头孢菌素C酰化酶突变体及其在头孢菌素母核合成中的应用
文献发布时间:2024-04-18 19:58:53
技术领域
本发明属于生物催化技术领域,涉及一种利用头孢菌素C酰化酶催化合成头孢菌素母核的方法。
背景技术
头孢类抗生素是多年来应用最为广泛的β-内酰胺类抗生素之一,其关键母核为7-氨基头孢烷酸(7-aminocephalosporanic acid,7-ACA)及其相关的衍生物。此类抗生素占据了全球抗生素市场的40%左右,具有重要的医用价值。
早期7-ACA的工业制备采用化学裂解法裂解头孢菌素C(Cephalosporin C,CPC),随后除去分子侧链得到7-ACA。其反应条件较为苛刻,且反应过程必须在超低温条件下进行。化学法涉及大量的有毒有害有机溶剂,工艺复杂且能耗很高,带来环境污染等问题。因此,近些年酶催化方法由于能耗低且对环境友好逐渐成为工业生产应用的新方向。早期,工业化生产采用两步酶法进行催化,主要利用D-氨基酸氧化酶(D-amino acid oxidase,DAAO)和戊二基-7-氨基头孢烷酸(glutaryl-7-aminocephalosporanicacid,GL-7-ACA)酰化酶。首先通过DAAO将CPC转化为GL-7-ACA;其次,通过GL-7-ACA酰化酶将GL-7-ACA转变为7-ACA。但是,两步酶法催化时,第一步产物过氧化氢与DAO底物或产物反应生成大量副产物,且其工艺过程长、难于控制且成本较高,因此目前更多地采用一步酶法催化CPC裂解反应。
一步酶法利用CPC酰化酶催化CPC酰胺键的水解反应。自20世纪80年代以来,人们从自然界发现了可分泌CPC酰化酶的菌株,如:假单胞菌SE83来源的acyII基因;假单胞菌N176来源的CPC酰化酶基因;假单胞菌V22来源的CPC酰化酶基因;以及假单胞菌P130来源的CPC酰化酶基因等。然而,目前头孢菌素C酰化酶基因水解活性亟待提高,与GL-7-ACA相比,其水解酶活力仅为2~4%。且新的头孢菌素C酰化酶仍有待挖掘,以开发出更具有商业应用潜力、且固定化后重复应用次数更多的酰化酶。
发明内容
本发明所要解决的技术问题是如何利用生物催化方法高效制备头孢菌素母核7-ACA。
为解决以上技术问题,本发明首先提供了一种酰化酶突变体,其中野生型的酰化酶的氨基酸序列如SEQ ID NO:1所示,其来源于Bosea sp.OK403,所述头孢菌素C酰化酶突变体是在所述野生型序列上进行在第L159、G160、L161、L162、I166、W167、H249、P259、V262、H294、F295、、V305F309、E308、A419、N479、H496、P603、Y704位进行突变得到的蛋白质。
所述突变头孢菌素C酰化酶进行的选自如下六种的任一种、任两种、任三种、任四种、任五种、任六种、任七种、任八种、任九种、任十种、任十一种、任十二种、任十三种、任十四种、任十五种、任十六种、任十七种、任十八种、或全部突变得到的蛋白质:
(1)L159为丙氨酸,赖氨酸、丝氨酸、半胱氨酸、谷氨酰胺、苏氨酸所取代;
(2)G160为苯丙氨酸、天冬酰胺、丝氨酸所取代;
(3)L161为苯丙氨酸、甲硫氨酸、丙氨酸、赖氨酸、丝氨酸、半胱氨酸所取代;
(4)L162为丙氨酸、甲硫氨酸、组氨酸所替代;
(5)I166为色氨酸所替代;
(6)W167为丙氨酸、丝氨酸、天冬酰胺所替代;
(7)H249为丙氨酸、丝氨酸所代替;
(8)P259为缬氨酸、丙氨酸、赖氨酸、天冬氨酸所替代;
(9)V262为亮氨酸、异亮氨酸、丝氨酸所替代;
(10)H294为丙氨酸、丝氨酸所代替;
(11)F295为缬氨酸、天冬酰胺、亮氨酸所替代;
(12)V305为精氨酸、赖氨酸、丝氨酸所替代;
(13)F309为天冬氨酸、谷氨酰胺所替代;
(14)E318为苏氨酸所替代;
(15)A419为缬氨酸所替代;
(16)N479为亮氨酸所替代;
(17)H496为苯丙氨酸所替代;
(18)P603为谷氨酰胺、天冬酰胺所取代;或
(19)Y704为丝氨酸、半胱氨酸所取代。
另外优选地,所述突变体存在下述组合的氨基酸替代:
L159A/E318T、L159Q/A419V、L159S/A419V、L159S/G160S、L159Q/G160S、L159S/P603Q、L159Q/P603Q、L159S/G160S/P603Q、L159Q/G160S/P603Q、L159S/A419V/G160S、L159S/A419V/P603Q、L159S/A419V/G160S/P603Q、G160S/P603Q、L161K/H249A、L161K/H249S、L161K/V305K、V262S/V305S、L159T/V262S、H294S/V305K或H294S/E318T。
本发明相应提供上述突变头孢菌素C酰化酶的编码核酸分子,其表达盒,含有所述编码基因或其表达盒的重组载体、含有所述基因或所述表达盒或所述重组载体的重组微生物。
其中,所述核酸分子可以是DNA,如cDNA、基因组DNA或重组DNA;所述核酸分子也可以是RNA,如mRNA或hnRNA等。
所述的核酸分子的表达盒是指能够在宿主细胞中表达所述突变头孢菌素C酰化酶的DNA,该DNA不但可包括启动突变头孢菌素C酰化酶编码基因转录的启动子,还可包括突变头孢菌素C酰化酶转录的终止子。更进一步的,所述表达盒还可包括增强子序列。
所述的核酸分子的重组载体可为携带有所述突变头孢菌素C酰化酶编码基因的细菌质粒(如在细菌中表达的基于T7启动子的表达载体,具体如pET-28a等)、噬菌体、酵母质粒(如YEp系列载体等)或逆转录病毒包装质粒。
所述重组载体为将所述核酸分子插入表达载体中得到的载体。
所述的含有编码所述突变头孢菌素C酰化酶的核酸分子的重组微生物可为携带有头孢菌素或所述头孢菌素突变蛋白质编码基因的酵母、细菌、藻或真菌,如大肠杆菌等。
本发明还提供了一种制备头孢菌素母核的方法,具体包括以化合物I为底物,利用如SEQ ID NO:1所示头孢菌素C酰化酶或所述的头孢菌素突变体的催化剂相接触进行催化反应,得到化合物II:
其中R为乙酰氧基、羟基、氢。
优选地,本发明的酰化酶采用固定化酶的方式,例如以环氧树脂,氨基树脂,大孔吸附树脂为载体,与酰化酶进行固定化,获得具有高转化活性的固定化酶。
上述方法中,所述重组细胞可通过向生物细胞中导入能表达所述突变头孢菌素C酰化酶的重组载体获得。
所述生物细胞可为微生物。所述微生物可为大肠杆菌,也可以是其他菌。在本发明的一个实施例中,所述微生物为大肠杆菌BL21(DE3)。
所述反应的温度可为10-40℃,具体可为20℃;所述反应的pH 6.5~9.5的磷酸盐缓冲液中进行的,具体为8.0。一般可为0.5-24小时,具体可为1小时。所述重组细胞的裂解产物可由裂解所述重组细胞得到。
同时,本发明也提供一种来源于Bosea sp.的头孢菌素C酰化酶在催化头孢菌素C或去乙酰氧基头孢菌素C水解或酰化反应中的应用,其中所述头孢菌素C酰化酶的氨基酸序列如SEQ ID NO:1所示。
本发明首先挖掘出了来源于Bosea sp.OK403的头孢菌素C酰化酶可用于催化头孢菌素合成头孢菌素母核。更进一步地,本发明半理性的设计,构建突变体库,获得了效果大为改善的突变体,所得突变体可以催化头孢菌素衍生物产生头孢菌素母核,在生物催化制备头孢菌素母核中具有较好的工业应用前景。
附图说明
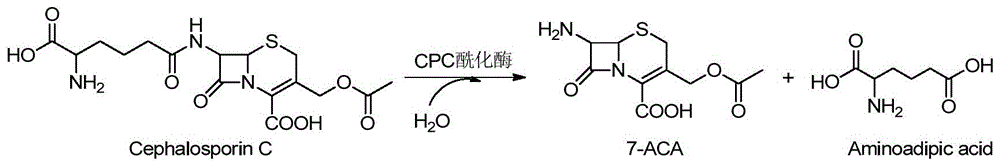
图1是本发明中酰化酶催化头孢菌素C水解反应路线图。
图2是本发明中7-ACA和CPC含量的标准曲线图。
图3是本发明中7-ACA和CPC的HPLC检测结果。
图4是本发明中CPC酰化酶E744用于底物饱和突变体库构建相关残基位点。
具体实施方式
下述实施例中所使用的实验方法如无特殊说明,均为常规方法。
下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。
实施例1酰化酶序列挖掘及酶活表征
1.1酰化酶序列挖掘
经NCBI检索,获取了来源于Bosea sp.OK403酰化酶序列SEQ ID NO.1(命名为E744)。
1.2.头孢菌素C酰化酶的诱导表达与纯化
将含有SEQ ID NO.1氨基酸序列的核酸序列转化到大肠杆菌BL21(DE3)菌液均匀的涂布在卡那霉素抗性(浓度为50μg/mL)的LB平板上,37℃恒温条件下培养14h后长出单菌落,即为含有头孢菌素C酰化酶核酸序列的转化子。
将上述转化子转接到含5mL的LB培养基(卡那霉素浓度为50μg/mL)的试管中,37℃、220rpm振荡培养过夜,即为种子液;然后按1%的接种量接种到100mL的TB培养基(卡那霉素浓度为50μg/mL)中,37℃,220rpm振荡培养3h,当菌液的OD
1.3酰化酶蛋白纯化
使用Ni
通过Bradford法对纯化后得到的目的蛋白浓度进行测定。使用SDS-PAGE凝胶电泳对目的蛋白进行分析。
1.4.头孢菌素C酰化酶催化CPC生成7-ACA
反应体系(1mL):40mM CPC,0.1mg酰化酶,补加磷酸缓冲液至1mL。反应条件:37℃1000rpm30 min,取100μL样品,加入9倍体积的甲醇终止并稀释反应体系,HPLC检测酶活。通过HPLC分析CPC转化7-ACA产物。
检测条件:色谱柱Agilent ZORBAX SB-C18(4.6*150mm 4um);流动相配为96%磷酸盐缓冲液(pH7.0,50mM)添加4%甲醇(v/v);流速0.8mL/min;检测波长为254nm,柱温箱恒定为25℃。
酶活定义:每分钟催化CPC产生1微摩尔(μmol)7-ACA所需要的酶量定义为1U。
表1酰化酶活性测试
实施例2构建酰化酶突变体
2.1单点饱和突变
以酰化酶E744氨基酸为模板,SWISS-MODEL进行建模,建模后蛋白与底物分子CPC建立置入模型,通过对接分析其作用位点,根据结果,在底物分子的6A范围内,酰化酶中位点L159、L161、L162、I166、W167、P259、V262、H294、F295、V305、F309、I413、P603严重影响CPC侧链的结合,影响底物在口袋的活动空间。在底物分子的6A范围内,位点G160、M163、G164、H260、R261、Y269、T306、H307、H313、H415、N479、H496、Y704对结构区域的氢键的形成,稳定底物结合口袋具有重要的作用。因此选择以上氨基酸进行单点饱和突变。
用所选的相应引物进行单点饱和突变,设计相应的引物如表2所示。
表2引物序列表
具体构建方法如下:
1、第一轮PCR
以质粒SEQ ID NO.1为模板,将表2中引物L159-F1、L159-F2、L159-F3、L159-F4按12:6:1:1比例混合,取2μL上述的混合液作为上游引物,取表4中KU-R1作为下游引物,扩增基因片段L1。位点G160、L161、L162、M163、G164、I166、W167扩增方法与上述基因片段L1相同。位点P259、H260、R261、V262、Y269以KU-R2作为下游引物,H294、F295、V305、T306、H307、F309、H313以KU-R3作为下游引物,I413、H415以KU-R4作为下游引物,N479、H496以KU-R5作为下游引物,P603以KU-R6作为下游引物,Y704以KU-R7作为下游引物进行基因片段扩增。
第一次轮PCR程序:94℃保温2min,(98℃保温15s,55℃保温30s,72℃保温30s)×28个循环,72℃保温5min。
2.第二轮PCR
分别取2μL第一轮PCR产物L1,L2,L3,L4等为引物,以质粒SEQ ID NO.1为模板,进行第二轮PCR,构建L159、G160、L161、L162、M163、G164、I166、W167、P259、H260、R261、V262、Y269、H294、F295、V305、T306、H307、F309、H313、I413、H415、N479、H496、P603、Y704单位点饱和突变库。第二轮PCR程序:94℃保温2min,(98℃保温15s,60℃保温30s,72℃保温5min)×28个循环,72℃保温10min。
3.基因突变体工程菌株构建
将获得的第二轮PCR产物进行如下操作:向第二轮PCR的产物中加入1μL的Dpn I用于消化质粒模板,37℃恒温条件下处理3h。取2μL酶消化后的第二轮PCR产物并经电转化进大肠杆菌BL21(DE3)中,将电转后的大肠杆菌BL21(DE3)菌液均匀的涂布在卡那霉素抗性(浓度为50μg/mL)的LB平板上,37℃恒温条件下培养14h后长出单菌落,即为酰化酶SEQ IDNO.1的基因突变体工程菌株。
2.2酰化酶突变体库高通量筛选及活力测定
将突变体库中的单菌落分别接种于96深孔板中,添加300μL(含卡那霉素50μg/mL)的LB培养基中,37℃培养8-12h,同时设置阴性对照组(pET-28a的空载体),然后将200μL种子液接种于的800μL TB培养基(含IPTG 0.1mM和卡那霉素50μg/mL)中。25℃表达16h,离心收集细胞菌体,置于-80℃冷冻1h,室温融化30min。加入200μL(0.1M pH8.0,含1mg/mL溶菌酶)的磷酸钾缓冲液,重悬菌体,37℃孵育1h。向细胞破碎液中加入200μL底物CPC(20mM),37℃的条件下反应4-6h。取40μL反应液至新的96孔板中,加入200μL终止反应液(20%冰醋酸:0.05M NaOH=2:1),离心10min(4℃,5000rpm)。取200μL上清,加入40μL显色剂(0.5%p-DAB溶于甲醇),室温反应10min后,检测415nm下的吸光度。
表3.随机突变体菌株相对活力测试
实施例3构建酰化酶组合突变体
基于实施例2筛选到活性的高突变体,构建如下的突变体:L159A/E318T、L159Q/A419V、L159S/A419V、L161K/H294A、L161K/H294S、L161K/V305S、V262L/V305S、L159T/V262L、H294S/V305S、H294S/E318T、L159Q/H294S/A419V,·为构建突变体设计引物,见表4。
表4.定点突变的引物
具体构建方法如下:
1)PCR
以质粒L159A为模板,取表4中E318T-F和E318T-R作为上、下游引物,扩增突变体L159A/E318T;以质粒L159T为模板,取表4中V262L-F和V262L-R作为上、下游引物,扩增突变体L159T/V262L;分别以质粒L159Q、L159S为模板,分别取表4中A419V-F/A419V-R、G160S-F/G160S-R、P603Q-F/P603Q-R作为上、下游引物,扩增突变体L159Q/A419V、L159S/A419V、L159Q/P603Q、L159S/P603Q、L159Q/G160S、L159S/G160S;以质粒G160S为模板,取表4中P603Q-F/P603Q-R作为上、下游引物,扩增突变体G160S/P603Q;以质粒L161K为模板,分别取表4中H294A-F和H294A-R、H294S-F和H294S-R作为上、下游引物,扩增突变体L161K/H294A、L161K/H294S和L161K/V305S;以质粒V262L为模板,取表4中V305S-F和V305S-R作为上、下游引物,扩增突变体V262L/V305S;以质粒H294S为模板,分别取表4中V305S-F和V305S-R、E318T-F和E318T-R作为上、下游引物,扩增突变体H294S/V305S和H294S/E318T以质粒L159S/A419V为模板,分别取表4中G160S-F/G160S-R、P603Q-F/P603Q-R作为上、下游引物,扩增突变体L159S/A419V/G160S、L159S/A419V/P603Q;以质粒L159S/A419V/G160S为模板,取表4中P603Q-F/P603Q-R作为上、下游引物,扩增突变体L159S/A419V/G160S/P603Q。
PCR程序:94℃保温2min,(98℃保温15s,55℃保温30s,72℃保温5min)×28个循环,72℃保温10min。
2)基因突变体工程菌株构建
将获得的PCR产物进行如下操作:向PCR的产物中加入1μL的Dpn I用于消化质粒模板,37℃恒温条件下处理3h。取2μL酶消化后的PCR产物并经电转化进大肠杆菌BL21(DE3)中,将电转后的大肠杆菌BL21(DE3)菌液均匀的涂布在卡那霉素抗性(浓度为50μg/mL)的LB平板上,37℃恒温条件下培养14h后长出单菌落,即为酰化酶SEQ ID NO.1的基因突变体菌株。
实施例4头孢菌素C酰化酶突变体全细胞催化7-ACA合成
4.1细胞培养
分别挑取实施例3中所构建的突变体的单克隆转接到含5mL的LB培养基(卡那霉素浓度为50μg/mL)的试管中,37℃、220rpm振荡培养过夜,即为种子液;按1%的接种量接种到100mL的TB培养基(卡那霉素浓度为50μg/mL)中,37℃、220rpm振荡培养3h,当菌液的OD
4.2全细胞反应
通过HPLC分析CPC转化产物,从而测定4.1中获得的全细胞的转化率的测定。反应为1mL体系,4.1中所得全细胞取800μL用于反应,添加底物CPC(40mM),37℃,1000rpm,反应30min。取100μL样品,使用甲醇稀释十倍后用HPLC测定转化率。
HPLC检测条件:所选用色谱柱为Agilent ZORBAX SB-C18(4.6*150mm 4um),流动相配比为:96%磷酸盐缓冲液(pH7.0,50mM),4%甲醇(v/v),流速0.8mL/min,检测波长为254nm,柱温箱恒定为25℃。
表5组合突变体全细胞对CPC的转化率
4.3粗酶液催化CPC合成7-ACA
按照实施例1的方式制备粗酶液,对获得突变体的粗酶液进行转化率验证。
反应体系(1mL):底物CPC(40mM),0.1ml酰化酶粗酶液,补加磷酸缓冲液至1mL。反应条件:37℃,1000rpm,30min,取100μL样品,使用甲醇稀释十倍后用HPLC测定转化率。
检测条件:色谱柱Agilent ZORBAX SB-C18(4.6*150mm 4um);流动相配为96%磷酸盐缓冲液(pH7.0,50mM)添加4%甲醇(v/v);流速0.8mL/min;检测波长为254nm,柱温箱恒定为25℃。
表6组合突变体粗酶对CPC的转化率
实施例5头孢菌素C酰化酶突变体全细胞催化去乙酰氧头孢菌素C合成7-ADCA
去乙酰氧头孢菌素C(Deacetoxycephalosporin C,DAOC)是CPC的类似物,其结构差别在于母核六元环侧链取代基,其中,DAOC为氢(-H),而CPC为乙酰氧基(-OCOCH3)。
通过HPLC分析DAOC转化产物,获得全细胞转化率。其反应为1mL体系,取800μL 3.1所得全细胞用于反应,加入底物DAOC(1mg/mL),37℃,1000rpm,反应30min。取100μL样品,使用乙腈稀释十倍后用HPLC测定转化率。
液相检测条件:色谱柱Agilent ZORBAX SB-C18(4.6*150mm 4um);流动相配为95%甲酸铵,5%乙腈,流速1ml/min,检测波长254nm,柱温箱恒定为25℃。
表7头孢菌素C酰化酶突变体对DAOC的转化率
实施例6头孢菌素C酰化酶突变体全细胞催化DAC合成去乙酰基7-ACA
去乙酰头孢菌素C(Deacetylcephalosporin C,DAC)是CPC的类似物。CPC与DAC、DAOC区别仅为母核六元环的侧链取代基,其中,DAC为羟基(-OH),而CPC为乙酰氧基(-OCOCH3)。
通过HPLC分析DAC转化产物,从而测定4.1中获得的全细胞的转化率的测定,所选用色谱柱为Hypersil BDS C18(4.6*200mm,5um)。反应为1mL体系,4.1所得全细胞取800μL用于反应,向体系中加入200μL的40mM底物DAC,37℃,1000rpm,反应30min。取100μL样品,使用乙腈稀释十倍后用HPLC测定转化率。
HPLC流动相配比为:96%乙酸铵缓冲液(pH6.2,50mM),4%乙腈(v/v),流速1mL/min,检测波长为254nm,柱温箱恒定为25℃。
表8头孢菌素突变体对GL-7-D-ACA的转化率
实施例7固定化头孢菌素C酰化酶突变体
(1)环氧树脂、大孔吸附树脂固定酰化酶突变体(L159S/A419V/G160S)
载体平衡:用固定化缓冲液,按照载体/缓冲液1:5(质量/体积比)的比例反复清洗3次,清洗完成后抽干。
载体和酰化酶比例调节:用固定化缓冲液溶解酶,载体和酶缓冲液的比例为1:4(质量/体积比),该比例可进一步的在1:1到1:4之间进行优化。
将含酶的缓冲液和载体加入到反应器中,25℃、800rpm转速下反应3h,之后停止搅拌静置2h。
固定化后过滤收集固定化酶,使用280nm吸收波长测定上清液中蛋白含量,计算固定化率及产率。用蒸馏水清洗,重复3次。随后使用HPLC检测环氧树脂固定化酶的反应活性。
(2)氨基树脂固定化酶处理
载体平衡:固定化缓冲液,按照载体/磷酸盐缓冲溶液(50mM,pH8.0)1:5(质量/体积比)的比例反复清洗3次,清洗完过滤抽干。
用固定化缓冲液配置2%戊二醛溶液,将载体和戊二醛缓冲液以比例1:4(质量/体积比)加入,22℃,180rpm条件下搅拌1h。后用缓冲液清洗载体3遍,然后抽滤备用,存放到4℃冰箱中。
(3)固定化酶活力测试
反应条件(总体系4ml):固定化酰化酶(400mg)加入反应瓶中,投入底物CPC(40mM),补加缓冲液至4ml,,37℃,1000rpm,反应30min。取100μL样品,使用甲醇稀释十倍后用HPLC测定转化率。
检测条件:色谱柱Agilent ZORBAX SB-C18(4.6*150mm 4um);流动相配为96%磷酸盐缓冲液(pH7.0,50mM)添加4%甲醇(v/v);流速0.8mL/min;检测波长为254nm,柱温箱恒定为25℃。检测转化率可以达到67%。
<110>中国科学院天津工业生物技术研究所
<120>头孢菌素C酰化酶突变体及其在头孢菌素母核合成中的应用
<160>1
<170>PatentIn version 3.5
<210> 1
<211>774
<212> PRT
<213>砷锑氧化菌株(Bosea sp.)
<400>1
MTTDADRDALEAALPPLSGSLSLSGLEAPVSVRRDAWGIPHIKAVGEADAYRALGFVHAQDRLFQMELTRRKALGRAAEWLGAEAAEADILVRRLGMEQACRRDYDALADAAKAMLQAYAAGVNAFLESGAPVPCEYTLLGATPETWEPWHSIAVMRRLGLLMGSIWFKLWRMLALPIVGAENALKLRYDDGGRDLLCIPPGAQADRFEADLAALAPTVDHLLKAMGGDASDAAGGGSNNWAVGPDRTATGRPILAGDPHRVFEIPGMYAQHHLACDRFDMIGLTVPGVPGFPHFAHNGSVAYCVTHAFMDIHDLYLEQFTDEGRAVRFGEGFEPVIHRRDRIAVRGGADREFEIFETRHGPVIAGDPREGVGLSLRSVQFAETDLSFDCLTRMPAASTVSELYEATRGWGLIDHNLVAGDTAGAIGHLVRARVPVRSRDNGWLPVPGWTAEHEWQGWIAHEAMPCVIDPPGGLIVTANNRVVADDHPDYLCTDCHPPYRAERILQRLDSGAPFAVADAAAIHADTQSPHLDLFRTRLAVLGLREQADAERLRQDLLAWDGRMDADSTSAAAYNAFRRALTRIVTVRSRLEQATAHRFAAVAPGVSPQGQVWWAVPTLLRDDDAGMLNGLTWDDVLSEALSEAAATLTGKTWGEEHQPRFAHPLSAQFPDWAQRLDPPSLPVGGDGDTVLAIGIVPSAGPTATYGALARYVFDVGNWDNSRWAVFHGTSGHPASPHYADQNTPWSACEMVPMLYDWERIAAEAKTVQELVPPRA 774
- 脱乙酰氧头孢菌素C合成酶突变体及其在β-内酰胺抗生素母核合成中的应用
- 脱乙酰氧头孢菌素C合成酶突变体及其在β-内酰胺抗生素母核合成中的应用