情况下使Y染色体暴露于Cas蛋白和CRISPR RNA,并且包括在暴露于Cas蛋白、CRISPR RNA和LTVEC后,将Y染色体修饰成包含至少10kb的核酸序列。LTVEC可包含至少20kb、至少30kb、至少40kb、至少50kb、至少60kb、至少70kb、至少80kb或至少90kb的核酸序列。在其他实施例中,LTVEC包含至少100kb、至少150kb或至少200kb的核酸序列。
还提供了一种用于修饰Y染色体上的靶基因组座位的方法,包括:(a)提供在Y染色体上包含靶基因组座位的哺乳动物细胞,其中所述靶基因组座位包含向导RNA(gRNA)靶序列;(b)向该哺乳动物细胞中引入:(i)包含第一核酸的大靶向载体(LTVEC),所述第一核酸侧接有与靶基因组座位同源的靶向臂,其中所述LTVEC为至少10kb;(ii)包含第一启动子的第一表达构建体,所述第一启动子有效连接至编码Cas蛋白的第二核酸,以及(iii)包含第二启动子的第二表达构建体,所述第二启动子有效连接至编码向导RNA(gRNA)的第三核酸,所述向导RNA包含杂交至gRNA靶序列和反式激活CRISPR RNA(tracrRNA)的核苷酸序列,其中所述第一启动子和所述第二启动子在该哺乳动物细胞中有活性;以及(c)鉴定在Y染色体上的靶基因组座位处包含靶向遗传修饰的经修饰的哺乳动物细胞。在其他实施例中,LTVEC为至少15kb、至少20kb、至少30kb、至少40kb、至少50kb、至少60kb、至少70kb、至少80kb或至少90kb。在其他实施例中,LTVEC为至少100kb、至少150kb或至少200kb。在一个实施例中,所述哺乳动物细胞为非人哺乳动物细胞。在一个实施例中,所述哺乳动物细胞为成纤维细胞。在一个实施例中,所述哺乳动物细胞来自啮齿动物。在一个实施例中,啮齿动物为大鼠、小鼠或仓鼠。在一个实施例中,所述哺乳动物细胞为多能细胞。在一个实施例中,所述多能细胞为诱导性多能干(iPS)细胞。在一个实施例中,所述多能细胞为小鼠胚胎干(ES)细胞或大鼠胚胎干(ES)细胞。在一个实施例中,所述多能细胞为发育受限的人祖细胞。在一个实施例中,Cas蛋白为Cas9蛋白。在一个实施例中,gRNA靶序列紧邻地侧接前间区序列邻近基序(PAM)序列。在一个实施例中,LTVEC的5’同源臂和3’同源臂的总和为约10kb至约150kb。在一个实施例中,LTVEC的5’同源臂和3’同源臂的总和为约10kb至约20kb、约20kb至约40kb、约40kb至约60kb、约60kb至约80kb、约80kb至约100kb、约100kb至约120kb、或约120kb至150kb。在一个实施例中,靶向遗传修饰包括:(a)用同源或直系同源核酸序列替换内源核酸序列;(b)内源核酸序列的缺失;(c)内源核酸序列的缺失,其中所述缺失在约5kb至约10kb、约10kb至约20kb、约20kb至约40kb、约40kb至约60kb、约60kb至约80kb、约80kb至约100kb、约100kb至约150kb、或约150kb至约200kb、约200kb至约300kb、约300kb至约400kb、约400kb至约500kb、约500kb至约1Mb、约1Mb至约1.5Mb、约1.5Mb至约2Mb、约2Mb至约2.5Mb、或约2.5Mb至约3Mb的范围内;(d)外源核酸序列的插入;(e)外源核酸序列的插入,所述插入在约5kb至约10kb、约10kb至约20kb、约20kb至约40kb、约40kb至约60kb、约60kb至约80kb、约80kb至约100kb、约100kb至约150kb、约150kb至约200kb、约200kb至约250kb、约250kb至约300kb、约300kb至约350kb、或约350kb至约400kb的范围内;(f)包含同源或直系同源核酸序列的外源核酸序列的插入;(g)包含人和非人核酸序列的嵌合核酸序列的插入;(h)侧接有位点特异性重组酶靶序列的条件性等位基因的插入;(i)有效连接至所述哺乳动物细胞中有活性的第三启动子的可选标记或报告基因的插入;或(j)它们的组合。在一个实施例中,所述靶基因组座位包含(i)与5’同源臂同源的5’靶序列,以及(ii)与3’同源臂同源的3’靶序列。在一个实施例中,所述5’靶序列和所述3’靶序列相隔至少5kb但少于3Mb。在一个实施例中,所述5’靶序列和所述3’靶序列相隔至少5kb但少于10kb、至少10kb但少于20kb、至少20kb但少于40kb、至少40kb但少于60kb、至少60kb但少于80kb、至少约80kb但少于100kb、至少100kb但少于150kb、或至少150kb但少于200kb、至少约200kb但少于约300kb、至少约300kb但少于约400kb、至少约400kb但少于约500kb、至少约500kb但少于约1Mb、至少约1Mb但少于约1.5Mb、至少约1.5Mb但少于约2Mb、至少约2Mb但少于约2.5Mb、或至少约2.5Mb但少于约3Mb。在一个实施例中,第一表达构建体和第二表达构建体位于单个核酸分子上。在一个实施例中,所述靶基因组座位包括Sry基因座。
还提供了一种用于非人动物Y染色体上的靶向遗传修饰的方法,包括:(a)根据本文所述的方法修饰非人多能细胞的Y染色体上的目标基因组座位,从而产生在Y染色体上包含靶向遗传修饰的经遗传修饰的非人多能细胞;(b)将(a)的所述经修饰的非人多能细胞引入非人宿主胚胎中;以及在代孕母体中孕育包含所述经修饰的多能细胞的非人宿主胚胎,其中所述代孕母体产生包含靶向遗传修饰的F0子代,其中所述靶向遗传修饰能够通过种系传递。在一个实施例中,所述目标基因组座位包括Sry基因座。
本发明提供了用于在Y染色体上生成靶向遗传修饰的方法与组合物。组合物包括体外培养物,所述体外培养物包含具有降低Sry蛋白水平和/或活性的修饰的XY多能和/或全能动物细胞(即,XY ES细胞或XY iPS细胞);以及在促进XY F0可育雌性发育的培养基中培养这些细胞。此类组合物可应用于在F0代中产生可育雌性XY非人哺乳动物的各种方法。
附图说明
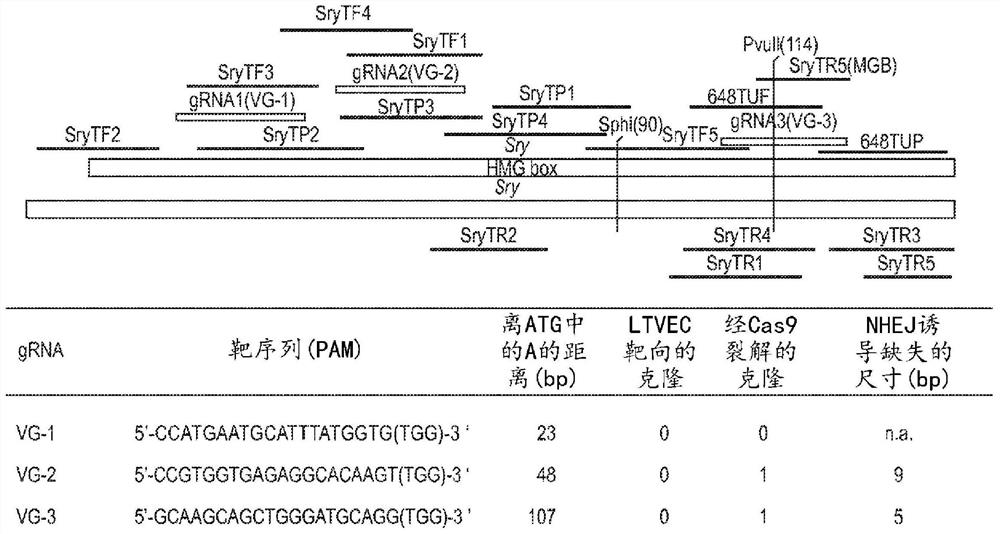
图1提供了靶向小鼠Sry基因的CRISPR Cas9/gRNA的示意图。VG-1(SEQ ID NO:10);VG-2(SEQ ID NO:11);VG-3(SEQ ID NO:12)。图1所示的引物和探针以SEQ ID NO:13至29提供。
图2提供了使用lacZ报告基因,用TALEN和CRISPR靶向Sry基因的示意图。用LTVEC和短臂载体(smallTVEC)两者靶向Sry基因,以避免攻击Y染色体上的基因座,所述短臂载体的同源臂比LTVEC小。
图3示出了胚胎中的LacZ表达。
图4提供了由ZFN或由CRISPR向导RNA结合Cas9 DNA内切核酸酶介导的在Y染色体上产生大于500kb的大段缺失的示意图。
图5A、图5B和图5C提供了对各种克隆中Y染色体的大段缺失的测序确认。图5A为克隆1-D5的测序结果。Kdm5上游和Uspy9下游序列以SEQ ID NO:30、1-D5 1500F(SEQ ID NO:31)、1-D5 1000R(SEQ ID NO:32)提供;图5B为克隆5-C4的测序结果。Kdm5上游和Uspy9下游序列以SEQ ID NO:33、1500F(SEQ ID NO:34)、1000R(SEQ ID NO:35)、1000F(SEQ ID NO:36)提供;图5C为克隆6-A12的测序结果。Kdm5上游和Uspy9下游序列以SEQ ID NO:37、1500F(SEQ ID NO:38)、1000R(SEQ ID NO:39)、1000F(SEQ ID NO:40)、1500R(SEQ ID NO:41)提供。图5B和图5C中的加框区域代表微同源区。
具体实施方式
在本文中可互换使用的术语“蛋白质”、“多肽”和“肽”包括任何长度的氨基酸聚合形式,包括编码和非编码的氨基酸、以及经化学或生物化学修饰的或衍生的氨基酸。这些术语还包括已经过修饰的聚合物,诸如具有经修饰的肽骨架的多肽。
在本文中可互换使用的术语“核酸”和“多核苷酸”包括任何长度的核苷酸聚合形式,包括核糖核苷酸、脱氧核糖核苷酸,或它们的类似物或修饰形式。这些术语包括单链、双链和多链的DNA或RNA,基因组DNA、cDNA、DNA-RNA杂合体,以及包含嘌呤碱基、嘧啶碱基或其他天然的、经化学修饰的、经生物化学修饰的、非天然的或衍生的核苷酸碱基的聚合物。为了简单起见,无论核酸是双链形式还是单链形式,核酸大小都可用bp来表示,如果核酸为单链形式,则bp是只在单链核酸与其完全互补链形成双链体时形成的那些。
“密码子优化”通常包括在维持天然氨基酸序列的同时,通过用在宿主细胞的基因中更频繁或最频繁使用的密码子替换天然序列的至少一个密码子来修饰核酸序列以增强特定宿主细胞中的表达的过程。例如,可修饰编码Cas蛋白的核酸,以替换成与天然存在的核酸序列相比在给定的原核或真核细胞(包括细菌细胞、酵母细胞、人细胞、非人细胞、哺乳动物细胞、啮齿动物细胞、小鼠细胞、大鼠细胞、仓鼠细胞或任何其他宿主细胞)中具有更高使用频率的密码子。密码子使用表可轻易获得,例如在“密码子使用数据库”处获得。这些表可按多种方式进行调整。参见Nakamura et al.(2000)Nucleic Acids Research 28:292(Nakamura等人,2000年,《核酸研究》,第28卷,第292页)。用于对在特定宿主中表达的特定序列进行密码子优化的计算机算法也是可得的(参见例如Gene Forge)。
“有效连接”或为“有效地连接的”包括两种或更多种组分(例如,启动子和另一个序列元件)的并置关系,使得两种组分都发挥正常功能并允许至少一种所述组分能够介导施加于至少一种其他组分的功能。例如,如果启动子响应于存在或不存在一个或多个转录调控因子而控制编码序列的转录水平,则该启动子可有效连接至该编码序列。
核酸的“互补性”是指核酸的一条链中的核苷酸序列由于其核碱基基团的取向,而与相对核酸链上的另一个序列形成氢键。DNA中的互补碱基通常为A与T、C与G。RNA中的互补碱基通常为C与G、U与A。互补性可以是完美的或显著的/充分的。两个核酸之间完美互补意味着这两个核酸可形成双链体,该双链体中的每个碱基通过沃森-克里克(Watson-Crick)配对而与互补碱基结合。“显著”或“充分”互补是指一条链中的序列与相对链中的序列并非完全和/或完美互补,而是在两条链上的碱基之间发生充分结合,从而在一组杂交条件(例如,盐浓度和温度)下形成稳定的杂交复合体。此类条件可通过以下方式预测:使用序列和标准数学计算来预测杂交链的Tm,或通过使用常规方法来凭经验确定Tm。Tm是指使两条核酸链之间形成的杂交复合体群体的50%变性的温度。在低于Tm的温度下,有利于杂交复合体形成,而在高于Tm的温度下,有利于杂交复合体中的链融化或分离。可通过使用例如Tm=81.5+0.41(%G+C)来估计在1M NaCl水溶液中具有已知G+C含量的核酸的Tm,但也可使用其他已知的Tm计算法来考虑核酸的结构特征。
“杂交条件”包括其中一条核酸链通过互补链相互作用和氢键与第二条核酸链结合而产生杂交复合体的累积环境。此类条件包括含有核酸的水溶液或有机溶液的化学组分及其浓度(例如盐、螯合剂、甲酰胺),以及混合物的温度。其他因素(诸如温育时长或反应室尺寸)可能对环境有影响。参见例如Sambrook et al.,Molecular Cloning,A LaboratoryManual,2.sup.nd ed.,pp.1.90-1.91,9.47-9.51,1 1.47-11.57(Cold Spring HarborLaboratory Press,Cold Spring Harbor,N.Y.,1989)(Sambrook等人,《分子克隆实验指南》,第2版增补版,第1.90-1.91页,第9.47-9.51页,第11.47-11.57页,冷泉港实验室出版社,纽约冷泉港,1989年)。
杂交需要两个核酸含有互补序列,但碱基之间可能存在错配。适于两个核酸之间进行杂交的条件取决于核酸长度和互补程度,这些都是本领域熟知的变量。两个核苷酸序列之间的互补程度越大,具有那些序列的核酸的杂合体的解链温度(Tm)值越大。对于具有短片段互补(例如,在35个或更少、30个或更少、25个或更少、22个或更少、20个或更少、或者18个或更少的核苷酸上互补)的核酸之间的杂交,错配位置变得重要(参见Sambrook等人,出处同上,第11.7-11.8页)。通常,可杂交核酸的长度为至少约10个核苷酸。可杂交核酸的示例性最小长度包括至少约15个核苷酸、至少约20个核苷酸、至少约22个核苷酸、至少约25个核苷酸,以及至少约30个核苷酸。此外,温度和洗涤溶液的盐浓度在必要时可根据多个因素进行调节,这些因素诸如互补区长度和互补程度。
多核苷酸序列不必与其可特异性杂交的靶核酸的序列100%互补。此外,多核苷酸可以在一个或多个区段上杂交,使得介入区段或相邻区段不参与杂交事件(例如,环结构或发夹结构)。多核苷酸(例如,gRNA)可与其所靶向的靶核酸序列内的靶区域具有至少70%、至少80%、至少90%、至少95%、至少99%或100%的序列互补性。例如,如果在gRNA的20个核苷酸中有18个与靶区域互补、因而将特异性杂交,则该gRNA代表90%的互补性。在该例子中,剩余的非互补核苷酸可以是成簇的或其间间隔互补核苷酸,不必彼此邻接、也不必与互补核苷酸邻接。
可使用BLAST程序(局部比对基本检索工具)和本领域已知的PowerBLAST程序(Altschul et al.(1990)J.Mol.Biol.215:403-410(Altschul等人,1990年,《分子生物学杂志》,第215卷,第403-410页);Zhang and Madden(1997)Genome Res.7:649-656(Zhang和Madden,1997年,《基因组研究》,第7卷,第649-656页))或通过使用Gap程序(Wisconsin序列分析程序包(Wisconsin Sequence Analysis Package),第8版,用于Unix操作系统,美国威斯康星州麦迪逊大学研究园遗传学计算机小组(Genetics Computer Group,UniversityResearch Park,Madison Wis.)),使用默认设置来常规地测定核酸内的特定核酸序列片段之间的互补性百分比,该测定使用史密斯-沃特曼算法(Adv.Appl.Math.,1981,2,482-489(《应用数学进展》,1981年,第2卷,第482-489页))。
本文所提供的方法与组合物采用多种不同的组分。已经认识到,在说明书通篇中,一些组分可具有活性变体和片段。此类组分包括例如Cas蛋白、CRISPR RNA、tracrRNA和向导RNA。这些组分各自的生物活性在本文别处描述。
在两个多核苷酸或多肽序列的上下文中,“序列同一性”或“同一性”是指在指定的比较窗中比对最大对应时,两个序列中的残基是相同的。在关于蛋白质使用序列同一性百分比时,已经认识到不相同的残基位置通常因保守氨基酸置换而不同,发生保守氨基酸置换时,氨基酸残基被置换成其他具有相似化学性质(例如,电荷或疏水性)的氨基酸残基,因而不改变分子的功能性质。当序列在保守置换方面不同时,可上调序列同一性百分比以校正所述置换的保守性。由于这类保守置换而不同的序列被称为具有“序列相似性”或“相似性”。用于进行这种调节的手段是本领域技术人员所熟知的。通常,这涉及将保守置换当作部分错配而不是完全错配来记分,从而增大序列同一性百分比。因此,例如,在将相同的氨基酸记1分并将非保守置换记0分的情况下,保守置换的得分介于0和1之间。计算保守置换的得分,例如,如在程序PC/GENE(美国加利福尼亚州山景城Intelligenetics公司(Intelligenetics,Mountain View,California))中实现。
“序列同一性百分比”包括通过在比较窗上比较两个最佳比对序列而测定的值,其中与参考序列(其不包含添加和缺失)相比,多核苷酸序列在比较窗中的部分可包含添加或缺失(即缺口)以实现两个序列的最佳比对。通过以下方式计算这种百分比:确定在两个序列中出现相同核酸碱基或氨基酸残基的位置的数目以得到匹配位置的数目,将匹配位置的数目除以比较窗中位置的总数目,然后将结果乘以100,得到序列同一性百分比。
除非另有说明,否则序列同一性/相似性值包括使用GAP版本10并使用以下参数获得的值:对于核苷酸序列的同一性%和相似性%,使用GAP权重50和长度权重3,以及nwsgapdna.cmp评分矩阵;对于氨基酸序列的同一性%和相似性%,使用GAP权重8和长度权重2,以及BLOSUM62评分矩阵;或它们的任何等同程序。“等同程序”包括对于任何两个所考虑的序列,当与由GAP版本10产生的相应比对比较时,产生具有相同的核苷酸或氨基酸残基匹配以及相同的序列同一性百分比的比对的任一种序列比较程序。
“包含”或“包括”一种或多种所述要素的组合物或方法可包括未明确述及的其他要素。例如,“包含”或“包括”一种蛋白质的组合物可含有单独的蛋白质,也可含有蛋白质与其他成分的组合。
指定的值范围包括在该范围内的或限定该范围的所有整数,以及由该范围内的整数限定的所有子范围。
术语“约”涵盖指定值的标准测量误差界限(例如,SEM)内的值,除非从上下文中明确地看出并非如此。
单数形式的冠词“一个”、“一种”和“该”包括多个指代物,除非上下文中明确地另行规定。例如,术语“一种Cas蛋白”或“至少一种Cas蛋白”可包括多种Cas蛋白,包括它们的混合物。
由供体ES细胞和宿主胚胎产生非人动物的方法是已知的。选择具有某些特征的供体ES细胞,这些特征增强细胞迁入宿主胚胎的能力,因此部分地或基本上部分地有助于用供体ES细胞和宿主胚胎形成动物。所形成的动物可以是雄性或雌性,这很大程度上基于ES细胞的基因型(例如,XY或XX)。
大部分用于产生转基因动物的ES细胞系具有雄性XY基因型。由于Y染色体在哺乳动物性别决定中占支配地位,所以,在将XY ES细胞引入宿主胚胎囊胚中并孕育时,这些细胞在第一代(F0)中几乎总是产生表型为雄性的动物,所述表型为雄性的动物为嵌合体,也就是说,含有来源于雄性供体ES细胞的细胞(XY)和来源于宿主胚胎的细胞,所述来源于宿主胚胎的细胞可以是雄性(XY)、也可以是雌性(XX)。XY ES细胞在通过VelociMouse方法引入8-细胞宿主胚胎中并孕育时,可以在第一代(F0)中产生完全来源于XY ES细胞、且表型为雄性的动物。
WO2011/156723提供了这样的方法和组合物:其采用在培养中维持XY供体细胞的培养基,使得在将XY供体细胞引入宿主胚胎并在合适的宿主中孕育之后,在F0群体中产生可育的XY雌性动物。此类组合物可用于产生对于给定的靶向遗传修饰为纯合的F1子代。
本申请提供了使用具有降低Sry蛋白水平和/或活性的修饰的XY供体细胞与促进产生解剖学上正常、可育且多产的XY F0雌性的培养基的组合的方法与组合物。此类方法和组合物允许在F0代中产生可育的雌性XY非人动物。具有降低Sry蛋白的水平和/或活性的修饰的XY ES细胞与本文所述培养基的组合显著增大了F0代中可育雌性XY子代的百分比。用于有效地将雄性性逆转为雌性的方法在家畜行业中很有价值。例如,对于奶牛行业来说,雌性小牛远比雄性小牛有价值。对于家禽也是如此。出于繁育目的,无论是牛、猪还是绵羊,优选地让多头雌性仅与少数几头公牛、公猪或公羊交配繁育。因此,本文所提供的各种方法可用于各种具有重要商业价值的繁育行业。
还提供了用于制备无需在雌性化培养基中培养,就能够在F0代中产生可育XY雌性非人哺乳动物的XY胚胎干(ES)细胞系的方法和组合物。在此类方法中,具有降低Sry蛋白水平和/或活性的修饰的XY ES细胞系可产生能够在缺乏本文别处所提供的雌性化培养基的情况下(例如,通过在基础培养基中培养,所述基础培养基诸如本文别处所描述的DMEM),在F0代中产生可育XY雌性非人哺乳动物的ES细胞系。
本文提供了包含来自动物的各种XY多能和/或全能细胞的各种组合物和方法。如本文所用的术语“多能细胞”,包括具有发育成不止一种分化细胞类型的能力的未分化细胞。此类多能和/或全能XY细胞可以是例如胚胎干(ES)细胞或诱导性多能干(iPS)细胞。如本文所用的术语“胚胎干细胞”或“ES细胞”包括胚胎衍生的全能或多能细胞,所述全能或多能细胞在引入胚胎后能够有助于胚胎发育成任何组织。
与细胞、多能和/或全能细胞、XY细胞、ES细胞、iPS细胞、供体细胞和/或宿主胚胎有关的术语“动物”,包括哺乳动物、鱼类和禽类。哺乳动物包括例如人、非人灵长类动物、猴、猿、猫、狗、马、公牛、鹿、野牛、绵羊、啮齿动物(例如,小鼠、大鼠、仓鼠、豚鼠)、牲畜(例如,牛物种,如奶牛、犍牛等;羊物种,如绵羊、山羊等;以及猪物种,例如小猪和公猪)。禽类包括例如鸡、火鸡、鸵鸟、鹅、鸭等。还包括家养动物和农业动物。与细胞、XY细胞、ES细胞、供体细胞和/或宿主胚胎有关的短语“非人动物”排除人类。
在具体实施例中,多能细胞为人XY ES细胞、人XY iPS细胞、成人XY ES细胞、发育受限的人祖ES细胞、非人XY ES细胞、非人XY iPS细胞、啮齿动物XY ES细胞、啮齿动物XYiPS细胞、小鼠XY ES细胞、小鼠XY iPS细胞、大鼠XY ES细胞、大鼠XY iPS细胞、仓鼠XY ES细胞、仓鼠XY iPS细胞、猴XY ES细胞、猴XY iPS细胞、农业哺乳动物XY ES细胞、农业哺乳动物XY iPS细胞、家养哺乳动物XY ES细胞或家养哺乳动物XY iPS细胞。此外,XY ES细胞或XYiPS细胞可来自近交品系、杂交品系或远交品系。还已经认识到,多能和/或全能XY细胞可具有XYY核型或XXY核型。
小鼠多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)可来自129品系、C57BL/6品系、129与C57BL/6的混合品系、BALB/c品系或Swiss Webster品系。在一个具体实施例中,小鼠多能和/或全能细胞一半来自129品系,另一半来自C57BL/6品系。在一个实施例中,小鼠多能和/或全能细胞为129品系,其中129品系选自下列品系:129P1、129P2、129P3、129X1、129S1(例如,129S1/SV、129S1/Svlm)、129S2、129S4、129S5、129S9/SvEvH、129S6(129/SvEvTac)、129S7、129S8、129T1、129T2。参见例如Festing et al.(1999)Mammalian Genome10:836(Festing等人,1999年,《哺乳动物基因组》,第10卷,第836页)。在一个实施例中,小鼠多能和/或全能细胞为C57BL品系,并且在一个具体实施例中来自C57BL/A、C57BL/An、C57BL/GrFa、C57BL/Kal_wN、C57BL/6、C57BL/6J、C57BL/6ByJ、C57BL/6NJ、C57BL/6NTac、C57BL/10、C57BL/10ScSn、C57BL/10Cr或C57BL/Ola。在一个具体实施例中,小鼠多能和/或全能细胞为上述129品系与上述C57BL/6品系的混合物。在另一个具体实施例中,小鼠多能和/或全能细胞为上述129品系的混合物,或上述BL/6品系的混合物。在一个具体实施例中,所述混合物中的129品系为129S6(129/SvEvTac)品系。在一些实施例中,小鼠XY ES细胞包含来源于129品系的Y染色体。
在另一个实施例中,小鼠XY ES细胞为VGF1小鼠ES细胞。VGF1(也称为F1H4)小鼠ES细胞来源于通过使雌性C57BL/6NTac小鼠与雄性129S6/SvEvTac小鼠杂交而产生的杂交胚胎。因此,VGF1 ES细胞包含来自129S6/SvEvTac小鼠的Y染色体。参见例如Auerbach,W.etal.(2000)Establishment and chimera analysis of 129/SvEv-and C57BL/6-derivedmouse embryonic stem cell lines.Biotechniques 29,1024–1028,1030,1032(Auerbach,W.等人,2000年,“129/SvEv和C57BL/6来源小鼠胚胎干细胞系的建立与嵌合体分析”,《生物技术》,第29卷,第1024–1028页、第1030页、第1032页),该文献全文以引用方式并入本文。
大鼠多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)可来自任何大鼠品系,包括但不限于ACI大鼠品系、Dark Agouti(DA)大鼠品系、Wistar大鼠品系、LEA大鼠品系、Sprague Dawley(SD)大鼠品系或Fischer大鼠品系(诸如Fisher F344或Fisher F6)。大鼠多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)也可获自来源于两种或更多种上述品系的混合物的品系。在一个实施例中,大鼠多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)来源于选自DA品系和ACI品系的品系。在一个具体实施例中,大鼠多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)来源于ACI品系。ACI大鼠品系的特征在于具有黑野鼠色、腹部和足部为白色,并具有RT1
在各种实施例中,所述多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)、供体细胞和/或宿主胚胎不来自下列一个或多个物种:南美原鼠属物种(Akodon spp.)、林旅鼠属物种(Myopus spp.)、田鼠属物种(Microtus spp.)、鼹鼠属物种(Talpa spp.)。在各种实施例中,所述供体细胞和/或宿主胚胎不来自其正常的野生型特征为XY雌性可育性的任何物种。在各种实施例中,在多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)、供体细胞或宿主胚胎中存在遗传修饰的情况下,所述遗传修饰不是XYY和XXY、Tdy-阴性性逆转、Tdy-阳性性逆转、X0修饰、非整倍性、fgf9
所述方法和组合物中采用的多能和/或全能XY细胞(即,XY ES细胞或XY iPS细胞)具有导致Sry蛋白的水平和/或活性降低的遗传修饰。“性别决定区Y”蛋白或“Sry”蛋白是作为DNA结合蛋白的高迁移率族(HMG)盒家族的成员的转录因子。Sry是引发雄性性别决定的睾丸决定因子。来自多种生物体的Sry蛋白的序列是已知的,包括来自小鼠的Sry蛋白的序列(登记号Q05738)、来自大鼠的Sry蛋白的序列(GenBank登记号CAA61882.1)、来自人的Sry蛋白的序列(登记号Q05066)、来自猫的Sry蛋白的序列(登记号Q67C50)以及来自马的Sry蛋白的序列(登记号P36389),这些Sry蛋白的序列各自以引用方式并入本文。
一般来讲,如果Sry蛋白的蛋白水平和/或活性水平在统计学上低于尚未经过遗传修饰或诱变处理以抑制Sry蛋白的表达和/或活性的适当对照细胞中Sry蛋白的水平,则称所述Sry蛋白的水平和/或活性降低。在具体实施例中,相对于尚未经过修饰以具有降低的Sry蛋白水平和/或活性的对照细胞,Sry蛋白的浓度和/或活性至少降低1%、5%、10%、20%、30%、40%、50%、60%、70%、80%或90%。
“受试细胞”是其中已实现了遗传改变(诸如本文所公开的遗传修饰)的细胞,或为实现了这种遗传改变的细胞的后代并包含这种改变的细胞。“对照”或“对照细胞”提供了用于测量受试细胞的表型变化的参考点。在一个实施例中,对照细胞与Sry活性降低的细胞尽可能密切地匹配,区别之处为对照细胞缺少导致活性降低的遗传修饰或突变(例如,相应的细胞可源自相同的细胞系)。在其他情况下,对照细胞可包括例如:(a)野生型细胞,即与用于引起受试细胞中出现遗传改变的起始物质具有相同基因型的细胞;(b)与起始物质具有相同基因型,但已用无效构建体(即用对目标性状无已知作用的构建体,诸如包含标记基因的构建体)进行遗传修饰的细胞;(c)作为受试细胞的未经遗传修饰子代的细胞(即,对照细胞和受试细胞源于相同的细胞系);(d)在遗传上与受试细胞相同但未暴露于将诱导目标基因表达的条件或刺激的细胞;或者(e)在其中的遗传修饰不会导致目标多核苷酸的表达改变的条件下,受试细胞本身。
可以直接地(例如通过测定细胞或生物体中的Sry多肽的水平)或间接地(例如通过测量Sry多肽的活性)测量Sry多肽的表达水平。用于测定Sry蛋白活性的各种方法是已知的。参见Wang et al.(2013)Cell 153:910-918(Wang等人,2013年,《细胞》,第153卷,第910-918页);Mandalos et al.(2012)PLOS ONE 7:e45768:1-9(Mandalos等人,2012年,《美国公共科学图书馆期刊》,第7卷,第e45768期,第1-9页);以及Wang et al.(2013)NatBiotechnol.31:530-532(Wang等人,2013年,《自然生物技术》,第31卷,第530-532页),这些文献各自以引用方式并入本文。
在其他情况下,使用包括但不限于Southern印迹分析、DNA测序、PCR分析或表型分析的方法来选择具有降低Sry多肽的活性和/或水平的靶向遗传修饰的细胞。然后将此类细胞用于本文所述的各种方法、组合物和试剂盒。
靶向遗传修饰可包括对目标多核苷酸的靶向改变,包括例如对Y染色体上的靶基因组座位的靶向改变、对Sry基因的靶向改变,或者对其他所需多核苷酸的靶向改变。此类靶向修饰包括但不限于一个或多个核苷酸的添加、一个或多个核苷酸的缺失、一个或多个核苷酸的置换、目标多核苷酸或其部分的敲除、目标多核苷酸或其部分的敲入、用异源核酸序列替换内源核酸序列,或它们的组合。在具体实施例中,改变至少1个、2个、3个、4个、5个、7个、8个、9个、10个或更多个核苷酸以形成靶向基因组修饰。
Sry蛋白的水平和/或活性降低可由Sry基因中的遗传修饰(即,调控区、编码区和/或内含子等中的遗传修饰)引起。此类遗传修饰包括但不限于基因组中核苷酸的添加、缺失和置换。此类遗传修饰可包括对Sry基因的改变,包括例如将一个或多个核苷酸插入Sry基因、从Sry基因缺失一个或多个核苷酸、置换Sry基因中的一个或多个核苷酸、敲除Sry基因或其部分、敲入Sry基因或其部分、用异源核酸序列替换内源核酸序列,或它们的组合。因此,在具体实施例中,可通过破坏编码Sry多肽的基因来降低或消除Sry多肽的活性。在具体实施例中,改变Sry基因中的至少1个、2个、3个、4个、5个、7个、8个、9个、10个或更多个核苷酸。可使用各种方法产生另外的靶向遗传修饰。参见例如Wang et al.(2013)Cell 153:910-918(Wang等人,2013年,《细胞》,第153卷,第910-918页);Mandalos et al.(2012)PLOSONE 7:e45768:1-9(Mandalos等人,2012年,《美国公共科学图书馆期刊》,第7卷,第e45768期,第1-9页);以及Wang et al.(2013)Nat Biotechnol.31:530-532(Wang等人,2013年,《自然生物技术》,第31卷,第530-532页),这些文献各自以引用方式并入本文。另外,本文所描述的用于修饰Y染色体上的基因组座位的各种方法可用于引入对Sry基因的靶向遗传修饰。
在其他实施例中,通过将抑制Sry多肽的水平或活性的多核苷酸引入细胞中来降低或消除Sry多肽的活性和/或水平。所述多核苷酸可通过阻止Sry信使RNA翻译来直接抑制Sry多肽表达,或通过编码抑制所述编码Sry蛋白的基因的转录或翻译的多肽来间接抑制Sry多肽表达。在其他实施例中,通过将编码抑制Sry多肽活性的多肽的序列引入细胞中来降低或消除Sry多肽的活性。
在一个实施例中,XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)包含降低Sry蛋白的活性和/或水平的条件性Sry等位基因。“条件性Sry等位基因”包括被设计成在期望的发育时间和/或期望的目标组织内具有降低的Sry蛋白水平和/或活性的经修饰Sry基因。可将降低的水平和/或活性与缺少产生条件性等位基因的修饰的对照细胞进行比较,或者在期望的发育时间活性降低的情况下与先前时间和/或后续时间进行比较,或在期望的组织中活性降低的情况下与所有组织的平均活性进行比较。在一个实施例中,条件性Sry等位基因包括Sry的条件性无效等位基因,其可以在期望的发育时间点和/或在特定的组织中关闭。这种条件性等位基因可用于产生源自任何基因靶向克隆的可育XY雌性。如本文别处所述,采用这种方法能够在F1代中产生所需的纯合遗传修饰。采用此类方法,不必繁育至F2代,就可快速查看表型。
在非限制性实施例中,条件性Sry等位基因为如US 2011/0104799中所述的多功能等位基因,该专利全文以引用方式并入。在具体实施例中,所述条件性等位基因包含:(a)相对于靶基因的转录呈有义取向的起动序列,以及呈有义或反义取向的药物选择盒(DSC);(b)呈反义取向的目标核苷酸序列(NSI)和逆转条件模块(conditional by inversionmodule)(COIN,其利用外显子断裂内含子和可逆转的基因诱捕样模块;参见例如US 2011/0104799,该专利全文以引用方式并入);以及(c)在暴露于第一重组酶后重组以形成条件性等位基因的可重组单元,所述条件性等位基因(i)缺乏起动序列和DSC,并且(ii)含有呈有义取向的NSI和呈反义取向的COIN。
Sry基因的条件性等位基因可以在任何细胞类型中产生,而不限于XY多能和/或全能细胞。此类细胞类型以及靶向Y染色体上的基因组座位的非限制性方法在本文别处进一步详细讨论。
如本文别处所讨论,具有降低Sry蛋白的水平和/或活性的遗传修饰的多能和/或全能XY细胞(即,XY ES细胞或XY iPS细胞)还可包含对目标多核苷酸的至少一种另外的靶向遗传修饰。所述至少一种另外的靶向遗传修饰可包括一个或多个核酸的置换、用异源核酸序列替换内源核酸序列、敲除和敲入。所述另外的靶向遗传修饰可位于Y染色体、X染色体或常染色体上。可使用各种方法产生另外的靶向遗传修饰,包括采用如本文别处所讨论的靶向质粒和大靶向载体。还参见US20080092249、WO/1999/005266A2、US20040177390、WO/2008/017234A1和美国专利No.7,612,250,这些专利各自以引用方式并入本文,供读者参考与核移植相关的方法。此外,本文所述的修饰Y染色体上的基因组座位(即,Sry基因)的各种方法也可用于将靶向遗传修饰引入不位于Y染色体上的目标多核苷酸。
促进F0代中产生XY可育雌性的各种方法和组合物中所采用的培养基是维持多能和/或全能细胞(即,ES细胞、iPS细胞、XY ES细胞、XY iPS细胞等)的培养基。术语“维持”是指稳定地保存本文所述多能和/或全能细胞(包括ES细胞或iPS细胞)的至少一种或多种特征或表型。此类表型可包括维持多能性和/或全能性、细胞形态、基因表达谱以及细胞的其他功能特性。术语“维持”还可涵盖细胞增殖,或正在培养的细胞的数目增加。该术语还设想了允许细胞保持多能性,同时细胞可继续分裂也可不继续分裂、其数目可继续增加也可不继续增加的培养条件。
在一些实施例中,通过以下方式来维持具有降低Sry蛋白的水平和/或活性的遗传修饰的XY细胞:在本领域已知的(通过添加补充物)适用于在培养中生长或维持多能和/或全能细胞(即ES细胞、iPS细胞、XY ES细胞、XY iPS细胞等)的任何基础培养基(例如,DMEM)中培养所述XY细胞。在此类情况下,所培养的XY ES细胞具有发育成可育雌性动物的潜能,但依然保留多能性和/或全能性,使得所述细胞可被应用于受体胚胎中并产生可育的雌性子代。
在其他实施例中,通过以下方式来维持具有降低Sry蛋白的水平和/或活性的遗传修饰的XY细胞:在如下文进一步定义的培养基中培养所述XY细胞足够长的时间,让一些细胞转化为具有发育成可育雌性动物的潜能、但依然保留多能性和/或全能性的XY细胞,使得所述细胞可被应用于受体胚胎中并产生可育的雌性子代。
用于维持具有降低Sry蛋白的水平和/或活性的遗传修饰的XY多能和/或全能细胞(即XY ES细胞、XY iPS细胞等)的培养基促进XY F0可育雌性的发育。因此,与在合适的对照培养基(诸如,基于DMEM的一种培养基)中培养相比,在这种培养基中培养增加了所获得的XY F0可育雌性的数量。因此,XY F0可育雌性的数量增加可包括:F0非人动物中的至少10%、15%、20%、25%、30%、40%、50%、60%、70%、75%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%(在将非人动物XY ES细胞引入宿主胚胎中并孕育该宿主胚胎之后)为XY雌性,并且在达到性成熟后,所述F0 XY雌性非人动物是可育的。
短语“基础培养基”包括例如本领域已知的基础培养基(例如,DMEM),其(通过添加补充物)适用于在培养中生长或维持多能和/或全能细胞(即ES细胞、iPS细胞、XY ES细胞、XY iPS细胞等)。适用于产生可育XY雌性的基础培养基(即“低盐DMEM”或“低渗透压培养基”)不同于通常用于在培养中维持ES细胞的基础培养基。一般来讲,出于讨论基础培养基的目的,不适用于产生可育XY雌性的基础培养基在本节中被描述为“DMEM”,并且在表1中说明(例如,典型的DMEM培养基)。出于讨论适用于产生可育XY雌性的基础培养基的目的,使用短语“低盐DMEM”或“低渗透压DMEM”。本文明确地阐述了通常用于在培养中维持多能和/或全能细胞的基础培养基(例如,DMEM)与适用于产生可育XY雌性的基础培养基(例如,“低盐DMEM”)之间的差异。为方便起见,使用短语“低盐DMEM”;用于产生可育XY雌性的合适DMEM表现出的特征不限于“低盐”,但包括本文所述的那些。例如,可通过以下方式使表1所示的DMEM适用于产生可育的XY雌性:将氯化钠和/或碳酸氢钠的浓度改变为本文所提供的浓度,这也会引起渗透压和电导率与表1所示的DMEM相比有所不同。基础培养基的例子是各种形式的达尔伯克改良伊格尔培养基(Dulbeco's Modified Eagle's Medium)(DMEM)(例如,英杰公司(Invitrogen)的DMEM,产品目录号1 1971-025)(表1)。合适的低盐DMEM可作为KO-DMEM
术语“补充物”或短语“+补充物”包括添加到基础培养基中以用于在培养中生长或维持多能和/或全能细胞(即,XY ES细胞或XY iPS细胞),例如用于在培养中维持供体细胞的多能性或全能性的元素。例如,适用于在培养中生长或维持多能和/或全能细胞的培养基补充物包括但不限于胎牛血清(FBS)、谷氨酰胺、抗生素(一种或多种)、青霉素和链霉素(例如,青链霉素)、丙酮酸盐(例如,丙酮酸钠)、非必需氨基酸(例如,MEM NEAA)、2-巯基乙醇以及白血病抑制因子(LIF)。
在一个实施例中,基础培养基包含适用于在培养中维持多能细胞的一种或多种补充物,所述多能细胞包括例如在注入胚胎以及宫腔内移植到代孕母体小鼠内之后促成雄性性别决定发育程序的能力降低的XY ES细胞或XY iPS细胞。
在一个具体实施例中,适用于在培养中维持多能细胞的一种或多种补充物为FBS(90ml FBS/0.5L基础培养基)、谷氨酰胺(2.4mmol/0.5L基础培养基)、丙酮酸钠(0.6mmol/0.5L基础培养基)、非必需氨基酸(<0.1mmol/0.5L基础培养基)、2-巯基乙醇、LIF,以及一种或多种抗生素。
在其他实施例中,用于在培养中维持多能细胞(包括例如在注入胚胎以及宫腔内移植到代孕母体小鼠内之后促成雄性性别决定发育程序的能力降低的XY ES细胞或XY iPS细胞)的培养基包含约500ml基础培养基,在该基础培养基中添加了以下补充物:约90mlFBS(例如,Hylcone FBS产品目录号SH30070.03)、约2.4毫摩尔谷氨酰胺(例如,约12ml200mM谷氨酰胺溶液,例如英杰公司(Invitrogen)产品目录号25030-081)、青霉素:链霉素(例如,60,000单位青霉素G钠和60mg硫酸链霉素,含有约51mg NaCl;例如约6ml英杰公司(Invitrogen)青链霉素,产品目录号15140-122)、约0.6毫摩尔丙酮酸钠(例如,6ml 100mM丙酮酸钠,英杰公司(Invitrogen)产品目录号1 1360-070)、约0.06毫摩尔非必需氨基酸(例如,约6ml MEM NEAA,例如购自英杰公司(Invitrogen)、产品目录号为1 1 140-050的MEM NEAA)、约1.2ml2-巯基乙醇,以及约1.2微克LIF(例如,约120微升106单位/mL的LIF制备物;例如约120微升Millipore ESGRO
在一些实施例中,补充物包括Wnt条件培养基,例如Wnt-3a条件培养基。
在一个实施例中,将多能细胞(包括例如在注入胚胎以及宫腔内移植到代孕母体小鼠内之后促成雄性性别决定发育程序的能力降低的XY ES细胞或XY iPS细胞)维持在包含基础培养基和补充物的培养基中进行体外培养,其中所述基础培养基表现出以下一种或多种特征:(a)约200mOsm/kg至小于约329mOsm/kg的渗透压;(b)约11mS/cm至约13mS/cm的电导率;(c)约50mM至约110mM的碱金属卤化物盐浓度;(d)约17mM至约30mM的碳酸盐浓度;(e)约85mM至约130mM的碱金属卤化物盐和碳酸盐总浓度;和/或(f)这些特征之中任何两种或更多种的组合。在其他实施例中,将XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)维持在如WO2011/156723(该专利全文以引用方式并入本文)中所述的培养基中进行体外培养。
在一个实施例中,基础培养基为低盐DMEM。在一个具体实施例中,低盐DMEM的NaCl浓度为85至130mM。在一个实施例中,基础培养基为低渗透压DMEM。在一个具体实施例中,低渗透压DMEM的渗透压为250至310mOsm/kg。在一个实施例中,基础培养基为低电导率DMEM。在一个具体实施例中,低电导率DMEM的电导率为11至13mS/cm。
在其他实施例中,基础培养基表现出不超过约320、310、300、290、280、275、270、260、250或240mOsm/kg的渗透压。在一个实施例中,基础培养基或包含基础培养基和补充物的培养基表现出不超过约240至320mOsm/kg、250至310mOsm/kg、275至295mOsm/kg、或260至300mOsm/kg的渗透压。在一个具体实施例中,基础培养基或包含基础培养基和补充物的培养基表现出约270mOsm/kg的渗透压。
在其他实施例中,基础培养基表现出不超过约10.0、10.5、11.0、11.5、12.0、12.5、13.0、13.5或14.0mS/cm的电导率。在一个实施例中,基础培养基表现出不超过约10至14mS/cm或11至13mS/cm的电导率。在一个具体实施例中,基础培养基表现出约12至13mS/cm的电导率。
在一个具体实施例中,基础培养基表现出约12至13mS/cm的电导率和约260至300mOsm/kg的渗透压。在另一个具体实施例中,基础培养基包含浓度为约90mM的氯化钠。在另一个具体实施例中,氯化钠的浓度为约70至95mM。在另一个具体实施例中,基础培养基包含浓度小于约35mM的碳酸氢钠。在另一个具体实施例中,碳酸氢钠的浓度为约20至30mM。
在一个实施例中,基础培养基表现出不超过约100mM的碱金属卤化物盐浓度。在一个实施例中,碱金属卤化物盐为NaCl。在一个实施例中,碱金属卤化物盐的浓度不高于90、80、70、60或50mM。在一个实施例中,基础培养基中碱金属卤化物盐的浓度为约60至105mM、70至95mM、或80至90mM。在一个具体实施例中,所述浓度为约85mM。
在一个实施例中,基础培养基表现出一定的碳酸盐浓度。在一个实施例中,所述碳酸盐为钠盐。在一个实施例中,所述钠盐为碳酸氢钠。在一个实施例中,基础培养基中碳酸盐的浓度不高于40、35、30、25或20mM。在一个实施例中,基础培养基中碳酸盐的浓度为约10至40mM,在另一个实施例中为约20至30mM。在一个具体实施例中,所述浓度为约25或26mM。在其他实施例中,碳酸氢钠浓度为约26mM、约18mM、约18mM至约26mM、或约18mM至约44mM。
在一个实施例中,基础培养基中碱金属卤化物盐和碳酸盐的总浓度不超过140、130、120、110、100、90或80mM。在一个实施例中,基础培养基中碱金属卤化物盐和碳酸盐的总浓度为约80至140mM、85至130mM、90至120mM、95至120mM、或100至120mM。在一个具体实施例中,基础培养基中碱金属卤化物盐和碳酸盐的总浓度为约115mM。
在一个实施例中,碱金属卤化物盐与碳酸盐的摩尔比大于2.5。在一个实施例中,所述摩尔比为约2.6至4.0、2.8至3.8、3至3.6、或3.2至3.4。在一个实施例中,所述摩尔比为3.3至3.5。在一个具体实施例中,所述摩尔比为3.4。
在一个实施例中,基础培养基表现出约250至310mOsm/kg的渗透压,以及约60至105mM的碱金属卤化物盐浓度。在另一个实施例中,基础培养基的碳酸盐浓度为约20至30mM。在另一个实施例中,碱金属卤化物盐和碳酸盐的总浓度为约80至140mM。在另一个实施例中,基础培养基的电导率为约12至13mS/cm。
在一个实施例中,基础培养基包含约50±5mM NaCl和约26±5mM碳酸盐,且渗透压为约218±22mOsm/kg。在一个具体实施例中,基础培养基包含约3mg/mL NaCl和2.2mg/mL碳酸氢钠,且渗透压为约218mOsm/kg。
在另一个实施例中,基础培养基包含约87±5mM NaCl和约18±5mM碳酸盐,且渗透压为约261±26mOsm/kg。在一个具体实施例中,基础培养基包含约5.1mg/mL NaCl和约1.5mg/mL碳酸氢钠,且渗透压为约261mOsm/kg。
在另一个实施例中,基础培养基包含约110±5mM NaCl和约18±5mM碳酸盐,且渗透压为约294±29mOsm/kg。在一个具体实施例中,基础培养基包含约6.4mg/mL NaCl和约1.5mg/mL碳酸氢钠,且渗透压为约294mOsm/kg。
在另一个实施例中,基础培养基表现出约87±5mM NaCl和约26±5mM碳酸盐,且渗透压为约270±27mOsm/kg。在一个具体实施例中,基础培养基表现出约5.1mg/mL NaCl和约2.2mg/mL碳酸氢钠,且渗透压为约270mOsm/kg。
在另一个实施例中,基础培养基包含约87±5mM NaCl、约26±5mM碳酸盐以及约86±5mM葡萄糖,且渗透压为约322±32mOsm/kg。在一个具体实施例中,基础培养基包含约5.1mg/mL NaCl、约2.2mg/mL碳酸氢钠以及约15.5mg/mL葡萄糖,且渗透压为约322mOsm/kg。
在本文所公开的各种方法和组合物中可采用的另外的基础培养基包括含有50±5mM NaCl和26±5mM碳酸盐、且渗透压为218±22mOsm/kg的基础培养基。在一个具体实施例中,基础培养基包含约3mg/mL NaCl和2.2mg/mL碳酸氢钠,且渗透压为约218mOsm/kg。
在其他实施例中,基础培养基包含50±5mM NaCl和26±5mM碳酸盐,且渗透压为218±22mOsm/kg。在一个具体实施例中,基础培养基包含约3mg/mL NaCl和2.2mg/mL碳酸氢钠,且渗透压为约218mOsm/kg。
在其他实施例中,具有如本文所公开的NaHCO
可使用各种方法来产生降低Sry蛋白的水平和/或活性的靶向遗传修饰。例如,在一种情况下,靶向遗传修饰采用了将经由同源重组事件产生靶向遗传修饰的系统。在其他情况下,可使用在靶向基因组位置处产生单链或双链断裂的核酸酶试剂来修饰动物细胞。然后通过非同源性末端接合途径(NHEJ)来修复单链或双链断裂。此类系统可用于例如产生靶向功能丧失的遗传修饰。用于产生此类靶向遗传修饰的非限制性方法在本文别处详细讨论,包括例如使用靶向质粒、小靶向载体(smallTVEC)或大靶向载体。还参见Wang et al.(2013)Cell 153:910-918(Wang等人,2013年,《细胞》,第153卷,第910-918页);Mandaloset al.(2012)PLOS ONE 7:e45768:1-9(Mandalos等人,2012年,《美国公共科学图书馆期刊》,第7卷,第e45768期,第1-9页);以及Wang et al.(2013)Nat Biotechnol.31:530-532(Wang等人,2013年,《自然生物技术》,第31卷,第530-532页),这些文献各自以引用方式并入本文。
已经认识到在具体实施例中,对Sry基因的靶向遗传修饰和/或对任何其他目标多核苷酸的靶向遗传修饰可在多能细胞(即,ES细胞)正被维持在本文所述的培养基(例如,促进XY F0可育雌性发育的培养基)中时发生。另选地,对Sry基因和/或任何其他目标多核苷酸的靶向遗传修饰可在多能细胞(即,ES细胞)被维持在不同的培养基中、随后被转移到本文所述的培养基(例如,促进XY F0可育雌性发育的培养基)中时发生。
本发明提供了在体外培养中维持或培养XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)的方法,其中所述细胞包含降低Sry蛋白水平和/或活性的修饰,并且所述细胞在本文所述的条件下维持体外培养。此类在体外培养中维持或培养XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)的方法用于在将非人动物XY ES细胞引入宿主胚胎中并接着孕育该宿主胚胎之后,促进XY F0可育雌性动物的数目增加。
虽然本文所公开的任何培养基都可用于此类维持或培养方法,但一个非限制性例子包括在适用于在培养中维持或培养XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)、包含基础培养基和补充物的培养基中培养,其中基础培养基或包含基础培养基和补充物的培养基表现出约200mOsm/kg至小于约329mOsm/kg的渗透压。
在一些实施例中,基础培养基或包含基础培养基和补充物的培养基表现出以下一种或多种特征:约11mS/cm至约13mS/cm的电导率;约50mM至约110mM的碱金属卤化物盐浓度;约17mM至约30mM的碳酸盐浓度;约85mM至约130mM的碱金属卤化物盐和碳酸盐总浓度;和/或这些特征之中任何两种或更多种的组合。
在一个实施例中,所述方法包括在包含基础培养基和补充物的合适培养基中维持或培养XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞),其中所述基础培养基或所述包含基础培养基和补充物的培养基具有约240至320mOsm/kg的渗透压、约10至14mS/cm的电导率、约50至105mM的碱金属卤化物盐浓度、10至40mM的碳酸盐浓度,以及/或者约80至140mM的碱金属盐和碳酸盐组合浓度。在一个实施例中,将XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)置于培养基(含有用于维持ES细胞的补充物)中维持1天、2天、3天、4天、5天、6天、7天、8天、9天、10天、11天、12天、13天、2周、3周或4周,然后引入宿主胚胎中。在一个具体实施例中,将XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)置于培养基(含有用于维持ES细胞的补充物的低盐基础培养基)中维持约2至4周,然后引入宿主胚胎中。
在另一个实施例中,将XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)置于含有低盐基础培养基的培养基中维持至少1天、2天、3天、4天、5天、6天、7天、8天、9天、10天、11天、12天、2周、3周或4周,然后将供体细胞引入宿主胚胎中。在一个具体实施例中,将XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)置于含有低盐基础培养基的培养基中维持至少2至4周,然后将所述细胞引入宿主胚胎中。
在另一个实施例中,将XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)置于促进产生XY可育F0雌性的培养基中维持(例如,冷冻),之后将供体细胞解冻,置于促进产生XY可育F0雌性的培养基中维持至少1天、2天、3天、4天或更多天,然后把所述XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)引入宿主胚胎中。在一个具体实施例中,将XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)置于促进产生XY可育F0雌性的培养基中至少传代一次,将所述细胞冷冻在促进产生XY可育F0雌性的培养基中,之后在促进产生XY可育F0雌性的培养基中将所述细胞解冻,使所述细胞生长1天、2天、3天、4天、5天、6天、7天、8天、9天、10天、11天、12天、13天、2周、3周、4周或更长时间,然后引入宿主胚胎中。
在另一个实施例中,将XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)置于促进产生XY可育F0雌性的培养基中维持一天、二天、三天或四天,然后引入宿主胚胎中。在一个实施例中,将XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)置于促进产生XY可育F0雌性的培养基中维持三天。
在一个实施例中,将XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)置于促进产生XY可育F0雌性的培养基中维持约1天、2天、3天、4天、5天、6天、7天、8天、9天、10天、11天、12天、2周、3周、4周或更长时间,然后引入宿主胚胎中。在一个具体实施例中,将供体细胞置于促进产生XY可育F0雌性的培养基中维持至少一周,然后引入宿主胚胎中。在一个具体实施例中,将XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)置于促进产生XY可育F0雌性的培养基中维持2至4周,然后引入宿主胚胎中。
因此,提供了一种用于在培养中维持或培养XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)的方法,其中将细胞维持在下述条件下:在将所述XY细胞引入宿主胚胎中并接着在合适的雌性宿主体内孕育该宿主胚胎之后,这些条件促进或有利于雌性XY动物发育。
在一个方面,提供了一种用于在本文所述的条件下在培养中维持或培养供体XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)的方法,其中在将供体XY ES细胞引入宿主胚胎中以形成F0胚胎并接着在合适的动物体内孕育该F0胚胎之后,该F0胚胎发育成至少70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或更高百分比为XY,并且在达到性成熟后为可育雌性的F0动物。
采用本文提供的Sry蛋白水平和/或活性降低的XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)的各种方法和组合物可用于产生经遗传修饰的动物。用于引入遗传修饰的各种方法在本文别处详细讨论。
提供了一种用于在F0代中产生可育雌性XY非人动物的方法。此类方法包括:(a)在促进XY可育雌性ES细胞发育的培养基中维持或培养具有降低Sry蛋白水平和/或活性的修饰的供体非人动物XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞);(b)将所述供体XY非人动物XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)引入宿主胚胎中;(c)孕育该宿主胚胎;以及(d)获得F0 XY雌性非人动物,其中在达到性成熟后,所述F0 XY雌性非人动物是可育的。在具体实施例中,所述供体非人动物的XY供体细胞可在目标多核苷酸中包含至少一个另外的靶向遗传修饰。此类修饰在本文别处详细讨论。
具有降低Sry蛋白水平和/或活性的修饰的XY ES细胞可以在没有低盐培养基的情况下维持并且可发育成XY可育雌性。
在一些实施例中,促进XY可育F0雌性动物发育的培养基可包括低盐基础培养基,所述低盐基础培养基适用于在培养中维持或培养非人哺乳动物ES细胞并包含基础培养基和补充物,其中所述低盐基础培养基表现出包括以下一者或多者的特征:约200mOsm/kg至小于约329mOsm/kg的渗透压;约11mS/cm至约13mS/cm的电导率;约50mM至约110mM的碱金属卤化物盐浓度;约17mM至约30mM的碳酸盐浓度;约85mM至约130mM的碱金属卤化物盐和碳酸盐总浓度;和/或这些特征之中任何两种或更多种的组合。
在其他实施例中,可使用本文所公开的培养基来执行此类用于在F0代中产生可育雌性XY非人动物的方法,所述培养基包括但不限于:(a)包含50±5mM NaCl、26±5mM碳酸盐,且渗透压为218±22mOsm/kg的基础培养基;(b)包含约3mg/mL NaCl、2.2mg/mL碳酸氢钠,且渗透压为218mOsm/kg的基础培养基;(c)包含87±5mM NaCl、18±5mM碳酸盐,且渗透压为261±26mOsm/kg的基础培养基;(d)包含约5.1mg/mL NaCl、1.5mg/mL碳酸氢钠,且渗透压为261mOsm/kg的基础培养基;(e)包含110±5mM NaCl、18±5mM碳酸盐,且渗透压为294±29mOsm/kg的基础培养基;(f)包含约6.4mg/mL NaCl、1.5mg/mL碳酸氢钠,且渗透压为294mOsm/kg的基础培养基;(g)包含87±5mM NaCl、26±5mM碳酸盐,且渗透压为270±27mOsm/kg的基础培养基;(h)包含约5.1mg/mL NaCl、2.2mg/mL碳酸氢钠,且渗透压为270mOsm/kg的基础培养基;(i)包含87±ì5mM NaCl、26±5mM碳酸盐、86±5mM葡萄糖,且渗透压为322±32mOsm/kg的基础培养基;以及/或者(j)包含约5.1mg/mL NaCl、2.2mg/mL碳酸氢钠、15.5mg/mL葡萄糖,且渗透压为322mOsm/kg的基础培养基。
可将具有降低Sry蛋白的水平和/或活性的修饰并且已在促进XY F0可育雌性发育的培养基中培养过的经遗传修饰的XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)植入宿主胚胎中。已植入宿主胚胎中的细胞在本文中称为“供体细胞”。在具体实施例中,经遗传修饰的XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)来自与宿主胚胎相同的品系或来自与宿主胚胎不同的品系。同样,代孕母体可来自与经遗传修饰的XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)和/或宿主胚胎相同的品系,或者代孕母体可来自与经遗传修饰的XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)和/或宿主胚胎不同的品系。在一个实施例中,将XY供体细胞植入XX宿主胚胎中。
在本文所公开的方法和组合物中可采用多种宿主胚胎。在一些实施例中,将具有导致Sry蛋白水平和/或活性降低的靶向遗传修饰的XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)引入来自相应生物体的桑椹胚前期胚胎(例如,8-细胞期胚胎)中。参见例如US 7,576,259、US 7,659,442、US 7,294,754和US 2008-0078000 A1,所有这些专利均全文以引用方式并入本文。在其他实施例中,可在宿主胚胎处于2-细胞期、4-细胞期、8-细胞期、16-细胞期、32-细胞期或64-细胞期时将供体ES细胞植入宿主胚胎中。在另一个实施例中,宿主胚胎为囊胚。在一个实施例中,宿主胚胎处于选自下列的时期:囊胚前期、桑椹胚前期、桑椹胚期、未致密化桑椹胚期和致密化桑椹胚期。在一个实施例中,当采用小鼠胚胎时,宿主胚胎期选自泰勒期(Theiler Stage)1(TS1)、TS2、TS3、TS4、TS5和TS6,参考Theiler(1989)“The House Mouse:Atlas of Mouse Development,”Springer-Verlag,New York(Theiler,1989年,《家鼠:小鼠发育数据集》,纽约施普林格出版公司)中所述的泰勒期。在一个具体实施例中,泰勒期选自TS1、TS2、TS3和TS4。在一个实施例中,宿主胚胎包含透明带,并且供体细胞为通过透明带中的孔引入宿主胚胎内的XY ES细胞;而在其他实施例中,宿主胚胎为无透明带的胚胎。在其他具体实施例中,桑椹胚期宿主胚胎为聚合胚胎。
也可使用核移植技术生成经遗传修饰的动物。简而言之,用于核移植的方法包括以下步骤:(1)将卵母细胞去核;(2)分离供体细胞或核,准备与去核卵母细胞合并;(3)将所述细胞或核插入所述去核卵母细胞中,以形成重建细胞;(4)将所述重建细胞植入动物的子宫,以形成胚胎;以及(5)允许所述胚胎发育。在此类方法中,一般从处死的动物体内取出卵母细胞,不过也可从活体动物的输卵管和/或卵巢中分离卵母细胞。卵母细胞可在去核之前在本领域普通技术人员已知的多种培养基中成熟。将卵母细胞去核可采用本领域普通技术人员熟知的多种方式执行。将供体细胞或核插入去核卵母细胞中以形成重建细胞,通常借助在融合之前在透明带下显微注射供体细胞而实现。融合可通过跨接触/融合平面施加直流电脉冲(电融合)、通过将细胞暴露于促进融合的化学品(诸如聚乙二醇)、或借助灭活病毒(诸如仙台病毒)来诱导。重建细胞通常在核供体和受体卵母细胞融合之前、期间和/或之后,通过电力方式和/或非电力方式激活。激活方法包括电脉冲、化学诱导冲击、精液渗透、增加二价阳离子在卵母细胞中的水平以及降低细胞蛋白在卵母细胞中的磷酸化(如借助激酶抑制剂)。激活的重建细胞或胚胎通常在本领域普通技术人员熟知的培养基中培养,然后移植到动物的子宫中。参见例如US20080092249、WO/1999/005266A2、US20040177390、WO/2008/017234A1以及美国专利No.7,612,250,这些专利各自以引用方式并入本文。
将包含Sry蛋白水平和/或活性降低的经遗传修饰的XY多能和/或全能细胞(即,XYES细胞或XY iPS细胞)的宿主胚胎孵育直至囊胚期,随后植入代孕母体中以产生F0动物。可经由如本文所述的等位基因修饰(MOA)测定法鉴定具有经遗传修饰的基因组座位的动物。
在一个实施例中,将包含Sry蛋白水平和/或活性降低的经遗传修饰的XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)的宿主胚胎置于促进XY可育雌性ES细胞发育的培养基(即,低盐基础培养基)中维持一天、两天、三天、四天或更多天,然后植入合适的宿主。此类方法对生成F0可育雌性动物提供支持。
在一个实施例中,将所培养的宿主胚胎植入代孕母体,然后在代孕母体中孕育所培养的宿主胚胎。
在具体实施例中,在将非人动物XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)引入宿主胚胎中并接着孕育该宿主胚胎之后,至少15%、20%、30%、40%、50%、60%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的F0非人动物为XY雌性,在达到性成熟后,所述F0 XY雌性非人哺乳动物是可育的。
还提供了F0胚胎,该F0胚胎包含具有至少一个异源干细胞的内细胞团,所述异源干细胞包括具有降低Sry蛋白水平和/或活性的靶向遗传修饰的XY ES细胞或XY iPS细胞。
本文所述的在F0代中产生可育雌性XY非人动物的各种方法可采用具有以下遗传修饰的XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞):(1)用于降低Sry多肽的水平和/或活性的遗传修饰;以及在具体实施例中,(2)目标多核苷酸中的一种或多种另外的靶向遗传修饰。如本文别处所概述,可在XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)中产生至少1种、2种、3种、4种、5种、6种、7种、8种、9种、10种或更多种另外的靶向遗传修饰。在此类情况下,F0可育雌性XY非人动物可包含这些另外的靶向遗传修饰中的一种或多种。
在其他实施例中,F0可育雌性XY非人动物在其一生中产下1窝、2窝、3窝、4窝、5窝、6窝、7窝、8窝或9窝幼仔。在一个实施例中,F0可育雌性XY非人动物每窝至少产下1只、2只、3只、4只、5只、6只、7只、8只、9只或10只后代。在一个实施例中,F0可育雌性XY非人动物每窝产下约4至6只后代。在一个实施例中,F0可育雌性XY非人动物产下2至6窝幼仔,其中每窝至少有2只、3只、4只、5只或6只后代。在一个实施例中,所述后代中的至少约10%、15%、20%、25%、30%、35%、40%、45%、50%、60%、70%、80%、85%、90%、95%或100%为XY可育雌性后代。
还提供了用于产生啮齿动物成窝幼仔(即,小鼠或大鼠的成窝幼仔)的方法,包括将根据本文所述方法制备的Sry蛋白水平和/或活性降低的XY多能和/或全能供体细胞(即,XY供体ES细胞或XY供体iPS细胞)引入宿主胚胎中,在合适的代孕母体中孕育所述胚胎,以及获得包括至少一只XY雌性啮齿动物的F0子代,所述XY雌性啮齿动物在达到性成熟后为可育的XY雌性啮齿动物。在一个实施例中,出生的在达到性成熟后可育的F0 XY雌性啮齿动物的百分比为约10%、15%、20%、25%、30%、35%、40%、45%、50%、60%、70%、75%、80%、85%、95%或100%。
在其他实施例中,用此类方法产生的F0子代中约3%、约10%或更多、或者约63%或更多来源于经遗传修饰的供体XY细胞。
本文所提供的方法和组合物允许F0动物中的至少1%、3%、5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%或更多具有靶向遗传修饰(即,降低Sry蛋白水平和/或活性的靶向遗传修饰和/或对目标多核苷酸的靶向遗传修饰),以将该遗传修饰传递给F1子代。
在一个实施例中,F0代雌性XY非人动物和/或雄性XY非人动物中的至少90%、92%、94%、96%、98%、99%或99.8%来源于供体细胞。在一个实施例中,F0雌性XY非人动物和/或F0雄性XY非人动物具有的毛色100%来源于供体细胞。
在一个实施例中,F0代中的非人雌性XY动物为啮齿动物(即,小鼠或大鼠),其毛色100%来源于供体细胞。在一个实施例中,F0代中所产生的非人雌性XY动物中的至少90%、92%、94%、96%、98%或99.8%来源于XY供体细胞。在一个实施例中,F0代中的非人雌性XY动物约100%来源于供体细胞。在一个实施例中,通过能够在2,000个细胞中检出1个细胞(0.05%)的定量测定法来确定宿主胚胎细胞对F0代中的非人雌性XY动物的贡献,雌性XY动物的任何组织都不会增大宿主胚胎细胞的贡献。
在具体实施例中,将来源于具有降低Sry蛋白水平和/或活性的遗传修饰的XY多能和/或全能细胞(即,XY ES细胞或XY iPS细胞)的所得雌性可育XY F0代与动物杂交,获得F1代后代。在具体实施例中,将雌性可育XY F0与野生型动物杂交。在一个实施例中,所述雌性XY F0非人哺乳动物在与野生型小鼠杂交时是可育的。在具体实施例中,所述野生型小鼠为C57BL/6。可使用特异性引物和/或探针对F1子代进行基因分型,以确定是否存在导致Sry蛋白水平和/或活性降低的靶向遗传修饰。此外,如果F0代中存在另外的靶向遗传修饰,则可使用用于确定是否存在此类修饰的特异性引物和/或探针对F1子代进行基因分型。然后可鉴定用于所需用途的适当F1子代。在具体实施例中,选择缺少用于降低Sry蛋白水平和/或活性的遗传修饰的F1子代。在其他实施例中,选择缺少用于降低Sry蛋白水平和/或活性的遗传修饰并且包含至少一种另外的靶向遗传修饰的F1子代。
在一个非限制性例子中,在用特异性引物和/或探针进行基因分型之后,将对于目标多核苷酸的靶向遗传修饰为杂合的并且缺少用于降低Sry蛋白水平和/或活性的靶向修饰的F1动物彼此杂交。这种杂交产生了对于经遗传修饰的目标基因组座位为纯合的并且不包含用于降低Sry蛋白水平和/或活性的遗传修饰的F2子代。
还提供了在F1代中产生对于靶向遗传修饰为纯合的转基因非人动物的方法。该方法包括:(a)使具有降低Sry蛋白水平和/或活性的靶向遗传修饰的F0 XY可育雌性非人动物与F0 XY雄性非人动物杂交,其中所述F0XY可育雌性非人动物和所述F0 XY雄性非人动物各自对于目标多核苷酸的相同遗传修饰均为杂合的,以及(b)获得对于目标多核苷酸中的靶向遗传修饰为纯合的F1子代。在一个具体实施例中,所选择的F1子代对于目标多核苷酸中的靶向遗传修饰为纯合的,并且缺少用于降低Sry蛋白活性和/或水平的靶向遗传修饰。此类方法可用于开发繁育用成对非人动物,其中每个非人动物完全来源于相同F0代中的供体ES细胞或iPS细胞。
可采用各种方法来获得上述F0动物。在一个非限制性实施例中,将在任何染色体上具有对目标多核苷酸的靶向修饰的XY细胞克隆分离。已经认识到,可使用各种方法在目标多核苷酸中产生靶向修饰。在第二步骤中,将靶向修饰引入Sry基因中,使得该修饰降低Sry蛋白的水平和/或活性。此类方法将进一步采用在促进XY F0可育雌性发育的培养基中培养XY ES细胞,如本文别处详细描述。靶向修饰Sry基因的方法在本文别处详细公开,可包括例如单独使用靶向载体(包括LTVEC),或组合使用靶向载体与本文别处所述的核酸酶(即,Talen、CRISPR或ZFN体系)。分离既包含对目标多核苷酸的第一靶向修饰,又包含用于降低Sry蛋白的水平和/或活性的第二靶向Sry基因修饰的亚克隆。将在目标多核苷酸中具有靶向修饰的原始XY克隆以及既包含对Sry基因的靶向修饰又包含对目标多核苷酸的靶向修饰的XY亚克隆都引入单独的非人宿主胚胎中,如本文别处所讨论。在具体实施例中,非人宿主胚胎包括桑椹胚前期胚胎(即,8-细胞期胚胎)。将包含经修饰多能细胞的每个非人宿主胚胎引入代孕母体内进行孕育。每个代孕母体均产生包含靶向基因组修饰的F0子代(即,在目标多核苷酸中具有靶向修饰的F0 XY雄性,以及具有对目标多核苷酸的靶向修饰并且具有降低Sry蛋白的水平和/或活性的遗传修饰的F0 XY可育雌性)。在具体实施例中,靶向基因组修饰中的每一种都能够通过种系传递。将这些F0动物中的每一个彼此交配,产生包含对目标多核苷酸的纯合靶向修饰的F1动物。预期F1代中的四分之一对于目标多核苷酸中的靶向修饰为纯合的。可选择保留了对Sry基因的靶向修饰的F1子代,也可选择不保留对Sry基因的靶向修饰的F1子代。
在另一个实施例中,采用靶向载体(在具体实施例中,还采用核酸酶诸如Talen、Crispr或Zfn)引入对Sry基因的靶向修饰可与载体靶向对目标多核苷酸的遗传修饰同时发生。此类方法允许产生具有降低Sry蛋白的水平和/或活性的遗传修饰并且还包含对目标多核苷酸的靶向修饰的XY ES细胞。
在一个实施例中,F1代子代包含完全来源于供体ES细胞的基因组。在其他实施例中,产生完全来源于ES细胞的小鼠的F0代雄性小鼠与F0代雌性小鼠的杂交频率为100%。
提供了允许修饰细胞中Y染色体上的靶基因组座位的方法和组合物。还提供了允许修饰“挑战性(challenging)”基因组座位的方法。术语“挑战性基因座”包括难以通过常规的基因靶向策略靶向的染色体区域。此类基因座可位于Y染色体、X染色体或常染色体上。在某些实施例中,挑战性基因座位于基因少、富含重复序列和/或大部分为异染色质的染色体区域内或附近。参见例如Bernardini et al.,Proc.Natl.Acad.Sci.USA 111:7600-7605(2014)(Bernardini等人,《美国国家科学院院刊》,第111卷,第7600-7605页,2014年),该文献全文以引用方式并入本文用于所有目的。在某些实施例中,挑战性基因座位于其中染色体DNA的可及性受染色质结构限制的染色体区域内或附近。在某些实施例中,挑战性基因座在特征为异染色质百分比高(诸如异染色质为至少约20%、至少约30%、至少约40%、少约50%、至少约60%或至少约70%)的染色体区域内或附近。在某些实施例中,挑战性基因座位于已经历复制和重排或者特征为存在重复序列或反向重复序列的染色体区域内或附近。参见例如Gubbay et al.,Proc.Natl.Acad.Sci.USA 89:7953-7957(1992)(Gubbay等人,《美国国家科学院院刊》,第89卷,第7953-7957页,1992年),该文献全文以引用方式并入本文用于所有目的。
术语“染色质”包括核蛋白复合物,其致密化并组织细胞遗传物质,从而在细胞内包含细胞遗传物质。术语“异染色质”包括基因组中处于高度浓缩状态并且通常转录沉默的区域。与常染色质相比,异染色质通常更紧密地盘绕并且通常具有重复性更高的DNA序列。术语“常染色质”包括基因组中由延伸得更长且浓缩度较低的染色质结构域表征的区域,所述染色质结构域通常具有转录活性并且是可及的。
术语“暴露”包括采用任何方法使所需组分变得紧邻或直接接触。
提供了允许修饰细胞中Y染色体上的挑战性靶基因组座位或靶基因组座位的方法和组合物。也许是由于Y染色体具有独特的结构特征,所以在小鼠胚胎干细胞中产生Y连锁基因突变的常规基因靶向策略已获得的成果有限。因此通常情况下,对鼠Y连锁基因功能的理解受限于从对携带自发缺失、随机基因捕获插入或常染色体转基因的小鼠的研究获得的见解。本文提供的方法允许通过在不存在核酸酶试剂的情况下使用靶向载体或通过与核酸酶试剂组合使用靶向载体,来靶向Y染色体上的基因组座位。
一些此类方法利用小靶向载体(或称smallTVEC)。“smallTVEC”包括含有短同源臂的靶向载体。smallTVEC上同源臂的长度可为约400bp至1000bp。smallTVEC的同源臂可具有足以促进与对应靶位点发生同源重组事件的任何长度,包括例如约400bp至约500bp、约500bp至约600bp、约600bp至约700bp、约700bp至约800bp、约800bp至约900bp、或约900bp至约1000bp。smallTVEC上同源臂的优选长度为约700bp至约800bp。在另一个实施例中,smallTVEC的5’和3’同源臂的总和为约0.5kb、1kb、1.5kb、2kb、3kb、4kb、5kb、6kb、7kb、8kb、9kb、约0.5kb至约1kb、约1kb至约1.5kb、约1.5kb至约2kb、约2kb至约3kb、约3kb至约4kb、约4kb至约5kb、约5kb至约6kb、约6kb至约7kb、约8kb至约9kb,或为至少10kb。在此类方法中,与具有较长同源臂的靶向载体相比,长度短的同源臂增大了靶向效率。由于Y染色体的性质为具有高度重复的序列,所以smallTVEC的短臂允许高度特异性地靶向Y染色体。
提供了用于修饰细胞中Y染色体上的靶基因组座位的方法,包括:(a)提供在Y染色体上包含靶基因组座位的细胞,所述靶基因组座位包含核酸酶试剂的识别位点;(b)向该细胞中引入包含第一插入多核苷酸的第一靶向载体,所述第一插入多核苷酸侧接与第一靶位点和第二靶位点对应的第一同源臂和第二同源臂;以及(c)鉴定在其基因组中包含在Y染色体上的所述靶基因组座位处整合的第一插入多核苷酸的至少一个细胞。在具体实施例中,靶向载体的第一同源臂和第二同源臂的总和为约0.5kb、1kb、1.5kb、2kb、3kb、4kb、5kb、6kb、7kb、8kb、9kb、约0.5kb至约1kb、约1kb至约1.5kb、约1.5kb至约2kb、约2kb至约3kb、约3kb至约4kb、约4kb至约5kb、约5kb至约6kb、约6kb至约7kb、约8kb至约9kb,或为至少10kb或至少10kb且少于150kb。在一些实施例中,采用smallTVEC。在具体实施例中,采用LTVEC。在靶向挑战性靶基因组座位时,可执行类似的方法。在一个非限制性实施例中,执行此类方法,其采用促进本文所公开的XY F0可育雌性发育的培养基,从而生成XY F0可育雌性动物。在其他情况下,本文所述的方法用于在Sry基因中产生靶向遗传修饰,如本文别处所讨论。
还提供了用于修饰细胞中Y染色体上的靶基因组座位的方法,包括:(a)提供在Y染色体上包含靶基因组座位的细胞,所述靶基因组座位包含核酸酶试剂的识别位点,(b)向该细胞中引入(i)核酸酶试剂,其中所述核酸酶试剂在第一识别位点处诱导切口或双链断裂;和(ii)包含第一插入多核苷酸的第一靶向载体,所述第一插入多核苷酸侧接第一同源臂和第二同源臂,所述第一同源臂和第二同源臂与位置足够接近第一识别位点的第一靶位点和第二靶位点相对应;以及(c)鉴定在其基因组中包含在Y染色体上的靶基因组座位处整合的第一插入多核苷酸的至少一个细胞。在具体实施例中,靶向载体的第一同源臂和第二同源臂的总和为约0.5kb、1kb、1.5kb、2kb、3kb、4kb、5kb、6kb、7kb、8kb、9kb、约0.5kb至约1kb、约1kb至约1.5kb、约1.5kb至约2kb、约2kb至约3kb、约3kb至约4kb、约4kb至约5kb、约5kb至约6kb、约6kb至约7kb、约8kb至约9kb,或为至少10kb或至少10kb且少于150kb。在一些实施例中,采用smallTVEC。在具体实施例中,采用LTVEC。在靶向挑战性靶基因组座位时,可执行类似的方法。在一个非限制性实施例中,执行此类方法,其采用促进本文所公开的XY F0可育雌性发育的培养基,从而生成XY F0可育雌性动物。在其他情况下,本文所述的方法用于在Sry基因中产生靶向遗传修饰,如本文别处所讨论。
已经认识到,本文所公开的采用靶向载体、smallTVEC或LTVEC在Y染色体的基因组座位(或任何挑战性基因组座位)中产生靶向修饰的各种方法可以在任何细胞类型中执行,并不限于XY多能和/或全能细胞。此类细胞类型包括但不限于人细胞、非人细胞、哺乳动物细胞、非人哺乳动物细胞、啮齿动物细胞、小鼠细胞、大鼠细胞、仓鼠细胞、成纤维细胞或任何其他宿主细胞。此类细胞包括多能细胞,包括例如诱导性多能干(iPS)细胞、小鼠胚胎干(ES)细胞、大鼠胚胎干(ES)细胞、人胚胎干(ES)细胞或发育受限的人祖细胞。
还公开了采用本文所提供的各种核酸酶试剂中的任一种(例如,CRISPR gRNA与Cas9、ZFN或TALEN的组合)在Y染色体上产生大段缺失的方法。Y染色体上的这种缺失可以是内源核酸序列的缺失。所述缺失可在约5kb至约10kb、约10kb至约20kb、约20kb至约40kb、约40kb至约60kb、约60kb至约80kb、约80kb至约100kb、约100kb至约150kb、约150kb至约200kb、约200kb至约300kb、约300kb至约400kb、约400kb至约500kb、约500kb至约600kb、约600kb至约700kb、约700kb至约800kb、约800kb至约900kb、约900kb至约1Mb、约500kb至约1Mb、约1Mb至约1.5Mb、约1.5Mb至约2Mb、约2Mb至约2.5Mb、或约2.5Mb至约3Mb的范围内。在一个实施例中,所述缺失大于500kb。在另一个实施例中,所述缺失为约500kb至约600kb。在一个具体实施例中,所述缺失为约500kb。Y染色体上的这种缺失可以是任意核酸序列的缺失。在一个实施例中,所述缺失包括与可育性/不育性相关联的基因。Y染色体上的所述缺失可包括多个基因的缺失。在此类方法中,可缺失1个、2个、3个、4个、5个、6个、7个、8个、9个、10个或更多个基因。在具体实施例中,Kdm5d基因(赖氨酸(K)特异性去甲基化酶5d;例如Entrez Gene ID 20592(小家鼠))和/或Usp9y基因(泛素特异性肽酶9,y连锁;例如EntrezGene ID 107868(小家鼠))是缺失所靶向的基因。在其他实施例中,Sry基因是缺失所靶向的基因。
术语“核酸酶试剂的识别位点”包括用核酸酶试剂在其处诱导切口或双链断裂的DNA序列。核酸酶试剂的识别位点对于细胞可为内源的(或天然的),或所述识别位点对于细胞可为外源的。在具体实施例中,所述识别位点对于细胞是外源的,因此并非天然存在于细胞的基因组中。在另外的实施例中,所述识别位点对于细胞是外源的,对于期望将其定位在靶基因座处的目标多核苷酸也是外源的。在另外的实施例中,宿主细胞的基因组中只存在一处外源或内源的识别位点。在具体实施例中,鉴定在基因组内仅出现一次的内源或天然位点。随后可使用这种位点来设计将在内源识别位点处产生切口或双链断裂的核酸酶试剂。
识别位点的长度可能有变,包括例如锌指核酸酶(ZFN)对的约30至36bp识别位点(即,针对每个ZFN约15至18bp)、转录激活因子样效应物核酸酶(TALEN)的约36bp识别位点,或者CRISPR/Cas9向导RNA的约20bp识别位点。
在一个实施例中,每种核酸酶试剂单体识别至少含9个核苷酸的识别位点。在其他实施例中,所述识别位点的长度为约9至约12个核苷酸、约12至约15个核苷酸、约15至约18个核苷酸、或约18至约21个核苷酸,以及此类子范围的任意组合(例如,9至18个核苷酸)。已经认识到,给定核酸酶试剂可结合所述识别位点并切割该结合位点;另选地,核酸酶试剂可结合至不同于所述识别位点的序列。此外,术语“识别位点”既包括核酸酶试剂结合位点,又包括切口/裂解位点(不管切口/裂解位点是在核酸酶试剂结合位点内、还是核酸酶试剂结合位点外)。在另一种变型中,核酸酶试剂引起的裂解可发生在彼此直接相对的核苷酸位置处,以产生平头末端切口,或者在其他情况下,所述切口可交错而产生单链悬垂(也称为“粘性末端”),其可为5'悬垂或3'悬垂。
用于诱导切口或双链断裂到期望识别位点中的任何核酸酶试剂都可用于本文所公开的方法和组合物。可采用天然存在的或天然的核酸酶试剂,只要这种核酸酶试剂在期望的识别位点中诱导切口或双链断裂即可。另选地,可采用经修饰的或经改造的核酸酶试剂。“经改造的核酸酶试剂”包括从其天然形式进行改造(修饰或衍生)以特异性地识别并诱导期望的识别位点中的切口或双链断裂的核酸酶。因此,经改造的核酸酶试剂可衍生自天然的、天然存在的核酸酶试剂,也可人工产生或合成。对核酸酶试剂的修饰可能只发生在蛋白裂解试剂中的一个氨基酸处或核酸裂解试剂中的一个核苷酸处。在一些实施例中,经改造的核酸酶在识别位点中诱导切口或双链断裂,其中所述识别位点是原本不会被天然的(未经改造或未经修饰的)核酸酶试剂识别的序列。在识别位点或其他DNA中产生切口或双链断裂在本文中可称为“切割”或“裂解”识别位点或其他DNA。
还提供了示例性识别位点的活性变体和片段。此类活性变体可与给定的识别位点至少具有65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或更高的序列同一性,其中所述活性变体保留生物活性,因此能够被核酸酶试剂以序列特异性方式识别和裂解。用核酸酶试剂测量识别位点的双链断裂的测定法是本领域已知的(例如,
在具体实施例中,所述识别位点位于编码选择标记的多核苷酸内。这种位置可位于选择标记的编码区内或影响选择标记表达的调控区内。因此,核酸酶试剂的识别位点可位于选择标记的内含子中,或者编码选择标记的多核苷酸的启动子、增强子、调控区或任何非蛋白编码区中。在具体实施例中,识别位点处出现的切口或双链断裂破坏了选择标记的活性。用来测定功能性选择标记存在与否的方法是已知的。
在一个实施例中,所述核酸酶试剂为转录激活因子样效应物核酸酶(TALEN)。TAL效应物核酸酶是一类序列特异性核酸酶,可用于在原核或真核生物体的基因组中的特定靶序列处产生双链断裂。TAL效应物核酸酶是通过将天然的或经改造的转录激活因子样(TAL)效应物或其功能部分融合至内切核酸酶(诸如FokI)的催化结构域而产生的。独特的模块化TAL效应物DNA结合结构域允许设计具有潜在的任何给定DNA识别特异性的蛋白质。因此,TAL效应物核酸酶的DNA结合结构域可被改造以识别特定的DNA靶位点,因而用于在期望的靶序列处产生双链断裂。参见WO 2010/079430;Morbitzer et al.(2010)PNAS 10.1073/pnas.1013133107(Morbitzer等人,2010年,《美国国家科学院院刊》,DOI号:10.1073/pnas.1013133107);Scholze&Boch(2010)Virulence 1:428-432(Scholze和Boch,2010年,《毒力》,第1卷,第428-432页);Christian et al.Genetics(2010)186:757-761(Christian等人,《遗传学》,2010年,第186卷,第757-761页);Li et al.(2010)Nuc.Acids Res.(2010)doi:10.1093/nar/gkq704(Li等人,2010年,《核酸研究》,DOI号:10.1093/nar/gkq704);以及Miller et al.(2011)Nature Biotechnology 29:143–148(Miller等人,2011年,《自然生物技术》,第29卷,第143-148页);所有这些文献均以引用方式并入本文。
合适TAL核酸酶的例子以及用于制备合适TAL核酸酶的方法公开于例如美国专利申请No.2011/0239315 A1、No.2011/0269234 A1、No.2011/0145940 A1、No.2003/0232410A1、No.2005/0208489 A1、No.2005/0026157 A1、No.2005/0064474 A1、No.2006/0188987A1以及No.2006/0063231 A1中(每份专利申请据此以引用方式并入)。在各种实施例中,TAL效应物核酸酶被改造成在例如目标基因座或目标基因组座位中的靶核酸序列之中或附近进行切割,其中所述靶核酸序列位于靶向载体将要修饰的序列之处或附近。适合与本文所提供的各种方法和组合物一起使用的TAL核酸酶包括被特别设计成在本文所述的靶向载体将要修饰的靶核酸序列之处或附近结合的那些TAL核酸酶。
在一个实施例中,TALEN的每个单体包含经由两个高变残基识别单碱基对的33至35个TAL重复序列。在一个实施例中,核酸酶试剂为包含有效连接至独立核酸酶的基于TAL重复序列的DNA结合结构域的嵌合蛋白。在一个实施例中,独立核酸酶为FokI内切核酸酶。在一个实施例中,核酸酶试剂包含第一基于TAL重复序列的DNA结合结构域和第二基于TAL重复序列的DNA结合结构域,其中所述第一基于TAL重复序列的DNA结合结构域和第二基于TAL重复序列的DNA结合结构域每一者均有效连接至FokI核酸酶,其中所述第一基于TAL重复序列的DNA结合结构域和第二基于TAL重复序列的DNA结合结构域识别DNA靶序列每条链中被不同长度(12-20bp)的间隔序列隔开的两条邻接DNA靶序列,并且其中所述FokI核酸酶亚基发生二聚化,从而生成能在靶序列处产生双链断裂的活性核酸酶。
在本文所公开的各种方法和组合物中采用的核酸酶试剂还可包括锌指核酸酶(ZFN)。在一个实施例中,ZFN的每个单体包含3个或更多个基于锌指的DNA结合结构域,其中每个基于锌指的DNA结合结构域结合于3bp亚位点。在其他实施例中,ZFN为包含有效连接至独立核酸酶的、基于锌指的DNA结合结构域的嵌合蛋白。在一个实施例中,独立内切核酸酶为FokI内切核酸酶。在一个实施例中,核酸酶试剂包含第一ZFN和第二ZFN,其中所述第一ZFN和第二ZFN每一者均有效连接至FokI核酸酶亚基,其中所述第一ZFN和第二ZFN识别DNA靶序列每条链中被约5至7bp间隔序列隔开的两条邻接DNA靶序列,并且其中所述FokI核酸酶亚基发生二聚化,从而生成能产生双链断裂的活性核酸酶。参见例如US20060246567、US20080182332、US20020081614、US20030021776、WO/2002/057308A2、US20130123484、US20100291048、WO/2011/017293A2,以及Gaj et al.(2013)Trends in Biotechnology,31(7):397-405(Gaj等人,2013年,《生物技术趋势》,第31卷,第7期,第397-405页);这些文献各自以引用方式并入本文。
在又一个实施例中,核酸酶试剂为大范围核酸酶。已基于保守序列基序将大范围核酸酶分类为四个家族,这些家族是LAGLIDADG、GIY-YIG、H-N-H和His-Cys框家族。这些基序参与金属离子的配位和磷酸二酯键的水解。大范围核酸酶以其长识别位点及耐受其DNA底物中的一些序列多态性而著称。大范围核酸酶的结构域、结构和功能是已知的,参见例如,Guhan and Muniyappa(2003)Crit Rev Biochem Mol Biol 38:199-248(Guhan和Muniyappa,2003年,《生物化学与分子生物学评论》,第38卷,第199-248页);Lucas et al.,(2001)Nucleic Acids Res 29:960-9(Lucas等人,2001年,《核酸研究》,第29卷,第960-969页);Jurica and Stoddard,(1999)Cell Mol Life Sci 55:1304-26(Jurica和Stoddard,1999年,《细胞和分子生命科学》,第55卷,第1304-1326页);Stoddard,(2006)Q RevBiophys 38:49-95(Stoddard,2006年,《生物物理学季评》,第38卷,第49-95页);以及Moureet al.,(2002)Nat Struct Biol 9:764(Moure等人,2002年,《自然结构生物学》,第9卷,第764页)。在一些例子中,使用天然存在的变体和/或经改造的衍生大范围核酸酶。用于调整动力学、辅因子相互作用、表达、最适条件和/或识别位点特异性以及筛选活性的方法是已知的,参见例如,Epinat et al.,(2003)Nucleic Acids Res 31:2952-62(Epinat等人,2003年,《核酸研究》,第31卷,第2952-2962页);Chevalier et al.,(2002)Mol Cell 10:895-905(Chevalier等人,2002年,《分子细胞》,第10卷,第895-905页);Gimble et al.,(2003)Mol Biol 334:993-1008(Gimble等人,2003年,《分子生物学》,第334卷,第993-1008页);Seligman et al.,(2002)Nucleic Acids Res 30:3870-9(Seligman等人,2002年,《核酸研究》,第30卷,第3870-3879页);Sussman et al.,(2004)J Mol Biol 342:31-41(Sussman等人,2004年,《分子生物学杂志》,第342卷,第31-41页);Rosen et al.,(2006)Nucleic Acids Res 34:4791-800(Rosen等人,2006年,《核酸研究》,第34卷,第4791-4800页);Chames et al.,(2005)Nucleic Acids Res 33:e178(Chames等人,2005年,《核酸研究》,第33卷,第e178页);Smith et al.,(2006)Nucleic Acids Res 34:e149(Smith等人,2006年,《核酸研究》,第34卷,第e149页);Gruen et al.,(2002)Nucleic Acids Res 30:e29(Gruen等人,2002年,《核酸研究》,第30卷,第e29页);Chen and Zhao,(2005)NucleicAcids Res 33:e154(Chen和Zhao,2005年,《核酸研究》,第33卷,第e154页);WO2005105989、WO2003078619、WO2006097854、WO2006097853、WO2006097784,以及WO2004031346。
可在本文中使用任何大范围核酸酶,包括但不限于I-SceI、I-SceII、I-SceIII、I-SceIV、I-SceV、I-SceVI、I-SceVII、I-CeuI、I-CeuAIIP、I-CreI、I-CrepsbIP、I-CrepsbIIP、I-CrepsbIIIP、I-CrepsbIVP、I-TliI、I-PpoI、PI-PspI、F-SceI、F-SceII、F-SuvI、F-TevI、F-TevII、I-AmaI、I-AniI、I-ChuI、I-CmoeI、I-CpaI、I-CpaII、I-CsmI、I-CvuI、I-CvuAIP、I-DdiI、I-DdiII、I-DirI、I-DmoI、I-HmuI、I-HmuII、I-HsNIP、I-LlaI、I-MsoI、I-NaaI、I-NanI、I-NcIIP、I-NgrIP、I-NitI、I-NjaI、I-Nsp236IP、I-PakI、I-PboIP、I-PcuIP、I-PcuAI、I-PcuVI、I-PgrIP、I-PobIP、I-PorI、I-PorIIP、I-PbpIP、I-SpBetaIP、I-ScaI、I-SexIP、I-SneIP、I-SpomI、I-SpomCP、I-SpomIP、I-SpomIIP、I-SquIP、I-Ssp6803I、I-SthPhiJP、I-SthPhiST3P、I-SthPhiSTe3bP、I-TdeIP、I-TevI、I-TevII、I-TevIII、I-UarAP、I-UarHGPAIP、I-UarHGPA13P、I-VinIP、I-ZbiIP、PI-MtuI、PI-MtuHIPPI-MtuHIIP、PI-PfuI、PI-PfuII、PI-PkoI、PI-PkoII、PI-Rma43812IP、PI-SpBetaIP、PI-SceI、PI-TfuI、PI-TfuII、PI-ThyI、PI-TliI、PI-TliII,或它们的任何活性变体或片段。
在一个实施例中,所述大范围核酸酶识别12至40个碱基对的双链DNA序列。在一个实施例中,所述大范围核酸酶识别基因组中的一个完全匹配的靶序列。在一个实施例中,所述大范围核酸酶为归巢核酸酶。在一个实施例中,所述归巢核酸酶为归巢核酸酶的LAGLIDADG家族。在一个实施例中,归巢核酸酶的LAGLIDADG家族选自I-SceI、I-CreI和I-Dmol。
核酸酶试剂还可包括限制性内切核酸酶,它们包括I型、II型、III型和IV型内切核酸酶。I型和III型限制性内切核酸酶识别特异性识别位点,但通常在离核酸酶结合位点的可变位置处裂解,该核酸酶结合位点离裂解位点(识别位点)可达数百个碱基对。在II型系统中,限制性酶切活性独立于任何甲基化酶活性,并且通常在结合位点之内或附近的特异性位点处发生裂解。大多数II型酶切割回文序列,但是IIa型酶识别非回文识别位点并在识别位点之外裂解,IIb型酶在识别位点之外的两个位点处切割序列两次,并且IIs型酶识别非对称识别位点并在一侧且离识别位点约1至20个核苷酸的限定距离处裂解。IV型限制性酶靶向甲基化DNA。限制性酶进一步在例如REBASE数据库中进行说明和分类(地址为rebase.neb.com的网页;Roberts et al.,(2003)Nucleic Acids Res 31:418-20(Roberts等人,2003年,《核酸研究》,第31卷,第418-420页),Roberts et al.,(2003)Nucleic AcidsRes 31:1805-12(Roberts等人,2003年,《核酸研究》,第31卷,第1805-1812页),以及Belfort et al.,(2002)in Mobile DNA II,pp.761-783,Eds.Craigie et al.,(ASMPress,Washington,DC)(Belfort等人,2002年,载于《可移动的DNA II》,第761-783页,Craigie等人编辑,华盛顿特区ASM出版社))。
在各种方法和组合物中采用的核酸酶试剂还可包括CRISPR/Cas系统。此类系统可采用Cas9核酸酶,其在一些情况下针对要在其中表达的所需细胞类型进行密码子优化。该系统还采用融合的crRNA-tracrRNA构建体,该构建体与经密码子优化的Cas9一起发挥作用。该单一RNA通常称为向导RNA或gRNA。在gRNA内,crRNA部分被确定为给定识别位点的“靶序列”,并且tracrRNA通常称为“支架”。已证实该系统可在多种真核细胞和原核细胞中发挥作用。简而言之,包含靶序列的短DNA片段被插入到向导RNA表达质粒中。gRNA表达质粒包含靶序列(在一些实施例中约20个核苷酸)、一种形式的tracrRNA序列(支架),以及在细胞中有活性的合适启动子和用于在真核细胞中正确加工的必要元件。这些系统中的多种系统依赖于定制的互补寡核苷酸,这些寡核苷酸退火而形成双链DNA,接着克隆到gRNA表达质粒中。然后将gRNA表达盒与Cas9表达盒引入细胞。参见例如Mali P et al.(2013)Science2013Feb 15;339(6121):823-6(Mali P等人,2013年,《科学》,2013年2月15日,第339卷,第6121期,第823-826页);Jinek M et al.Science 2012Aug 17;337(6096):816-21(Jinek M等人,《科学》,2012年8月17日,第337卷,第6096期,第816-821页);Hwang WY et al.NatBiotechnol 2013Mar;31(3):227-9(Hwang WY等人,《自然生物技术》,2013年3月,第31卷,第3期,第227-229页);Jiang W et al.Nat Biotechnol 2013Mar;31(3):233-9(Jiang W等人,《自然生物技术》,2013年3月,第31卷,第3期,第233-239页);以及Cong L etal.Science 2013Feb 15;339(6121):819-23(Cong L等人,《科学》,2013年2月15日,第339卷,第6121期,第819-823页),这些文献各自以引用方式并入本文。
本文所公开的方法和组合物可利用成簇规律间隔短回文重复序列(CRISPR)/CRISPR相关(Cas)系统或此类系统的组分来修饰细胞内的基因组。CRISPR/Cas系统包括参与Cas基因的表达或指导Cas基因的活性的转录物和其他元件。CRISPR/Cas系统可为I型、II型或III型系统。本文所公开的方法和组合物通过利用CRISPR复合物(包含与Cas蛋白复合的向导RNA(gRNA))来采用CRISPR/Cas系统对核酸进行定点裂解。
用于本文所公开的方法中的一些CRISPR/Cas系统为非天然存在的。“非天然存在的”系统包括指示受到人工干预的任何系统,诸如系统的一种或多种组分从其天然存在的状态改变或突变,至少基本上不含其在自然界中与之天然关联的至少一种其他组分,或和其不与之天然关联的至少一种其他组分相关联。例如,一些CRISPR/Cas系统采用非天然存在的CRISPR复合物,这些复合物包含在天然情况下不会同时存在的gRNA和Cas蛋白。
Cas蛋白一般包含至少一个RNA识别或结合结构域。此类结构域可与向导RNA(gRNA,下文更详细说明)相互作用。Cas蛋白还可包含核酸酶结构域(例如,DNA酶或RNA酶结构域)、DNA结合结构域、解旋酶结构域、蛋白-蛋白相互作用结构域、二聚化结构域以及其他结构域。核酸酶结构域具有用于核酸裂解的催化活性。裂解包括核酸分子共价键的断裂。裂解可产生平头末端或交错末端,并且其可为单链或双链裂解。
Cas蛋白的例子包括Cas1、Cas1B、Cas2、Cas3、Cas4、Cas5、Cas5e(CasD)、Cas6、Cas6e、Cas6f、Cas7、Cas8a1、Cas8a2、Cas8b、Cas8c、Cas9(Csn1或Csx12)、Cas10、Casl0d、CasF、CasG、CasH、Csy1、Csy2、Csy3、Cse1(CasA)、Cse2(CasB)、Cse3(CasE)、Cse4(CasC)、Csc1、Csc2、Csa5、Csn2、Csm2、Csm3、Csm4、Csm5、Csm6、Cmr1、Cmr3、Cmr4、Cmr5、Cmr6、Csb1、Csb2、Csb3、Csx17、Csx14、Csx10、Csx16、CsaX、Csx3、Csx1、Csx15、Csf1、Csf2、Csf3、Csf4和Cu1966,以及它们的同源物或修饰形式。
Cas蛋白可来自II型CRISPR/Cas系统。例如,Cas蛋白可为Cas9蛋白或衍生自Cas9蛋白。这些Cas9蛋白通常共用具有保守架构的四个关键基序。基序1、2和4为RuvC样基序,基序3为HNH基序。Cas9蛋白可来自例如化脓性链球菌(Streptococcus pyogenes)、嗜热链球菌(Streptococcus thermophilus)、链球菌属物种(Streptococcus sp.)、金黄色葡萄球菌(Staphylococcus aureus)、达氏拟诺卡氏菌(Nocardiopsis dassonvillei)、始旋链霉菌(Streptomyces pristinaespiralis)、绿色产色链霉菌(Streptomycesviridochromogenes)、绿色产色链霉菌(Streptomyces viridochromogenes)、粉红链孢囊菌(Streptosporangium roseum)、粉红链孢囊菌(Streptosporangium roseum)、酸热脂环酸芽孢杆菌(AlicyclobacHlus acidocaldarius)、假蕈状芽孢杆菌(Bacilluspseudomycoides)、还原硒酸盐芽孢杆菌(Bacillus selenitireducens)、西伯利亚微小杆菌(Exiguobacterium sibiricum)、德氏乳杆菌(Lactobacillus delbrueckii)、唾液乳杆菌(Lactobacillus salivarius)、海洋微颤菌(Microscilla marina)、伯克氏菌(Burkholderiales bacterium)、萘降解极地单胞菌(Polaromonas naphthalenivorans)、极地单胞菌属物种(Polaromonas sp.)、瓦氏鳄球藻(Crocosphaera watsonii)、蓝杆藻属物种(Cyanothece sp.)、铜绿微囊藻(Microcystis aeruginosa)、聚球藻属物种(Synechococcus sp.)、阿拉伯糖醋盐杆菌(Acetohalobium arabaticum)、制氨菌(Ammonifex degensii)、热解纤维素菌(Caldicelulosiruptor becscii)、CandidatusDesulforudis、肉毒梭菌(Clostridium botulinum)、艰难梭菌(Clostridium difficile)、大芬戈尔德菌(Finegoldia magna)、嗜热盐碱厌氧菌(Natranaerobius thermophilus)、丙酸互营细菌(Pelotomaculum thermopropionicum)、喜温嗜酸硫杆菌(Acidithiobacilluscaldus)、嗜酸氧化亚铁硫杆菌(Acidithiobacillus ferrooxidans)、紫色硫细菌(Allochromatium vinosum)、海杆菌属物种(Marinobacter sp.)、嗜盐亚硝化球菌(Nitrosococcus halophilus)、瓦氏亚硝化球菌(Nitrosococcus watsoni)、游海假交替单胞菌(Pseudoalteromonas haloplanktis)、纤线杆菌(Ktedonobacter racemifer)、甲烷盐菌(Methanohalobium evestigatum)、多变鱼腥藻Anabaena variabilis)、泡沫节球藻(Nodularia spumigena)、念珠藻属物种(Nostoc sp.)、极大节螺藻(Arthrospiramaxima)、钝顶节螺藻(Arthrospira platensis)、节螺藻属物种(Arthrospira sp.)、鞘丝藻属物种(Lyngbya sp.)、原型微鞘藻(Microcoleus chthonoplastes)、颤藻属物种(Oscillatoria sp.)、运动石袍菌(Petrotoga mobilis)、非洲栖热腔菌(Thermosiphoafricanus)或深海单细胞蓝细菌(Acaryochloris marina)。Cas9家族成员的附加例子在WO2014/131833中有所描述,该专利全文以引用方式并入本文。来自化脓性链球菌(S.pyogenes)或从其衍生的Cas9蛋白是优选的酶。来自化脓性链球菌的Cas9蛋白被分配了SwissProt登记号Q99ZW2。
Cas蛋白可为野生型蛋白(即,自然界存在的蛋白)、经修饰的Cas蛋白(即,Cas蛋白变体),或者野生型或经修饰的Cas蛋白的片段。Cas蛋白也可以是野生型或经修饰的Cas蛋白的活性变体或片段。活性变体或片段可与野生型或经修饰的Cas蛋白或者其一部分至少具有80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或更高的序列同一性,其中所述活性变体保留了在所需裂解位点处切割的能力,从而保留了切口诱导活性或双链断裂诱导活性。对切口诱导活性或双链断裂诱导活性的测定法是已知的,并且一般测量Cas蛋白对包含裂解位点的DNA底物的总体活性和特异性。
可修饰Cas蛋白以提高或降低核酸结合亲和力、核酸结合特异性和/或酶活性。还可修饰Cas蛋白以改变蛋白的任何其他活性或特性,诸如稳定性。例如,Cas蛋白的一个或多个核酸酶结构域可以被修饰、缺失或失活,或者Cas蛋白可以被截短以去除对于蛋白质的功能并非必要的结构域、或优化(例如,增强或降低)Cas蛋白的活性。
一些Cas蛋白包含至少两个核酸酶结构域,诸如DNA酶结构域。例如,Cas9蛋白可包含RuvC样核酸酶结构域和HNH样核酸酶结构域。RuvC结构域和HNH结构域各自可切割双链DNA的不同链,从而在DNA中产生双链断裂。参见例如Jinek et al.(2012)Science 337:816-821(Jinek等人,2012年,《科学》,第337卷,第816-821页),该文献全文据此以引用方式并入。
这些核酸酶结构域中的一者或两者可以被缺失或突变,使得它们不再有功能或具有降低的核酸酶活性。如果这两个核酸酶结构域之一被缺失或突变,则所得的Cas蛋白(例如,Cas9)可称为切口酶,并且可在双链DNA内的CRISPR RNA识别序列处生成单链断裂,而不会生成双链断裂(即,其可裂解互补链或非互补链,但无法同时裂解两者)。如果这两个核酸酶结构域都被缺失或突变,则所得的Cas蛋白(例如,Cas9)裂解双链DNA两条链的能力会降低。将Cas9转变为切口酶的突变的例子是来自化脓性链球菌的Cas9的RuvC结构域中的D10A(Cas9的第10位处天冬氨酸至丙氨酸)突变。同样,来自化脓性链球菌的Cas9的HNH结构域中的H939A(氨基酸位置839处组氨酸至丙氨酸)或H840A(氨基酸位置840处组氨酸至丙氨酸)可将Cas9转变为切口酶。将Cas9转变为切口酶的突变的其他例子包括来自嗜热链球菌(S.thermophilus)的Cas9的对应突变。参见例如Sapranauskas et al.(2011)NucleicAcids Research 39:9275-9282(Sapranauskas等人,2011年,《核酸研究》,第39卷,第9275-9282页)和WO 2013/141680,这些文献每一者的全文均以引用方式并入本文。此类突变可使用诸如定点诱变、PCR介导的诱变或全基因合成的方法来生成。其他形成切口酶的突变的例子可见于例如WO/2013/176772A1和WO/2013/142578A1中,这些专利各自以引用方式并入本文。
Cas蛋白也可为融合蛋白。例如,Cas蛋白可融合到裂解结构域、表观遗传修饰结构域、转录激活结构域或转录阻遏物结构域。参见WO 2014/089290,该专利全文以引用方式并入本文。Cas蛋白也可融合到异源多肽,从而提供增强或减弱的稳定性。融合的结构域或异源多肽可位于N端、C端或Cas蛋白的内部。
Cas蛋白可融合到有助于亚细胞定位的异源多肽。此类异源肽包括例如用于靶向细胞核的核定位信号(NLS)诸如SV40 NLS、用于靶向线粒体的线粒体定位信号、ER滞留信号,等等。参见例如Lange et al.(2007)J.Biol.Chem.282:5101-5105(Lange等人,2007年,《生物化学杂志》,第282卷,第5101-5105页)。此类亚细胞定位信号可位于N端、C端或Cas蛋白内的任何位置处。NLS可包含一段碱性氨基酸,并且可为单分型序列或双分型序列。
Cas蛋白也可连接至细胞穿透结构域。例如,细胞穿透结构域可衍生自HIV-1TAT蛋白、来自人乙肝病毒的TLM细胞穿透基序、MPG、Pep-1、VP22、来自单纯性疱疹病毒的细胞穿透肽,或多聚精氨酸肽序列。参见例如WO 2014/089290,该专利全文以引用方式并入本文。细胞穿透结构域可位于N端、C端或Cas蛋白内的任何位置处。
Cas蛋白还可包含便于示踪或纯化的异源多肽,诸如荧光蛋白、纯化标签或表位标签。荧光蛋白的例子包括绿色荧光蛋白(例如,GFP、GFP-2、tagGFP、turboGFP、eGFP、Emerald、Azami Green、Monomeric Azami Green、CopGFP、AceGFP、ZsGreenl)、黄色荧光蛋白(例如,YFP、eYFP、Citrine、Venus、YPet、PhiYFP、ZsYellowl)、蓝色荧光蛋白(例如,eBFP、eBFP2、Azurite、mKalamal、GFPuv、Sapphire、T-sapphire)、青色荧光蛋白(例如,eCFP、Cerulean、CyPet、AmCyanl、Midoriishi-Cyan)、红色荧光蛋白(mKate、mKate2、mPlum、DsRedmonomer、mCherry、mRFP1、DsRed-Express、DsRed2、DsRed-Monomer、HcRed-Tandem、HcRedl、AsRed2、eqFP611、mRaspberry、mStrawberry、Jred)、橙色荧光蛋白(mOrange、mKO、Kusabira-Orange、Monomeric Kusabira-Orange、mTangerine、tdTomato),以及任何其他合适的荧光蛋白。标签的例子包括谷胱甘肽-S-转移酶(GST)、几丁质结合蛋白(CBP)、麦芽糖结合蛋白、硫氧还蛋白(TRX)、多聚(NANP)、串联亲和纯化(TAP)标签、myc、AcV5、AU1、AU5、E、ECS、E2、FLAG、血凝素(HA)、nus、Softag 1、Softag 3、Strep、SBP、Glu-Glu、HSV、KT3、S、S1、T7、V5、VSV-G、组氨酸(His)、生物素羧基载体蛋白(BCCP)以及钙调蛋白。
Cas蛋白可以任何形式提供。例如,Cas蛋白可以蛋白的形式提供,诸如与gRNA复合的Cas蛋白。另选地,Cas蛋白可以编码Cas蛋白的核酸的形式提供,诸如RNA(例如,信使RNA(mRNA))或DNA。任选地,编码Cas蛋白的核酸可进行密码子优化,以便在特定的细胞或生物体中有效地翻译成蛋白。
编码Cas蛋白的核酸可稳定地整合在细胞的基因组中,并有效连接至细胞中有活性的启动子。另选地,编码Cas蛋白的核酸可有效连接至表达构建体中的启动子。表达构建体包括能够指导目标基因或其他核酸序列(例如,Cas基因)的表达并且可将这种目标核酸序列转移到靶细胞中的任何核酸构建体。可用于表达构建体的启动子包括例如在大鼠、真核、哺乳动物、非人哺乳动物、人、啮齿动物、小鼠或仓鼠多能细胞中有活性的启动子。其他启动子的例子在本文别处有所描述。
“向导RNA”或“gRNA”包括结合于Cas蛋白并使Cas蛋白靶向靶DNA内的特定位置的RNA分子。向导RNA可包含两个区段:“DNA靶向区段”和“蛋白结合区段”。“区段”包括分子的区段、部分或区域,诸如RNA中的一个邻接核苷酸段。一些gRNA包含两个单独的RNA分子:“激活因子-RNA”和“靶向因子-RNA”。其他gRNA为单个RNA分子(单条RNA多核苷酸),其也可称为“单分子gRNA”、“单向导RNA”或“sgRNA”。参见例如WO/2013/176772A1、WO/2014/065596A1、WO/2014/089290A1、WO/2014/093622A2、WO/2014/099750A2、WO/2013142578A1以及WO2014/131833A1,这些专利各自以引用方式并入本文。术语“向导RNA”和“gRNA”包括双分子gRNA和单分子gRNA两者。
示例性双分子gRNA包含crRNA样(“CRISPR RNA”或“靶向因子-RNA”或“crRNA”或“crRNA重复序列”)分子以及对应的tracrRNA样(“反式作用CRISPR RNA”或“激活因子-RNA”或“tracrRNA”或“支架”)分子。crRNA包含gRNA的DNA靶向区段(单链)和一段核苷酸,这段核苷酸形成gRNA的蛋白结合区段的dsRNA双链体的一半。
对应的tracrRNA(激活因子-RNA)包含一段核苷酸,这段核苷酸形成gRNA的蛋白结合区段的dsRNA双链体的另一半。crRNA的一段核苷酸与tracrRNA的一段核苷酸互补并杂交,从而形成gRNA的蛋白结合结构域的dsRNA双链体。因此,每个crRNA可以说成具有对应的tracrRNA。
crRNA和对应的tracrRNA杂交而形成gRNA。crRNA另外提供了与CRISPR RNA识别序列杂交的单链DNA靶向区段。如果用于细胞内的修饰,则给定crRNA或tracrRNA分子的准确序列可被设计成对于将在其中使用这些RNA分子的物种具有特异性。参见例如Mali et al.(2013)Science 339:823-826(Mali等人,2013年,《科学》,第339卷,第823-826页);Jineket al.(2012)Science 337:816-821(Jinek等人,2012年,《科学》,第337卷,第816-821页);Hwang et al.(2013)Nat.Biotechnol.31:227-229(Hwang等人,2013年,《自然生物技术》,第31卷,第227-229页);Jiang et al.(2013)Nat.Biotechnol.31:233-239(Jiang等人,2013年,《自然生物技术》,第31卷,第233-239页);以及Cong et al.(2013)Science 339:819-823(Cong等人,2013年,《科学》,第339卷,第819-823页),这些文献各自以引用方式并入本文。
给定gRNA的DNA靶向区段(crRNA)包含与靶DNA中的序列互补的核苷酸序列。gRNA的DNA靶向区段通过杂交(即,碱基配对)以序列特异性方式与靶DNA相互作用。因此,DNA靶向区段的核苷酸序列可能有变,并且决定gRNA和靶DNA将与之相互作用的靶DNA内的位置。可修饰对象gRNA的DNA靶向区段,以与靶DNA内的任何所需序列杂交。天然存在的crRNA因Cas9系统和生物体不同而异,但通常包含21至72个核苷酸长的靶向区段,该靶向区段侧接21至46个核苷酸长的两个正向重复序列(DR)(参见例如WO2014/131833)。就化脓性链球菌而言,DR为36个核苷酸长,并且靶向区段为30个核苷酸长。位于3’的DR与对应的tracrRNA互补并杂交,该tracrRNA又结合于Cas9蛋白。
DNA靶向区段的长度可为约12个核苷酸至约100个核苷酸。例如,DNA靶向区段的长度可为约12个核苷酸(nt)至约80nt、约12nt至约50nt、约12nt至约40nt、约12nt至约30nt、约12nt至约25nt、约12nt至约20nt、或约12nt至约19nt。另选地,DNA靶向区段的长度可为约19nt至约20nt、约19nt至约25nt、约19nt至约30nt、约19nt至约35nt、约19nt至约40nt、约19nt至约45nt、约19nt至约50nt、约19nt至约60nt、约19nt至约70nt、约19nt至约80nt、约19nt至约90nt、约19nt至约100nt、约20nt至约25nt、约20nt至约30nt、约20nt至约35nt、约20nt至约40nt、约20nt至约45nt、约20nt至约50nt、约20nt至约60nt、约20nt至约70nt、约20nt至约80nt、约20nt至约90nt、或约20nt至约100nt。
DNA靶向区段内与靶DNA的核苷酸序列(CRISPR RNA识别序列)互补的核苷酸序列的长度可为至少约12nt。例如,DNA靶向序列(即,DNA靶向区段内与靶DNA内的CRISPR RNA识别序列互补的序列)的长度可为至少约12nt、至少约15nt、至少约18nt、至少约19nt、至少约20nt、至少约25nt、至少约30nt、至少约35nt、或至少约40nt。另选地,DNA靶向序列的长度可为约12个核苷酸(nt)至约80nt、约12nt至约50nt、约12nt至约45nt、约12nt至约40nt、约12nt至约35nt、约12nt至约30nt、约12nt至约25nt、约12nt至约20nt、约12nt至约19nt、约19nt至约20nt、约19nt至约25nt、约19nt至约30nt、约19nt至约35nt、约19nt至约40nt、约19nt至约45nt、约19nt至约50nt、约19nt至约60nt、约20nt至约25nt、约20nt至约30nt、约20nt至约35nt、约20nt至约40nt、约20nt至约45nt、约20nt至约50nt、或约20nt至约60nt。在一些情况下,DNA靶向序列的长度可为约20nt。
TracrRNA可为任何形式(例如,全长tracrRNA或活性的部分tracrRNA)并具有不同长度。它们可包括初级转录物或加工形式。例如,tracrRNA(作为单向导RNA的一部分或作为双分子gRNA一部分形式的单独分子)可包含以下部分或由以下部分组成:野生型tracrRNA序列的全部或一部分(例如,野生型tracrRNA序列的约或大于约20、26、32、45、48、54、63、67、85个或更多个核苷酸)。来自化脓性链球菌的野生型tracrRNA序列的例子包括171个核苷酸、89个核苷酸、75个核苷酸以及65个核苷酸的形式。参见例如Deltcheva et al.(2011)Nature 471:602-607(Deltcheva等人,2011年,《自然》,第471卷,第602-607页);WO 2014/093661,这些文献每一者的全文均以引用方式并入本文。单向导RNA(sgRNA)内的tracrRNA的例子包括存在于+48、+54、+67和+85形式的sgRNA内的tracrRNA区段,其中“+n”指示野生型tracrRNA的至多+n核苷酸包含在sgRNA中。参见US 8,697,359,该专利全文以引用方式并入本文。
DNA靶向序列与靶DNA内的CRISPR RNA识别序列之间的互补性百分比可为至少60%(例如,至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少95%、至少97%、至少98%、至少99%或100%)。DNA靶向序列与靶DNA内的CRISPR RNA识别序列之间的互补性百分比在约20个邻接核苷酸内可为至少60%。例如,DNA靶向序列与靶DNA内的CRISPR RNA识别序列之间的互补性百分比在靶DNA的互补链内的CRISPR RNA识别序列的5’端的14个邻接核苷酸内为100%,而在其余部分内低至0%。在这种情况下,DNA靶向序列可被视为14个核苷酸长。又如,DNA靶向序列与靶DNA内的CRISPR RNA识别序列之间的互补性百分比在靶DNA的互补链内的CRISPR RNA识别序列的5’端的7个邻接核苷酸内为100%,而在其余部分内低至0%。在这种情况下,DNA靶向序列可被视为7个核苷酸长。
gRNA的蛋白结合区段可包含彼此互补的两段核苷酸。所述蛋白结合区段的两条互补核苷酸杂交而形成双链RNA双链体(dsRNA)。对象gRNA的蛋白结合区段与Cas蛋白相互作用,并且gRNA经由DNA靶向区段将结合的Cas蛋白引导至靶DNA内的特异性核苷酸序列。
向导RNA可包括提供额外所需特征(例如,经改良或调控的稳定性;亚细胞靶向;用荧光标记示踪;蛋白或蛋白复合物的结合位点;等等)的修饰或序列。此类修饰的例子包括例如5'帽(例如,7-甲基鸟苷酸帽(m7G));3'多聚腺苷酸化尾(即,3'多聚(A)尾);核糖开关序列(例如,以实现经调控的稳定性和/或经调控的蛋白和/或蛋白复合物可及性);稳定性控制序列;形成dsRNA双链体(即,发夹)的序列);使RNA靶向亚细胞位置(例如,细胞核、线粒体、叶绿体等)的修饰或序列;提供示踪的修饰或序列(例如,与荧光分子的直接缀合、与有利于荧光检测的部分的缀合、允许荧光检测的序列等);为蛋白(例如,作用于DNA的蛋白,包括转录激活因子、转录阻遏物、DNA甲基转移酶、DNA去甲基化酶、组蛋白乙酰转移酶、组蛋白去乙酰化酶等)提供结合位点的修饰或序列;以及它们的组合。
向导RNA可以任何形式提供。例如,gRNA可以RNA的形式(作为两个分子(单独的crRNA和tracrRNA)或作为一个分子(sgRNA))提供,并任选地以与Cas蛋白的复合物的形式提供。gRNA也可以编码RNA的DNA形式提供。编码gRNA的DNA可编码单个RNA分子(sgRNA)或单独的RNA分子(例如,单独的crRNA和tracrRNA)。在后一种情况下,编码gRNA的DNA可作为分别编码crRNA和tracrRNA的单独DNA分子提供。
编码gRNA的DNA可稳定地整合在细胞的基因组中,并有效连接至细胞中有活性的启动子。另选地,编码gRNA的DNA可有效连接至表达构建体中的启动子。此类启动子可例如在大鼠、真核、哺乳动物、非人哺乳动物、人、啮齿动物、小鼠或仓鼠多能细胞中有活性。在一些情况下,所述启动子为RNA聚合酶III启动子,诸如人U6启动子、大鼠U6聚合酶III启动子或小鼠U6聚合酶III启动子。其他启动子的例子在本文别处有所描述。
另选地,可通过各种其他方法制备gRNA。例如,可通过采用例如T7 RNA聚合酶的体外转录来制备gRNA(参见例如WO 2014/089290和WO 2014/065596)。向导RNA也可为通过化学合成制备的合成产生的分子。
术语“CRISPR RNA识别序列”包括只要存在结合的充分条件,gRNA的DNA靶向区段就将与之结合的、存在于靶DNA中的核酸序列。例如,CRISPR RNA识别序列包括向导RNA被设计成与之具有互补性的序列,其中CRISPR RNA识别序列与DNA靶向序列之间的杂交促进CRISPR复合物形成。完全互补性不是必需的,只要存在引起杂交并促进CRISPR复合物形成的充分互补性即可。CRISPR RNA识别序列还包括下文更详细说明的Cas蛋白裂解位点。CRISPR RNA识别序列可包含任何多核苷酸,所述多核苷酸可位于例如细胞的细胞核或细胞质中、或细胞的细胞器(诸如线粒体或叶绿体)内。
靶DNA内的CRISPR RNA识别序列可被Cas蛋白或gRNA靶向(即,被Cas蛋白或gRNA结合,或者与Cas蛋白或gRNA杂交,或者与Cas蛋白或gRNA互补)。合适的DNA/RNA结合条件包括通常存在于细胞中的生理条件。其他合适的DNA/RNA结合条件(例如,无细胞系统中的条件)是本领域已知的(参见例如Molecular Cloning:A Laboratory Manual,3rd Ed.(Sambrooket al.,Harbor Laboratory Press 2001)(《分子克隆实验指南》,第3版,Sambrook等人,冷泉港实验室出版社,2001年))。与Cas蛋白或gRNA互补并杂交的靶DNA链可称为“互补链”,而与“互补链”互补(因此不与Cas蛋白和gRNA互补)的靶DNA链可称为“非互补链”或“模板链”。
Cas蛋白可在gRNA的DNA靶向区段将与之结合的、存在于靶DNA中的核酸序列之内或之外的位点处裂解核酸。“裂解位点”包括Cas蛋白产生单链断裂或双链断裂的核酸位置。例如,CRISPR复合物(包含与CRISPR RNA识别序列杂交并与Cas蛋白复合的gRNA)的形成,可导致gRNA的DNA靶向区段将与之结合的、存在于靶DNA中的核酸序列之中或附近(例如,在相距1、2、3、4、5、6、7、8、9、10、20、50个或更多个碱基对内)的一条或两条链裂解。如果裂解位点位于gRNA的DNA靶向区段将与之结合的核酸序列之外,则裂解位点仍被视为在“CRISPRRNA识别序列”内。裂解位点可只位于核酸的一条链上,或可位于核酸的两条链上。裂解位点可位于核酸的两条链上的相同位置处(产生平头末端),或可位于每条链上的不同位点处(产生交错末端)。可例如通过使用两种Cas蛋白来产生交错末端,每种Cas蛋白在每条链上的不同裂解位点处产生单链断裂,从而产生双链断裂。例如,第一切口酶可在双链DNA(dsDNA)的第一链上形成单链断裂,而第二切口酶可在dsDNA的第二链上形成单链断裂,因此形成悬垂序列。在一些情况下,第一链上的切口酶CRISPR RNA识别序列与第二链上的切口酶CRISPR RNA识别序列相隔至少2、3、4、5、6、7、8、9、10、15、20、25、30、40、50、75、100、250、500或1,000个碱基对。
Cas9对靶DNA的位点特异性裂解可在由以下两者决定的位置处发生:(i)gRNA与靶DNA之间的碱基配对互补性,以及(ii)靶DNA中的短基序,称为前间区序列邻近基序(PAM)。PAM可在CRISPR RNA识别序列的旁侧。任选地,CRISPR RNA识别序列可侧接PAM。例如,Cas9的裂解位点可为PAM序列上游或下游的约1至约10个、或者约2至约5个碱基对(例如,3个碱基对)。在一些情况下(例如,当使用来自化脓性链球菌的Cas9或密切相关的Cas9时),非互补链的PAM序列可为5'-N
CRISPR RNA识别序列的例子包括与gRNA的DNA靶向区段互补的DNA序列、或除PAM序列之外的这种DNA序列。例如,靶基序可为紧接在Cas蛋白所识别的NGG基序前面的含20个核苷酸的DNA序列(参见例如WO 2014/165825)。5’端的鸟嘌呤可有利于RNA聚合酶在细胞中进行转录。CRISPR RNA识别序列的其他例子可包括5’端处的两个鸟嘌呤核苷酸(例如,GGN
CRISPR RNA识别序列可为细胞内源或外源的任何核酸序列。CRISPR RNA识别序列可为编码基因产物(例如,蛋白)的序列或非编码序列(例如,调控序列),或者可包括这两者。
在一个实施例中,靶序列紧邻地侧接前间区序列邻近基序(PAM)序列。在一个实施例中,目标基因座包含SEQ ID NO:1的核苷酸序列。在一个实施例中,gRNA包含编码成簇规律间隔短回文重复序列(CRISPR)RNA(crRNA)和反式激活CRISPR RNA(tracrRNA)的第三核酸序列。在另一个实施例中,大鼠多能细胞的基因组包含与靶序列互补的靶DNA区。在一些此类方法中,Cas蛋白为Cas9。在一些实施例中,gRNA包括(a)SEQ ID NO:2的核酸序列的嵌合RNA;或(b)SEQ ID NO:3的核酸序列的嵌合RNA。在一些此类方法中,crRNA包含SEQ IDNO:4、SEQ ID NO:5或SEQ ID NO:6所示的序列。在一些此类方法中,tracrRNA包含SEQ IDNO:7或SEQ ID NO:8所示的序列。
还提供了核酸酶试剂的活性变体和片段(即,经改造的核酸酶试剂)。此类活性变体可与天然核酸酶试剂至少具有65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或更高的序列同一性,其中所述活性变体保留在所需识别位点处切割的能力,从而保留了切口或双链断裂诱导活性。例如,本文所述的任何核酸酶试剂可由天然内切核酸酶序列修饰而成,并且可设计成在不被天然核酸酶试剂识别的识别位点处识别并诱导切口或双链断裂。因此,在一些实施例中,经改造的核酸酶具有在与对应天然核酸酶试剂识别位点不同的识别位点处诱导切口或双链断裂的特异性。对切口或双链断裂诱导活性的测定法是已知的,并且一般测量内切核酸酶对包含识别位点的DNA底物的总体活性和特异性。
可通过本领域已知的任何方式将核酸酶试剂引入细胞。可将编码核酸酶试剂的多肽直接引入细胞。另选地,可将编码核酸酶试剂的多核苷酸引入细胞。将编码核酸酶试剂的多核苷酸引入细胞时,核酸酶试剂可在细胞内瞬时地、有条件地或组成性地表达。因此,编码核酸酶试剂的多核苷酸可包含在表达盒中,并有效连接至条件启动子、诱导型启动子、组成型启动子或组织特异性启动子。此类目标启动子在本文别处进一步详细讨论。另选地,将核酸酶试剂作为编码核酸酶试剂的mRNA引入细胞。
在具体实施例中,编码核酸酶试剂的多核苷酸稳定地整合在细胞的基因组中,并有效连接至细胞中有活性的启动子。在其他实施例中,编码核酸酶试剂的多核苷酸位于包含插入多核苷酸的同一靶向载体中,而在其他情况下,编码核酸酶试剂的多核苷酸位于与包含插入多核苷酸的靶向载体分离的载体或质粒中。
当通过引入编码核酸酶试剂的多核苷酸来向细胞提供核酸酶试剂时,可修饰这种编码核酸酶试剂的多核苷酸,以替换成与编码核酸酶试剂的天然存在的多核苷酸序列相比在目标细胞中具有更高使用频率的密码子。例如,可修饰编码核酸酶试剂的多核苷酸,以替换成与天然存在的多核苷酸序列相比在给定的目标原核或真核细胞(包括细菌细胞、酵母细胞、人细胞、非人细胞、哺乳动物细胞、啮齿动物细胞、小鼠细胞、大鼠细胞或任何其他目标宿主细胞)中具有更高使用频率的密码子。
用于修饰挑战性基因组座位或Y染色体基因座的非限制性方法包括在存在包含至少10kb核酸序列的大靶向载体(LTVEC)的情况下使染色体(即,Y染色体)暴露于Cas蛋白和CRISPR RNA,其中在暴露于Cas蛋白、CRISPR RNA和LTVEC后,染色体(即,Y染色体)被修饰成包含至少10kb的核酸序列。
该方法可采用本文所述的LTVEC或smallTVEC中的任何一种。在非限制性实施例中,LTVEC或smallTVEC包含至少20kb、至少30kb、至少40kb、至少50kb、至少60kb、至少70kb、至少80kb、至少90kb、至少100kb、至少150kb或至少200kb的核酸序列。在其他实施例中,LTVEC的5’和3’同源臂的总和为约10kb至约150kb、约10kb至约20kb、约20kb至约40kb、约40kb至约60kb、约60kb至约80kb、约80kb至约100kb、约100kb至约120kb、或约120kb至150kb。在另一个实施例中,smallTVEC的5’和3’同源臂的总和为约0.5kb、1kb、1.5kb、2kb、3kb、4kb、5kb、6kb、7kb、8kb、9kb、约0.5kb至约1kb、约1kb至约1.5kb、约1.5kb至约2kb、约2kb至约3kb、约3kb至约4kb、约4kb至约5kb、约5kb至约6kb、约6kb至约7kb、约8kb至约9kb,或为至少10kb。
还提供了用于修饰挑战性靶基因座或Y染色体上的靶基因组座位的方法,包括:(a)提供包含挑战性靶基因座或Y染色体上的靶基因组座位的哺乳动物细胞,其中所述靶基因组座位包含向导RNA(gRNA)靶序列;(b)向该哺乳动物细胞中引入:(i)包含第一核酸的大靶向载体(LTVEC),所述第一核酸侧接有与靶基因组座位同源的靶向臂,其中所述LTVEC为至少10kb;(ii)包含第一启动子的第一表达构建体,所述第一启动子有效连接至编码Cas蛋白的第二核酸,以及(iii)包含第二启动子的第二表达构建体,所述第二启动子有效连接至编码向导RNA(gRNA)的第三核酸,所述向导RNA包含杂交至gRNA靶序列和反式激活CRISPRRNA(tracrRNA)的核苷酸序列,其中所述第一启动子和第二启动子在该哺乳动物细胞中有活性;以及(c)鉴定在挑战性靶基因组座位处或在Y染色体上的靶基因组座位处包含靶向遗传修饰的经修饰的哺乳动物细胞。在具体实施例中,第一表达构建体和第二表达构建体位于单个核酸分子上。在其他实施例中,Y染色体的靶基因组座位为Sry基因座。
如上文概述,在一个实施例中,Cas蛋白可包括Cas9蛋白。在另一个实施例中,gRNA靶序列紧邻地侧接前间区序列邻近基序(PAM)序列。
该方法可采用本文所述的LTVEC或smallTVEC中的任何一种。在非限制性实施例中,LTVEC或smallTVEC为至少0.5kb、至少1kb、至少5kb、至少10kb、至少15kb、至少20kb、至少30kb、至少40kb、至少50kb、至少60kb、至少70kb、至少80kb、至少90kb、至少100kb、至少150kb或至少200kb。在其他实施例中,LTVEC的5’和3’同源臂的总和为约10kb至约150kb、约10kb至约20kb、约20kb至约40kb、约40kb至约60kb、约60kb至约80kb、约80kb至约100kb、约100kb至约120kb、或约120kb至150kb。
可对例如哺乳动物细胞、非人哺乳动物细胞、成纤维细胞、啮齿动物细胞、大鼠细胞、小鼠细胞或仓鼠细胞执行采用CRISPR/Cas系统的各种方法(或本文所公开的任何方法)。所述细胞可为多能细胞、诱导性多能干(iPS)细胞、小鼠胚胎干(ES)细胞、大鼠胚胎干(ES)细胞、人胚胎干(ES)细胞或发育受限的人祖细胞。
如下文详细讨论,在采用例如使用上文概述的CRISPR/Cas系统对非人多能细胞的挑战性基因组座位或Y染色体上的目标基因组座位(即,Sry基因座)进行修饰之后,可将所产生的经遗传修饰的非人多能细胞引入非人宿主胚胎;然后在代孕母体中孕育包含经修饰的多能细胞的非人宿主胚胎。代孕母体产生包含靶向遗传修饰的F0子代。在具体实施例中,靶向遗传修饰能够通过种系传递。
可在本文所公开的用于修饰Y染色体上的靶基因组座位或挑战性靶基因组座位的方法和组合物中使用各种选择标记。此类标记在本文别处公开,包括但不限于赋予对诸如G418、潮霉素、杀稻瘟菌素、新霉素或嘌呤霉素等抗生素的抗性的选择标记。编码选择标记的多核苷酸有效连接至细胞中有活性的启动子。此类表达盒及其各种调控组分在本文别处进一步详细讨论。
提供了可实现在Y染色体上的靶基因组座位或挑战性靶基因组座位处整合至少一个插入多核苷酸的各种方法和组合物。如本文所用,“Y染色体上的靶基因组座位”包含Y染色体上希望整合插入多核苷酸的任何DNA区段或区域。
要靶向的Y染色体上的基因组座位或挑战性靶基因组座位对于细胞可为天然的,或另选地可包含整合到细胞染色体中的异源或外源DNA区段。此类异源或外源DNA区段可包括转基因、表达盒、编码选择标记的多核苷酸,或基因组DNA的异源或外源区域。Y染色体上的靶基因组座位或挑战性靶基因组座位可包含所靶向基因组整合系统中的任何一种,包括例如识别位点、选择标记、此前整合的插入多核苷酸、编码核酸酶试剂的多核苷酸、启动子等。另选地,Y染色体上的靶基因组座位或挑战性靶基因组座位可位于适当宿主细胞中所含的酵母人工染色体(YAC)、细菌人工染色体(BAC)、人类人工染色体或任何其他经改造的基因组区域内。因此,在具体实施例中,Y染色体上的所靶向基因组座位或挑战性靶基因组座位可包含来自非人哺乳动物、非人细胞、啮齿动物、人、大鼠、小鼠、仓鼠、兔、猪、牛、鹿、绵羊、山羊、鸡、猫、狗、白鼬、灵长类动物(例如,狨猴、恒河猴)、家养哺乳动物或农业哺乳动物、或任何其他目标生物体、或它们的组合的天然、异源或外源基因组核酸序列。
Y染色体上的靶基因组座位的非限制性例子包括Sry基因、Uty基因、Eif2s3y基因、Ddx3y基因、基因、Ube1y基因、Tspy基因、Usp9y基因、Zfy1基因和Zfy2基因,以及Y染色体上包含Kdm5d基因、Eif2s3y基因、Tspy基因、Uty基因、Ddx3y基因和Usp9y基因的区域。Y染色体上的这种基因座可来自非人哺乳动物、哺乳动物、啮齿动物、人、大鼠、小鼠、仓鼠、兔、猪、牛、鹿、绵羊、山羊、鸡、猫、狗、白鼬、灵长类动物(例如,狨猴、恒河猴)、家养哺乳动物或农业哺乳动物、或任何其他目标生物体、或它们的组合。此类细胞包括多能细胞,包括例如诱导性多能干(iPS)细胞、小鼠胚胎干(ES)细胞、大鼠胚胎干(ES)细胞、人胚胎干(ES)细胞或发育受限的人祖细胞。
如本文别处所述,提供了包含Sry蛋白活性或水平降低的XY多能细胞和/或全能细胞(诸如XY ES细胞或iPS细胞)的各种方法和组合物。本文所述的修饰Y染色体上的基因组座位的各种方法也可用于将靶向的遗传修饰引入不位于Y染色体上的目标多核苷酸。
如上文概述,本文所提供的方法和组合物采用了单独的或与核酸酶试剂组合的靶向载体。按照惯例,“同源重组”用来指在同源区内的交叉位点处两个DNA分子之间的DNA片段交换。
如本文所用,术语“插入多核苷酸”包含希望在靶基因组座位处整合的DNA区段。在具体实施例中,靶基因组座位位于Y染色体上。在其他实施例中,靶基因组座位为挑战性基因组座位。在一个实施例中,插入多核苷酸包含一个或多个目标多核苷酸。在其他实施例中,插入多核苷酸可包含一个或多个表达盒。给定表达盒可包含目标多核苷酸、编码选择标记和/或报告基因的多核苷酸,以及影响表达的各种调控组分。可包含在插入多核苷酸内的目标多核苷酸、选择标记和报告基因的非限制性例子在本文别处详细讨论。
在具体实施例中,插入多核苷酸可包含基因组核酸。在一个实施例中,基因组核酸衍生自动物、小鼠、人、非人、啮齿动物、非人、大鼠、仓鼠、兔、猪、牛、鹿、绵羊、山羊、鸡、猫、狗、白鼬、灵长类动物(例如,狨猴、恒河猴)、家养哺乳动物或农业哺乳动物、鸟类、或任何其他目标生物体、或它们的组合。
在另外的实施例中,插入多核苷酸包含条件性等位基因。在一个实施例中,条件性等位基因为如US 2011/0104799中所述的多功能等位基因,该专利全文以引用方式并入。在具体实施例中,条件性等位基因包含:(a)相对于靶基因的转录呈有义取向的起动序列,以及呈有义或反义取向的药物选择盒;(b)呈反义取向的目标核苷酸序列(NSI)和倒转条件模块(COIN,其利用外显子断裂内含子和可倒转的基因诱捕样模块;参见例如US 2011/0104799,该专利全文以引用方式并入);以及(c)在暴露于第一重组酶后重组以形成条件性等位基因的可重组单元,所述条件性等位基因(i)缺乏起动序列和DSC,并且(ii)含有呈有义取向的NSI和呈反义取向的COIN。
插入多核苷酸可为约5kb至约200kb、约5kb至约10kb、约10kb至约20kb、约20kb至约30kb、约30kb至约40kb、约40kb至约50kb、约60kb至约70kb、约80kb至约90kb、约90kb至约100kb、约100kb至约110kb、约120kb至约130kb、约130kb至约140kb、约140kb至约150kb、约150kb至约160kb、约160kb至约170kb、约170kb至约180kb、约180kb至约190kb、约190kb至约200kb、约200kb至约250kb、约250kb至约300kb、约300kb至约350kb、或约350kb至约400kb。
在具体实施例中,插入多核苷酸包含侧接有位点特异性重组靶序列的核酸。已经认识到,尽管整条插入多核苷酸可侧接这种位点特异性重组靶序列,但该插入多核苷酸内的任何区域或单独的目标多核苷酸也可侧接此类位点。如本文所用,术语“重组位点”包括由位点特异性重组酶识别且可充当重组事件的底物的核苷酸序列。如本文所用,术语“位点特异性重组酶”包括可促进在重组位点之间的重组的一组酶,其中这两个重组位点在单个核酸分子内或在分离的核酸分子上物理分隔。位点特异性重组酶的例子包括但不限于Cre、Flp和Dre重组酶。位点特异性重组酶可通过任何方式引入细胞,包括将重组酶多肽引入细胞或将编码位点特异性重组酶的多核苷酸引入宿主细胞。编码位点特异性重组酶的多核苷酸可位于插入多核苷酸内或单独的多核苷酸内。位点特异性重组酶可有效连接至细胞中有活性的启动子,包括例如诱导型启动子、对于细胞为内源的启动子、对于细胞为异源的启动子、细胞特异性启动子、组织特异性启动子或发育阶段特异性启动子。可在插入多核苷酸旁侧或在插入多核苷酸中的任何目标多核苷酸旁侧的位点特异性重组靶序列可包括但不限于loxP、lox511、lox2272、lox66、lox71、loxM2、lox5171、FRT、FRT11、FRT71、attp、att、FRT、rox以及它们的组合。
在其他实施例中,位点特异性重组位点在插入多核苷酸内所含的编码选择标记和/或报告基因的多核苷酸旁侧。在此类情况下,在所靶向的基因组座位处整合插入多核苷酸之后,可除去在位点特异性重组位点之间的序列。
在一个实施例中,插入多核苷酸包含编码选择标记的多核苷酸。此类选择标记包括但不限于新霉素磷酸转移酶(neo
插入多核苷酸还可包含有效连接至启动子的报告基因,其中所述报告基因编码报告蛋白,所述报告蛋白选自LacZ、mPlum、mCherry、tdTomato、mStrawberry、J-Red、DsRed、mOrange、mKO、mCitrine、Venus、YPet、增强型黄色荧光蛋白(EYFP)、Emerald、增强型绿色荧光蛋白(EGFP)、CyPet、青色荧光蛋白(CFP)、Cerulean、T-Sapphire、荧光素酶、碱性磷酸酶以及它们的组合。此类报告基因可有效连接至细胞中有活性的启动子。此类启动子可为诱导型启动子、对于报告基因或细胞为内源的启动子、对于报告基因或细胞为异源的启动子、细胞特异性启动子、组织特异性启动子或发育阶段特异性启动子。
靶向载体用于将插入多核苷酸引入Y染色体上的所靶向基因组座位中,或引入挑战性靶基因座中,或引入另一个目标染色体上。靶向载体包含插入多核苷酸,并且还包含上游同源臂和下游同源臂,这些同源臂在插入多核苷酸的旁侧。在插入多核苷酸旁侧的同源臂对应于所靶向基因组座位内的基因组区域。为了便于提及,所靶向基因组座位内的对应基因组区域在本文中称为“靶位点”。因此,在一个例子中,靶向载体可包含第一插入多核苷酸,所述第一插入多核苷酸侧接第一同源臂和第二同源臂,所述第一同源臂和第二同源臂与位置足够接近编码选择标记的多核苷酸内的第一识别位点的第一靶位点和第二靶位点相对应。因此,靶向载体由此有助于经由在细胞的基因组内的同源臂与对应靶位点之间发生的同源重组事件将插入多核苷酸整合到所靶向的基因组座位中。
靶向载体的同源臂可具有足以促进与对应靶位点发生同源重组事件的任何长度,包括例如约400bp至约500bp、约500bp至约600bp、约600bp至约700bp、约700bp至约800bp、约800bp至约900bp、或约900bp至约1000bp;或至少5-10、5-15、5-20、5-25、5-30、5-35、5-40、5-45、5-50、5-55、5-60、5-65、5-70、5-75、5-80、5-85、5-90、5-95、5-100、100-200、或200-300千碱基长或更长。在具体实施例中,靶向臂的总和为至少0.5kb、1kb、1.5kb、2kb、3kb、4kb、5kb、6kb、7kb、8kb、9kb或至少10kb。在其他实施例中,同源臂的总和为约0.5kb至约1kb、约1kb至约1.5kb、约1.5kb至约2kb、约2kb至约3kb、约3kb至约4kb、约4kb至约5kb、约5kb至约6kb、约6kb至约7kb、约7kb至约8kb、约8kb至约9kb、或约10kb至约150kb。如下文进一步详细概述,大靶向载体可采用更大长度的靶向臂。
与靶向载体的上游同源臂和下游同源臂对应的、所靶向基因组座位内的靶位点的位置“足够接近识别位点”,该识别位点位于编码选择标记的多核苷酸中。如本文所用,靶向载体的上游同源臂和下游同源臂“位置足够接近”识别位点,这一说法在所述距离能促进在识别位点处出现切口或双链断裂后在靶位点与同源臂之间发生同源重组事件时成立。因此,在具体实施例中,与靶向载体的上游同源臂和/或下游同源臂相对应的靶位点位于给定识别位点至少10个核苷酸至约14kb内。在具体实施例中,识别位点紧邻靶位点中的至少一者或两者。
对应于靶向载体的同源臂的靶位点与编码选择标记的多核苷酸内的识别位点的空间关系可能有变。例如,这两个靶位点可位于识别位点的5’,这两个靶位点可位于识别位点的3’,或靶位点可在识别位点的旁侧。
在具体实施例中,靶基因组座位包含(i)与5’同源臂同源的5’靶序列,以及(ii)与3’同源臂同源的3’靶序列。在具体实施例中,5’靶序列和3’靶序列相隔至少5kb但少于3Mb、至少5kb但少于10kb、至少10kb但少于20kb、至少20kb但少于40kb、至少40kb但少于60kb、至少60kb但少于80kb、至少约80kb但少于100kb、至少100kb但少于150kb、或至少150kb但少于200kb、至少约200kb但少于约300kb、至少约300kb但少于约400kb、至少约400kb但少于约500kb、至少约500kb但少于约1Mb、至少约1Mb但少于约1.5Mb、至少约1.5Mb但少于约2Mb、至少约2Mb但少于约2.5Mb、或至少约2.5Mb但少于约3Mb。
如本文所用,在两个区域彼此共有水平足够高的序列同一性时,同源臂和靶位点彼此“对应”,从而充当同源重组反应的底物。所谓“同源性”,意指DNA序列与对应序列相同或共有序列同一性。给定靶位点与存在于靶向载体上的对应同源臂之间的序列同一性,可为允许同源重组发生的任何程度的序列同一性。例如,靶向载体的同源臂(或其片段)与靶位点(或其片段)共有的序列同一性的大小可为至少50%、55%、60%、65%、70%、75%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%,使得所述序列经历同源重组。此外,同源臂与对应靶位点之间的对应同源区可具有足以促进在裂解的识别位点处发生同源重组的任何长度。例如,给定同源臂和/或对应靶位点可包含对应同源区,所述对应同源区为约400bp至约500bp、约500bp至约600bp、约600bp至约700bp、约700bp至约800bp、约800bp至约900bp、或约900bp至约1000bp(诸如针对本文别处所述的smallTVEC载体描述的);或至少约5-10、5-15、5-20、5-25、5-30、5-35、5-40、5-45、5-50、5-55、5-60、5-65、5-70、5-75、5-80、5-85、5-90、5-95、5-100、100-200、或200-300千碱基长或更长(诸如本文别处所述的LTVEC载体中描述的),使得同源臂与在细胞的基因组内的对应靶位点具有足以经历同源重组的同源性。
为了便于提及,同源臂在本文中指上游同源臂和下游同源臂。该术语涉及同源臂与插入多核苷酸在靶向载体内的相对位置。
靶向载体的同源臂因此被设计成与具有Y染色体上的所靶向基因组座位或位于挑战性靶基因座内的靶位点相对应。因此,同源臂可与对于细胞为天然的基因组座位相对应,或另选地,同源臂可与整合到Y染色体中的异源或外源DNA区段的区域相对应,所述区域包括但不限于转基因、表达盒、或者异源或外源基因组DNA区域。另选地,靶向载体的同源臂可与酵母人工染色体(YAC)、细菌人工染色体(BAC)、人类人工染色体的区域、或在适当宿主细胞中包含的任何其他经改造的基因组区域相对应。更进一步,靶向载体的同源臂可与BAC文库、粘粒文库或P1噬菌体文库的区域相对应,或可衍生自BAC文库、粘粒文库或P1噬菌体文库的区域。因此,在具体实施例中,靶向载体的同源臂与对于以下生物为天然、异源或外源的Y染色体上的基因组座位或挑战性靶基因座相对应:非人哺乳动物、啮齿动物、人、大鼠、小鼠、仓鼠、兔、猪、牛、鹿、绵羊、山羊、鸡、猫、狗、白鼬、灵长类动物(例如,狨猴、恒河猴)、家养哺乳动物或农业哺乳动物、鸟类或任何其他目标生物体。在另外的实施例中,同源臂与细胞中这样的基因组座位相对应:在不存在核酸酶试剂所诱导的切口或双链断裂的情况下,该基因组座位无法使用常规方法靶向,或者只可不正确地靶向或只以明显较低的效率靶向。在一个实施例中,同源臂衍生自合成DNA。
在其他实施例中,上游同源臂和下游同源臂对应于与所靶向基因组相同的基因组。在一个实施例中,同源臂来自相关基因组,例如,所靶向基因组为第一品系的小鼠基因组,而靶向臂来自第二品系的小鼠基因组,其中所述第一品系与第二品系不同。在其他实施例中,同源臂来自相同动物的基因组或来自相同品系的基因组,例如,所靶向基因组为第一品系的小鼠基因组,而靶向臂来自相同小鼠的小鼠基因组或来自相同品系的小鼠基因组。
靶向载体(诸如大靶向载体)还可包含如本文别处所讨论的选择盒或报告基因。选择盒可包含编码选择标记的核酸序列,其中所述核酸序列有效连接至启动子。此类启动子可为诱导型启动子、对于报告基因或细胞为内源的启动子、对于报告基因或细胞为异源的启动子、细胞特异性启动子、组织特异性启动子或发育阶段特异性启动子。在一个实施例中,选择标记选自新霉素磷酸转移酶(neo
在一个实施例中,靶向载体(诸如大靶向载体)包含有效连接至启动子的报告基因,其中所述报告基因编码报告蛋白,所述报告蛋白选自LacZ、mPlum、mCherry、tdTomato、mStrawberry、J-Red、DsRed、mOrange、mKO、mCitrine、Venus、YPet、增强型黄色荧光蛋白(EYFP)、Emerald、增强型绿色荧光蛋白(EGFP)、CyPet、青色荧光蛋白(CFP)、Cerulean、T-Sapphire、荧光素酶、碱性磷酸酶以及它们的组合。此类报告基因可有效连接至细胞中有活性的启动子。此类启动子可为诱导型启动子、对于报告基因或细胞为内源的启动子、对于报告基因或细胞为异源的启动子、细胞特异性启动子、组织特异性启动子或发育阶段特异性启动子。
在一个非限制性实施例中,与单独使用靶向载体相比,联合使用靶向载体(包括例如大靶向载体)与核酸酶试剂导致靶向效率提高。在一个实施例中,与单独使用靶向载体时相比,将靶向载体与核酸酶试剂结合使用时,靶向载体的靶向效率提高至少两倍、至少三倍、至少四倍或至少十倍。
如本文所用,术语“大靶向载体”或“LTVEC”包括这样的大靶向载体:其包含对应于且衍生自比通常由意欲在细胞中执行同源靶向的其他方法使用的那些核酸序列大的核酸序列的同源臂,并且/或者包含具有比通常由意欲在细胞中执行同源重组靶向的其他方法使用的那些核酸序列大的核酸序列的插入多核苷酸。在具体实施例中,LTVEC的同源臂和/或插入多核苷酸包含真核细胞的基因组序列。LTVEC的尺寸过大,无法通过例如Southern印迹和长片段(例如,1kb至5kb)PCR的常规测定法来筛选靶向事件。LTVEC的例子包括但不限于衍生自细菌人工染色体(BAC)、人类人工染色体或酵母人工染色体(YAC)的载体。LTVEC及其制备方法的非限制性例子描述于例如美国专利No.6,586,251、No.6,596,541、No.7,105,348和WO 2002/036789(PCT/US01/45375)中,这些专利各自以引用方式并入本文。
LTVEC可具有任何长度,包括但不限于至少约10kb、约15kb、约20kb、约30kb、约40kb、约50kb、约60kb、约70kb、约80kb、约90kb、约100kb、约150kb、约200kb、约10kb至约15kb、约15kb至约20kb、约20kb至约30kb、约30kb至约50kb、约50kb至约300kb、约50kb至约75kb、约75kb至约100kb、约100kb至125kb、约125kb至约150kb、约150kb至约175kb、约175kb至约200kb、约200kb至约225kb、约225kb至约250kb、约250kb至约275kb、或约275kb至约300kb。
在一个实施例中,LTVEC包含在约5kb至约200kb、约5kb至约10kb、约10kb至约20kb、约20kb至约30kb、约30kb至约40kb、约40kb至约50kb、约60kb至约70kb、约80kb至约90kb、约90kb至约100kb、约100kb至约110kb、约120kb至约130kb、约130kb至约140kb、约140kb至约150kb、约150kb至约160kb、约160kb至约170kb、约170kb至约180kb、约180kb至约190kb、或约190kb至约200kb、约200kb至约250kb、约250kb至约300kb、约300kb至约350kb、或约350kb至约400kb范围内的插入多核苷酸。
在其他情况下,LTVEC设计可使得允许替换如本文所述的约5kb至约200kb、或约5kb至约3.0Mb的给定序列。在一个实施例中,所述替换为约5kb至约10kb、约10kb至约20kb、约20kb至约30kb、约30kb至约40kb、约40kb至约50kb、约50kb至约60kb、约60kb至约70kb、约80kb至约90kb、约90kb至约100kb、约100kb至约110kb、约110kb至约120kb、约120kb至约130kb、约130kb至约140kb、约140kb至约150kb、约150kb至约160kb、约160kb至约170kb、约170kb至约180kb、约180kb至约190kb、约190kb至约200kb、约5kb至约10kb、约10kb至约20kb、约20kb至约40kb、约40kb至约60kb、约60kb至约80kb、约80kb至约100kb、约100kb至约150kb、或约150kb至约200kb、约200kb至约300kb、约300kb至约400kb、约400kb至约500kb、约500kb至约1Mb、约1Mb至约1.5Mb、约1.5Mb至约2Mb、约2Mb至约2.5Mb、或约2.5Mb至约3Mb。
在一个实施例中,LTVEC的同源臂衍生自BAC文库、粘粒文库或P1噬菌体文库。在其他实施例中,同源臂衍生自细胞的所靶向基因组座位;在一些情况下,LTVEC被设计用来靶向其的靶基因组座位无法使用常规方法靶向。在其他实施例中,同源臂衍生自合成DNA。
在一个实施例中,LTVEC中的上游同源臂和下游同源臂的总和为至少10kb。在其他实施例中,上游同源臂在约5kb至约100kb的范围内。在一个实施例中,下游同源臂在约5kb至约100kb的范围内。在其他实施例中,上游同源臂和下游同源臂的总和为约5kb至约10kb、约10kb至约20kb、约20kb至约30kb、约30kb至约40kb、约40kb至约50kb、约50kb至约60kb、约60kb至约70kb、约70kb至约80kb、约80kb至约90kb、约90kb至约100kb、约100kb至约110kb、约110kb至约120kb、约120kb至约130kb、约130kb至约140kb、约140kb至约150kb、约150kb至约160kb、约160kb至约170kb、约170kb至约180kb、约180kb至约190kb、或约190kb至约200kb。在一个实施例中,所述缺失的尺寸与LTVEC的5'同源臂和3'同源臂的总和的尺寸相同或相似。
在一个实施例中,LTVEC包含如本文别处所讨论的选择盒或报告基因。
提供了用于修饰细胞中Y染色体上的靶基因组座位的方法,包括:(a)提供在Y染色体上包含靶基因组座位的细胞,(b)向该细胞中引入包含第一插入多核苷酸的第一靶向载体,所述第一插入多核苷酸侧接与第一靶位点和第二靶位点对应的第一同源臂和第二同源臂;以及(c)鉴定在其基因组中包含在Y染色体上的靶基因组座位处整合的第一插入多核苷酸的至少一个细胞。可执行类似方法以靶向挑战性染色体基因座。如本文别处详细讨论,在具体实施例中,靶向载体的第一同源臂和第二同源臂的总和为约0.5kb、1kb、1.5kb、2kb、3kb、4kb、5kb、6kb、7kb、8kb、9kb、约0.5kb至约1kb、约1kb至约1.5kb、约1.5kb至约2kb、约2kb至约3kb、约3kb至约4kb、约4kb至约5kb、约5kb至约6kb、约6kb至约7kb、约8kb至约9kb,或为至少10kb或至少10kb且少于150kb。在具体实施例中,采用LTVEC。在其他具体实施例中,采用smallTVEC。在一个非限制性实施例中,执行此类方法,其采用促进本文所公开的XYF0可育雌性发育的培养基,从而生成XY F0可育雌性动物。在其他情况下,本文所述的方法用于在Sry基因中产生靶向遗传修饰,如本文别处所讨论。
还提供了用于修饰细胞中Y染色体上的靶基因组座位的方法,包括:(a)提供在Y染色体上包含靶基因组座位的细胞,所述靶基因组座位包含核酸酶试剂的识别位点,(b)向该细胞中引入(i)核酸酶试剂,其中所述核酸酶试剂在第一识别位点处诱导切口或双链断裂;和(ii)包含第一插入多核苷酸的第一靶向载体,所述第一插入多核苷酸侧接第一同源臂和第二同源臂,所述第一同源臂和第二同源臂与位置足够接近第一识别位点的第一靶位点和第二靶位点相对应;以及(c)鉴定在其基因组中包含在Y染色体上的靶基因组座位处整合的第一插入多核苷酸的至少一个细胞。可执行类似方法以靶向挑战性靶基因座。如本文别处详细讨论,在具体实施例中,靶向载体的第一同源臂和第二同源臂的总和为约0.5kb、1kb、1.5kb、2kb、3kb、4kb、5kb、6kb、7kb、8kb、9kb、约0.5kb至约1kb、约1kb至约1.5kb、约1.5kb至约2kb、约2kb至约3kb、约3kb至约4kb、约4kb至约5kb、约5kb至约6kb、约6kb至约7kb、约8kb至约9kb,或为至少10kb或至少10kb且少于150kb。在具体实施例中,采用LTVEC。在其他具体实施例中,采用smallTVEC。在一个非限制性实施例中,执行此类方法,其采用促进本文所公开的XY F0可育雌性发育的培养基,从而生成XY F0可育雌性动物。在其他情况下,本文所述的方法用于在Sry基因中产生靶向遗传修饰,如本文别处所讨论。
还可采用各种方法来鉴定具有在靶基因组座位处整合的插入多核苷酸的细胞。插入多核苷酸在靶基因组座位处的插入产生“等位基因修饰”。术语“等位基因修饰”或“MOA”包括对基因组中一个或多个基因或染色体基因座的一个等位基因的精确DNA序列的修饰。“等位基因修饰(MOA)”的例子包括但不限于缺失、置换或插入少至单个核苷酸,或跨一个或多个目标基因或染色体基因座缺失数千碱基,以及这两个末端之间的任何以及所有可能的修饰。
在各种实施例中,为了促进对靶向修饰的鉴定,采用高通量定量测定法,也就是等位基因修饰(MOA)测定法。本文所述的MOA测定法允许在遗传修饰之后大规模筛选在亲本染色体中的一个或多个经修饰等位基因。MOA测定法可经由各种分析技术进行,包括但不限于定量PCR,例如实时PCR(qPCR)。例如,实时PCR包括识别靶基因座的第一引物探针组和识别非靶向参考基因座的第二引物探针组。此外,引物探针组包含识别扩增序列的荧光探针。定量测定法还可经由多种分析技术进行,包括但不限于荧光介导原位杂交(FISH)、比较基因组杂交、等温DNA扩增、与固定探针的定量杂交、Invader
在各种实施例中,在存在切口或双链断裂的情况下,靶向载体(诸如LTVEC或smallTVEC)在靶基因组座位处的靶向效率是不存在切口或双链断裂(使用例如相同的靶向载体和相同的同源臂以及在目标基因组座位处的对应靶位点,但不存在会造成切口或双链断裂的所添加核酸酶试剂)时的至少约2倍高、至少约3倍高、至少约4倍高。
可依次地重复上述各种方法,以允许任何数量的插入多核苷酸在Y染色体上的给定所靶向基因组座位中或在挑战性靶基因座中靶向整合。因此,所述各种方法可以实现在Y染色体上的靶基因组座位中或在挑战性靶基因座中插入至少1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20个或更多个插入多核苷酸。在特定实施例中,此类顺序拼接方法允许将来自动物细胞或来自哺乳动物细胞(即,人、非人、啮齿动物、小鼠、猴、大鼠、仓鼠、家养哺乳动物或农业动物)的大基因组区域重建到Y染色体上的所靶向基因组座位中。在此类情况下,包含编码区和非编码区两者的基因组区域的转移和重建允许通过至少部分地保留在天然基因组区域内发现的编码区、非编码区和拷贝数变异来保持给定区域的复杂性。因此,所述各种方法提供了例如在任何哺乳动物细胞或目标动物内生成“异源”或“外源”基因组区域的方法。在一个非限制性例子中,在非人动物内生成了“人源化”基因组区域。
还已经认识到,除了修饰Y染色体上的靶基因组座位以外,本文所公开的各种方法和组合物还可用于在另一个染色体上生成所靶向的遗传修饰。
任何目标多核苷酸都可包含在各种插入多核苷酸中,由此在Y染色体上的靶基因组座位处或在挑战性靶基因座中整合。本文所公开的方法提供整合到所靶向基因组座位中的至少1、2、3、4、5、6个或更多个目标多核苷酸。
插入多核苷酸内的目标多核苷酸在Y染色体上的靶基因组座位处或在挑战性靶基因座处整合时,可将一个或多个遗传修饰引入细胞中。所述遗传修饰可包括缺失内源核酸序列以及/或者将外源或异源或直系同源多核苷酸添加到靶基因组座位中。在一个实施例中,所述遗传修饰包括在靶基因组座位处用外源目标多核苷酸替换内源核酸序列。因此,本文所提供的方法允许在Y染色体上的靶基因组座位中生成遗传修饰,所述遗传修饰包括敲除、缺失、插入、替换(“敲入”)、点突变、结构域交换、外显子交换、内含子交换、调控序列交换、基因交换或它们的组合。此类修饰可在将第一、第二、第三、第四、第五、第六、第七或任何后续的插入多核苷酸整合到靶基因组座位中之后发生。
在插入多核苷酸内的和/或在靶基因组座位处整合的目标多核苷酸可包括对于其引入的细胞为天然或同源的序列、对于其引入的细胞可为异源的目标多核苷酸、对于其引入的细胞可为外源的目标多核苷酸、对于其引入的细胞可为直系同源的目标多核苷酸,或者所述目标多核苷酸可来自与其引入的细胞不同的物种。如本文所用,与序列有关的“同源”,是指序列对于细胞来说是天然的。如本文所用,与序列有关的“异源”,是指序列来源于外来物种,而如果来源于相同物种,则通过有意人为干预,从其天然形式进行组成和/或基因组座位方面的实质修饰。如本文所用,与序列有关的“外源”,是指序列来源于外来物种。如本文所用,“直系同源”是指来自一种物种的多核苷酸在功能上等同于另一物种(即,物种变体)中的已知参考序列。目标多核苷酸可来自任何目标生物体,包括但不限于非人、啮齿动物、仓鼠、小鼠、大鼠、人、猴、鸟类、农业哺乳动物或非农业哺乳动物。目标多核苷酸还可包含编码区、非编码区、调控区或基因组DNA。因此,第1、第2、第3、第4、第5、第6、第7插入多核苷酸和/或任何后续插入多核苷酸可包含此类序列。
在一个实施例中,在插入多核苷酸内和/或在Y染色体上的靶基因组座位处整合的目标多核苷酸与小鼠核酸序列、人核酸、非人核酸、啮齿动物核酸、大鼠核酸、仓鼠核酸、猴核酸、农业哺乳动物核酸或非农业哺乳动物核酸是同源的。在更进一步的实施例中,在靶基因座处整合的目标多核苷酸为基因组核酸的片段。在一个实施例中,基因组核酸为小鼠基因组核酸、人基因组核酸、非人核酸、啮齿动物核酸、大鼠核酸、仓鼠核酸、猴核酸、农业哺乳动物核酸或非农业哺乳动物核酸,或它们的组合。
在一个实施例中,如上所述,目标多核苷酸可在约500个核苷酸至约200kb的范围内。目标多核苷酸可为约500个核苷酸至约5kb、约5kb至约200kb、约5kb至约10kb、约10kb至约20kb、约20kb至约30kb、约30kb至约40kb、约40kb至约50kb、约60kb至约70kb、约80kb至约90kb、约90kb至约100kb、约100kb至约110kb、约120kb至约130kb、约130kb至约140kb、约140kb至约150kb、约150kb至约160kb、约160kb至约170kb、约170kb至约180kb、约180kb至约190kb、或约190kb至约200kb。
在插入多核苷酸内的和/或在Y染色体上的靶基因组座位处或挑战性靶基因座之中插入的目标多核苷酸可编码多肽,可编码miRNA,可编码长非编码RNA,或者可包含任何目标调控区或非编码区,包括例如调控序列、启动子序列、增强子序列、转录阻遏物结合序列,或非蛋白编码序列的缺失,但不包含蛋白编码序列的缺失。另外,在插入多核苷酸内的和/或在Y染色体上的靶基因组座位处或在挑战性靶基因座处插入的目标多核苷酸可编码在神经系统、骨骼系统、消化系统、循环系统、肌肉系统、呼吸系统、心血管系统、淋巴系统、内分泌系统、泌尿系统、生殖系统或它们的组合中表达的蛋白。
在插入多核苷酸内的和/或在Y染色体上的靶基因组座位处或在挑战性靶基因座处整合的目标多核苷酸可在编码序列中包含遗传修饰。此类遗传修饰包括但不限于编码序列的缺失突变或两个编码序列的融合。
在插入多核苷酸内的和/或在Y染色体上的靶基因组座位处或在挑战性靶基因座处整合的目标多核苷酸可包括编码突变体蛋白的多核苷酸。在一个实施例中,所述突变体蛋白的特征在于结合特性改变、定位改变、表达改变和/或表达模式改变。在一个实施例中,在插入多核苷酸内的和/或在Y染色体上的靶基因组座位处或在挑战性靶基因座处整合的目标多核苷酸包含至少一个疾病等位基因。在此类情况下,所述疾病等位基因可为显性等位基因或隐性等位基因。此外,所述疾病等位基因可包括单核苷酸多态性(SNP)等位基因。编码突变体蛋白的目标多核苷酸可来自任何生物体,包括但不限于编码突变体蛋白的哺乳动物、非人哺乳动物、啮齿动物、小鼠、大鼠、人、猴、农业哺乳动物或家养哺乳动物多核苷酸。
在插入多核苷酸内的和/或在Y染色体上的靶基因组座位处或在挑战性靶基因座处整合的目标多核苷酸还可包含调控序列,包括例如启动子序列、增强子序列、转录阻遏物结合序列或转录终止子序列。在具体实施例中,在插入多核苷酸内的和/或在Y染色体上的靶基因组座位处或在挑战性靶基因座处整合的目标多核苷酸包含具有非蛋白编码序列的缺失的多核苷酸,但不包含蛋白编码序列的缺失。在一个实施例中,所述非蛋白编码序列的缺失包括调控序列的缺失。在另一个实施例中,所述调控元件的缺失包括启动子序列的缺失。在一个实施例中,所述调控元件的缺失包括增强子序列的缺失。这种目标多核苷酸可来自任何生物体,包括但不限于编码突变体蛋白的哺乳动物、非人哺乳动物、啮齿动物、小鼠、大鼠、人、猴、农业哺乳动物或家养哺乳动物多核苷酸。
本文所公开的各种方法可用于在挑战性基因组座位中或在Y染色体基因座(诸如Sry)中生成多种修饰。此类修饰包括例如用同源或直系同源核酸序列替换内源核酸序列;内源核酸序列的缺失;内源核酸序列的缺失,其中所述缺失在约5kb至约10kb、约10kb至约20kb、约20kb至约40kb、约40kb至约60kb、约60kb至约80kb、约80kb至约100kb、约100kb至约150kb、约150kb至约200kb、约200kb至约300kb、约300kb至约400kb、约400kb至约500kb、约500kb至约600kb、约600kb至约700kb、约700kb至约800kb、约800kb至约900kb、约900kb至约1Mb、约500kb至约1Mb、约1Mb至约1.5Mb、约1.5Mb至约2Mb、约2Mb至约2.5Mb、或约2.5Mb至约3Mb的范围内;外源核酸序列的插入;外源核酸序列的插入,所述插入在约5kb至约10kb、约10kb至约20kb、约20kb至约40kb、约40kb至约60kb、约60kb至约80kb、约80kb至约100kb、约100kb至约150kb、约150kb至约200kb、约200kb至约250kb、约250kb至约300kb、约300kb至约350kb、或约350kb至约400kb的范围内;包含同源或直系同源核酸序列的外源核酸序列的插入;包含人和非人核酸序列的嵌合核酸序列的插入;侧接有位点特异性重组酶靶序列的条件性等位基因的插入;有效连接至哺乳动物细胞中有活性的第三启动子的可选标记或报告基因的插入;或它们的组合。
如上文概述,本文提供了实现位于Y染色体上的、位于挑战性靶基因座处的一个或多个目标多核苷酸的靶向遗传修饰,或者Sry蛋白水平和/或活性降低的方法和组合物。还已经认识到,除了对Y染色体上或挑战性靶染色体基因座上的序列进行靶向遗传修饰以外,还可对其他染色体进行附加的靶向遗传修饰。可实现这些靶向遗传修饰的此类系统可采用多种组分,为了便于提及,本文中的术语“靶向基因组整合系统”通常包括整合事件所需的所有组分(即,各种核酸酶试剂、识别位点、插入DNA多核苷酸、靶向载体、靶基因组座位和目标多核苷酸)。
本文所提供的方法包括向细胞中引入包含靶向基因组整合系统的各种组分的一个或多个多核苷酸或多肽构建体。“引入”意指以使得序列(多肽或多核苷酸)能够进入细胞内部的方式将序列呈递到细胞。本文所提供的方法并不取决于将靶向基因组整合系统的任何组分引入细胞中的特定方法,只要使多核苷酸能够进入至少一个细胞的内部即可。用于将多核苷酸引入各种细胞类型中的方法是本领域已知的,包括但不限于稳定转染方法、瞬时转染方法和病毒介导的方法。
在一些实施例中,在所述方法和组合物中采用的细胞具有稳定地掺入其基因组中的DNA构建体。“稳定地掺入”或“稳定地引入”意指将多核苷酸引入细胞中,使得核苷酸序列整合到细胞的基因组中且能够被其子代遗传。可使用任何方案稳定地掺入DNA构建体或靶向基因组整合系统的各种组分。
转染方案以及用于将多肽或多核苷酸序列引入细胞中的方案可能有变。非限制性转染方法包括基于化学的转染方法,其包括使用脂质体、纳米粒子、磷酸钙(Graham et al.(1973).Virology 52(2):456–67(Graham等人,1973年,《病毒学》,第52卷,第2期,第456–467页),Bacchetti et al.(1977)Proc Natl Acad Sci USA 74(4):1590–4(Bacchetti等人,1977年,《美国国家科学院院刊》,第74卷,第4期,第1590–1594页)以及Kriegler,M(1991).Transfer and Expression:A Laboratory Manual.New York:W.H.Freeman andCompany.pp.96–97(Kriegler,M,1991年,《转移和表达:实验室手册》,纽约W.H.弗里曼公司,第96–97页))、树状体或阳离子聚合物(诸如DEAE-葡聚糖或聚乙烯亚胺)。非化学方法包括电穿孔、声孔效应和光学转染。基于颗粒的转染包括使用基因枪、磁铁辅助转染(Bertram,J.(2006)Current Pharmaceutical Biotechnology 7,277–28(Bertram,J.,2006年,《当今药物生物技术》,第7卷,第277–28页))。病毒方法也可用于转染。
在一个实施例中,将核酸酶试剂与靶向载体、smallTVEC或大靶向载体(LTVEC)同时引入细胞中。另选地,在一段时间内将核酸酶试剂与靶向载体、smallTVEC或LTVEC分别引入。在一个实施例中,在引入靶向载体、smallTVEC或LTVEC之前引入核酸酶试剂,而在其他实施例中,在引入靶向载体、smallTVEC或LTVEC之后引入核酸酶试剂。
可采用本文所公开的各种方法生成非人动物。此类方法包括:(1)采用本文所公开的方法在非人动物多能细胞的Y染色体的靶基因组座位处整合一个或多个目标多核苷酸,以生成在Y染色体的所靶向基因组座位中包含插入多核苷酸的经遗传修饰的多能细胞;(2)选择在Y染色体的靶基因组座位处具有一个或多个目标多核苷酸的经遗传修饰的多能细胞;(3)将经遗传修饰的多能细胞引入处于桑椹胚前期的非人动物宿主胚胎中;以及(4)将包含经遗传修饰的多能细胞的宿主胚胎植入代孕母体中,以生成衍生自经遗传修饰的多能细胞的F0代。可采用类似方法靶向挑战性靶染色体基因座。所述非人动物可为非人哺乳动物、啮齿动物、小鼠、大鼠、仓鼠、猴、农业哺乳动物或家养哺乳动物、或鱼类、或禽类。
所述多能细胞可为人ES细胞、非人ES细胞、啮齿动物ES细胞、小鼠ES细胞、大鼠ES细胞、仓鼠ES细胞、猴ES细胞、农业哺乳动物ES细胞或家养哺乳动物ES细胞。在其他实施例中,所述多能细胞为哺乳动物细胞、人细胞、非人哺乳动物细胞、人多能细胞、人ES细胞、人成体干细胞、发育受限的人祖细胞、人iPS细胞、人细胞、啮齿动物细胞、大鼠细胞、小鼠细胞、仓鼠细胞。在一个实施例中,所述靶向遗传修饰降低Sry蛋白的水平和/或活性。在此类情况下,所述多能细胞可包括XY ES细胞或XY iPS细胞。培养此类细胞以促进F0可育XY雌性动物发育的方法在本文别处详细描述。
也可使用核移植技术生成非人哺乳动物。简而言之,用于核移植的方法包括以下步骤:(1)将卵母细胞去核;(2)分离供体细胞或核,准备与去核卵母细胞合并;(3)将所述细胞或核插入所述去核卵母细胞中,形成重建细胞;(4)将所述重建细胞植入动物的子宫,以形成胚胎;以及(5)允许所述胚胎发育。在此类方法中,一般从处死的动物体内取出卵母细胞,不过也可从活体动物的输卵管和/或卵巢中分离卵母细胞。卵母细胞可在去核之前在本领域普通技术人员已知的多种培养基中成熟。将卵母细胞去核可采用本领域普通技术人员熟知的多种方式执行。将供体细胞或核插入去核卵母细胞中以形成重建细胞,通常借助在融合之前在透明带下显微注射供体细胞而实现。融合可通过跨接触/融合平面施加直流电脉冲(电融合)、通过将细胞暴露于促进融合的化学品(诸如聚乙二醇)、或借助灭活病毒(诸如仙台病毒)来诱导。重建细胞通常在核供体和受体卵母细胞融合之前、期间和/或之后,通过电力方式和/或非电力方式激活。激活方法包括电脉冲、化学诱导冲击、精液渗透、增加二价阳离子在卵母细胞中的水平以及降低细胞蛋白在卵母细胞中的磷酸化(如借助激酶抑制剂)。激活的重建细胞或胚胎通常在本领域普通技术人员熟知的培养基中培养,然后移植到动物的子宫中。参见例如US20080092249、WO/1999/005266A2、US20040177390、WO/2008/017234A1以及美国专利No.7,612,250,这些专利各自以引用方式并入本文。
提供了用于制备在其种系中包含一个或多个如本文所述的遗传修饰的非人动物的其他方法,包括:(a)采用本文所述的各种方法在原核细胞中修饰非人动物的Y染色体上的所靶向基因组座位;(b)选择在所靶向基因组座位处包含遗传修饰的经修饰的原核细胞;(c)从经修饰的原核细胞的基因组分离经遗传修饰的靶向载体;(d)将经遗传修饰的靶向载体引入非人动物的多能细胞中,以生成在Y染色体上的所靶向基因组座位处包含插入核酸的经遗传修饰的多能细胞;(e)选择经遗传修饰的多能细胞;(f)将经遗传修饰的多能细胞引入处于桑椹胚前期的非人动物宿主胚胎中;以及(g)将包含经遗传修饰的多能细胞的宿主胚胎植入代孕母体中,以生成衍生自经遗传修饰的多能细胞的F0代。在此类方法中,所述靶向载体可包括大靶向载体或smallTVEC。可采用类似方法靶向挑战性靶基因座。所述非人动物可为非人哺乳动物、啮齿动物、小鼠、大鼠、仓鼠、猴、农业哺乳动物或家养哺乳动物。所述多能细胞可为人ES细胞、非人ES细胞、啮齿动物ES细胞、小鼠ES细胞、大鼠ES细胞、仓鼠ES细胞、猴ES细胞、农业哺乳动物ES细胞或家养哺乳动物ES细胞。在其他实施例中,所述多能细胞为哺乳动物细胞、人细胞、非人哺乳动物细胞、人多能细胞、人ES细胞、人成体干细胞、发育受限的人祖细胞、人iPS细胞、人细胞、啮齿动物细胞、大鼠细胞、小鼠细胞、仓鼠细胞。在一个实施例中,所述靶向遗传修饰降低Sry蛋白的水平和/或活性。在此类情况下,所述多能细胞可包括XY ES细胞或XY iPS细胞。培养此类细胞以促进F0可育XY雌性动物发育的方法在本文别处详细描述。
在另外的方法中,分离步骤(c)还包括(c1)使经遗传修饰的靶向载体(即,经遗传修饰的LTVEC)线性化。在更进一步的实施例中,引入步骤(d)还包括(d1)将如本文所述的核酸酶试剂引入多能细胞中。在一个实施例中,选择步骤(b)和/或(e)通过将如本文所述的选择性试剂施加到所述原核细胞或所述多能细胞来进行。在一个实施例中,选择步骤(b)和/或(e)经由如本文所述的等位基因修饰(MOA)测定法进行。
提供了经由在原核细胞中的细菌同源重组(BHR)来修饰动物细胞的靶基因组座位的另外的方法,包括:(a)提供包含Y染色体的靶基因组座位的原核细胞;(b)向所述原核细胞中引入包含侧接有第一上游同源臂和第一下游同源臂的插入多核苷酸的靶向载体(如上所述),其中所述插入多核苷酸包含哺乳动物基因组区域,然后向所述原核细胞中引入在第一识别位点处或附近产生切口或双链断裂的核酸酶试剂,以及(c)选择在所述染色体的靶基因组座位处包含插入多核苷酸的所靶向原核细胞,其中所述原核细胞能够表达介导BHR的重组酶。可采用类似方法靶向挑战性靶基因座。步骤(a)至(c)可如本文公开的那样连续地重复,以允许在原核细胞中的所靶向基因组座位处引入多个插入多核苷酸。一旦所靶向基因组座位用原核细胞“构建”,则包含Y染色体的经修饰靶基因组座位的靶向载体就可从原核细胞分离并引入哺乳动物细胞内的Y染色体的靶基因组座位中。随后可将包含Y染色体的经修饰基因组座位的哺乳动物细胞制成非人转基因动物。
提供了经由在原核细胞中的细菌同源重组(BHR)来修饰动物细胞的靶基因组座位的另外的方法,包括:(a)提供包含Y染色体的靶基因组座位的原核细胞;(b)向所述原核细胞中引入包含侧接有第一上游同源臂和第一下游同源臂的插入多核苷酸的靶向载体(如上所述),其中所述插入多核苷酸包含哺乳动物基因组区域,以及(c)选择在所述染色体的靶基因组座位处包含插入多核苷酸的所靶向原核细胞,其中所述原核细胞能够表达介导BHR的重组酶。可采用类似方法靶向挑战性靶基因座。步骤(a)至(c)可如本文公开的那样连续地重复,以允许在原核细胞中的所靶向基因组座位处引入多个插入多核苷酸。一旦所靶向基因组座位用原核细胞“构建”,包含Y染色体的经修饰靶基因组座位的靶向载体就可从原核细胞分离并引入哺乳动物细胞内的Y染色体的靶基因组座位中。随后可将包含Y染色体的经修饰基因组座位的哺乳动物细胞制成非人转基因动物。
在一些实施例中,本文所述的靶基因组座位的各种遗传修饰可使用衍生自细菌人工染色体(BAC)DNA的LTVEC,通过
在一些实施例中,将包含如本文所述的各种遗传修饰的所靶向XY多能细胞和/或全能细胞(即,XY ES细胞或XY iPS细胞)用作插入供体细胞,经由
本文所述的各种方法采用细胞中对于Y染色体或对于挑战性靶基因座的基因组座位靶向系统。此类细胞包括原核细胞,诸如细菌细胞,包括大肠杆菌(E.coli);或真核细胞,诸如酵母、昆虫、两栖动物、植物或哺乳动物细胞,包括但不限于小鼠细胞、大鼠细胞、兔细胞、猪细胞、牛细胞、鹿细胞、绵羊细胞、山羊细胞、鸡细胞、猫细胞、狗细胞、白鼬细胞、灵长类动物(例如,狨猴、恒河猴)细胞等,以及来自家养哺乳动物的细胞或来自农业哺乳动物的细胞。一些细胞为非人细胞,特别是非人哺乳动物细胞。在一些实施例中,对于不易获得合适的可遗传修饰的多能细胞的那些哺乳动物而言,采用其他方法将体细胞重新编程为多能细胞,例如经由向体细胞中引入多能诱导性因子的组合来重新编程,所述多能诱导性因子包括但不限于Oct3/4、Sox2、KLF4、Myc、Nanog、LIN28和Glis1。在此类方法中,所述细胞也可为哺乳动物细胞、人细胞、非人哺乳动物细胞、非人细胞,来自啮齿动物、大鼠、小鼠、仓鼠的细胞,成纤维细胞或任何其他宿主细胞。在其他实施例中,所述细胞为多能细胞、诱导性多能干(iPS)细胞、非人胚胎干(ES)细胞。此类细胞包括多能细胞,包括例如诱导性多能干(iPS)细胞、小鼠胚胎干(ES)细胞、大鼠胚胎干(ES)细胞、人胚胎(ES)细胞或发育受限的人祖细胞、啮齿动物胚胎干(ES)细胞、小鼠胚胎干(ES)细胞或大鼠胚胎干(ES)细胞。
术语“多核苷酸”、“多核苷酸序列”、“核酸序列”和“核酸片段”在本文中可互换使用。这些术语涵盖核苷酸序列等。多核苷酸可为任选地含有合成、非天然或改变的核苷酸碱基的单链或双链的RNA或DNA聚合物。DNA聚合物形式的多核苷酸可由cDNA、基因组DNA、合成DNA或它们的混合物的一个或多个区段构成。多核苷酸可包含脱氧核糖核苷酸和核糖核苷酸,包括天然存在的分子与合成类似物两者、以及这些的任意组合。本文提供的多核苷酸还涵盖所有形式的序列,包括但不限于单链形式、双链形式、发夹、茎环结构等。
还提供了重组多核苷酸。术语“重组多核苷酸”和“重组DNA构建体”在本文中可互换使用。重组构建体包含核酸序列的人工或异源组合,例如自然界中不共存的调控序列和编码序列。在其他实施例中,重组构建体可包含衍生自不同来源的调控序列和编码序列,或衍生自相同来源但以与自然界中所存在的方式不同的方式排列的调控序列和编码序列。这种构建体可独自使用,也可结合载体使用。如果使用载体,则载体的选择取决于如本领域技术人员所熟知的用以转化宿主细胞的方法。例如,可使用质粒载体。筛选尤其可通过DNA的Southern分析、mRNA表达的Northern分析、蛋白表达的免疫印迹分析或表型分析来实现。
在具体实施例中,可在表达盒中提供本文所述组分中的一者或多者,以便在多能和/或全能细胞中表达。所述盒可包含有效连接至本文所提供的多核苷酸的5'调控序列和3'调控序列。“有效连接”意指两个或更多个元件之间的功能性连接。例如,目标多核苷酸与调控序列(即,启动子)之间的有效连接为允许表达目标多核苷酸的功能性连接。有效连接的元件可为邻接的或不邻接的。使用“有效连接”来提及两个蛋白编码区的连接时,意指编码区位于同一个阅读框中。在另一种情况下,编码蛋白的核酸序列可有效连接至调控序列(例如,启动子、增强子、沉默子序列等)以保持恰当的转录调控。所述盒可另外包含将共同引入ES细胞中的至少一个额外的目标多核苷酸。另选地,所述额外的目标多核苷酸可在多个表达盒上提供。这种表达盒具有多个限制位点和/或重组位点,以使重组多核苷酸的插入处于调控区的转录调控之下。所述表达盒可另外包含选择标记基因。
所述表达盒在5'至3'转录方向上可包含在哺乳动物细胞或目标宿主细胞中起作用的转录和翻译起始区(即,启动子)、本文所提供的重组多核苷酸,以及转录和翻译终止区(即,终止区)。所述调控区(即,启动子、转录调控区以及转录和翻译终止区)和/或本文所提供的多核苷酸对于宿主细胞或对于彼此可为天然/类似的。另选地,所述调控区和/或本文所提供的多核苷酸对于宿主细胞或对于彼此可为异源的。例如,有效连接至异源多核苷酸的启动子来自与多核苷酸来源物种不同的物种,而如果来自相同/类似的物种,则一者或两者从其原始形式和/或基因组座位进行实质修饰,或者启动子不是有效连接的多核苷酸的天然启动子。另选地,本文所提供的调控区和/或重组多核苷酸可以是完全合成的。
所述终止区对于转录起始区而言可为天然的,对于有效连接的重组多核苷酸而言可为天然的,对于宿主细胞而言可为天然的,或者可衍生自对于启动子、重组多核苷酸、宿主细胞或它们的任何组合而言为另一种的(即,外来的或异源的)来源。
在制备表达盒时,可对各种DNA片段进行操纵,以便提供处于正确取向的DNA序列。为此目的,可采用衔接子或接头将DNA片段连接在一起,或者可涉及其他的操纵以提供便利的限制位点、去除多余的DNA、去除限制位点,等等。出于这个目的,可能涉及体外诱变、引物修复、限制性酶切、退火、再置换(例如转换和颠换)。
多种启动子可用于本文所提供的表达盒中。可根据期望的结果来选择启动子。已经认识到,不同的应用可通过在表达盒中使用不同的启动子来增强,从而调整目标多核苷酸表达的时机、位置和/或水平。如果需要,此类表达构建体还可包含启动子调控区(例如,赋予可诱导的、组成型的、环境或发育调控的、或细胞或组织特异性/选择性的表达)、转录起始位点、核糖体结合位点、RNA加工信号、转录终止位点和/或多聚腺苷酸化信号。
非限制性实施例包括:
1.一种体外培养物,包含:
(a)具有降低Sry蛋白水平和/或活性的修饰的非人哺乳动物XY胚胎干(ES)细胞;以及
(b)适用于在培养中维持非人哺乳动物ES细胞、包含基础培养基和补充物的培养基,其中所述培养基表现出以下一种或多种特征:约200mOsm/kg至小于约329mOsm/kg的渗透压;约11mS/cm至约13mS/cm的电导率;约50mM至约110mM的碱金属卤化物盐浓度;约17mM至约30mM的碳酸盐浓度;约85mM至约130mM的碱金属卤化物盐和碳酸盐总浓度;和/或这些特征之中任何两种或更多种的组合。
2.根据实施例1所述的体外培养物,其中所述非人哺乳动物XY ES细胞来自啮齿动物。
3.根据实施例2所述的体外培养物,其中所述啮齿动物为小鼠。
4.根据实施例3所述的体外培养物,其中所述小鼠XY ES细胞为VGF1小鼠ES细胞。
5.根据实施例2所述的体外培养物,其中所述啮齿动物为大鼠或仓鼠。
6.根据实施例1至5中任一项所述的体外培养物,其中所述Sry蛋白的水平和/或活性降低源自Sry基因中的遗传修饰。
7.根据实施例6所述的体外培养物,其中所述Sry基因中的所述遗传修饰包括一个或多个核苷酸的插入、一个或多个核苷酸的缺失、一个或多个核苷酸的置换、敲除、敲入、用异源核酸序列替换内源核酸序列,或它们的组合。
8.根据实施例1至7中任一项所述的体外培养物,其中所述非人哺乳动物ES细胞包含一种、两种、三种或更多种靶向遗传修饰。
9.根据实施例8所述的体外培养物,其中所述靶向遗传修饰包括插入、缺失、敲除、敲入、点突变或它们的组合。
10.根据实施例8所述的体外培养物,其中所述靶向遗传修饰包括异源多核苷酸在所述XY ES细胞的基因组中的至少一个插入。
11.根据实施例8至10中任一项所述的体外培养物,其中所述靶向遗传修饰位于常染色体上。
12.根据实施例1至11中任一项所述的体外培养物,其中所述基础培养基表现出50±5mM NaCl、26±5mM碳酸盐以及218±22mOsm/kg。
13.根据实施例1至11中任一项所述的体外培养物,其中所述基础培养基表现出约3mg/mL NaCl、2.2mg/mL碳酸氢钠以及218mOsm/kg。
14.根据实施例1至11中任一项所述的体外培养物,其中所述基础培养基表现出87±5mM NaCl、18±5mM碳酸盐以及261±26mOsm/kg。
15.根据实施例1至11中任一项所述的体外培养物,其中所述基础培养基表现出约5.1mg/mL NaCl、1.5mg/mL碳酸氢钠以及261mOsm/kg。
16.根据实施例1至11中任一项所述的体外培养物,其中所述基础培养基表现出110±5mM NaCl、18±5mM碳酸盐以及294±29mOsm/kg。
17.根据实施例1至11中任一项所述的体外培养物,其中所述基础培养基表现出约6.4mg/mL NaCl、1.5mg/mL碳酸氢钠以及294mOsm/kg。
18.根据实施例1至11中任一项所述的体外培养物,其中所述基础培养基表现出87±5mM NaCl、26±5mM碳酸盐以及270±27mOsm/kg。
19.根据实施例1至11中任一项所述的体外培养物,其中所述基础培养基表现出约5.1mg/mL NaCl、2.2mg/mL碳酸氢钠以及270mOsm/kg。
20.根据实施例1至11中任一项所述的体外培养物,其中所述基础培养基表现出87±5mM NaCl、26±5mM碳酸盐、86±5mM葡萄糖以及322±32mOsm/kg。
21.根据实施例1至11中任一项所述的体外培养物,其中所述基础培养基表现出约5.1mg/mL NaCl、2.2mg/mL碳酸氢钠、15.5mg/mL葡萄糖以及322mOsm/kg。
22.根据实施例1至21中任一项所述的体外培养物,其中在将所述非人哺乳动物XYES细胞引入宿主胚胎中并接着孕育所述宿主胚胎之后,至少80%的F0非人哺乳动物为XY雌性,在达到性成熟后,所述F0 XY雌性非人哺乳动物是可育的。
23.一种用于在F0代中产生可育雌性XY非人哺乳动物的方法,包括:
(a)在适用于在培养中维持非人哺乳动物ES细胞、包含基础培养基和补充物的培养基中,培养具有降低Sry蛋白水平和/或活性的修饰的供体非人哺乳动物XY胚胎干(ES)细胞,其中所述培养基表现出包括以下一者或多者的特征:约200mOsm/kg至小于约329mOsm/kg的渗透压;约11mS/cm至约13mS/cm的电导率;约50mM至约110mM的碱金属卤化物盐浓度;约17mM至约30mM的碳酸盐浓度;约85mM至约130mM的碱金属卤化物盐和碳酸盐总浓度;和/或这些特征之中任何两种或更多种的组合;
(b)将所述供体XY非人哺乳动物ES细胞引入宿主胚胎中;
(c)孕育所述宿主胚胎;以及
(d)获得F0 XY雌性非人哺乳动物,其中在达到性成熟后,所述F0XY雌性非人哺乳动物是可育的。
24.根据实施例23所述的方法,其中所述非人哺乳动物XY ES细胞来自啮齿动物。
25.根据实施例24所述的方法,其中所述啮齿动物为小鼠。
26.根据实施例25所述的方法,其中所述小鼠XY ES细胞为VGF1小鼠ES细胞。
27.根据实施例24所述的方法,其中所述啮齿动物为大鼠或仓鼠。
28.根据实施例23至27中任一项所述的方法,其中所述Sry蛋白的水平和/或活性降低源自Sry基因中的遗传修饰。
29.根据实施例28所述的方法,其中所述Sry基因中的所述遗传修饰包括一个或多个核苷酸的插入、一个或多个核苷酸的缺失、一个或多个核苷酸的置换、敲除、敲入、用异源核酸序列替换内源核酸序列,或它们的组合。
30.根据实施例23至29中任一项所述的方法,其中所述非人哺乳动物ES细胞包含一种、两种、三种或更多种靶向遗传修饰。
31.根据实施例30所述的方法,其中所述靶向遗传修饰包括插入、缺失、敲除、敲入、点突变或它们的组合。
32.根据实施例30所述的方法,其中所述靶向遗传修饰包括异源多核苷酸在所述XY ES细胞的基因组中的至少一个插入。
33.根据实施例30至32中任一项所述的方法,其中所述靶向遗传修饰位于常染色体上。
34.根据实施例23至33中任一项所述的方法,其中所述基础培养基表现出50±5mMNaCl、26±5mM碳酸盐以及218±22mOsm/kg。
35.根据实施例23至33中任一项所述的方法,其中所述基础培养基表现出约3mg/mL NaCl、2.2mg/mL碳酸氢钠以及218mOsm/kg。
36.根据实施例23至33中任一项所述的方法,其中所述基础培养基表现出87±5mMNaCl、18±5mM碳酸盐以及261±26mOsm/kg。
37.根据实施例23至33中任一项所述的方法,其中所述基础培养基表现出约5.1mg/mL NaCl、1.5mg/mL碳酸氢钠以及261mOsm/kg。
38.根据实施例23至33中任一项所述的方法,其中所述基础培养基表现出110±5mM NaCl、18±5mM碳酸盐以及294±29mOsm/kg。
39.根据实施例23至33中任一项所述的方法,其中所述基础培养基表现出约6.4mg/mL NaCl、1.5mg/mL碳酸氢钠以及294mOsm/kg。
40.根据实施例23至33中任一项所述的方法,其中所述基础培养基表现出87±5mMNaCl、26±5mM碳酸盐以及270±27mOsm/kg。
41.根据实施例23至33中任一项所述的方法,其中所述基础培养基表现出约5.1mg/mL NaCl、2.2mg/mL碳酸氢钠以及270mOsm/kg。
42.根据实施例23至33中任一项所述的方法,其中所述基础培养基表现出87±5mMNaCl、26±5mM碳酸盐、86±5mM葡萄糖以及322±32mOsm/kg。
43.根据实施例23至33中任一项所述的方法,其中所述基础培养基表现出约5.1mg/mL NaCl、2.2mg/mL碳酸氢钠、15.5mg/mL葡萄糖以及322mOsm/kg。
44.一种在F1代中产生对于靶向遗传突变为纯合的转基因非人哺乳动物的方法,包括:(a)使所述Sry蛋白的水平和/或活性降低的F0 XY可育雌性与衍生自相同ES细胞克隆的同龄群无性系同胞F0 XY雄性非人哺乳动物杂交,其中所述F0 XY可育雌性非人哺乳动物和所述F0 XY雄性非人哺乳动物各自对于所述遗传突变均为杂合的;以及(b)获得对于所述遗传修饰为纯合的F1子代小鼠。
45.一种用于修饰细胞中Y染色体上的靶基因组座位的方法,包括:(a)提供在Y染色体上包含靶基因组座位的细胞,所述靶基因组座位包含核酸酶试剂的识别位点,(b)向所述细胞中引入(i)所述核酸酶试剂,其中所述核酸酶试剂在第一识别位点处诱导切口或双链断裂;和(ii)包含第一插入多核苷酸的第一靶向载体,所述第一插入多核苷酸侧接第一同源臂和第二同源臂,所述第一同源臂和所述第二同源臂与位置足够接近第一识别位点的第一靶位点和第二靶位点相对应,其中所述第一同源臂和所述第二同源臂的总和为至少4kb但少于150kb;以及(c)鉴定在其基因组中包含在所述靶基因组座位处整合的第一插入多核苷酸的至少一个细胞。
46.一种用于修饰细胞中Y染色体上的靶基因组座位的方法,包括:
(a)提供在Y染色体上包含靶基因组座位的细胞,所述靶基因组座位包含核酸酶试剂的识别位点,
(b)向所述细胞中引入包含第一插入多核苷酸的第一靶向载体,所述第一插入多核苷酸侧接与第一靶位点和第二靶位点相对应的第一同源臂和第二同源臂,其中所述第一同源臂和所述第二同源臂的总和为至少4kb但少于150kb;以及
(c)鉴定至少一个在其基因组中包含在所述靶基因组座位处整合的所述第一插入多核苷酸的细胞。
47.根据实施例45或46所述的方法,其中所述细胞为哺乳动物细胞。
48.根据实施例47所述的方法,其中所述哺乳动物细胞为非人细胞。
49.根据实施例47所述的方法,其中所述哺乳动物细胞来自啮齿动物。
50.根据实施例49所述的方法,其中所述啮齿动物为大鼠、小鼠或仓鼠。
51.根据实施例45至50中任一项所述的方法,其中所述细胞为多能细胞。
52.根据实施例45至50中任一项所述的方法,其中所述哺乳动物细胞为诱导性多能干(iPS)细胞。
53.根据实施例51所述的方法,其中所述多能细胞为非人胚胎干(ES)细胞。
54.根据实施例51所述的方法,其中所述多能细胞为啮齿动物胚胎干(ES)细胞、小鼠胚胎干(ES)细胞或大鼠胚胎干(ES)细胞。
55.根据实施例45以及47至54中任一项所述的方法,其中所述核酸酶试剂为编码核酸酶的mRNA。
56.根据实施例45以及47至54中任一项所述的方法,其中所述核酸酶试剂为锌指核酸酶(ZFN)。
57.根据实施例45以及47至54中任一项所述的方法,其中所述核酸酶试剂为转录激活因子样效应物核酸酶(TALEN)。
58.根据实施例45以及47至54中任一项所述的方法,其中所述核酸酶试剂为大范围核酸酶。
59.根据实施例45以及47至54中任一项所述的方法,其中所述核酸酶试剂为CRISPR RNA指导的Cas9内切核酸酶。
60.一种用于修饰Y染色体的方法,包括在存在包含至少10kb核酸序列的大靶向载体(LTVEC)的情况下使Y染色体暴露于Cas蛋白和CRISPR RNA,其中在暴露于Cas蛋白、CRISPR RNA和LTVEC后,Y染色体被修饰成包含至少10kb的核酸序列。
61.根据实施例60所述的方法,其中所述LTVEC包含至少20kb、至少30kb、至少40kb、至少50kb、至少60kb、至少70kb、至少80kb或至少90kb的核酸序列。
62.根据实施例60所述的方法,其中所述LTVEC包含至少100kb、至少150kb或至少200kb的核酸序列。
63.一种用于修饰Y染色体上的靶基因组座位的方法,包括:(a)提供在Y染色体上包含靶基因组座位的哺乳动物细胞,其中所述靶基因组座位包含向导RNA(gRNA)靶序列;(b)向该哺乳动物细胞中引入:(i)包含第一核酸的大靶向载体(LTVEC),所述第一核酸侧接有与靶基因组座位同源的靶向臂,其中所述LTVEC为至少10kb;(ii)包含第一启动子的第一表达构建体,所述第一启动子有效连接至编码Cas蛋白的第二核酸,以及(iii)包含第二启动子的第二表达构建体,所述第二启动子有效连接至编码向导RNA(gRNA)的第三核酸,所述向导RNA包含杂交至gRNA靶序列和反式激活CRISPR RNA(tracrRNA)的核苷酸序列,其中所述第一启动子和所述第二启动子在该哺乳动物细胞中有活性;以及(c)鉴定在Y染色体上的靶基因组座位处包含靶向遗传修饰的经修饰的哺乳动物细胞。
64.根据实施例63所述的方法,其中所述LTVEC为至少15kb、至少20kb、至少30kb、至少40kb、至少50kb、至少60kb、至少70kb、至少80kb或至少90kb。
65.根据实施例63所述的方法,其中所述LTVEC为至少100kb、至少150kb或至少200kb。
66.根据实施例63所述的方法,其中所述哺乳动物细胞为非人哺乳动物细胞。
67.根据实施例63所述的方法,其中所述哺乳动物细胞为成纤维细胞。
68.根据实施例63所述的方法,其中所述哺乳动物细胞来自啮齿动物。
69.根据实施例68所述的方法,其中所述啮齿动物为大鼠、小鼠或仓鼠。
70.根据实施例63所述的方法,其中所述哺乳动物细胞为多能细胞。
71.根据实施例70所述的方法,其中所述多能细胞为诱导性多能干(iPS)细胞。
72.根据实施例70所述的方法,其中所述多能细胞为小鼠胚胎干(ES)细胞或大鼠胚胎干(ES)细胞。
73.根据实施例70所述的方法,其中所述多能细胞为发育受限的人祖细胞。
74.根据实施例63所述的方法,其中所述Cas蛋白为Cas9蛋白。
75.根据实施例74所述的方法,其中所述gRNA靶序列紧邻地侧接前间区序列邻近基序(PAM)序列。
76.根据实施例63所述的方法,其中所述LTVEC的5’同源臂和3’同源臂的总和为约10kb至约150kb。
77.根据实施例76所述的方法,其中所述LTVEC的5’同源臂和3’同源臂的总和为约10kb至约20kb、约20kb至约40kb、约40kb至约60kb、约60kb至约80kb、约80kb至约100kb、约100kb至约120kb、或约120kb至150kb。
78.根据实施例63所述的方法,其中所述靶向遗传修饰包括:(a)用同源或直系同源核酸序列替换内源核酸序列;(b)内源核酸序列的缺失;(c)内源核酸序列的缺失,其中所述缺失在约5kb至约10kb、约10kb至约20kb、约20kb至约40kb、约40kb至约60kb、约60kb至约80kb、约80kb至约100kb、约100kb至约150kb、或约150kb至约200kb、约200kb至约300kb、约300kb至约400kb、约400kb至约500kb、约500kb至约1Mb、约1Mb至约1.5Mb、约1.5Mb至约2Mb、约2Mb至约2.5Mb、或约2.5Mb至约3Mb的范围内;(d)外源核酸序列的插入;(e)外源核酸序列的插入,所述插入在约5kb至约10kb、约10kb至约20kb、约20kb至约40kb、约40kb至约60kb、约60kb至约80kb、约80kb至约100kb、约100kb至约150kb、约150kb至约200kb、约200kb至约250kb、约250kb至约300kb、约300kb至约350kb、或约350kb至约400kb的范围内;(f)包含同源或直系同源核酸序列的外源核酸序列的插入;(g)包含人和非人核酸序列的嵌合核酸序列的插入;(h)侧接有位点特异性重组酶靶序列的条件性等位基因的插入;(i)有效连接至在所述哺乳动物细胞中有活性的第三启动子的可选标记或报告基因的插入;或(j)它们的组合。
79.根据实施例63所述的方法,其中所述靶基因组座位包含(i)与5’同源臂同源的5’靶序列,以及(ii)与3’同源臂同源的3’靶序列。
80.根据实施例79所述的方法,其中所述5’靶序列和所述3’靶序列相隔至少5kb但少于3Mb。
81.根据实施例79所述的方法,其中所述5’靶序列和所述3’靶序列相隔至少5kb但少于10kb、至少10kb但少于20kb、至少20kb但少于40kb、至少40kb但少于60kb、至少60kb但少于80kb、至少约80kb但少于100kb、至少100kb但少于150kb、或至少150kb但少于200kb、至少约200kb但少于约300kb、至少约300kb但少于约400kb、至少约400kb但少于约500kb、至少约500kb但少于约1Mb、至少约1Mb但少于约1.5Mb、至少约1.5Mb但少于约2Mb、至少约2Mb但少于约2.5Mb、或至少约2.5Mb但少于约3Mb。
82.根据实施例63所述的方法,其中所述第一表达构建体和所述第二表达构建体位于单个核酸分子上。
83.根据实施例63所述的方法,其中所述靶基因组座位包括所述Sry基因座。
84.一种用于非人动物Y染色体上的靶向遗传修饰的方法,包括:(a)根据实施例4所述的方法修饰非人多能细胞的Y染色体上的目标基因组座位,从而产生在Y染色体上包含靶向遗传修饰的经遗传修饰的非人多能细胞;(b)将(a)的所述经修饰的非人多能细胞引入非人宿主胚胎中;以及(c)在代孕母体中孕育包含所述经修饰的多能细胞的所述非人宿主胚胎,其中所述代孕母体产生包含所述靶向遗传修饰的F0子代,其中所述靶向遗传修饰能够通过种系传递。
85.根据实施例84所述的方法,其中所述目标基因组座位包括所述Sry基因座。
86.一种用于修饰细胞中Y染色体上的靶基因组座位的方法,包括:(a)提供在Y染色体上包含靶基因组座位的细胞,所述靶基因组座位包含核酸酶试剂的识别位点,(b)向所述细胞中引入(i)所述核酸酶试剂,其中所述核酸酶试剂在第一识别位点处诱导切口或双链断裂;和(ii)包含第一插入多核苷酸的第一靶向载体,所述第一插入多核苷酸侧接第一同源臂和第二同源臂,所述第一同源臂和所述第二同源臂与位置足够接近第一识别位点的第一靶位点和第二靶位点相对应,其中所述第一同源臂和/或所述第二同源臂的长度为至少400bp但少于1000bp;以及(c)鉴定在其基因组中包含在所述靶基因组座位处整合的第一插入多核苷酸的至少一个细胞。
87.根据实施例86所述的方法,其中所述第一同源臂和/或所述第二同源臂的长度为约700bp至约800bp。
88.根据实施例86所述的方法,其中所述修饰包括内源核酸序列的缺失。
89.根据实施例88所述的方法,其中所述缺失在约5kb至约10kb、约10kb至约20kb、约20kb至约40kb、约40kb至约60kb、约60kb至约80kb、约80kb至约100kb、约100kb至约150kb、或约150kb至约200kb、约200kb至约300kb、约300kb至约400kb、约400kb至约500kb、约500kb至约1Mb、约1Mb至约1.5Mb、约1.5Mb至约2Mb、约2Mb至约2.5Mb、或约2.5Mb至约3Mb的范围内。
90.根据实施例88所述的方法,其中所述缺失为至少500kb。
本发明的方法和组合物可按许多不同的形式实施,并且不应理解为局限于本文所给出的实施例;相反,提供这些实施例是为了使得本公开满足适用的法律要求。全文中类似的数字指代类似的要素。
本文所述方法和组合物所属领域的技术人员在获得以上说明和相关的附图中给出的教导的益处后,可以想到这些方法和组合物的许多修改形式和其他实施例。因此,应当理解,所述方法和组合物并不局限于所公开的具体实施例,并且这些修改形式和其他实施例也包含在所附权利要求的范围内。尽管本文采用了专用术语,但是这些术语只以一般的说明性意义使用,而非用于限制。
以下实例是以说明性方式而非限制性方式提供的。
使用靶向载体形成包含Sry的lacZ替换等位基因的靶向缺失,该靶向载体依次包含大约700bp的上游同源臂;β-半乳糖苷酶编码序列(lacZ)后接多聚腺苷酸化信号;侧接包含人泛素C启动子的loxP位点的新霉素抗性盒,其包含第一外显子、第一内含子及部分第二外显子、新霉素磷酸转移酶编码序列和多聚腺苷酸化信号;以及大约650bp的下游同源臂。使用靶向载体正确靶向Sry基因所形成的等位基因,包含大约1kb Sry开放阅读框的缺失以及用lacZ-neo盒进行的替换,使得β-半乳糖苷酶编码序列在Sry起始密码子处框内融合。使用靶向载体来靶向VGB6(亦称B6A6)C57BL/6和VGF1(亦称F1H4)C57BL6/129F1杂交ES细胞系中的Sry基因。VGF1(F1H4)小鼠ES细胞来源于通过使雌性C57BL/6NTac小鼠与雄性129S6/SvEvTac小鼠杂交而产生的杂交胚胎。因此,VGF1 ES细胞包含来自129S6/SvEvTac小鼠的Y染色体。由VGF1细胞系产生的雌性XY小鼠包含来源于129S6/SvEvTac小鼠的Y染色体。
在TALEN或CRISPR向导RNA与Cas9 DNA内切核酸酶联合作用下,在Sry基因中形成在3bp至1.2kb及更大的范围内的缺失突变,这大概是双链DNA断裂的非同源性末端接合(NHEJ)修复的结果(参见图1)。在Sry基因中包含TALEN诱导突变和CRISPR诱导突变的ES细胞还携带了NIH KOMP项目VG12778 LTVEC(可经由互联网在万维网(www)上以URL“velocigene.com/komp/detail/12778”获取)的随机转基因插入,其包括Sry编码序列的缺失,以及用包含lacZ的插入盒进行的替换,所述lacZ与Sry起始密码子和新霉素抗性基因框内融合,所述新霉素抗性基因侧接38kb同源臂和37kb同源臂并基于来自bMQ文库的BAC(129S7/SvEv Brd-Hprt b-m2)。LTVEC在其同源臂中包含用于Sry表达的所有已知控制元件。其lacZ编码的β-半乳糖苷酶充当Sry基因的组织特异性和发育阶段特异性表达的报告子。在VGB6(亦称B6A6)和VGF1(亦称F1H4)ES细胞系两者中均形成了伴随LTVEC插入的TALEN诱导突变和CRISPR诱导突变。
我们获得了TALEN(TALEN-1),其被设计成靶向Sry基因中的部分HMG框DNA结合基序编码序列(上游识别序列:5'-TCCCGTGGTGAGAGGCAC-3';下游识别序列:5'-TATTTTGCATGCTGGGAT-3')。在多个实验中,TALEN-1都在Sry基因座处有效形成了NHEJ突变。
VGB6和VGF1小鼠ES细胞均形成有TALEN诱导突变。表1包含所有克隆及其携带的缺失突变的尺寸的列表。(表1中的ND指示通过qPCR测定法检测到突变,但尚未确定该突变的确切分子性质。)所有克隆也携带至少一个拷贝的NIH KOMP项目VG12778 LTVEC。
*还包含50bp插入
Sry突变体克隆的显微注射结果示于表2中,而性逆转雌性的繁育结果示于表3中。
*必须在产前对XY雌性ID#1460406实施安乐死,因为其有近足月危象,
无法分娩。通过解剖取出其死亡幼仔(4只雄性,5只雌性),任何一只幼仔均未携带Sry突变。
所有来源于VGB6 ES细胞的带有Sry突变的VelociMice均为雌性,正如对Sry失活所预料的那样(表2)。那些不带有Sry突变但携带至少一个拷贝的NIH KOMP VG12778 LTVEC的克隆仅产生雄性VelociMice(表2,克隆GB4和GG1)。当对17只Sry突变体雌性B6VelociMice进行测交时,在繁育启动约四个月后仅一只受孕(表3),并且必须在产前对该雌性实施安乐死,因为其有近足月危象,无法分娩。通过解剖取出的其死亡幼仔(4只雄性,5只雌性)全为野生型,任何一只幼仔均未携带Sry突变。得出如下结论:几乎所有由VGB6 ES细胞形成的Sry突变体小鼠都是不育的,这与有关Sry突变的文献一致。然而,我们的数据显示用VGF1克隆得出了迥然不同的结果。
首先,像往常一样,在我们的KO-DMEM样低渗透压强度生长培养基中维持VGF1 ES细胞,该培养基具有雌性化作用:在该培养基中生长的一些经显微注射的XY克隆将产生可育XY雌性,即XY雌性现象,即便它们并未携带突变也是如此。一个例子是克隆TH4,其不具有Sry突变但携带至少一个拷贝的NIH KOMP VG12778 LTVEC。该克隆产生2只雌性和6只雄性VelociMice(表2)。另两个不带有Sry突变的VGF1克隆(TA3和TA4,表2)仅产生雄性VelociMice。我们希望确定在Sry中具有突变的VGF1 XY ES细胞是否也可能通过该培养基进行雌性化。换句话讲,它们是否与VGB6 Sry突变体ES细胞不同,会产生一些可育XY Sry突变体雌性?(应当注意,VGB6 ES细胞无法在KO-DMEM样低渗透压强度培养基中维持并保留产生小鼠的能力。)答案是肯定的,如表3中所示。
显微注射了六个VGF1 ES细胞克隆,这些克隆具有5bp至超过1kb范围内的TALEN诱导的小段缺失。所有克隆都产生了雌性VelociMice,让其中32只交配。结果引人注目,所有Sry突变体XY雌性VelociMice都是可育的;每只至少产下一窝幼仔(表3)。许多Sry突变体XY雌性产下幼仔数正常的多窝幼仔,而一些XY雌性仅产下幼仔数较少的一窝或两窝幼仔。在由这些繁育操作得到的经过基因分型的299只F1小鼠之中,大约一半(146只,49%)是正常的XY雄性或正常的XX雌性。174只(58%)F1小鼠为表型雌性,而125只(42%)为表型雄性。26只雌性(雌性中的15%,总F1代中的8.7%)为遗传了突变体Sry等位基因的XY雌性。由于与XY卵母细胞相关的减数分裂不分离事件,在Sry突变体XY雌性VelociMice的F1子代中观察到许多异常的基因型-XXY、XYY、XO、XXYY-其中一些包含突变体Sry等位基因。
已发现了由XY ES细胞有效形成可育XY雌性VelociMice的方法。如果在ES细胞中形成Sry基因中的失活性突变而这些ES细胞一直维持在雌性化生长培养基中,则得到高比例的可育XY雌性小鼠,这些小鼠在与雄性交配时产下大多数具有正常的X染色体和Y染色体的雄性小鼠和雌性小鼠。
通过对来源于F0雌性(其为XY且携带Sry突变)的F1后代进行基因分型,确认或否定LTVEC已正确地靶向小鼠Sry。F1小鼠中LacZ/Neo盒与Sry突变的共分离(如通过Sry LOA测定法评估)非常明确地提示靶向正确。LacZ/Neo未能与该突变共分离则指示初始克隆包含Sry缺失突变(由TALEN诱导),外加基因组中别处的LacZ/Neo转基因插入。
来自带有Sry突变的XY雌性的后代高频率地表现出多种异常核型(包括XXY、XYY和XO)。通过对X染色体和Y染色体上的基因使用不相关等位基因丢失(LOA)测定法,来评估性染色体计数。然后使用LOA测定法确定Sry的拷贝数。推断在Y染色体拷贝数超过Sry拷贝数(例如,1个拷贝的Y和0个拷贝的Sry,或2个拷贝的Y和1个拷贝的Sry)的小鼠中存在突变体Sry等位基因。最后使用TaqMan测定法确定是否存在LacZ和Neo。
在初始组克隆(由Sry LTVEC与TALEN核酸酶一起形成并在KO-DMEM中生长)中,明显的是,LacZ/Neo盒未与Sry突变共分离。来自这些克隆的样本窝仔示于表4中。
在后续组克隆(由Sry LTVEC与TALEN核酸酶一起形成并在DMEM中生长)中,LacZ/Neo盒完全与Sry突变共分离,表明靶向正确。来自这些克隆的典型窝仔示于表5中。
如图2所示,一起使用LTVEC或小靶向载体(smallTVEC)与TALEN核酸酶或CRISPR向导RNA,并联合Cas9 DNA内切核酸酶来形成包含Sry的lacZ替换等位基因的靶向缺失。smallTVEC依次包含大约700至800bp的上游同源臂;β-半乳糖苷酶编码序列(lacZ)后接多聚腺苷酸化信号;侧接包含人泛素C启动子的loxP位点的新霉素抗性盒,其包含第一外显子、第一内含子及部分第二外显子、新霉素磷酸转移酶编码序列和多聚腺苷酸化信号;以及大约700至800bp的下游同源臂。使用靶向载体正确靶向Sry基因所形成的等位基因,包含大约1kb Sry开放阅读框的缺失以及用lacZ-neo盒进行的替换,使得β-半乳糖苷酶编码序列在Sry起始密码子处框内融合。使用靶向载体来靶向VGF1(亦称F1H4)C57BL6/129F1杂交ES细胞系中及VGB6 ES细胞系(亦称B6A6)中的Sry基因。
如表6中所示,产生了使用四个不同gRNA和一个TALEN对而产生的克隆,并通过TaqMan测定法针对裂解和等位基因丢失进行筛选。
LTVEC转基因克隆产生了具有相同lacZ模式的胚胎。图3示出了胚胎中的LacZ表达。
表7报告了来源于生长在常规DMEM类培养基中的ES细胞的XY雌性的可育性结果,这些ES细胞具有在TALEN协助下对Sry的LTVEC靶向缺失-替换突变。与采用生长在KO-DEMEM类培养基中的ES细胞进行的类似实验的结果(表3)相比,我们意外地发现,DMEM类培养基中的LTVEC靶向产生了具有正确靶向的Sry缺失和lacZ-neo插入的克隆。来源于四个靶向克隆的41只XY
如图4中所示,使用ZFN靶向Kdm5d和Usp9y基因,在Y染色体上产生大段缺失(500kb或更大)。表8提供了Y染色体上的锌指序列的例子。
在一个实验中,使用ZFN mRNA对Kdm5d-ZFN5(NM011419-r17347a1)/ZFN6(NM011419-17353a1)和Usp9y-ZFN3(NM148943-r11830a1)/ZFN4(NM148943-11836a1)(各10μg)并使用靶向Ch25h基因的LTVEC(0.67μg),对来自VGB6克隆D-G5和E-G7(表3)的330万个ES细胞进行电穿孔,从而提供对嘌呤霉素抗性的选择。挑取嘌呤霉素抗性集落,针对缺失进行筛选。结果示于表9中。
表10示出了大于500kb缺失的确切尺寸,这些尺寸针对来源于E-G7亲本克隆(表3)的一个缺失克隆(4306A-D5)以及来源于D-G5亲本克隆(表3)的两个缺失克隆(4306E-C4和4306F-A12)进行精确确定。
Kdm5d、Eif2s3y、Uty、Ddx3y和Usp9y基因的缺失(图4)在缺失等位基因丢失测定法及如图5中所示的DNA测序中进行确认。克隆4306A-D5在显微注射到8-细胞期胚胎中并移植进代孕母体之后,产生了九只XY雌性全ES细胞来源的VelociMice。来自克隆4306A-D5的XY雌性均不可育。
联合利用CRISPR向导RNA和Cas9 DNA内切核酸酶,来形成靶向Kdm5d基因与Usp9y基因之间的区域的Y染色体上的大段缺失。gRNA被设计成靶向Kdm5d基因和Usp9y基因。以下gRNA被设计成靶向Kdm5d:Kdm5dgA(向导#1)UUUGCCGAAUAUGCUCUCGU(SEQ ID NO:66);Kdm5dgB(向导#2)UUGCCGAAUAUGCUCUCGUG(SEQ ID NO:67);以及Kdm5dgC(向导#5)CGGGCAUCUCCAUACUCCCU(SEQ ID NO:68)。以下gRNA被设计成靶向Usp9y:Usp9ygA(向导#1)UAGCUCGUUGUGUAGCACCU(SEQ ID NO:69);Usp9ygB(向导#1)UAUAGUUUCUUCGGGGUAAC(SEQ IDNO:70);以及Usp9ygC(向导#2)GGAUACCCUUCUAUAGGCCC(SEQ ID NO:71)。
使用5μg表达Cas9的质粒及各10μg表达Kdm5d gRNA B和Usp9y gRNA C的质粒并使用靶向Ch25h基因的LTVEC(0.67μg),对VGF1小鼠ES细胞进行电穿孔,从而提供对嘌呤霉素抗性的选择。
如图4中所示,使用Kdm5dgB(gRNA B)和Usp9ygC(gRNA C)来靶向Kdm5d基因和Usp9y基因的缺失。通过对Kdm5d基因、Usp9y基因、这两者间的基因(Eif2s3y、Uty和Ddx3y)以及靶向缺失外部的基因(Zfy2和Sry)的序列执行等位基因丢失测定法,来筛选所得克隆的缺失。如表11中所示,获得了包含大段缺失的四个克隆。克隆R-A8在显微注射到8-细胞期胚胎中并移植进代孕母体之后,产生了七只XY雄性和3只XY雌性全ES细胞来源的VelociMice。
本说明书中提到的所有出版物和专利申请都表示本发明所属领域的技术人员的水平。所有出版物和专利申请均以引用方式并入本文,其程度就如同每篇单独的出版物或专利申请被具体且单独地指明以引用方式并入。如果与引用相关的信息诸如保藏号随时间而变化,则指的是在本申请的有效提交日期时生效的信息版本,所述有效提交日期意指实际提交日期或首先提供该引用的优先申请的日期。本发明的方面、步骤或特征可与任何其他方面、步骤或特征组合使用,除非从任何实施例的上下文可以明显看出不是这样。对范围的提及包括该范围内的任何整数、该范围内的任何子范围。对多个范围的提及包括此类范围的组合。
序列表
<110> 大卫·弗伦杜威
古斯塔沃·德罗格特
安东尼·加戈利亚地
琼科·库诺
沃基特克·奥尔巴赫
大卫·M.·巴伦苏埃拉
<120>用于所靶向遗传修饰的方法和组合物,
以及这些组合物的使用方法
<130> 057766-463545
<150> US 62/017,582
<151> 2014-06-26
<150> US 62/017,627
<151> 2014-06-26
<160> 71
<170>适用于Windows 4.0版的FastSEQ
<210> 1
<211> 23
<212> DNA
<213>人工序列
<220>
<223>连接至向导RNA (gRNA)
的靶基因座
<220>
<221> misc_feature
<222> 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18,
19, 20, 21
<223> n = A、T、C或G
<400> 1
gnnnnnnnnn nnnnnnnnnn ngg 23
<210> 2
<211> 80
<212> RNA
<213>人工序列
<220>
<223>向导RNA (gRNA)
<400> 2
guuuuagagc uagaaauagc aaguuaaaau aaggcuaguc cguuaucaac uugaaaaagu 60
ggcaccgagu cggugcuuuu 80
<210> 3
<211> 42
<212> RNA
<213>人工序列
<220>
<223>向导RNA (gRNA)
<400> 3
guuuuagagc uagaaauagc aaguuaaaau aaggcuaguc cg 42
<210> 4
<211> 30
<212> RNA
<213>人工序列
<220>
<223> a crRNA
<400> 4
guuuuagagc uagaaauagc aaguuaaaau 30
<210> 5
<211> 33
<212> RNA
<213>人工序列
<220>
<223> a crRNA
<400> 5
guuuuagagc uagaaauagc aaguuaaaau aag 33
<210> 6
<211> 26
<212> RNA
<213>人工序列
<220>
<223> a crRNA
<400> 6
gaguccgagc agaagaagaa guuuua 26
<210> 7
<211> 12
<212> RNA
<213>人工序列
<220>
<223> a tracrRNA
<400> 7
aaggcuaguc cg 12
<210> 8
<211> 50
<212> RNA
<213>人工序列
<220>
<223> a tracrRNA
<400> 8
aaggcuaguc cguuaucaac uugaaaaagu ggcaccgagu cggugcuuuu 50
<210> 9
<211> 23
<212> DNA
<213>人工序列
<220>
<223>连接至向导RNA (gRNA)
的靶基因座
<220>
<221> misc_feature
<222> 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18,
19, 20, 21
<223> n = A、T、C或G
<400> 9
gnnnnnnnnn nnnnnnnnnn ngg 23
<210> 10
<211> 23
<212> DNA
<213>人工序列
<220>
<223> VG-1 gRNA靶序列
<400> 10
ccatgaatgc atttatggtg tgg 23
<210> 11
<211> 23
<212> DNA
<213>人工序列
<220>
<223> VG-2 gRNA靶序列
<400> 11
ccgtggtgag aggcacaagt tgg 23
<210> 12
<211> 23
<212> DNA
<213>人工序列
<220>
<223> VG-3 gRNA靶序列
<400> 12
gcaagcagct gggatgcagg tgg 23
<210> 13
<211> 20
<212> DNA
<213>人工序列
<220>
<223>合成sryTF1引物
<400> 13
cgtggtgaga ggcacaagtt 20
<210> 14
<211> 19
<212> DNA
<213>人工序列
<220>
<223>合成sryTR1引物
<400> 14
gagatcagca agcagctgg 19
<210> 15
<211> 22
<212> DNA
<213>人工序列
<220>
<223>合成sryTP1探针
<400> 15
cccagcagaa tcccagcatg ca 22
<210> 16
<211> 18
<212> DNA
<213>人工序列
<220>
<223>合成sryTF2引物
<400> 16
tggagggcca tgtcaagc 18
<210> 17
<211> 18
<212> DNA
<213>人工序列
<220>
<223>合成sryTR2引物
<400> 17
acaagttggc ccagcaga 18
<210> 18
<211> 26
<212> DNA
<213>人工序列
<220>
<223>合成sryTP2探针
<400> 18
tgaatgcatt tatggtgtgg tcccgt 26
<210> 19
<211> 20
<212> DNA
<213>人工序列
<220>
<223>合成sryTF3引物
<400> 19
atgaatgcat ttatggtgtg 20
<210> 20
<211> 18
<212> DNA
<213>人工序列
<220>
<223>合成sryTR3引物
<400> 20
aggtggaaaa gccttaca 18
<210> 21
<211> 23
<212> DNA
<213>人工序列
<220>
<223>合成sryTP3探针
<400> 21
ccgtggtgag aggcacaagt tgg 23
<210> 22
<211> 19
<212> DNA
<213>人工序列
<220>
<223>合成sryTF4引物
<400> 22
gtgtggtccc gtggtgaga 19
<210> 23
<211> 20
<212> DNA
<213>人工序列
<220>
<223>合成sryTR4引物
<400> 23
agatcagcaa gcagctggga 20
<210> 24
<211> 24
<212> DNA
<213>人工序列
<220>
<223>合成sryTP4探针
<400> 24
aagttggccc agcagaatcc cagc 24
<210> 25
<211> 24
<212> DNA
<213>人工序列
<220>
<223>合成sryTF5引物
<400> 25
catgcaaaat acagagatca gcaa 24
<210> 26
<211> 14
<212> DNA
<213>人工序列
<220>
<223>合成sryTR5引物
<400> 26
ggaaaagcct taca 14
<210> 27
<211> 15
<212> DNA
<213>人工序列
<220>
<223>合成sryTP5探针
<400> 27
cagctgggat gcagg 15
<210> 28
<211> 20
<212> DNA
<213>人工序列
<220>
<223>合成648TUF引物
<400> 28
gatcagcaag cagctgggat 20
<210> 29
<211> 19
<212> DNA
<213>人工序列
<220>
<223>合成648TUP探针
<400> 29
caggtggaaa agccttaca 19
<210> 30
<211> 73
<212> DNA
<213>人工序列
<220>
<223>合成
<400> 30
aattaccaca tcttttgcct attgccaccg cttgaggagt tgttggagaa gtctcattgg 60
aagaatcaga tga 73
<210> 31
<211> 65
<212> DNA
<213>人工序列
<220>
<223>合成
<400> 31
aattaccaca tctttttgcc tattgtccaa atactggaga agtctcattg caacgaatca 60
gatga 65
<210> 32
<211> 63
<212> DNA
<213>人工序列
<220>
<223>合成
<400> 32
aattaccaca tcttttgcct attgacaaaa tgttggagaa gtctcatggg aagaatcaaa 60
ttg 63
<210> 33
<211> 90
<212> DNA
<213>人工序列
<220>
<223>合成
<400> 33
ctatcagata ggatatttta gagttttcat attgtatgga ggagttgttg gagaagtctc 60
attggaagaa tcagatgatg agatcttaat 90
<210> 34
<211> 69
<212> DNA
<213>人工序列
<220>
<223>合成
<400> 34
ctatcagata ggatatttta gagttgttgg agaagtctca ttggaagaat cagatgatga 60
gatcttaat 69
<210> 35
<211> 69
<212> DNA
<213>人工序列
<220>
<223>合成
<400> 35
ctatcagata ggatatttta gagttgttgg agaagtctca ttggaagaat cagatgatga 60
gatcttaat 69
<210> 36
<211> 69
<212> DNA
<213>人工序列
<220>
<223>合成
<400> 36
ctatcacata ggatatttta gagttgttgg cgaagtctca ttggaagaat cagatgatga 60
gatcttaat 69
<210> 37
<211> 69
<212> DNA
<213>人工序列
<220>
<223>合成
<400> 37
tcagatagga tattttagag ttttcatatt gtatggttat aggaggagtt gttggagaag 60
tctcattgg 69
<210> 38
<211> 49
<212> DNA
<213>人工序列
<220>
<223>合成
<400> 38
tcagatagga tattttagag ttttcatatt gttggagaag tctcattgg 49
<210> 39
<211> 49
<212> DNA
<213>人工序列
<220>
<223>合成
<400> 39
tcagataggg tattttagag ttttcatatt gttggagaag tctcatggg 49
<210> 40
<211> 49
<212> DNA
<213>人工序列
<220>
<223>合成
<400> 40
tcccatagga tattttagag ttctcatatt gctgccgaag tctcattgc 49
<210> 41
<211> 39
<212> DNA
<213>人工序列
<220>
<223>合成
<400> 41
ttttagagtt tttcatattg ttgggagaag tctcattgg 39
<210> 42
<211> 28
<212> DNA
<213>人工序列
<220>
<223>合成KDM5D ZFN1靶序列
<400> 42
ttaggtaggt agacagggat gttttctg 28
<210> 43
<211> 28
<212> DNA
<213>人工序列
<220>
<223>合成KDM5D ZFN2靶序列
<400> 43
atccagtctc tgaaggaagc tctgacta 28
<210> 44
<211> 28
<212> DNA
<213>人工序列
<220>
<223>合成KDM5D ZFN3靶序列
<400> 44
caaaagcttc agggggactc ttacactc 28
<210> 45
<211> 28
<212> DNA
<213>人工序列
<220>
<223>合成KDM5D ZFN4靶序列
<400> 45
tttgagcagg ctacacagga gtatactt 28
<210> 46
<211> 28
<212> DNA
<213>人工序列
<220>
<223>合成KDM5D ZFN5靶序列
<400> 46
aagcggtggc aataggcaaa agatgtgg 28
<210> 47
<211> 28
<212> DNA
<213>人工序列
<220>
<223>合成KDM5D ZFN6靶序列
<400> 47
ctgaagtccc caagggagta tggagatg 28
<210> 48
<211> 28
<212> DNA
<213>人工序列
<220>
<223>合成KDM5D ZFN7靶序列
<400> 48
agaaagcggt ggcaataggc aaaagatg 28
<210> 49
<211> 28
<212> DNA
<213>人工序列
<220>
<223>合成KDM5D ZFN8靶序列
<400> 49
aagtccccaa gggagtatgg agatgccc 28
<210> 50
<211> 28
<212> DNA
<213>人工序列
<220>
<223>合成DDX3Y ZFN1靶序列
<400> 50
actccaacga ctatgaccac tccgttca 28
<210> 51
<211> 28
<212> DNA
<213>人工序列
<220>
<223>合成DDX3Y ZFN2靶序列
<400> 51
acagatcaga tgaagatgac tggtcaaa 28
<210> 52
<211> 28
<212> DNA
<213>人工序列
<220>
<223>合成DDX3Y ZFN3靶序列
<400> 52
ctttcaagga aaaaaagaac aaaaccca 28
<210> 53
<211> 28
<212> DNA
<213>人工序列
<220>
<223>合成DDX3Y ZFN4靶序列
<400> 53
ggtctgtgat aaggacagtt caggatgg 28
<210> 54
<211> 28
<212> DNA
<213>人工序列
<220>
<223>合成DDX3Y ZFN5靶序列
<400> 54
taaatctgac tgagaatggg tagtagaa 28
<210> 55
<211> 28
<212> DNA
<213>人工序列
<220>
<223>合成DDX3Y ZFN6靶序列
<400> 55
cagatggtcc aggagaggct ttgaaggc 28
<210> 56
<211> 28
<212> DNA
<213>人工序列
<220>
<223>合成DDX3Y ZFN7靶序列
<400> 56
attgggcttc cctctggaat cacgagat 28
<210> 57
<211> 28
<212> DNA
<213>人工序列
<220>
<223>合成DDX3Y ZFN8靶序列
<400> 57
tttcagtgat cgtggaagtg gatccagg 28
<210> 58
<211> 28
<212> DNA
<213>人工序列
<220>
<223>合成USP9Y ZFN1靶序列
<400> 58
ctggtttgga aatcgtactg taaaagac 28
<210> 59
<211> 28
<212> DNA
<213>人工序列
<220>
<223>合成USP9Y ZFN2靶序列
<400> 59
gcaaagaggt tgaggatttg gacatatt 28
<210> 60
<211> 28
<212> DNA
<213>人工序列
<220>
<223>合成USP9Y ZFN3靶序列
<400> 60
gaggagttgt tggagaagtc tcattgga 28
<210> 61
<211> 28
<212> DNA
<213>人工序列
<220>
<223>合成USP9Y ZFN4靶序列
<400> 61
atatgaacaa ggccaaggtg atgctcca 28
<210> 62
<211> 28
<212> DNA
<213>人工序列
<220>
<223>合成USP9Y ZFN5靶序列
<400> 62
actcagaaga aggattagga atgctttg 28
<210> 63
<211> 28
<212> DNA
<213>人工序列
<220>
<223>合成USP9Y ZFN6靶序列
<400> 63
atgcttagaa atgtatcagt tcatcttg 28
<210> 64
<211> 28
<212> DNA
<213>人工序列
<220>
<223>合成USP9Y ZFN7靶序列
<400> 64
tccataagga ttttggaaaa agacacag 28
<210> 65
<211> 28
<212> DNA
<213>人工序列
<220>
<223>合成USP9Y ZFN8靶序列
<400> 65
aggctgtgag tggatggaag tttgaaat 28
<210> 66
<211> 20
<212> RNA
<213>人工序列
<220>
<223>合成Kdm5dgA gRNA 1
<400> 66
uuugccgaau augcucucgu 20
<210> 67
<211> 20
<212> RNA
<213>人工序列
<220>
<223>合成Kdm5dgB gRNA 2
<400> 67
uugccgaaua ugcucucgug 20
<210> 68
<211> 20
<212> RNA
<213>人工序列
<220>
<223>合成Kdm5dgC gRNA 5
<400> 68
cgggcaucuc cauacucccu 20
<210> 69
<211> 20
<212> RNA
<213>人工序列
<220>
<223>合成Usp9ygA gRNA 1
<400> 69
uagcucguug uguagcaccu 20
<210> 70
<211> 20
<212> RNA
<213>人工序列
<220>
<223>合成Usp9ygB gRNA 1
<400> 70
uauaguuucu ucgggguaac 20
<210> 71
<211> 20
<212> RNA
<213>人工序列
<220>
<223>合成Usp9ygC gRNA 2
<400> 71
ggauacccuu cuauaggccc 20
- 用于靶向遗传修饰的方法和组合物,以及这些组合物的使用方法
- 用于靶向遗传修饰的方法和组合物,以及这些组合物的使用方法