一种新型透明质酸酶及其基因工程菌、构建方法与应用
文献发布时间:2024-04-18 19:58:53
技术领域
本发明属于生物工程技术领域,具体涉及一种新型透明质酸酶及其基因工程菌、构建方法与应用。
背景技术
透明质酸(Hyaluronic acid,HA)是由D-葡萄糖醛酸与N-乙酰氨基葡萄糖以β-1,3和β-1,4糖苷键交替连接而成的线性多糖。因其具有独特的黏弹性、吸湿性、非免疫原性和生物相容性,在医药、化妆品和食品领域具有广泛的应用。随着对HA研究的不断深入,越来越多的研究表明HA的生物学功能与其分子量大小密切相关。高分子量HA具有良好的保水性,主要用于骨科伤口愈合和化妆品添加等方面;中分子量HA具有很好的润滑性,常用于滴眼液和皮肤烧伤愈合等方面;低分子量HA可以口服吸收,能应用于保健品和养颜美容等方面;HA寡糖生理活性较高,具有促进骨和血管生成,增强免疫调等功能。因此,如何制备不同分子量的HA及HA寡糖以满足不同领域的需求是当前研究的热点问题。
目前,市售的HA多为发酵法生产的高分子量和中分子量的HA。低分子量和HA寡糖主要通过物理法、化学法和酶解法获得。但物理法效率比较低,需要同其他方法结合使用;化学法反应条件较为剧烈,对特定分子量的可控性差、易破坏产物的特有结构,造成HA生物活性降低等问题;而酶解法温和高效,能较好保留底物活性,且可以控制反应时间来控制目的产物的分子量。因此,酶解法是制备有生物活性低分子量HA和HA寡糖的重要方法。
透明质酸酶(hyaluronidase,HAase)是一种内源性糖苷酶,能将HA分解成小分子HA或二糖。根据酶的结构、作用机制、底物特异性以及降解产物的差异性,HAase常被分为三类:哺乳动物透明质酸酶(分类号:EC 3.2.1.35)、无脊椎动物透明质酸酶(分类号EC3.2.1.36)和微生物透明质酸酶(分类号:EC 4.2.2.1)。其中,微生物来源的HAase因其来源不受限制,且研究机制相对成熟,成为了不少学者的研究对象。不少研究曾尝试引入HAase来实现对特定HA分子量的生产。虽然,利用HAase能够实现对HA分子量范围的调控,但目前主要是水解已经纯化的高分子量HA来获得,这种先合成再降解的生产模式在一定程度上造成了生物能量上的浪费。加之HAase的产业化一直未得到突破,市场上尚没有针对特定低分子量HA降解的商业用酶,少量上市HAase的价格也较为昂贵;且已发现的HAase种类较少、活性较低,这也就极大地限制了微生物来源HAase的工业化应用。因此,未来挖掘更多新颖的微生物来源的HAase,以及更加深入地了解其功能和性质,并构建能够一步法发酵生产高、中、低分子量HA及HA寡糖的工程菌,能够极大地简化工业程序的复杂性,降低生产成本,具有重要的研究意义。
发明内容
本发明所要解决的技术问题是针对现有技术的不足,提供一种新型透明质酸酶及其基因工程菌、构建方法与应用。
为了解决上述技术问题,本发明采用的技术方案如下:
一种新型透明质酸酶编码基因SthylO,所述透明质酸酶编码基因SthylO来源于耐盐放线菌S.thermolilacinus SPC6,其经密码子优化后的核苷酸序列如SEQ ID NO.1所示。该基因与已知的HAase基因同源性低,与水蛭来源的HAase同源性为14.89%,与兽疫链球菌来源的HAase同源性为19.68%。
上述新型透明质酸酶编码基因SthylO编码的透明质酸酶也在本发明所保护的范围之内。
其中,所述的透明质酸酶具有25kDa的分子量,属于透明质酸裂解酶、透明质酸-4-糖苷水解酶。
一种高产透明质酸酶的基因工程菌,是以质粒pET-28a(+)为载体,以E.coli BL21为宿主,通过克隆透明质酸酶编码基因SthylO得到的。
上述基因工程菌的构建方法,包括如下步骤:
(1)以SthylO基因序列为模板,通过PCR反应进行目的基因扩增,并用限制酶Nde I和Xho I对载体pET-28a(+)和目的基因进行线性化处理,再连接得到重组质粒pET-28a(+)-SthylO;
(2)将步骤(1)获得的重组质粒pET-28a(+)-SthylO通过化学转化法转入至中间克隆宿主E.coli DH5α中,对验证正确的阳性转化子进行质粒提取,再将完整质粒通过化学转化法转入至表达宿主E.coli BL21的感受细胞中,复苏1h后将转化液离心并保留适当液体重悬菌体,随后将其涂布到含有硫酸卡那霉素(25μg/mL)的LB琼脂平板上,37℃,静置培养12h。最后对克隆子进行验证,验证成功的阳性克隆子即为高产透明质酸酶的基因工程菌。
上述基因工程菌在生产透明质酸酶中的应用也在本发明所保护的范围之内。
具体的,将高产透明质酸酶的基因工程菌活化后接种到LB培养基中,37℃,200rpm条件下种子培养12h得到种子液;将种子液以6%v/v的接种量转接至LB培养基中进行发酵;当菌体的OD
其中,所述的LB培养基,其配方为:NaCl 5~15g/L、胰蛋白胨5~15g/L、酵母粉2~7g/L。
其中,所述的发酵,其条件为:20~37℃,200rpm;所述的诱导培养,其条件为:20~37℃,200rpm,诱导18~34h,pH 6.0~8.0。
优选的,当菌体的OD
优选的,所述的发酵,其条件为:37℃,200rpm;所述的诱导培养,其条件为:30℃,200rpm,诱导28h,pH 7.0。
对诱导培养获得的含有透明质酸酶的发酵液进行低温离心,去除上清液,将菌体用pH值6.0的PBS缓冲液洗涤2~3次,再溶入PBS缓冲液中形成菌体重悬液,利用超声破碎仪对菌体重悬液进行破碎,得到粗酶液;将粗酶液离心,收集上清用0.22μm滤膜进行抽滤,采用AKTA纯化系统,利用镍柱亲和层析法进行纯化得到纯酶液;将纯酶液置于超滤管中进行浓缩,并除去金属离子,随后对其进行酶学性质分析,包括最适反应温度及稳定性、最适反应pH及稳定性、金属离子对酶的影响、酶的底物特异性研究、动力学参数的测定。
其中,反应温度为20~70℃,优选的最适反应温度为40℃;在20~40℃的温度范围内,透明质酸酶的相对活力值均在70%以上,说明其在该范围内具有较好的热稳定性。
其中,反应pH为4.0~8.9,优选的最适反应pH为6.0;pH值在5.0~7.2之间,透明质酸酶的相对活力都在70%以上,说明其在该范围内相对稳定。
其中,在研究金属离子对酶的影响中,在酶反应体系里,分别额外添加含有不同金属离子,浓度为0.1M的KCl、NaCl、CoCl
其中,在酶的底物特异性研究中,发现透明质酸酶具有一定的特异性,只能显著降解HA,而不能降解同系的甘露聚糖、硫酸软骨素、硫酸软骨素(鸡)、肝素钠和结冷胶。
一种高产透明质酸的重组解淀粉芽孢杆菌,是以解淀粉芽孢杆菌CF为宿主,以质粒pMA5-sthasA-cghasB为表达载体,通过克隆透明质酸酶编码基因SthylO、信号肽编码基因SpyncM和IPTG诱导型启动子编码基因P
其中,所述的信号肽编码基因SpyncM来源于枯草芽孢杆菌B.subtilis 168,其核苷酸序列如SEQ ID NO.2所示;所述的IPTG诱导型启动子编码基因P
上述重组解淀粉芽孢杆菌的构建方法,包括如下步骤:
(a)分别利用各自引物单独扩增IPTG诱导型启动子编码基因P
(b)将步骤(a)获得的P
上述重组解淀粉芽孢杆菌在发酵制备高、中、低分子量透明质酸及透明质酸寡糖中的应用也在本发明所保护的范围之内。
首先利用上述重组解淀粉芽孢杆菌发酵生产高、中、低分子量的透明质酸及透明质酸寡糖,并对IPTG添加浓度、IPTG添加时间进行了优化。
具体的,将重组解淀粉芽孢杆菌37℃活化后接种到种子培养基中,200rpm、37℃下培养10~12h;将种子液以6%v/v的接种量接种于发酵培养基中32℃,pH 7.0,200rpm发酵,并在发酵开始后的0~48h添加0.1~1.2mM的IPTG进行诱导培养,32℃,pH 7.0,200rpm,诱导12~48h;
其中,所述的种子培养基,其配方为:蔗糖10~30g/L、酵母粉5~15g/L、牛肉膏5~15g/L、MgSO
优选的,所述的IPTG,其最佳添加浓度为0.5mM。
优选的,所述的IPTG,其最佳添加时间为发酵开始36h后添加。
然后将种子液以4~8%v/v的接种量接种于装有发酵培养基的发酵罐中,在30~34℃,pH 6.0~8.0,400~600rpm,通气量4~6vvm条件下进行5L发酵罐放大培养,当发酵开始36h后,添加0.5mM的IPTG进行诱导培养,通过收集诱导开始12h、24h、48h后的发酵液上清,对不同诱导时间透明质酸的相对分子量进行了研究。经高效液相分析,并结合葡聚糖标准曲线,换算得到:未诱导时透明质酸的产量为和分子量分别为5.57g/L,1.14×10
最后对不同分子量的透明质酸及透明质酸寡糖进行分离纯化,并对其抗氧化活性进行分析。结果发现,(1)在DPPH自由基清除活性测定中,高、中、低分子量HA的清除效率较低,仅有20~30%,而HA寡糖对DPPH自由基的清除效率较高,当HA浓度为5g/L时,清除率为57.66%,是相同浓度下高、中、低分子量HA清除率的2.1~3.0倍。(2)在羟自由基清除活性测定中,当HA浓度<4g/L时,不同分子量的HA对羟自由基的清除效率与HA浓度呈正相关,在5g/L逐渐趋于平稳,此时高、中、低分子量HA及HA寡糖的清除效率分别67.59、76.21、90.62、99.60%。同时,5g/L的HA寡糖与2g/L Vc的清除率相当,具有明显的清除效果。(3)在ABTS自由基清除活性测定中,不同分子量的HA对ABTS自由基的清除效率随浓度的升高而逐渐增加。其中高、中、低分子量HA的清除效率较HA寡糖低,有48~82%;HA寡糖对ABTS自由基的清除效率最高,当HA浓度为5g/L时,清除率为99.91%,与3g/L的Vc具有同等的清除效果。有益效果:
(1)本发明提供了一种来源于耐盐放线菌SPC6的新型HAase编码基因SthylO,该基因与已知的HAase基因同源性低,编码的HAase具有优异的HA降解功能。
(2)本发明提供了一株高产HAase的基因工程菌E.coli BL21 pET-28a(+)-SthylO,该基因工程菌发酵生产的HAase具有良好的热稳定性及酸碱稳定性,同时还具有底物特异性。
(3)本发明提供了一株合成高、中、低分子量HA及HA寡糖的重组解淀粉芽孢杆菌B.amyloliquefaciens CF pMA5-sthasA-cghasB-P
(4)本发明提供的发酵生产透明质酸的工艺水平较高,能够简化工业化程序的复杂性,降低生产成本,极具工业化应用价值。
附图说明
下面结合附图和具体实施方式对本发明做更进一步的具体说明,本发明的上述和/或其他方面的优点将会变得更加清楚。
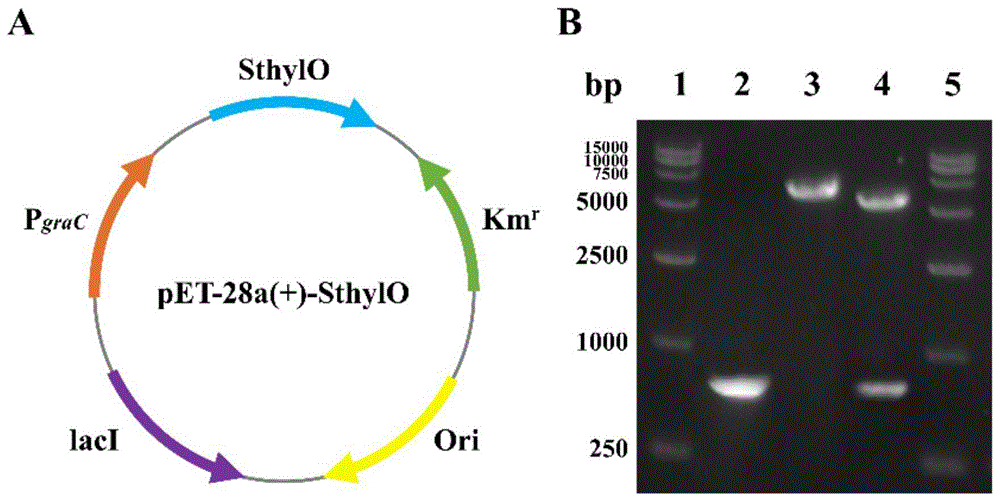
图1(A)重组质粒pET-28a(+)-SthylO构建示意图;(B)重组质粒琼脂糖凝胶分析验证图,注:1:DL 15000DNA marker;2:基因SthylO的PCR扩增产物;3:重组质粒Xho I单酶切验证;4:重组质粒Nde I和Xho I双酶切验证;5:DL 15000DNA marker。
图2SthylO表达条件优化。(A)诱导剂浓度对SthylO表达的影响;(B)诱导初始菌体浓度对SthylO表达的影响;(C)诱导时间对SthylO表达的影响;(D)诱导温度对SthylO表达的影响。
图3(A)SthylO最适温度及其稳定性探究;(B)SthylO最适pH及其稳定性探究。
图4(A)金属离子对SthylO活性影响的研究;(B)SthylO底物特异性的研究,注:1:HA;2:甘露聚糖;3:硫酸软骨素;4:硫酸软骨素(鸡);5:肝素钠;6:结冷胶。
图5(A)SthylO反应速率曲线;(B)SthylO的双倒数曲线。
图6(A)重组质粒pMA5-sthasA-cghasB-P
图7(A)IPTG浓度对重组解淀粉芽孢杆菌发酵产HA的影响;(B)IPTG添加时间对重组解淀粉芽孢杆菌发酵产HA的影响。
图8(A)5L发酵罐扩大培养发酵进程图;(B)不同诱导时间对HA分子量大小的影响。
图9高、中、低分子量HA及HA寡糖的抗氧化活性分析。(A)DPPH自由基清除活性测定;(B)羟自由基清除活性测定;(C)ABTS自由基清除活性测定。
具体实施方式
下述实施例中所述实验方法,如无特殊说明,均为常规方法;所述试剂和材料,如无特殊说明,均可从商业途径获得。
下面结合具体实施例对本发明进一步进行描述。
下述实施例中所用的主要实验材料和试剂盒如下表1所示。
表1主要实验材料
实施例1:高产透明质酸酶工程菌的构建
(1)SthylO密码子优化
一种透明质酸酶SthylO,来源于耐盐放线菌(S.thermolilacinus)SPC6。按照解淀粉芽孢杆菌的密码子偏好性将该酶优化后全基因合成SthylO基因,其核苷酸序列如SEQ IDNO.1所示。目的基因密码子偏好性优化和全基因合成均由通用生物系统(安徽)有限公司完成。
(2)高产透明质酸酶工程菌的构建
以人工合成的SthylO基因序列为模板,引物Sthyl-F/Sthyl-R用于扩增SthylO基因,并用限制酶Nde I和Xho I对载体pET-28a(+)进行线性化处理,再通过一步克隆试剂盒连将线性片段和载体进行连接,得到重组质粒pET-28a(+)-SthylO,如图1A所示。再将表达质粒pET-28a(+)-SthylO通过化学转化法转入中间克隆宿主E.coli DH5α中,涂布在含有25mg/mL硫酸卡那霉素的LB固体培养基上,37℃恒温培养12h;挑取阳性克隆子于5mL含有25mg/mL硫酸卡那霉素的LB液体培养基中,37℃、200rpm培养12h获得转化子,利用引物pET-F/R对转化子进行验证,验证正确的阳性转化子进行质粒提取,并送测序。如图1B所示,SthylO基因已经成功连接在表达载体pET-28a(+)上。最后再将验证成功的完整质粒通过化学转化法转入表达宿主E.coli BL21中,即获得高产透明质酸酶的基因工程菌E.coli BL21pET-28a(+)-SthylO。
其中,扩增编码基因SthylO及转化子验证所用引物序列如表2所示;PCR扩增反应体系为:5×SF Buffer 10μL、dNTP Mix 2μL、Phanta HS Super-Fidelity DNAPolymerase2μL、上游引物2μL、下游引物2μL、模板DNA 2μL、ddH
表2扩增编码基因SthylO及转化子验证所用引物序列
实施例2:高产透明质酸酶工程菌发酵合成透明质酸酶及酶学性质研究
(1)将实施例1构建得到的基因工程菌在37℃培养温度下活化后,将菌株接种到LB培养基中,37℃,200rpm培养12h,得到种子液;将种子液以6%v/v的接种量转接至发酵培养基(配方为:NaCl 10g/L、胰蛋白胨10g/L、酵母粉5g/L)中,37℃,200rpm进行发酵;当菌体的OD
通过考察诱导剂浓度、诱导初始菌体浓度、诱导时间、诱导温度对基因工程菌表达SthylO的影响,优化发酵生产透明质酸酶的工艺。
1、诱导剂浓度对基因工程菌表达SthylO的影响
当初始菌体浓度OD
2、诱导初始菌体浓度对基因工程菌表达SthylO的影响
选择OD
3、诱导时间对基因工程菌表达SthylO的影响
当OD
4、诱导温度对基因工程菌表达SthylO的影响
当OD
综上,发酵生产透明质酸酶的最佳发酵条件为:当OD
(2)诱导结束后,将发酵液进行低温离心,去除上清液,将菌体用pH值6.0的PBS缓冲液洗涤2~3次,再溶入PBS缓冲液中形成菌体重悬液,利用超声破碎仪对菌体重悬液进行破碎,得到粗酶液;将粗酶液离心,收集上清用0.22μm滤膜进行抽滤,采用AKTA层析系统,利用镍柱亲和层析法进行纯化得到纯酶液;将纯酶液置于超滤管中进行浓缩,并除去金属离子。经SDS-PAGE电泳显示SthylO的分子量大小约为25kDa。随后将纯酶液进行收集,并至于-80℃保存,方便后续透明质酸酶SthylO的酶学性质分析。
通过考察最适反应温度及稳定性、最适反应pH及稳定性、金属离子对酶活力的影响,以及酶的底物特异性和动力学参数的研究,来进行酶学性质分析。
1、透明质酸酶SthylO最适反应温度及稳定性的测定
最适反应温度为:取200μL纯酶液与200μL的HA溶液(10g/L的HA标准品溶于pH值6.0的PBS缓冲溶液)分别在20、25、30、35、40、45、50、55、60、65、70℃条件下反应30min,使用DNS法测定酶活,每个温度设置3组平行。酶活定义为:在pH 6.0、40℃下,每分钟生成1μmolN乙酰氨基葡萄糖所需的酶量为一个酶活单位。温度稳定性测定为:提前将纯酶液于20、25、30、35、40、45、50、55、60、65、70℃条件下保温6h,再测定残余酶活。结果如图3A所示,20~40℃之间SthylO的相对活力随温度的升高而不断增加,在40℃达到最大值,当温度超过40℃时,其相对活力开始骤降,这是因为高温条件下会破坏酶的结构,使得蛋白质变性从而失去活力。在20~40℃的温度范围内,酶的相对活力值均在70%以上,说明SthylO具有较好的热稳定性,有利于酶的运输和存储,从而降低生产成本。
2、SthylO最适反应pH及稳定性的测定
最适反应pH测定为:将HA标品溶解在不同pH的PBS缓冲液中形成不同pH的HA溶液(10g/L),再取200μL纯酶液分别与pH为4.0、5.0、6.0、6.5、7.0、7.2、7.4、7.5、7.6、8.0、8.9的200μL HA溶液反应,40℃反应30min后测定酶活。酶活定义为:每分钟生成1μmol N乙酰氨基葡萄糖所需的酶量为一个酶活单位。pH稳定性的测定为:先将纯酶液分别加入不同pH的HA溶液中于4℃保温6h后测定残余酶活。结果如图3B所示,SthylO的相对活力在pH值4.0~6.0之间不断增加,并在pH值6.0达到最大值。当pH小于5.0和高于7.2时,SthylO的相对活力明显降低,这是因为在强酸和强碱的条件下,酶的空间结构和催化基团的解离状态受到影响,从而会影响酶和底物结合,进而影响产物的合成,故活力降低。pH值在5.0~7.2之间,SthylO的相对活力都在70%以上,说明其在该范围内相对稳定。
3、金属离子对SthylO活力的影响
金属离子对SthylO活力影响的测定为:配制浓度为0.1M的KCl、NaCl、CoCl
4、SthylO底物特异性的研究
底物特异性的测定为:用pH值为6.0的PBS缓冲液配制浓度为10g/L的不同底物溶液。所选取的底物为含有β-1,4糖苷键二糖单位组成的聚合物,包括HA、甘露聚糖、硫酸软骨素、硫酸软骨素(鸡)、肝素钠、结冷胶中的任意一种。将200μL分离纯化超滤后的纯酶液与200μL不同底物溶液于40℃,pH值6.0下反应30min后测定SthylO的酶活。酶活定义为:每分钟生成1μmol N乙酰氨基葡萄糖所需的酶量为一个酶活单位。结果如图4B所示,SthylO具有一定的特异性,只能显著降解HA,而不能降解同系的甘露聚糖、硫酸软骨素、硫酸软骨素(鸡)、肝素钠、结冷胶。Sthyl的高底物特异性也使得其具有较高度催化效能,能够高效地催化底物获得所需的不同分子量的HA。
5、SthylO动力学参数的研究
动力学参数的测定为:配制一系列浓度的HA底物溶液(1.25、2.5、3.75、6.25、8.75、12.5、15、18.75mg/mL),采用DNS法测定酶活,并根据标准曲线得到反应所消耗的底物浓度,从而计算反应速率(图5A),对米氏方程进行转换,作Lineweaver-Burk图。结果如图5B所示,获得拟合曲线方程y=2.0805x+0.3458,从而根据公式
实施例3:合成不同分子量透明质酸工程菌的构建
将SEQ ID NO.1、SEQ ID NO.2和SEQ ID NO.3所示的核苷酸序列分别单独扩增,经过重叠PCR后克隆到表达质粒pMA5-sthasA-cghasB(表达质粒pMA5-sthasA-cghasB的构建过程已公开在专利CN116004496A中)中得到重组质粒pMA5-sthasA-cghasB-P
(1)以质粒pHT01为模板,利用引物P
上述所用引物序列如表3所示;PCR扩增体系为:5×SF Buffer 10μL、dNTP Mix 2μL、Phanta HS Super-Fidelity DNA Polymerase 2μL、上游引物2μL、下游引物2μL、模板DNA2μL、ddH
①P
②SpyncM:95℃预变性3min,95℃变性15s;然后60℃退火15s,72℃延伸15s,循环30次;
③SthylO:95℃预变性3min,95℃变性15s;然后63℃退火15s,72℃延伸30s,循环30次。
利用胶回收试剂盒回收PCR扩增产物,进行重叠PCR反应。
重叠PCR所用上下游引物为:P
利用胶回收试剂盒回收重叠PCR扩增产物,与经过限制性内切酶Kpn I和Hind III双酶切后的质粒pMA5-sthasA-cghasB(表达质粒pMA5-sthasA-cghasB的构建过程已公开在专利CN116004496A中),使用一步克隆法在同源重组酶ExnaseⅡ作用下进行连接,得到重组质粒pMA5-sthasA-cghasB-P
(2)E.coli GM2163感受态制备
将保存在甘油管中的E.coli GM2163在LB固体培养基上进行划线,37℃过夜培养。挑取单菌落于5mL的LB培养基中,37℃培养12h,作为种子液。取6mL种子液接种到装有100mL的LB培养基中,37℃培养至OD
然后将重组质pMA5-sthasA-cghasB-P
表3扩增编码基因P
(3)解淀粉芽孢杆菌(B.amyloliquefaciens)CF感受态的制备
将保存在甘油管中的解淀粉芽孢杆菌(B.amyloliquefaciens)CF(解淀粉芽孢杆菌CF的构建及其感受态细胞的制备已公开在专利CN116004496A中)进行LB平板划线,37℃过夜培养。挑取单菌落于5mL感受态制备培养基中,37℃培养12h,作为种子液。取6mL种子液接种到装含有100mL的感受态培养基中,37℃培养震荡至OD
用75%酒精清洗电转杯(规格:2mm),在超净工作台中晾干,电转前于冰上预冷。向100μL感受态细胞中加入约100ng重组质粒,混匀后转移至预冷的电转杯中冰浴20~30min。电击转化条件为:电压3kV,电击时间4ms。电击结束后于室温下迅速加入1mL复苏培养基到电转杯中,混匀后将液体转移到2mL离心管中。复苏3h后将转化液离心,保留适当液体重悬菌体,随后将重悬液涂布到含有硫酸卡那霉素(25μg/mL)的LB琼脂平板上,37℃,静置培养12h。从平板上挑取表面水润有光泽的单菌落并用引物P-St-F/R进行PCR验证,验证成功的阳性菌株即为重组解淀粉芽孢杆菌B.amyloliquefaciens CF pMA5-sthasA-cghasB-P
其中,感受态制备培养基组成为:NaCl 10g/L、胰蛋白胨10g/L、酵母粉5g/L、山梨醇0.5M。电转缓洗涤液组成为:山梨醇0.5M、甘露醇0.5M、甘油15%;悬浮液组成为:山梨醇0.5M、甘露醇0.5M、甘油15%、聚乙二醇600 14%;复苏培养基组成为:NaCl 10g/L、胰蛋白胨10g/L、酵母粉5g/L、山梨醇0.5M、甘露醇0.38M;摇瓶发酵,其培养基组成为:蔗糖45g/L、酵母粉10g/L、MgSO
实施例4:重组解淀粉芽孢杆菌发酵合成不同分子量的透明质酸
(1)将实施例3得到的重组解淀粉芽孢杆菌于37℃活化后,挑取单菌落接种到种子培养基中,200rpm、37℃培养10h作为种子液。
(2)将步骤(1)获得的种子液以6%v/v的接种量分别接种于发酵培养基,在32℃、pH值7.0、200rpm条件下进行发酵,并添加适当的IPTG进行诱导优化。
(3)利用5L发酵罐进行扩大培养,并收集不同时间段的发酵产物进行液相分析。
其中,种子培养基组成为:蔗糖20g/L、酵母粉10g/L、牛肉膏10g/L、MgSO
通过考察诱导剂不同浓度、诱导剂不同添加时间下对透明质酸的影响,以及5L发酵扩大培养工艺来优化发酵合成不同分子质量透明质酸的发酵工艺。
1、诱导剂不同浓度对重组解淀粉芽孢杆菌发酵合成透明质酸的影响
选择0.1、0.3、0.5、0.7、1.0、1.2mM的IPTG分别诱导36h后,以不加IPTG组为对照。发酵结束后,将发酵液稀释5倍离心收集上清,并用0.22μm滤膜对进行过滤,然后利用高效液相色谱仪示差检测器对峰面积进监测,并结合标准曲线换算出透明质酸的产量。结果如图7A所示,当IPTG浓度为0.3mM时,透明质酸的产量最高为10.14g/L,但有肩峰存在,表明所添加的IPTG不能完全解除抑制;当IPTG浓度为0.5mM时,透明质酸的产量为10.03g/L,且峰型较为单一,故选择0.5mM的IPTG作为最佳的诱导剂浓度。
2、诱导剂不同添加时间对重组解淀粉芽孢杆菌发酵合成透明质酸的影响
在发酵开始0、6、12、24、36、48h后分别添加0.5mM的IPTG进行诱导。发酵72h后,将发酵液稀释5倍离心收集上清,并用0.22μm滤膜对进行过滤,然后利用高效液相色谱仪示差检测器对峰面积进监测,并结合标准曲线换算出透明质酸的产量。结果如图7B所示,当诱导剂IPTG的添加时间为发酵开始36h后,透明质酸的产量最高达到11.79g/L,而诱导剂添加时间过早或过晚都对重组解淀粉芽孢杆菌产透明质酸有不利影响,故选择诱导剂IPTG的最佳添加时间为发酵开始36h后。
3、5L发酵罐分批发酵透明质酸
将种子液以6%v/v的接种量接种于装有发酵培养基的发酵罐中,在32℃,pH 7.0,400~600rpm,通气量4vvm条件下进行5L发酵罐放大培养,当发酵开始36h后,添加0.5mM的IPTG进行诱导培养,并搜集诱导开始12h、24h、48h后的发酵液上清,经过滤除菌后,利用高效液相色谱仪示差检测器进行检测,并结合分子量标准曲线,对不同诱导时间段透明质酸的分子量大小进行初步判断。
其中,高效液相色谱仪示差检测方法为:将发酵液稀释5倍后,离心至去除细菌;上清用0.22μm滤膜过滤,保存在色谱进样瓶中以便采集。以0.1M的Na
结果如图8A所示,发酵开始后,细胞快速繁殖,菌体量也随着不断增加,但在发酵36h时增加幅度较小,且透明质酸的增加量也较为缓慢,这可能是因为在发酵过程中透明质酸黏性大分子的持续合成和分泌,使发酵液呈现黏稠的状态,不利于菌体对氧气的摄取。在36h添加诱导剂后,发酵溶氧问题得到解决,细胞代谢极为旺盛,透明质酸也在高效合成,最终在发酵96h时趋于稳定,此时透明质酸的产量为15.20g/L。随后对不同诱导时间段的发酵产物进行高效液相分析,并结合葡聚糖标准曲线
4、高、中、低分子量HA及HA寡糖的抗氧化活性分析
将不同诱导时间段的发酵产物高速离心去除菌体,然后将上清液体进行反复醇沉,经烘干后得到各自的粗多糖;将粗多糖分别进行复溶后,利用Sevag方法除去杂蛋白质,在4℃条件下透析两天除去小分子物质以及盐离子,每4h更换一次透析液(透析液为蒸馏水);最后将透析液高速离心,并取上清液进行冷冻干燥,备用。
1)DPPH自由基清除活性测定
分别配置高、中、低分子量HA及HA寡糖0.1、1、2、3、4、5g/L的HA溶液,无水乙醇配制0.1mM DPPH溶液于棕色瓶中保存备用。96孔板中分别加入100μL不同浓度的待测样品,与100μL DPPH溶液混匀,室温下,避光反应30min,然后测定其在517nm波长处的吸光度(A
2)羟自由基清除活性测定
分别配置高、中、低分子量HA及HA寡糖0.1、1、2、3、4、5g/L的HA溶液,配置9mM的FeSO
3)ABTS自由基清除活性测定
分别配置高、中、低分子量HA及及HA寡糖0.1、1、2、3、4、5g/L的HA溶液,配置7.4mM的ABTS溶液和2.6mM的过硫酸钾溶液。将ABTS和过硫酸钾溶液按1:1等体积混匀后,在室温条件下,于黑暗环境放置15h,然后再用蒸馏水将混合液稀释40倍得到ABTS工作液。96孔板中分别加入20μL不同浓度的待测样品,与200μL ABTS工作溶液混匀,室温下,反应20min,然后测定其在734nm波长处的吸光度(A
综上,本发明实现了一菌制备高、中、低不同分子质量的透明质酸及透明质酸寡糖,能够简化工业程序的复杂性,降低生产成本;且制备的HA寡糖在抗氧化分析中具有显著的清除效率,极具工业化应用价值。
本发明提供了一种新型透明质酸酶及其基因工程菌、构建方法与应用的思路及方法,具体实现该技术方案的方法和途径很多,以上所述仅是本发明的优选实施方式,应当指出,对于本技术领域的普通技术人员来说,在不脱离本发明原理的前提下,还可以做出若干改进和润饰,这些改进和润饰也应视为本发明的保护范围。本实施例中未明确的各组成部分均可用现有技术加以实现。