植物病原卵菌氧化固醇结合蛋白上PH结构域和START结构域的功能及其应用
文献发布时间:2024-04-18 19:58:21
技术领域
本发明涉及分子生物学和基因工程技术领域,具体涉及一种疫霉菌氧化固醇结合蛋白ORP1上PH结构域和START结构域的功能及结构,以及两种结构域在杀菌剂靶标中的应用。
背景技术
卵菌(Oomycetes)是一类与真菌相似的真核生物,由于两者营养汲取方式、菌丝形态类似且均能够通过有丝分裂产生孢子,在早期的分类系统中,卵菌一直被认为属于真菌。直到20世纪80代,化石证据及分子系统生物学数据表明卵菌与硅藻和褐藻的亲缘关系更近,因此微生物学家将其归为假菌界(Chromista)的成员(Cavalier-Smith,1998)。目前,研究人员已发现至少1800种卵菌,除了一些腐生卵菌外,大部分卵菌是植物、动物以及其他生物的病原菌,给人类造成了巨大的经济损失(Thines and Kamoun,2010)。植物病原卵菌主要包括霜霉(Downy mildews)、疫霉(Phytophthora)、腐霉(Pythium)和白锈病菌(Albugo)(Thines and Kamoun,2010)。2014年所公布的十大植物病原卵菌中包括6种疫霉属病原菌:致病疫霉(P.infestans)、橡树疫霉(P.ramorum)、大豆疫霉(P.sojae)、辣椒疫霉(P.capsici)、樟疫霉(P.cinnamomi)和寄生疫霉(P.parasitica)(Kamoun et al.,2015)。
由于缺少抗病品种或抗性易丧失等问题,化学防治仍是植物卵菌病害控制的主要措施。以氟噻唑吡乙酮和Fluoxapiprolin为代表的OSBPI类杀菌剂是迄今为止生物活性最高的一类卵菌抑制剂,也是21世纪最有代表性并有望形成系列产品的新型杀菌剂。前期研究结果表明,氟噻唑吡乙酮和Fluoxapiprolin通过靶向卵菌中ORP1蛋白上的ORD结构域而发挥杀菌作用,且ORD上14种氨基酸点突变可引起氟噻唑吡乙酮和Fluoxapiprolin的高水平的抗性(Miao et al.,2021;Li etal.,2022),该类药剂在推广应用2年后已陆续有田间氟噻唑吡乙酮抗性的监测报道(FRAC),因此开发新型杀菌剂用于已有OSBPI类杀菌剂抗药性治理,延长OSBPI类杀菌剂使用寿命已迫在眉睫。
氧化固醇结合蛋白ORPs存在相似的特点:(1)在羧基端均具有一个主要功能结构域—ORD结构域。(2)长链ORPs羧基端具有PH结构域,少部分ORPs还存在锚蛋白重复序列及高尔基体动态结构域等,主要介导ORPs与其互作蛋白或配体的相互作用(Raychaudhuriand Prinz,2010;Levine and Olkkonen,2004)。Raychaudhuri和Prinz(2010)总结了人类及酵母中氧化固醇结合蛋白ORPs的4种功能机制:(1)在细胞质膜之间(主要是在MCS)提取及运送脂类物质;(2)通过添加及移除质膜特定位置的脂类分子,建立质膜上的脂类分子的瞬时分布变化,通过这种方式ORPs对于膜的弯曲或由膜上特定脂类位置的聚集或分散所驱动的信号途径起作用;(3)ORPs作为脂类分子的感应分子,通过与脂类分子的结合或分开调控ORPs与其它互作蛋白的相互作用,进而影响相关的信号途径或调控蛋白与质膜的相互作用;(4)ORPs能够调节其他脂类结合蛋白与质膜的结合,既可以将膜上的脂类分子传递给其他的脂类结合蛋白,也可以阻止其他脂类分子与质膜的结合。四种机制不是相互独立的,一些ORPs能够同时利用多种功能机制。基于ORPs的重要性,靶向ORPs的药物分子不断在医药和农药中得到开发。
卵菌中ORP1蛋白相对于真菌、植物和动物中的氧化固醇结合蛋白相关蛋白ORPs同源性均低于30%,与此同时,卵菌的ORP1均包括了3个保守结构域PH、START和ORD结构域,相比于其他真核生物的长链ORPs多了一个START结构域。目前,已有的OSBPI类杀菌剂均是仅靶向结合ORD结构域。
发明内容
本发明要解决的技术问题是研究卵菌(辣椒疫霉、大豆疫霉、致病疫霉、寄生疫霉、荔枝霜疫霉、黄瓜霜霉、葡萄霜霉、瓜果腐霉等重要植物病原卵菌)关键生长发育蛋白ORP1上PH结构域和/或START结构域的结构和功能,明确卵菌中ORP1蛋白中PH结构域和START结构域缺失会导致植物病原卵菌死亡,表明卵菌ORP1上的PH结构域和START结构域是潜在的植物病原卵菌抑制剂的分子靶标。进而发掘ORP1上PH结构域和/或START结构域在防治卵菌病害防治中的应用,即可以通过设计靶向抑制两个结构域功能的农药分子,从而抑制卵菌ORP1生物学功能,控制植物病原卵菌生长或发育的某一环节,进而达到控制病害发生与传播。这些研究工作的开展对控制植物病原卵菌具有重要的理论和实践价值,为开发以干扰病原卵菌生长发育为目标的新型高效、低毒的杀菌剂具有重要的意义。
基于此,本发明提供植物病原卵菌氧化固醇结合蛋白ORP1上PH结构域和/或START结构域的新应用。
所述植物病原卵菌为疫霉、腐霉和霜霉菌中的一种或两种以上任意组合;优选的,所述植物病原卵菌为辣椒疫霉、大豆疫霉、致病疫霉、寄生疫霉、荔枝霜疫霉、黄瓜霜霉、葡萄霜霉、瓜果腐霉中的一种或两种以上任意组合。
所述植物病原卵菌PH结构域的氨基酸序列为与SEQ ID NO.1所示的氨基酸序列相似度大于90%的氨基酸序列;START结构域的氨基酸序列为SEQ ID NO.2所示的氨基酸序列相似度大于90%的氨基酸序列;其中,辣椒疫霉ORP1上PH结构域的氨基酸序列为SEQ IDNO.1所示的氨基酸序列;START结构域的氨基酸序列为SEQ ID NO.2所示的氨基酸序列。
其中,本发明的目的之一是提供植物病原卵菌氧化固醇结合蛋白ORP1上PH结构域和/或START结构域作为药剂分子靶标在卵菌病害防治中的应用。
辣椒疫霉、大豆疫霉、寄生疫霉、荔枝霜疫霉、瓜果腐霉等重要植物病原卵菌中的ORP1上的PH结构域和/或START结构域缺失能够引起病原菌死亡。
辣椒疫霉、大豆疫霉、寄生疫霉、荔枝霜疫霉、瓜果腐霉等重要植物病原卵菌中的ORP1上的PH结构域和/或START结构域对于ORP1在辣椒疫霉菌体中的定位尤为关键。
本发明还提供植物病原卵菌氧化固醇结合蛋白ORP1上PH结构域和/或START结构域杀灭辣椒疫霉中的应用。
本发明还提供了植物病原卵菌氧化固醇结合蛋白ORP1上PH结构域和/或START结构域在作为筛选杀菌剂的靶标中的应用。
本发明还提供了辣椒疫霉ORP1上PH结构域的晶体结构和AlphaFold2预测及校验的PcORP1-START结构域的结构,从而为后期基于开展靶向卵菌ORP1上PH结构域和START结构域的杀菌剂分子设计相关的研究提供了结构指导。因此,植物病原卵菌中氧化固醇结合蛋白ORP1上PH结构域和/或START结构域以及相似度>90%的卵菌PH结构域和/或START结构域在杀菌剂分子设计中的应用,也属于本发明的保护范围。
本发明的有益效果是:本发明提供了辣椒疫霉氧化固醇结合蛋白ORP1上PH结构域的晶体结构以及通过AlphaFold2预测及校验的PcORP1-START结构域的结构,植物病原卵菌ORP1上PH结构域和START结构域对于ORP1的亚细胞定位和功能具有关键作用,以卵菌ORP1上PH结构域或START结构域为作用靶标,开发的杀菌剂对控制辣椒疫病等植物病原卵菌病害的发生与流行具有重要的实践意义。
附图说明
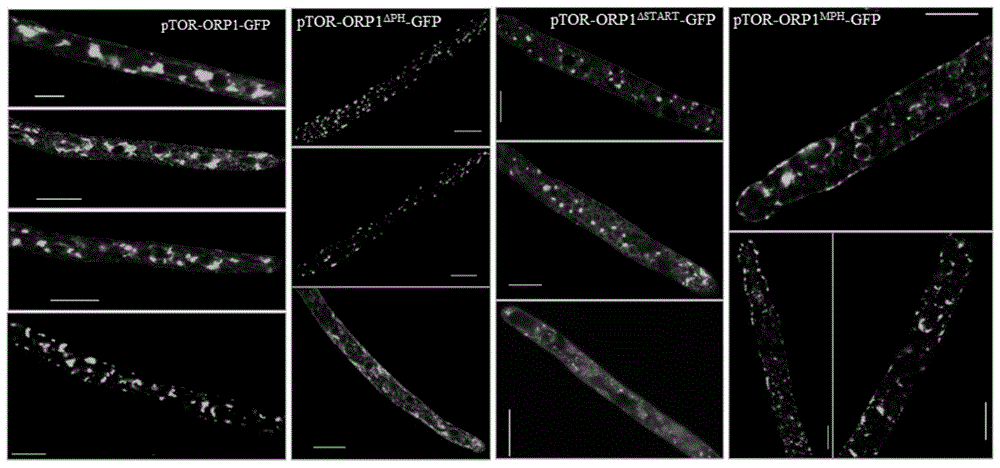
图1辣椒疫霉ORP1及其突变体在菌丝体定位
图2不同卵菌ORP1蛋白中PH结构域的相似度比对分析
图3不同卵菌ORP1蛋白中START结构域的相似度比对分析
图4是根据发明的辣椒疫霉ORP1上PH结构域的空间结构。(A、D和G)PH-SO4的蛋白晶体、衍射图样和
图5AlphaFold2预测的PcORP1-START结构域的结构
具体实施方式
下面参照附图,结合具体的实施例对本发明作进步一步地说明,以更好地理解本发明。实施例中未注明具体技术或条件者,按照本领域内的文献所描述的技术或条件或者按照产品说明书进行。所用试剂或仪器未注明生产厂商者,均为可以通过市购获得的常规产品。
下面结合附图,对本发明的具体实施方式进行详细描述,但应当理解本发明的保护范围并不够具体实施方式的限制。
下面实施例中所使用的实验方法如无特殊说明,均为常规方法。
下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径可购得。
为了探讨辣椒疫霉氧化固醇结合蛋白ORP1上PH结构域和START结构域是否可作为药剂靶标而开展抑制剂方面的研究,本发明明确了ORP1上PH结构域和START结构域对辣椒疫霉及辣椒疫霉ORP1蛋白的细胞定位的重要作用,同时解析了辣椒疫霉ORP1上PH结构域的晶体结构。
实施例1、辣椒疫霉ORP1全长及截断体的亚细胞定位:
pTOR-ORP1-GFP的构建:以辣椒疫霉LT1534的cDNA为模板,设计表1带有酶切位点(Apa1和Spe1)的特异性引物F1/R1扩增PsORP1的全长;pYF2.2-GFP质粒(Fang and Tyler,2015)为模板,利用表1所示F2/R2引物扩增获得的GFP片段;利用诺唯赞的ClonExpressUltra One Step Cloning Kit将上述两个片段插入克隆载体pTOR上,连接产物转入大肠杆菌DH5α感受态细胞中,37℃过夜培养后,利用通用引物pTOR-F(序列:5’-AAACCACCTGTTTGTGGGGTTGTG-3’)和pTOR-R(序列:5’-ACGGCCTTCTTTTGAAAACAATC-3’)扩增,将有目的条带大小的转化子送入公司测序。正确转化子提质粒备用。
TOR-ORP1Δ
采用PEG-CaCl
表1 ORP1及其突变体融合GFP表达载体构建引物序列
实施例2、辣椒疫霉,大豆疫霉ORP1杂合敲除转化子自交后代分析
前期通过CRISPR/Cas9基因敲除试验,尝试敲除辣椒疫霉、大豆疫霉、寄生疫霉、荔枝霜疫霉、瓜果腐霉等重要植物病原卵菌中的ORP1全长、PH结构域敲除、START结构域,但均为获得纯合敲除转化子,仅仅获得杂合敲除转化子,分析推测ORP1全长、PH结构域敲除、START结构域敲除后破坏ORP1生物学功能,可能导致病原菌死亡。其中,大豆疫霉,辣椒疫霉共验证发生同源重组交换的ORP1全长、PH结构域敲除、START结构域敲除转化子近2000株,但均为杂合敲除转化子。进一步以杂合敲除转化子为亲本,通过自交方法获得F1代,按照孟德尔的遗传规律,F1代中应该包括一定比例的纯合敲除转化子。如果F1中没有纯合敲除转化子出现,即可以证明目标基因不能被敲除,一旦ORP1的纯合缺失、辣椒疫霉ORP1上的PH结构域或START结构域的纯合缺失会即可导致病原菌死亡。
本实施例中,进一步分别利用获得的辣椒疫霉ORP1杂合敲除转化子、PH结构域敲除、START结构域敲除转化子,通过聚碳酸酯膜间隔配对自交试验产生卵孢子,1)25℃黑暗培养45天;2)取菌丝琼脂块,粉碎机粉碎,转移至50mL离心管;3)4000rpm离心5min,倒去上层培养基,加无菌水30mL,650×g离心5min,倒去上清;4)经40μm细胞过滤器过滤,获得卵孢子;5)使用0.25%的高锰酸钾处理20min去除残余菌丝,刺激卵孢子萌发,650×g离心5min,弃去上清;6)加入20mL无菌水混匀,400×g离心5min,弃去上清;加入5mL无菌水悬浮卵孢子,涂布于半选择性S+L培养基上,黑光灯照射2d;7)在无菌操作台中,使用显微镜挑取单个萌发的卵孢子于V8板上,用于后续测序分析。
共获得510株辣椒疫霉ORP1杂合敲除转化子自交后代后代菌株,对510株辣椒疫霉ORP1杂合敲除转化子自交后代进行测序分析,得到161株野生型菌株,349株ORP1杂合敲除转化子,没有获得ORP1纯合敲除转化子;获得486株辣椒疫霉ORP1-PH结构域杂合敲除转化子自交后代,得到134株野生型菌株,352株ORP1-PH结构域杂合敲除转化子,没有获得ORP1-PH结构域纯合敲除转化子;获得686株辣椒疫霉ORP1-START结构域杂合敲除转化子自交后代,得到196株野生型菌株,490株ORP1-PH结构域杂合敲除转化子,没有获得ORP1-START结构域纯合敲除转化子;近一步说明辣椒疫霉ORP1的纯合缺失、辣椒疫霉ORP1上的PH结构域或START结构域的纯合缺失会导致辣椒疫霉死亡。
实施例3、ORP1上PH结构域的结构解析及药效团筛选方法
序列相似度分析结果显示,不同卵菌ORP1中PH结构域的氨基酸序列相似度>90%(图2),说明PH结构域在不同卵菌ORP1中保守存在。
通过晶体生长筛选与优化,最终仅切除6His-MBP标签的PH16-134在3种不同条件下获得的晶体能够用于X-ray衍射收集数据。PH-SO
PH
基于PcORP1-PHD-SO4对接的最优构象PcORP1-PHD-SO4-B进行药效特征确定——采用MOE 2019软件中的Pharmacophore模块构建基于受体的药效团模型,建立了含8个药效团的初始模型。对specs和chemdiv数据库进行虚拟筛选,经过类农药五规则[MW(Weight)<435Da,Clogp(logp)<6,0 实例4:PcORP1中START结构域的结构预测与分析 序列相似度分析结果显示,不同卵菌ORP1中START结构域的氨基酸序列相似度>90%(图3),说明START结构域在不同卵菌ORP1中保守存在。 使用AlphaFold2在线服务器(https://colab.research.google.com/github/sokrypton/ColabFold/blob/main/AlphaF old2.ipynb)进行START结构域结构特征预测,结果如图5所示。pLDDT值基本在70以上。将AlphaFold2预测的PcORP1-START结构与PDB数据库中检索到的START结构域进行比对,结果表明,PcORP1-START与其他物种的START结构域的序列同源性介于12.4%~19.7%,三维结构RMSD介于1.439~5.524。分析发现PcORP1-START结构域与3H3Q、2Z9Z、1LN1、6CAY叠合后,显示其配体结合腔紧邻两个跨膜螺旋上方,这暗示PcORP1-START结构域可能通过近膜端获取并运输脂质;PcORP1-START结构域与3H3Q、2Z9Z具有更接近的配体结合口袋,基于叠合构象,使用MOE进行初步分子对接测试后,推测PcORP1-START结构域的底物有可能为神经酰胺(ceramide)或二酰基甘油(diacylglycerol)类似物。 以上对本发明的具体实施例进行了详细描述,但其只是作为范例,本发明并不限制于以上描述的具体实施例。对于本领域技术人员而言,任何对本发明进行的等同修改和替代也都在本发明的范畴之中。因此,在不脱离本发明的精神和范围下所作的均等变换和修改,都应涵盖在本发明的范围内。
- 将由A蛋白或A蛋白的A至E结构域中的至少一个结构域所组成的蛋白质结合到基材上的方法
- 靶向结合CDP上Sox2-CDP蛋白复合物中结合结构域的多肽及其合成方法和应用