泛素连接酶Trim23在调控树突状细胞分化成熟中的应用
文献发布时间:2023-06-19 19:28:50
技术领域
本发明属于细胞分化技术领域,具体涉及一种泛素连接酶Trim23在调控树突状细胞分化成熟中的应用。
背景技术
树突状细胞(dendritic cell, DC)是免疫系统中数量少但功能强的专职抗原提呈细胞,不仅可激活初始T细胞启动特异性免疫应答,还可诱导机体的体液免疫及非特异免疫,在抗肿瘤免疫应答中发挥着重要作用。由于DC在抗肿瘤免疫应答中不可或缺的地位及良好的临床疗效,其发现者RM Steinman在2011年被授予诺贝尔生理/医学奖。经典的 DC疫苗免疫治疗通常是在体外修饰或诱导激发性的DC,常用的方法有肿瘤细胞抗原与DCs共培养、合成的肿瘤抗原肽致敏DCs和肿瘤相关基因修饰DC等。目前以DC为基础的激发性疫苗已应用于多种肿瘤疾病并展现出许多优势,然而由于DC疫苗有效性的局限,如体外诱导的激发性DC在体内仍存在免疫活性降低、数量减少等现象,限制了它的临床客观应答率。如何有效地增强DC疫苗的抗肿瘤免疫应答进而提高其临床客观应答率,一直都是国内外科研努力研究的目标和动力。而增强DC疫苗抗肿瘤免疫效应的重要因素之一就是寻找到调控DC免疫功能的相关分子。
近年来,泛素化修饰作为蛋白质翻译后修饰的一种重要方式,参与了多种免疫细胞的生物学功能,其中在 DC 活化、成熟过程中扮演重要角色。已有研究发现,泛素连接酶MARCH1作为DC免疫功能的调控分子,可直接泛素化共刺激分子CD86和MHC-II,抑制DC的成熟度。Yang等研究发现,TLR受体介导的DC分化成熟中,泛素连接酶 Hrd1能诱导核内BLIMP泛素化降解,进而增强TLR的信号转导,促进DC表面MHC-II分子的表达及其诱导CD4
发明内容
本发明针对现有技术中的不足,目的是提供一种泛素连接酶Trim23在调控树突状细胞分化成熟中的应用。
为达到上述目的,本发明的解决方案是:
泛素连接酶Trim23在调控树突状细胞分化成熟中的应用。
一种泛素连接酶Trim23过表达载体,其包括上述的泛素连接酶Trim23基因和pcDNA3.1载体。
优选地,泛素连接酶Trim23基因序列(SEQ ID NO.1)如下:
ATGGCTGCCCTCGCTGTAAACAAGCCCGGAGCCGGAGTAGACAGTGGGCGGCAGGGCAGCCGGGGGACGGCTGTGGTGAAGGTTCTGGAGTGTGGGGTTTGTGAAGATGTCTTCTCTTTGCAAGGAGATAAAGTTCCTCGCCTCCTCCTCTGTGGCCACACAGTCTGTCATGACTGTCTTACTCGCTTGCCTCTGCATGGAAGAGCAATACGTTGCCCATTTGATCGACAAGTAACAGACCTAGGAGACTCAGGTGTATGGGGTTTGAAAAAGAACTTTGCTTTACTGGAGCTTTTAGAACGTTTGCAAAATGGGCACATTGGTCAGTATGGAGCTGCAGAAGAGGCCATTGGGACATCTGGAGAGAGCATCATTCGCTGTGATGAAGATGAAGCTCACGTTGCATCTGTATATTGCACTGTGTGTGCAACTCACTTGTGCTCAGACTGTTCTCAAGTTACTCATTCTACAAAGACATTAGCAAAGCATAGGCGTGTGCCTCTAGCTGATAAACCTCATGAGAAAACCATGTGCTGTCAGCACCAGGTGCATGCCATTGAGTTTGTTTGCTTGGAAGAAGGTTGTCAAACTAGTCCACTCATGTGCTGTGTCTGCAAAGAATATGGAAAGCACCAAGGTCACAAGCATTCAGTACTGGAACCAGAAGCTAATCAGATCCGAGCATCAATTTTAGACATGGCTCACTGCATACGGACCTTCACTGAGGAAATCTCAGATTATTCCAGAAAATTAGTTGGGATTGTTCAGCACATTGAAGGAGGAGAACAAATAGTAGAAGATGGAATTGGAATGGCTCACACAGAACATGTCCCAGGTACTGCAGAGAATGCCCGATCATGTGTCAGAGCTTATTTTTCTGATTTACATGAAACTCTTTGTCGTCAAGAAGAAATGGCTCTAAGTGTTGTTGATGCCCATGTTCGAGAAAAACTGATTTGGCTTAGGCAGCAACAAGAGGATATGACTATTCTCTTGTCCCAGGTCTCAACAGCCTGTCTCCACTGTGAAAAAACTCTGCAGCAGGATGATTGCAGAGTTGTCTTGGCAAAACAAGAAATCACAAGATTACTAGAAACACTGCAGAAACAGCAGCAGCAGTTTACAGAGGTTGCAGATCATATTCAGTTGGATGCCAGTATTCCAGTCACTTTTACAAAGGACAATAGAGTTCATATTGGACCCAAAATGGAAATCCGAGTAGTCACATTAGGATTAGATGGTGCTGGAAAAACTACCATTTTGTTCAAGTTAAAACAAGATGAATTTATGCAACCTATTCCAACAATTGGTTTTAATGTGGAAACTGTGGAATACAAAAATCTAAAATTCACCATTTGGGATGTGGGTGGAAAACACAAATTAAGACCATTGTGGAAACATTATTACCTCAATACACAAGCTGTTGTATTTGTTGTTGATAGTAGTCATAGAGACAGAATTAGTGAAGCACACAGTGAACTTGCAAAGTTGTTAACGGAAAAAGAACTCCGAGATGCCTTACTCTTGATTTTTGCTAACAAACAGGATGTAGCTGGAGCACTTTCAGTAGAAGAAATCACTGAACTTCTCAGTCTCCATAAACTATGCTGTGGAAGGAGTTGGTATATTCAGGGCTGTGATGCTCGAAGTGGAATGGGGCTCTACGAAGGGTTGGACTGGCTGTCCCGGCAACTTGTGGCTGCTGGAGTGTTGGATGTTGCTTGA。
一种如上述的泛素连接酶Trim23过表达载体抑制LPS诱导的树突状细胞的分化成熟。
一种如上述的泛素连接酶Trim23过表达载体抑制LPS激发的树突状细胞诱导Th1细胞的分化以及抗原特异性CD8
一种如上述的泛素连接酶Trim23过表达载体通过MAPK信号途径调控LPS诱导的树突状细胞分化成熟。
由于采用上述方案,本发明的有益效果是:
本发明通过免疫学、细胞生物学等技术发现泛素连接酶Trim23分子在DC分化成熟过程中发挥着重要的调控作用,过表达Trim23可显著抑制LPS诱导的DC表面共刺激分子的表达及炎性细胞因子的分泌水平,并明显降低DC诱导Th1细胞增殖分化的能力,进一步通过分子生物学技术发现Trim23主要通过MAPK信号通路影响DC的分化成熟,其作用机制依赖于Trim23的泛素化连接酶活性。发现了DC分化成熟过程中一个重要的负调控因子,为以后设计靶向Trim23的小分子抑制剂并联合免疫激活剂进一步增强DC疫苗的免疫学功能提供实验基础,也为临床DC肿瘤疫苗的开发及优化提供新的思路。
附图说明
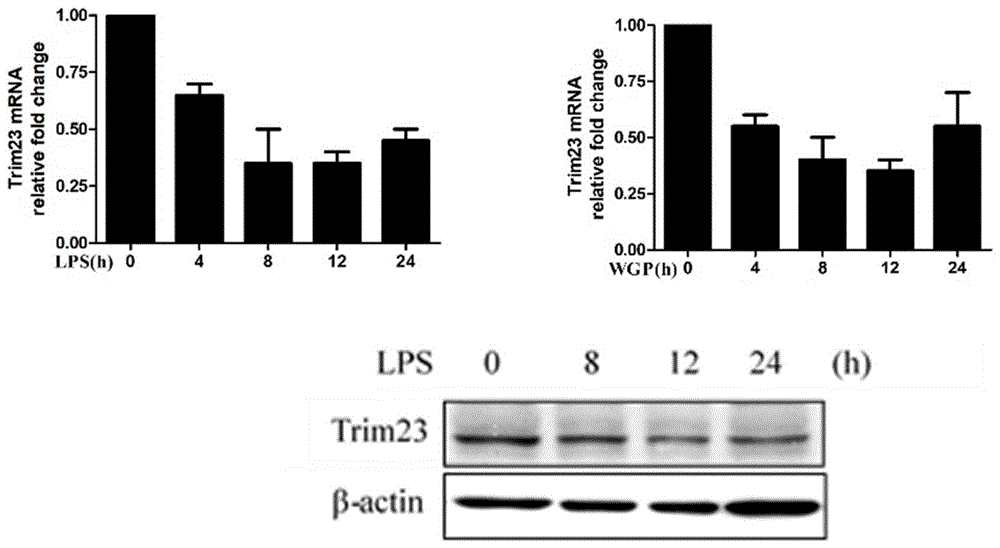
图1为Trim23在DC诱导成熟过程中的表达图。
图2为过表达Trim23对LPS诱导的DC表型及细胞因子分泌的影响图。
图3为过表达Trim23对LPS激发的DC诱导T细胞增殖分化的影响图。
图4为Western blot 检测DC中MAPK及AKT信号通路相关信号分子的磷酸化水平图。
图5为Trim23通过泛素连接酶活性抑制LPS激发的DC成熟图。
具体实施方式
本发明提供了一种泛素连接酶Trim23在调控树突状细胞分化成熟中的应用。
DC的免疫激活功能与其成熟状态密切相关,未成熟的DC通过模式识别受体(Toll样受体(TLR)、 C型凝集素受体(CLR)等)对病原体及细胞内容物等产生应答后逐渐成熟,并伴随上调表达主要组织相容性复合物(MHC-II)、共刺激分子(CD86、CD80 等)和分泌细胞因子(IL-12、TNF-α、IL-6等),为T细胞提供激活信号。研究表明DC的免疫激活受到泛素化修饰系统的严密调控,泛素连接酶能够通过调控 TLR、CLR 等信号转导通路影响DC的成熟及免疫应答功能。
三元基序蛋白(TRIMs)属于E3泛素连接酶家族,与固有免疫、适应性免疫和抗菌自噬的活化均有联系。大多数的TRIMs蛋白含有保守的RBCC结构域,包括一个泛素连接酶结构域(RING),一个或两个B盒结构域和一个螺线圈结构域。Trim23是TRIM家族的一个特殊成员,蛋白结构上除了含有RING结构域,还包含具有GTPase水解酶活性的ARF结构域。研究发现,Trim23在抗病毒免疫中扮演着重要角色,例如Trim23可泛素化修饰NEMO激活NF-κB,增强TLR3介导的抗病毒免疫反应,后有学者进一步研究发现Trim23调控病毒感染引起的自噬信号,同时依赖于RING结构域和ARF结构域,Trim23首先通过RING结构域介导自身泛素化后激活自身的GTP水解酶活性,从而诱导自噬相关分子TBK1的活化。近期研究报道,Trim23的高表达与肿瘤患者的不良预后相关,Trim23通过NF-κB信号通路促进肿瘤细胞的增殖。本发明主要研究Trim23对DC分化成熟及免疫功能活化的调控作用,并初步探讨相关的作用机制。
实施例:
实验动物:4-6周龄C57BL/6野生型小鼠购自常州卡文斯实验动物有限公司,CD4和CD8卵清蛋白T细胞受体转基因OT-Ⅱ和OT-I小鼠由清华大学祁海教授慷慨惠赠,所有小鼠在无特定病原体的条件下饲养。
主要试剂:重组小鼠FMS样酪氨酸激酶3配体(FMS-like tyrosine kinase 3ligand,Flt3L)购自PeproTech公司,RPMI 1640培养基、PBS购自美国GIBCO公司;胎牛血清(fetal bovine serum, FBS)购自BI公司;脂多糖(LPS)、卵清蛋白(ovalbumin,OVA)购自用sigma公司;ELISA试剂盒、流式细胞术抗体CD11c-PE、CD86-PerCp-Cy5.5、CD86-PerCp-Cy5.5、MHC Ⅱ-FITC、CD4-FITC、CD8-APC、IFN-γ-PE/Cy7、Permeabilization wash buffer及胞内染色固定液均购于Biolegend公司;BD Golgiplug
BMDC的培养及诱导成熟:将C57BL/6小鼠以颈椎脱臼法处死,取其腿骨。用PBS反复吹洗骨髓腔,收集所以细胞,经红细胞裂解液处理后,离心去上清,并用70 μmol/L细胞筛过滤去除杂质和细胞碎片,再次离心,并将细胞培养于含有100 ng/mL Flt3L的 RPMI1640完全培养基(100 mL/L FBS、非必需氨基酸、0.02 mM L-谷氨酰胺和0.05 mM 2-ME)。每2天换液一次,培养5天,加入LPS(100 ng/mL),继续培养2天,收获成熟DC。
过表达载体的构建及转染:小鼠Trim23过表达载体由吉玛基因公司构建完成,其中包括Trim23全基因过表达载体(aa 1-574,Trim23 OE)、敲除RING结构域的Trim23短截体(aa 77-574,Trim23ΔRING)、敲除ARF结构域的Trim23短截体(aa 403-574,Trim23 ΔARF)及对照pcDNA3.1空载体(CON)。细胞培养第5天,利用Lipofectamine
实时荧光定量PCR检测DC中Trim23 mRNA表达水平:采用TRIzol法提取细胞的总RNA,酶标仪检测其纯度和含量,并进行逆转录合成cDNA。定量PCR反应总体系参照说明书,其中上下游引物各1 μL,2×SYBR Green Supermix 10 μL,模板cDNA 2 μL。每组样品设2个复孔。PCR的反应程序为:95℃预变性3 min;95℃变性15 s,60℃退火30 s,总共进行 40个循环;融解曲线设定为60℃、10 s、95℃、10 s。mRNA的相对表达量计算公式为 2
流式细胞术检测:DC表面分子表达分析:收集不同转染组的DC,PBS洗涤2次,按照操作说明进行染色。在细胞中分别加入10 μg/mL的荧光基团标记的抗体:CD11c-PE、CD86-PerCP-Cy5.5、CD40-FITC、CD80-PerCP-Cy5.5、MHC-Ⅱ-FITC和同型对照抗体,4℃避光染色30 min。PBS洗涤2次,将细胞重悬于400 μL含20 mL/L FBS的PBS中,流式细胞仪检测分析。
检测DC诱导T细胞增殖分化的能力:按照前述方法培养BMDC,在第5天将Trim23 OE及CON组过表达载体分别转入DC。在实验第6天,无菌条件下取OT-I/OT-Ⅱ小鼠脾脏和淋巴结,研磨制备成单细胞悬液,磁珠分选出CD8
ELISA 检测细胞上清中TNF-α,IL-12p40,IL-6和IL-10的水平:各转染组BMDC经LPS诱导培养48 h,收集细胞培养上清,参照ELISA试剂盒的操作说明,应用双抗体夹心法检测各组细胞中IL12p40、IL-10、IL6和TNF-α在562nm处的A值。以上实验均进行3次独立重复实验。
蛋白质印迹(Western blot):收集待测细胞,PBS洗涤2次,弃上清。以下步骤均在冰上进行:样品中加入100 μL RIPA后冰上放置30 min,每隔 5 min震荡1次,12000rpm离心15min,吸取蛋白上清。蛋白样品用10% SDS-PAGE分离后转到PVDF膜。脱脂奶粉封闭1.5 h,PBST缓冲液洗涤3次,加入一抗(1:1000)4℃摇床过夜。次日,PBST洗膜后与HRP标记的二抗(1:2000)37℃孵育2 h,采用化学发光检测,曝光。β-actin作为内参蛋白。
统计学分析:应用 Graphpad Prism 9.0软件绘制柱状图,Flowjo10.7.2软件分析流式细胞术数据,并进行统计分析。两组间差异采用T检验,P<0.05为差异有统计学意义。
结果:
1.Trim23在小鼠骨髓来源的树突状细胞分化成熟中的表达
小鼠骨髓来源的单核细胞经Flt3L诱导为未成熟BMDC,LPS刺激4h、8h、12h、24h后,荧光定量PCR结果显示DC中Trim23的基因表达水平下调(图1)。前期的研究已证实,β-葡聚糖(WGP)作为DC表面CLR受体的配体分子可显著促进DC的成熟。接着进一步用WGP刺激BMDC4h、8h、12h、24h,结果显示Trim23的基因表达水平同样有所减少。Western blot检测Trim23蛋白的表达水平,结果显示,DC经LPS诱导后,Trim23蛋白水平显著下调(图1),这些结果说明Trim23可能在DC的分化成熟中起着重要作用。
2.过表达Trim23对LPS激发的DC表面共刺激分子表达及细胞因子分泌的影响
为了进一步确定Trim23对DC分化成熟的影响,选用DC诱导成熟的经典因子LPS进行后续实验。按照前述方法培养BMDC,分别转染过表达载体(Trim23 OE)和对照载体(CON)24 h,经LPS激发24 h后,荧光定量PCR和Westernblot结果显示,与对照组比较,Trim23 OE组中Trim23的表达水平显著上调(图2中A),表明Trim23过表达载体转染成功。流式结果显示,与CON组比较,过表达Trim23的DC表面MHC-II、CD86、CD80的表达水平明显下调(图2中B);ELISA结果显示,过表达Trim23可显著抑制DC对细胞因子TNF-α,IL-6的表达(图2中C),而IL-12p40,IL-10的表达在两组之间并无统计学差异(图2中C)。以上结果表明过表达Trim23对LPS诱导的DC分化成熟具有抑制作用。
3.过表达Trim23对LPS诱导的DC激发抗原特异性T细胞增殖分化的影响
不同转染组的DC与OVA特异性的T细胞共培养,流式结果显示,与CON组比较,过表达Trim23可显著抑制LPS激发的DC诱导CD4
4.Trim23对DC胞内MAPK及AKT信号通路中主要信号分子磷酸化水平的影响
为了进一步探索Trim23对LPS诱导DC分化成熟的分子机制,采用Western blot检测LPS处理不同时间时DC中p38 MAPK、ERK1/2和AKT的磷酸化水平。结果显示,LPS激发DC 15min后,ERK1/2和p38的磷酸化水平都有所增加,而AKT的磷酸水平尚无变化(图4)。当过表达Trim23后,在LPS未激发时DC中ERK1/2的磷酸化水平已有所降低,而激发15 min后显著大幅降低。此外,p38的磷酸化水平在LPS激发的Trim23 OE组中也明显下降,而AKT的磷酸化水平在对照组和过表达组中无变化(图4)。结果表明,Trim23影响DC内p38和ERK1/2蛋白的磷酸化,LPS刺激后进一步加剧其作用效果,说明Trim23可能通过MAPK信号途径调控LPS诱导的DC分化成熟。
5.Trim23通过泛素连接酶功能域抑制LPS诱导的DC成熟
研究表明,RING结构域是Trim23蛋白发挥泛素化修饰的功能域,而含有GTPase水解酶活性的ARF结构域在Trim23调控细胞生物学功能中也起着重要作用。本发明进一步探讨Trim23调控DC成熟的关键功能域。将缺失RING结构域或ARF结构域的Trim23突变序列分别插入pcDNA3.1载体中构建Trim23 ΔRING过表达载体(aa 77-574)和Trim23 ΔARF过表达载体(aa 403-574)。CON、Trim23 OE、Trim23ΔRING和Trim23 ΔARF过表达载体分别转染DC,经LPS激发后,流式结果显示,与Trim23 OE组比较,转染Trim23ΔRING载体的DC表面分子MHC-II和CD86的表达明显增加(图5中A),ELISA结果显示,细胞因子TNF-α,IL-6的分泌水平也显著上调(图5中B),而转染Trim23 ΔARF载体的DC表面分子及细胞因子的表达水平与Trim23 OE组一致(图5),说明敲除RING结构域的Trim23对DC分化成熟的抑制作用明显减弱。以上结果表明E3连接酶活性在Trim23调控LPS诱导的DC成熟中可能起着重要作用。
综上可知,在LPS或WGP(CLR相关受体Dectin-1的激活配体)诱导小鼠骨髓来源的DC成熟过程中,泛素连接酶Trim23的表达均显著降低。为了探索Trim23对DC分化成熟的影响,本发明构建了Trim23过表达载体,并选取DC的经典激活剂LPS作为其成熟的诱导因子,结果显示过表达Trim23可显著抑制LPS诱导的DC表面共刺激分子CD86、CD80、MHC-II的表达及细胞因子TNF-a、IL-6的分泌水平,这些结果表明泛素连接酶Trim23是LPS诱导DC成熟过程中的重要调控因子。
DC的成熟与其免疫功能的活化密切相关,进一步检测过表达Trim23的DC诱导T细胞增殖分化的能力。众所周知,在DC肿瘤疫苗开发中,评价DC肿瘤疫苗的疗效最重要的是看DC是否可高效地将肿瘤抗原提呈给T细胞并激发强大的Th1和肿瘤特异性CTL效应。本发明采用了OVA抗原特异性的OT-I和OT-II转基因小鼠。结果发现,上调Trim23表达后,DC介导的抗原特异性CD4
前期研究发现,Trim23主要参与调控I 型干扰素、自噬和NF-κB信号通路。在这些信号转导过程中,Trim23既可作为正向调节因子也可作为抑制分子发挥作用。LPS与细胞表面的TLR4受体结合,激活下游MyD88依赖的以及TRIF依赖的两条信号通路,其中My88介导的信号可激活下游MAPK、AKT等信号,引起相关蛋白发生磷酸化,并进一步引起NF-κB、AP-1的活化,诱导促炎因子TNF-α、IL-1β、IL-6等的分泌。前期研究发现,在WGP诱导DC的成熟中,WGP激活DC表面Dectin-1信号后,同样可引起其下游MAPK和AKT信号通路的活化。因此,本发明检测了DC胞内MAPK和AKT信号通路中主要信号分子的磷酸化水平,并发现过表达Trim23的DC中磷酸化的p38和ERK1/2蛋白含量显著减少,说明Trim23可能通过降低MAPK信号转导来抑制DC对细胞因子TNF-α、IL-6的分泌。
Trim23在功能上主要通过泛素化修饰来调控蛋白的激活参与细胞的生物学功能,其泛素化作用方式包括K27、K11和K48位修饰。随着对Trim23功能的深入研究,报道具有GTPase水解酶活性的ARF结构域在Trim23调控抗病毒免疫应答中起着不可或缺的作用。因此,本发明构建了缺失RING或ARF结构域的两个Trim23短截体,并发现缺失RING结构域的Trim23对DC分化成熟的抑制效果明显减弱,而缺失ARF结构域的Trim23对LPS诱导的DC成熟依然存在明显的抑制作用,说明RING结构域是Trim23发挥调控作用的关键功能域,Trim23可能通过泛素化修饰模式参与LPS诱导的DC成熟。
则本发明证明了Trim23可抑制LPS诱导的DC成熟,并减弱其抗肿瘤免疫应答能力,其作用机制依赖于DC胞内MAPK信号以及Trim23的泛素连接酶功能域。为后续的体内研究以及临床DC肿瘤疫苗的开发及应用提供了一定的实验基础。
对于本领域技术人员而言,显然本发明不限于上述示范性实施例的细节,而且在不背离本发明精神或基本特征的情况下,能够以其他的具体形式实现本发明。因此,无论从哪一点来看,均应将实施例看作是示范性的,而且是非限制性的,本发明的范围由所附权利要求而不是上述说明限定,因此旨在将落在权利要求的等同要件的含义和范围内的所有变化囊括在本发明内。不应将权利要求中的任何附图标记视为限制所涉及的权利要求。