双特异性抗CD28 X抗CD22抗体以及其用途
文献发布时间:2023-06-19 11:57:35
本申请要求于2018年12月19日提交的美国临时申请第62/781,689号的优先权的权益,所述美国临时申请的全部内容通过引用并入本文。
本申请含有序列表,所述序列表已经以ASCII格式以电子方式提交并且通过全文引用的方式特此并入。创建于2019年12月16日的所述ASCII副本命名为118003_49220_SL.txt并且大小为104,353字节。
技术领域
本发明涉及结合CD28和如CD22等靶分子的双特异性抗原结合分子以及其使用方法。
背景技术
CD28是在T细胞的表面上表达的I型跨膜蛋白,所述I型跨膜蛋白具有组装成同源二聚体的单个细胞外Ig-V样结构域。CD28是CD80(B7.1)蛋白和CD86(B7.2)蛋白的受体并且由在抗原呈递细胞(APC)上表达的CD80或CD86活化。CD28与CD80或CD86的结合提供了对于T细胞活化和存活而言具有重要性的共刺激信号。除了T细胞受体(TCR)之外,通过CD28的T细胞刺激也为各种白细胞介素的产生提供了强效信号。CD28在TCR活化之后还加强细胞信号,如由NFκB转录因子控制的通路。CD28共信号对于有效的T细胞活化如T细胞分化、增殖、细胞因子释放和细胞死亡而言具有重要性。
已提出抗CD28抗体用于涉及T细胞的活化的治疗性目的。在2006年的临床试验中使用一种特定的抗CD28抗体TGN1412(抗CD28超激动剂)。以0.1mg/kg的剂量向六名健康志愿者静脉给药TGN1412(抗CD28超激动剂)。在两个小时内,所有六名患者均有显著的炎症应答(细胞因子风暴),并且在十六个小时内,所有患者均出现多器官衰竭。用皮质类固醇治疗受试者,并且细胞因子水平在2-3天内恢复到正常水平。在1期研究(与CRS相关)中,0.1mg/kg的起始剂量是基于50mg/kg猕猴“NOAEL”的500倍的倍数(Suntharalingam等人,1期试验中抗CD28单克隆抗体TGN1412的细胞因子风暴(Cytokine Storm in a Phase 1Trial ofthe Anti-CD28 Monoclonal Antibody TGN1412),《NEJM》355:1018-1028(2006))。遗憾的是,TGN1412诱导了细胞因子风暴,这是通过毒理学研究在食蟹猴(cynomolgus macaques)中或通过离体人PBMC研究未预测到的。
Siglec家族成员CD22(也被称为Siglec-2)特异性地识别α2,6唾液酸并且是在B淋巴细胞(B细胞)上优先表达的跨膜蛋白。
CD22有许多先赋功能,包含例如B细胞稳态、B细胞存活和迁移、阻尼TLR和CD40信号传导和通过胞质区中的免疫受体酪氨酸基抑制基序(ITIM)的磷酸化作用招募含SH2结构域的磷酸酶来抑制B细胞受体(BCR)信号传导以及有助于B细胞和其它细胞类型之间的细胞粘着。
既没有在发育的早期阶段期间在B细胞的表面上发现CD22,也没有在干细胞中表达CD22。然而,所有B细胞淋巴瘤和白血病中的60-70%表达CD22。
已经研究了用于治疗B细胞淋巴瘤和白血病的抗CD22抗体。然而,单克隆抗体依帕珠单抗(Epratuzumab)取得的成功有限。(Grant等人(2013)《癌症(Cancer)》119(21):10.1002/cncr.28299)。
因此,本领域中需要改进的抗CD22抗体。还需要安全用于在药物组合物中使用的抗CD28抗体。此外,结合CD28和靶抗原(如CD22)两者的双特异性抗原结合分子在这样的治疗环境中将会是有用的:其中期望对表达靶抗原的细胞进行特异性靶向和T细胞介导的杀伤。
发明内容
在第一方面,本发明提供了结合CD28和靶抗原的双特异性抗原结合分子。根据某些示例性实施例,所述双特异性抗原结合分子结合CD28和CD22;这种双特异性抗原结合分子在本文中也称为“抗CD28/抗CD22双特异性分子”。所述抗CD28/抗CD22双特异性分子的抗CD22部分可用于靶向表达CD22的癌细胞(例如,癌性B细胞),并且所述双特异性分子的抗CD28部分可用于活化T细胞。癌细胞上的CD22和T细胞上的CD28的同时结合促进了通过活化的T细胞(例如,在T细胞的TCR活化之后)对靶向的癌细胞的定向杀伤(细胞裂解)。因此,本发明的抗CD28/抗CD22双特异性分子尤其可用于治疗与CD22表达性肿瘤相关或由其引起的疾病和病症(例如,B细胞增殖性病症,例如,B细胞淋巴瘤,例如弥漫性大B细胞淋巴瘤(DLBCL)、滤泡性淋巴瘤(FL)、边缘带淋巴瘤)。
根据本发明的此方面的双特异性抗原结合分子包括特异性地结合人CD28的第一抗原结合结构域和特异性地结合CD22第二抗原结合结构域。本发明包含抗CD28/抗CD22双特异性分子(例如,双特异性抗体),其中每个抗原结合结构域包括与轻链可变区(LCVR)配对的重链可变区(HCVR)。在本发明的某些示例性实施例中,抗CD28抗原结合结构域和抗CD22抗原结合结构域各自包括与共同LCVR配对的不同的独特HCVR。
本发明提供了抗CD28/抗CD22双特异性分子,其中特异性地结合CD28的第一抗原结合结构域包括如表6中所示的HCVR氨基酸序列中的任何一个。特异性地结合CD28的第一抗原结合结构域还可以包括如表6中所示的LCVR氨基酸序列中的任何一个。根据某些实施例,特异性地结合CD28的第一抗原结合结构域包括如表6中所示的HCVR/LCVR氨基酸序列对中的任何一个。本发明还提供了抗CD28/抗CD22双特异性分子,其中特异性地结合CD28的第一抗原结合结构域包括如表6中所示的重链CDR1-CDR2-CDR3氨基酸序列中的任何一个和/或如表6中所示的轻链CDR1-CDR2-CDR3氨基酸序列中的任何一个。
根据某些实施例,本发明提供了抗CD28/抗CD22双特异性分子,其中特异性地结合CD28的第一抗原结合结构域包括重链可变区(HCVR),所述HCVR具有选自由SEQ ID NO:28和26组成的组的氨基酸序列或其具有至少90%、至少95%、至少98%或至少99%序列同一性的基本上类似的序列。
本发明还提供了抗CD28/抗CD22双特异性分子,其中特异性地结合CD28的第一抗原结合结构域包括轻链可变区(LCVR),所述LCVR具有SEQ ID NO:10的氨基酸序列或其具有至少90%、至少95%、至少98%或至少99%序列同一性的基本上类似的序列。
本发明还提供了抗CD28/抗CD22双特异性分子,其中特异性地结合CD28的第一抗原结合结构域包括选自由SEQ ID NO:28/10和26/10组成的组的HCVR和LCVR(HCVR/LCVR)氨基酸序列对。
本发明还提供了抗CD28/抗CD22双特异性分子,其中特异性地结合CD28的第一抗原结合结构域包括:重链CDR3(HCDR3)结构域,所述HCDR3结构域具有SEQ ID NO:32的氨基酸序列或与其具有至少90%、至少95%、至少98%或至少99%序列同一性的基本上类似的序列;以及轻链CDR3(LCDR3)结构域,所述LCDR3结构域具有SEQ ID NO:16的氨基酸序列或其具有至少90%、至少95%、至少98%或至少99%的序列同一性的基本上类似的序列。
在某些实施例中,特异性地结合CD28的第一抗原结合结构域包括SEQ ID NO:32/16的HCDR3/LCDR3氨基酸序列对。
本发明还提供了抗CD28/抗CD22双特异性抗原结合分子,其中特异性地结合CD28的第一抗原结合结构域包括:重链CDR1(HCDR1)结构域,所述HCDR1结构域具有SEQ ID NO:28的氨基酸序列或其具有至少90%、至少95%、至少98%或至少99%序列同一性的基本上类似的序列;重链CDR2(HCDR2)结构域,所述HCDR2结构域具有SEQ ID NO:30的氨基酸序列或其具有至少90%、至少95%、至少98%或至少99%序列同一性的基本上类似的序列;轻链CDR1(LCDR1)结构域,所述LCDR1结构域具有SEQ ID NO:12的氨基酸序列或其具有至少90%、至少95%、至少98%或至少99%序列同一性的基本上类似的序列;以及轻链CDR2(LCDR2)结构域,所述LCDR2结构域具有SEQ ID NO:14的氨基酸序列或其具有至少90%、至少95%、至少98%或至少99%序列同一性的基本上类似的序列。
本发明的某些非限制性、示例性抗CD28/抗CD22双特异性抗原结合分子包含特异性地结合CD28的第一抗原结合结构域,所述第一抗原结合结构域包括分别具有以下的氨基酸序列的HCDR1-HCDR2-HCDR3-LCDR1-LCDR2-LCDR3结构域:SEQ ID NO:28-30-32-12-14-16。
本发明还提供了抗CD28/抗CD22双特异性分子,其中特异性地结合CD22的第二抗原结合结构域包括重链可变区(HCVR),所述HCVR具有选自由SEQ ID NO:2和18组成的组的氨基酸序列或其具有至少90%、至少95%、至少98%或至少99%序列同一性的基本上类似的序列。
本发明还提供了抗CD28/抗CD22双特异性分子,其中特异性地结合CD22的第二抗原结合结构域包括轻链可变区(LCVR),所述LCVR具有选自SEQ ID NO:10的氨基酸序列或其具有至少90%、至少95%、至少98%或至少99%序列同一性的基本上类似的序列。
本发明还提供了抗CD28/抗CD22双特异性分子,其中特异性地结合CD22的第二抗原结合结构域包括选自由SEQ ID NO:2/10和18/10组成的组的HCVR和LCVR(HCVR/LCVR)氨基酸序列对。
本发明还提供了抗CD28/抗CD22双特异性分子,其中特异性地结合CD22的第二抗原结合结构域包括:重链CDR3(HCDR3)结构域,所述HCDR3结构域具有选自由SEQ ID NO:8和24组成的组的氨基酸序列或与其具有至少90%、至少95%、至少98%或至少99%序列同一性的基本上类似的序列;以及轻链CDR3(LCDR3)结构域,所述LCDR3结构域具有选自SEQ IDNO:16的氨基酸序列或其具有至少90%、至少95%、至少98%或至少99%序列同一性的基本上类似的序列。
在某些实施例中,特异性地结合CD22的第二抗原结合结构域包括选自由SEQ IDNO:8/16和24/16组成的组的HCDR3/LCDR3氨基酸序列对。
本发明还提供了抗CD28/抗CD22双特异性抗原结合分子,其中特异性地结合CD22的第二抗原结合结构域包括:重链CDR1(HCDR1)结构域,所述HCDR1结构域具有选自由SEQID NO:4和20组成的组的氨基酸序列或其具有至少90%、至少95%、至少98%或至少99%序列同一性的基本上类似的序列;重链CDR2(HCDR2)结构域,所述HCDR2结构域具有选自由SEQID NO:6和22组成的组的氨基酸序列或其具有至少90%、至少95%、至少98%或至少99%序列同一性的基本上类似的序列;轻链CDR1(LCDR1)结构域,所述LCDR1结构域具有SEQ IDNO:12的氨基酸序列或其具有至少90%、至少95%、至少98%或至少99%序列同一性的基本上类似的序列;以及轻链CDR2(LCDR2)结构域,所述LCDR2结构域具有SEQ ID NO:14的氨基酸序列或其具有至少90%、至少95%、至少98%或至少99%序列同一性的基本上类似的序列。
本发明的某些非限制性、示例性抗CD28/抗CD22双特异性抗原结合分子包含特异性地结合CD22的第二抗原结合结构域,所述第二抗原结合结构域包括分别具有选自由以下组成的组的氨基酸序列的HCDR1-HCDR2-HCDR3-LCDR1-LCDR2-LCDR3结构域:SEQ ID NO:4-6-8-12-14-16和20-22-24-12-14-16。
在相关实施例中,本发明包含抗CD28/抗CD22双特异性抗原结合分子,其中特异性地结合CD22的第二抗原结合结构域包括选自由SEQ ID NO:2/10和18/10组成的组的重链可变区和轻链可变区(HCVR/LCVR)序列内含有的重链和轻链CDR结构域。
在另一个方面,本发明提供了对本文公开的抗CD28/抗CD22双特异性抗原结合分子的HCVR、LCVR或CDR序列中的任何一个进行编码的核酸分子,所述核酸分子包含:包括如本文表7中所示的多核苷酸序列的核酸分子以及包括如表7中所示的多核苷酸序列中的两个或更多个的核酸分子(以其任何功能组合或布置)。携带本发明的核酸的重组表达载体和引入了此类载体的宿主细胞也涵盖在本发明中,通过在允许产生抗体的条件下培养宿主细胞并回收产生的抗体来产生抗体的方法也是如此。
本发明包含抗CD28/抗CD22双特异性抗原结合分子,其中特异性地结合CD28的上述抗原结合结构域中的任何一个与特异性地结合CD22的上述抗原结合结构域中的任何一个组合、连接或以其它方式缔合,以形成结合CD28和CD22的双特异性抗原结合分子。
本发明包含具有经修饰的糖基化模式的抗CD28/抗CD22双特异性抗原结合分子。在一些应用中,用于去除不期望的糖基化位点的修饰可以是有用的,或者例如用于增加抗体依赖性细胞毒性(ADCC)功能的缺乏寡糖链上存在的岩藻糖部分的抗体(参见Shield等人(2002),《JBC》277:26733)。在其它应用中,可以进行半乳糖基化的修饰,以修饰补体依赖性细胞毒性(CDC)。
在另一个方面,本发明提供了一种药物组合物,其包括如本文所公开的抗CD28/抗CD22双特异性抗原结合分子以及药学上可接受的载体。在相关方面,本发明的特征在于一种组合物,其是抗CD28/抗CD22双特异性抗原结合分子和第二治疗剂的组合。在一个实施例中,第二治疗剂是有利地与抗CD28/抗CD22双特异性抗原结合分子组合的任何药剂。可以有利地与抗CD28/抗CD22双特异性抗原结合分子组合的示例性药剂在本文的其它地方详细讨论。
在又另一个方面,本发明提供了用于使用本发明的抗CD28/抗CD22双特异性抗原结合分子靶向/杀伤表达CD22的癌细胞的治疗性方法,其中所述治疗性方法包括向有需要的受试者施用治疗有效量的包括本发明的抗CD28/抗CD22双特异性抗原结合分子的药物组合物。
本发明还包含本发明的抗CD28/抗CD22双特异性抗原结合分子在制造用于治疗与CD22表达相关或由其引起的疾病或病症的药物中的用途。
在又另一个方面,本发明提供了用于使用本发明的抗CD28/抗CD22双特异性抗原结合分子靶向/杀伤表达CD22的癌细胞的治疗性方法,其中所述抗CD28/抗CD22双特异性抗原结合分子与同CD3结合的其它抗肿瘤双特异性抗原结合分子组合(例如,抗CD28/抗CD22与抗CD3/抗CD20抗体组合)。
在仍另一个方面,本发明提供了用于使用本发明的抗CD28/抗CD22双特异性抗原结合分子靶向/杀伤表达CD22的癌细胞的治疗性方法,其中所述抗CD28/抗CD22双特异性抗原结合分子与靶向PD-1、PD-L1或CTLA-4的检查点抑制剂组合(例如,抗CD28/抗CD22与抗PD1抗体组合)。例如,在某些实施例中,本发明的抗CD28/抗CD22抗体可以与靶向PD-1的药剂如派姆单抗(Pembrolizumab)
在仍另一个方面,本发明提供了用于使用本发明的抗CD28/抗CD22双特异性抗原结合分子靶向/杀伤表达CD22的癌细胞的治疗性方法,其中所述抗CD28/抗CD22双特异性抗原结合分子与同CD3结合的其它抗肿瘤双特异性抗原结合分子(例如,抗CD28/抗CD22与抗CD3/抗CD20双特异性抗体例如REGN1979(参见US9,657,102,其中抗CD20臂包括SEQ ID NO:1242/1258的HCVR/LCVR氨基酸对并且抗CD3臂包括SEQ ID NO:1250/1258的氨基酸对))组合和/或与靶向PD-1、PD-L1或CTLA-4的检查点抑制剂组合(即,抗CD28/抗CD22与抗PD-1抗体组合)。例如,在某些实施例中,本发明的抗CD28/抗CD22抗体可以与靶向PD-1的药剂如派姆单抗
通过阅读随后的详细描述,其它实施例将变得显而易见。
附图说明
图1是描绘了抗CD28/抗CD22双特异性抗体与在细胞表面上表达CD28的人CD4+T细胞和表达人CD22的靶细胞的结合的一组图。
图2A和2B是描绘了抗CD28/抗CD22双特异性抗体在原代T细胞刺激和CD22靶向表达存在的情况下示出增加的荧光素酶产生的一组图。图2A是描绘了如通过荧光素酶产生所评估的除了200pM恒定REGN1945(阴性hIgG4同种型对照)之外还与HEK293/hCD20、HEK293/hCD20/hCD22或Raji/CD80和CD86阴性细胞共温育的工程化报告T细胞的活化的一组图。图2B是描绘了如通过荧光素酶产生所评估的除了200pM恒定REGN2281(抗CD20 x抗CD3)之外还与HEK293/hCD20、HEK293/hCD20/hCD22或Raji/CD80和CD86阴性细胞共温育的工程化报告T细胞的活化的一组图。
图3A和图3B是描绘了抗CD28/抗CD22双特异性抗体在原代T细胞刺激和CD22靶向表达存在的情况下增加IL-2产生的一组图。更具体地,图3A是描绘了如通过IL-2产生所评估的在2nM恒定REGN1945(hIgG4同种型对照)存在的情况下与HEK293/hCD20、HEK293/hCD20/hCD22或Raji/CD80和CD86阴性细胞共温育的CD4
图4是示出了在工程化以表达PD-L1的细胞中REGN5837与西米普利单抗的组合将IL-2释放增强超过仅REGN5837治疗的一组图。
图5A是示出了REGN5837与西米普利单抗的组合在工程化以表达PD-L1的NALM6细胞存在的情况下增强IL-2释放的一组图。
图5B是示出了在工程化以表达PD-L1的RAJI细胞中REGN5837与西米普利单抗的组合将IL-2释放增强超过单独的REGN5837治疗的一组图。
图6是示出了在REGN1979(抗CD20 x抗CD3)存在的情况下用REGN5837治疗携带NALM-6-Luc肿瘤的小鼠与显著的肿瘤抑制相关的一组图。简而言之,向NSG小鼠(每组n=6到9)移植人PBMC、然后在移植后(第0天)12天植入NALM-6-luc B细胞白血病细胞。在植入后(箭头)8天、15天和22天向小鼠施用4mg/kg REGN5837+0.04mg/kg REGN1979(散列圆形)、0.4mg/kg REGN5837+0.04mg/kg REGN1979(封闭的直立三角形)、0.04mg/kg REGN5837+0.04mg/kg REGN1979(菱形)、4mg/kg non-TAAxCD28+0.04mg/kg REGN1979(正方形)、4mg/kg REGN5837+0.4mg/kg非-TAAxCD3(开放圆形)或4mg/kg非TAAxCD28+0.4mg/kg非TAAxCD3(封闭的倒三角形)。在植入后6天、10天、14天、17天、20天和23天通过对肿瘤体积的生物发光成像来监测肿瘤生长。将组合数据表示为组平均值±SEM。使用双向ANOVA和图基事后检验(Tukey's post hoc test)确定统计学显著性。使用以下符号来指示相对于非TAAxCD28+非TAAxCD3对照的统计学显著差异:
图7A-7C是示出了REGN1979活化并指导人T细胞以剂量依赖性方式杀伤Nalm6细胞的图。更具体地,图7A是描绘了在指示的抗体存在的情况下Nalm6细胞的存活率百分比的图。图7B是描绘了表达CD25(CD25+)的CD8+细胞在指示的抗体存在的情况下的百分比的图。图7C是描绘了在指示的抗体存在的情况下如通过CellTrace Violet稀释所评估的CD25+CD8+细胞的增殖的图。
图8A、8B和8C是示出了REGN1979活化并指导人T细胞以剂量依赖性方式杀伤WSU-DLCL2细胞的图。更具体地,图8A是示出了在指示的抗体存在的情况下WSU-DLCL2细胞的存活率百分比的图。图8B是描绘了表达CD25(CD25+)的CD8+细胞在指示的抗体存在的情况下的百分比的图。图8C是描绘了表示为已分裂%的在指示的抗体存在的情况下的CD8+细胞增殖的图。
图9是示出了在利用人PBMC和WSU-DLCL2细胞的测定中REGN1979诱导了人细胞因子IL-2、IL-4、IL-6和IL-10的释放的一组图。与仅由REGN1979诱导的细胞因子释放相比,在固定浓度的CD22 X CD28存在的情况下增强了用REGN1979观察到的细胞因子释放。
图10A-10E是示出了REGN1979活化并指导人T细胞以剂量依赖性方式耗竭NHL的图。与具有单臂CD28对照抗体或没有共刺激对照的REGN1979相比,将固定浓度的CD22xCD28双特异性抗体添加到REGN1979使REGN1979的细胞毒性功效(EC50)增强了2.3倍和3.5倍。如分别通过CD8+和CD4+细胞上的CD25上调或CellTrace Violet稀释测量的,观察到的由REGN1979介导的靶细胞裂解与T细胞活化和增殖相关。更具体地,图10A是描绘了在指示的抗体存在的情况下来自患者骨髓的NHL细胞的存活率百分比的图。图10B是描绘了表达CD25(CD25+)的CD8+细胞在指示的抗体存在的情况下的百分比的图。图10C是描绘了在指示的抗体存在的情况下如通过CellTrace Violet稀释评估的CD8+细胞的增殖的图。图10D是描绘了表达CD25(CD25+)的CD4+细胞在指示的抗体存在的情况下的百分比的图。图10E是描绘了在指示的抗体存在的情况下如通过CellTrace Violet稀释评估的CD4+细胞的增殖的图。
图11A-11E是示出了REGN5837以浓度依赖性方式增强了REGN1979介导细胞毒性的效力、CD25的细胞表面表达和T细胞增殖的图。简而言之,WSU-DLCL2细胞与淋巴细胞富集的人PBMC以1:5的靶细胞:PBMC比并且与抗CD20xCD3(REGN1979)在一定的浓度范围内(4.8fM到10nM)作为单一药剂(即,无REGN5837)或在固定浓度的REGN5837(范围为0.01到15μg/mL)存在的情况下在37℃温育72小时。缺乏REGN1979的情况仅含有指示的浓度下的REGN5837并且绘制为0.04pM。使用活/死细胞染色(11A)通过流式细胞术检测活细胞。使用Violet CellTracker染料以及针对CD2、CD4、CD8和CD25的的荧光团标记抗体的表型混合物来检测T细胞活化(如CD25表达所测量的;11B、11D)和通过流式细胞术)的CD4
更具体地,图11A是描绘了在所指示浓度的REGN5837下的死细胞%的图。图11B是描绘了在所指示浓度的REGN5837下的CD25+CD4+细胞的百分比的图。图11C是描绘了在所指示浓度的REGN5837下如通过CellTrace Violet稀释评估的CD4+细胞的增殖的图。图11D是描绘了CD25+CD8+细胞的百分比的图。图11E是描绘了在所指示浓度的REGN5837下如通过CellTrace Violet稀释评估的CD8+细胞的增殖的图。
图12A-12G是示出了REGN5837在WSU-DLCL2 B细胞淋巴瘤细胞存在的情况下以浓度依赖性方式增强了来自人T细胞的REGN1979介导的细胞因子释放的效力和最大水平的图。简而言之,WSU-DLCL2细胞与淋巴细胞富集的人PBMC以1:5的靶细胞:PBMC比并且与抗CD20xCD3(REGN1979)在一定的浓度范围内(4.8fM到10nM)作为单一药剂(即,无REGN5837)或在固定浓度的REGN5837(范围为0.01到15μg/mL)存在的情况下在37℃温育72小时。缺乏REGN1979的情况仅含有指示的浓度下的REGN5837并且绘制为0.04pM。使用BD流式细胞珠阵列人Th1/Th2/Th17细胞因子试剂盒评估上清液的(12A)IL-2、(12B)IL-4、(12C)IL-6、(12D)IL-10、(12E)TNF-α、(12F)IFN-γ和(12G)IL-17α的细胞因子释放。
更具体地,图12A是描绘了在所指示浓度的REGN5837下在WSU-DLCL2细胞存在的情况下从人T细胞释放的IL-2的水平的图。图12B是描绘了在所指示浓度的REGN5837下在WSU-DLCL2细胞WSU-DLCL2细胞存在的情况下从人T细胞释放的IL-4的水平的图。图12C是描绘了在所指示浓度的REGN5837下在WSU-DLCL2细胞WSU-DLCL2细胞存在的情况下从人T细胞释放的IL-6的水平的图。图12D是描绘了在所指示浓度的REGN5837下在WSU-DLCL2细胞WSU-DLCL2细胞存在的情况下从人T细胞释放的IL-10的水平的图。图12E是描绘了在所指示浓度的REGN5837下在WSU-DLCL2细胞WSU-DLCL2细胞存在的情况下从人T细胞释放的TNF-α的水平的图。图12F是描绘了在所指示浓度的REGN5837下在WSU-DLCL2细胞WSU-DLCL2细胞存在的情况下从人T细胞释放的IFN-γ的水平的图。图12G是描绘了在所指示浓度的REGN5837下在WSU-DLCL2细胞WSU-DLCL2细胞存在的情况下从人T细胞释放的IL-17α的水平的图。
图13A和13B是示出了在REGN1979以0.4或4mg/kg存在的情况下用REGN5837治疗携带WSU-DLCL2肿瘤的NSG小鼠与显著肿瘤抑制相关的图。简而言之,将WSU-DLCL2 B细胞淋巴瘤细胞和人PBMC的1:1混合物植入雌性NSG小鼠(每组n=6到7)(第0天)。在植入后第1天、第8天和第15天(箭头)向小鼠施用1mg/kg REGN5837和0.4mg/kg(13A)或4mg/kg(13B)REGN1979(或非桥接对照)的组合在植入后第7天、第10天、第14天、第16天、第28天、第31天、第35天、第38天、第43天、第46天、第49天、第53天、第57天和第63天通过卡尺测量来监测肿瘤生长。将组合数据表示为组平均值±SEM。使用双向ANOVA和图基事后检验确定统计学显著性。以下符号用于指示组之间的统计学上显著的差异:
更具体地,图13A是描绘了施用了1mg/kg REGN5837和0.4mg/kg REGN1979(或非桥接对照,非TAAxCD3)的小鼠的肿瘤生长的图。图13B是描绘了施用了1mg/kg REGN5837和4mg/kg(或非桥接对照,非TAAxCD3)的小鼠的肿瘤生长的图。
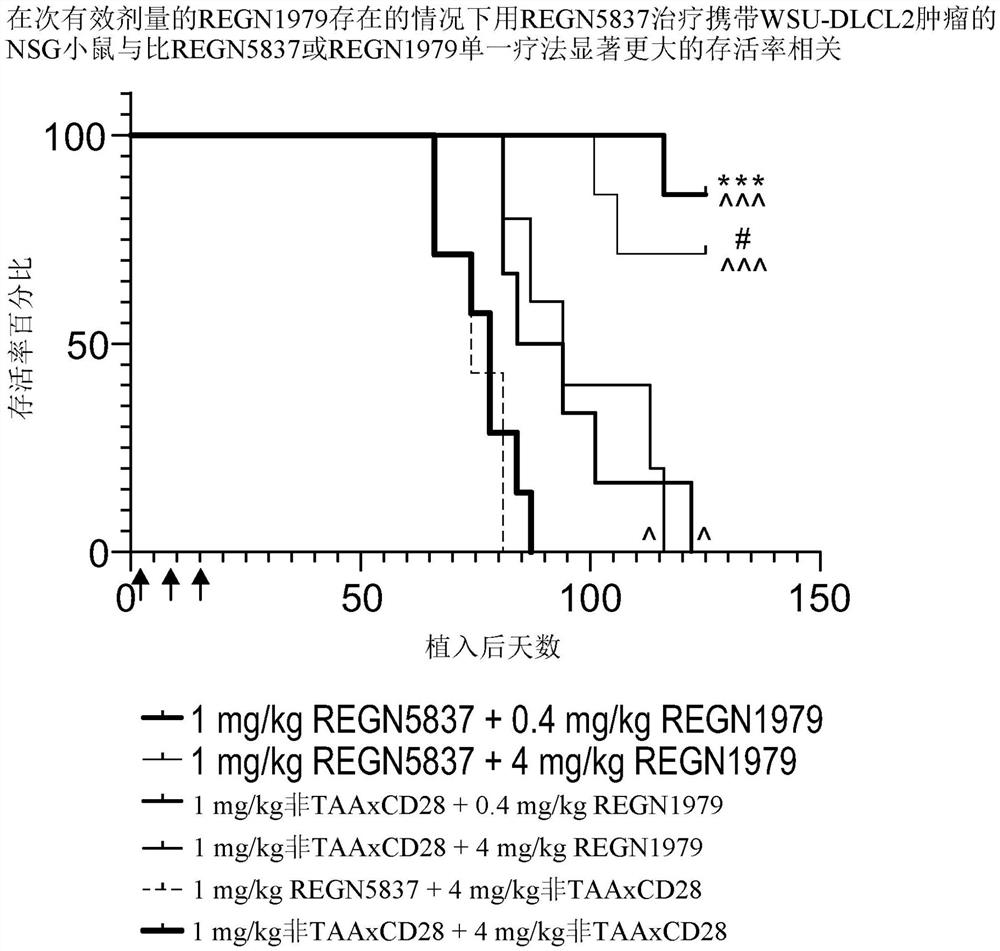
图14是示出了在次有效剂量的REGN1979存在的情况下用REGN5837治疗携带WSU-DLCL2肿瘤的小鼠与比REGN5837或REGN1979单一疗法显著更大的存活率相关的图。简而言之,将WSU-DLCL2 B细胞淋巴瘤细胞和人PBMC的1:1混合物植入雌性NSG小鼠(每组n=6到7)(第0天)。在植入后第1天、第8天和第15天(箭头)向小鼠施用REGN5837和REGN1979或对照的组合。使用Mantel-Cox检验确定统计学显著性。以下符号用于指示组之间的统计学上显著的差异:
具体实施方式
在描述本发明之前,应当理解,本发明不限于所描述的特定方法和实验条件,因为此类方法和条件可以变化。还应当理解,本文所使用的术语仅出于描述特定实施例的目的,而并不旨在进行限制,因为本发明的范围仅由所附权利要求限制。
除非另外定义,否则本文所使用的所有技术术语和科学术语具有与本发明所属领域的普通技术人员通常所理解的含义相同的含义。如本文所使用的,当用于提及特定所列举数值时,术语“约”意味着值可以与所列举值相差不超过1%。例如,如本文所使用的,表述“约100”包含99和101以及介于两者之间的所有值(例如,99.1、99.2、99.3、99.4等)。
尽管在本发明的实践或测试中可以使用类似于或等同于本文所述的方法和材料的任何方法和材料,但现在描述优选的方法和材料。本说明书中提及的所有专利、申请和非专利出版物均通过全文引用的方式并入本文。
定义
如本文所使用的,表述“CD28”是指作为共刺激受体在T细胞上表达的抗原。人CD28包括如SEQ ID NO:74中所示的氨基酸序列,和/或具有如NCBI登录号NP_006130.1中所示的氨基酸序列。除非明确指定为来自非人物种,否则本文对蛋白质、多肽和蛋白质片段的所有提及旨在指代相应蛋白质、多肽或蛋白质片段的人形式。因此,除非指定为来自非人物种例如“小鼠CD28”、“猴CD28”等,否则表述“CD28”意指人CD28。
如本文所使用的,“结合CD28的抗体”或“抗CD28抗体”包含特异性地识别单体CD28的抗体和其抗原结合片段以及特异性地识别二聚体CD28的抗体和其抗原结合片段。本发明的抗体和抗原结合片段可以结合可溶性CD28和/或细胞表面表达的CD28。可溶性CD28包含天然CD28蛋白以及缺乏跨膜结构域或以其它方式与细胞膜未缔合的重组CD28蛋白变体,例如单体和二聚体CD28构建体。
如本文所使用的,表述“细胞表面表达的CD28”意指在体外或体内在细胞的表面上表达使得CD28蛋白的至少一部分暴露于细胞膜的细胞外侧的并且可被抗体的抗原结合部分接近的一个或多个CD28蛋白。“细胞表面表达的CD28”包含在细胞的膜中的功能性T细胞共刺激受体的背景下所含有的CD28蛋白。表述“细胞表面表达的CD28”包含在细胞表面上作为同源二聚体的一部分表达的CD28蛋白。“细胞表面表达的CD28”可以包括在通常表达CD28蛋白的细胞表面上表达的CD28蛋白或由其组成。可替代地,“细胞表面表达的CD28”可以包括在这样的细胞的表面上表达的CD28蛋白或由其组成:所述细胞通常在其表面上不表达人CD28,但已被人工工程化成在其表面上表达CD28。
如本文所使用的,表述“抗CD28抗体”包含具有单一特异性的单价抗体以及包括结合CD28的第一臂和结合第二(靶)抗原的第二臂的双特异性抗体,其中抗CD28臂包括如本文表1中所示的HCVR/LCVR或CDR序列中的任何一个。抗CD28双特异性抗体的实例在本文的其它地方描述。术语“抗原结合分子”包含抗体和抗体的抗原结合片段,包含例如双特异性抗体。
如本文所使用的,除非指定为来自非人物种(例如,“小鼠CD22”、“猴CD22”等),否则术语“CD22”是指人CD22蛋白。人CD22蛋白具有如登录号CAA42006中所示的氨基酸序列。带有myc myc六组氨酸标签(“六组氨酸”被公开为SEQ ID NO:60)的重组人CD22 ecto(D20-R687)的序列在登录号NP_001762.2中示出并且还被示为SEQ ID NO:50。hCD22胞外结构域(D20-R687).hFc也可以从R&D系统公司(R&D Systems)购买,目录号为1968-SL-050。
如本文所使用的,“结合CD22的抗体”或“抗CD22抗体”包含可以结合可溶性CD22和/或细胞表面表达的CD22的抗体和其抗原结合片段。可溶性CD22包含天然CD22蛋白以及缺乏跨膜结构域或以其它方式与细胞膜未缔合的重组CD22蛋白变体,例如CD22构建体。
如本文所使用的,表述“抗CD22抗体”包含具有单一特异性的单价抗体以及包括结合CD22的第一臂和结合第二(靶)抗原的第二臂的双特异性抗体,其中抗CD22臂包括如本文表1中所示的HCVR/LCVR或CDR序列中的任何一个。抗CD22双特异性抗体的实例在本文的其它地方描述。术语“抗原结合分子”包含抗体和抗体的抗原结合片段,包含例如双特异性抗体。
术语“抗原结合分子”包含抗体和抗体的抗原结合片段,包含例如双特异性抗体。
如本文所使用的,术语“抗体”意指包括与特定抗原(例如,CD28)特异性地结合或相互作用的至少一个互补决定区(CDR)的任何抗原结合分子或分子复合物。术语“抗体”包含免疫球蛋白分子以及其多聚体(例如,IgM),所述免疫球蛋白分子包括通过二硫键相互连接的两条重(H)链和两条轻(L)链四条多肽链。每条重链包括重链可变区(在本文中缩写为HCVR或VH)和重链恒定区。重链恒定区包括三个结构域C
如本文所使用的,术语“抗体”还包含完整抗体分子的抗原结合片段。如本文所使用的,术语抗体的“抗原结合部分”、抗体的“抗原结合片段”等包含特异性地结合抗原以形成复合物的任何天然存在的、可酶促获得的、合成的或基因工程化的多肽或糖蛋白。抗体的抗原结合片段可以例如使用任何合适的标准技术衍生自完整抗体分子,所述标准技术如涉及操纵和表达对抗体可变结构域和任选地恒定结构域进行编码的DNA的蛋白水解消化或重组基因工程技术。此类DNA是已知的和/或易于从例如商业来源、DNA文库(包含例如噬菌体-抗体文库)获得或可以合成。可以以化学方式或通过使用分子生物学技术对DNA进行测序和操纵,例如,以将一个或多个可变结构域和/或恒定结构域布置成合适的构型或引入密码子、产生半胱氨酸残基、修饰、添加或缺失氨基酸等。
抗原结合片段的非限制性实例包含:(i)Fab片段;(ii)F(ab')2片段;(iii)Fd片段;(iv)Fv片段;(v)单链Fv(scFv)分子;(vi)dAb片段;以及(vii)由模拟抗体的高变区的氨基酸残基组成的最小识别单位(例如,分离的互补决定区(CDR),如CDR3肽)或受约束的FR3-CDR3-FR4肽。其它工程化分子如结构域特异性抗体、单结构域抗体、结构域缺失抗体、嵌合抗体、CDR移植抗体、双抗体、三抗体、四抗体、微抗体、纳米抗体(例如,单价纳米抗体、二价纳米抗体等)、小型模块化免疫药物(SMIP)和鲨鱼可变IgNAR结构域也涵盖在如本文所使用的表述“抗原结合片段”内。
抗体的抗原结合片段通常包括至少一个可变结构域。可变结构域可以具有任何大小或氨基酸组成并且通常包括与一个或多个框架序列相邻或同框的至少一个CDR。在具有与V
在某些实施例中,抗体的抗原结合片段可以含有至少一个与至少一个恒定结构域共价连接的可变结构域。可以在本发明的抗体的抗原结合片段内发现的可变结构域和恒定结构域的非限制性示例性构型包含:(i)V
与完整抗体分子一样,抗原结合片段可以是单特异性的或多特异性的(例如,双特异性的)。抗体的多特异性抗原结合片段通常包括至少两个不同的可变结构域,其中每个可变结构域能够与单独的抗原或同一抗原上的不同表位特异性地结合。可以使用本领域可用的常规技术将任何多特异性抗体形式(包含本文公开的示例性双特异性抗体形式)适配成用于在本发明的抗体的抗原结合片段的背景下使用。
本发明的抗体可以通过补体依赖性细胞毒性(CDC)或抗体依赖性细胞介导的细胞毒性(ADCC)起作用。“补体依赖性细胞毒性”(CDC)是指在补体存在的情况下本发明的抗体对表达抗原的细胞的裂解。“抗体依赖性细胞介导的细胞毒性”(ADCC)是指这样的细胞介导的反应:其中表达Fc受体(FcR)的非特异性细胞毒性细胞(例如,自然杀伤(NK)细胞、嗜中性粒细胞和巨噬细胞)识别靶细胞上的结合抗体并且由此导致靶细胞的裂解。可以使用本领域熟知和可用的测定测量CDC和ADCC。(参见例如,美国专利第5,500,362号和第5,821,337号以及Clynes等人,(1998),《美国国家科学院院刊(Proc.Natl.Acad.Sci.(USA))》95:652-656)。抗体的恒定区对于抗体固定补体和介导细胞依赖性细胞毒性的能力是重要的。因此,可以基于抗体是否需要介导细胞毒性来选择抗体的同种型。
在本发明的某些实施例中,本发明的抗CD28抗体和/或抗CD22抗体(单特异性或双特异性)是人抗体。如本文所使用的,术语“人抗体”旨在包含具有衍生自人种系免疫球蛋白序列的可变区和恒定区的抗体。本发明的人抗体可以包含并非由人种系免疫球蛋白序列编码的氨基酸残基(例如,通过体外随机或位点特异性诱变或通过体内体细胞突变而引入的突变),例如在CDR中,特别是在CDR3中。然而,如本文所使用的,术语“人抗体”不旨在包含其中衍生自如小鼠等另一种哺乳动物物种的种系的CDR序列已经移植到人框架序列上的抗体。
在一些实施例中,本发明的抗体可以是重组人抗体。如本文所使用的,术语“重组人抗体”旨在包含通过重组方式制备、表达、产生或分离的所有人抗体,如使用转染到宿主细胞内的重组表达载体表达的抗体(在下文中进一步描述)、从重组的组合人抗体文库中分离的抗体(在下文中进一步描述)、从人免疫球蛋白基因转基因动物(例如,小鼠)中分离的抗体(参见例如,Taylor等人,(1992),《核酸研究(Nucl.Acids Res.)》,20:6287-6295)或通过任何其它方式制备、表达、产生或分离的抗体,所述其它方式涉及将人免疫球蛋白基因序列剪接到其它DNA序列。此类重组人抗体具有衍生自人种系免疫球蛋白序列的可变区和恒定区。然而,在某些实施例中,对此类重组人抗体进行体外诱变(或当使用人Ig序列转基因动物时,进行体内体细胞诱变),并且因此重组抗体的V
人抗体可以以与铰链异质性相关的两种形式存在。在一种形式中,免疫球蛋白分子包括约150-160kDa的稳定四链构建体,其中二聚体通过链间重链二硫键保持在一起。在第二种形式中,二聚体不通过链间二硫键连接,并且形成约75-80kDa的分子,所述分子由共价偶联的轻链和重链(半抗体)构成。即使在亲和纯化之后,这些形式也极难分离。
在各种完整IgG同种型中出现第二种形式的频率是由于但不限于与抗体的铰链区同种型相关的结构差异。人IgG4铰链的铰链区中的单个氨基酸取代可以将第二种形式的出现显著地降低(Angal等人,(1993),《分子免疫学(Molecular Immunology)》30:105)到通常使用人IgG1铰链所观察到的水平。本发明涵盖在铰链区、CH2区或CH3区具有一个或多个突变的抗体,所述突变可能是例如在生产中所期望的,以便提高期望的抗体形式的产率。
本发明的抗体可以是分离的抗体。如本文所使用的,“分离的抗体”意指已经从其天然环境的至少一种组分中鉴定和分离和/或回收的抗体。例如,已经从生物体的至少一种组分或从天然存在或天然产生抗体的组织或细胞中分离或去除的抗体是用于本发明的目的的“分离的抗体”。分离的抗体还包含重组细胞内的原位抗体。分离的抗体是已经进行至少一个纯化或分离步骤的抗体。根据某些实施例,分离的抗体可以基本上不含其它细胞物质和/或化学品。
本发明还包含结合CD28和/或CD22的单臂抗体。如本文所使用的,“单臂抗体”意指包括单抗体重链和单抗体轻链的抗原结合分子。本发明的单臂抗体可以包括表1中所示的HCVR/LCVR或CDR氨基酸序列中的任何一个。
如与衍生出抗原结合蛋白或抗原结合结构域的对应种系序列相比,本文的抗CD28抗体和/或抗CD22抗体或其抗原结合结构域可以在重链和轻链可变结构域的框架和/或CDR区中包括一个或多个氨基酸取代、插入和/或缺失。通过将本文所公开的氨基酸序列与可从例如公共抗体序列数据库获得的种系序列进行比较,可以容易地确定此类突变。本发明包含衍生自本文公开的氨基酸序列中的任何一个的抗体和其抗原结合结构域,其中一个或多个框架和/或CDR区内的一个或多个氨基酸突变为衍生出抗体的种系序列的一个或多个对应的残基或突变为另一个人种系序列的一个或多个对应的残基或突变为一个或多个对应的种系残基的保守氨基酸取代(此类序列变化在本文中统称为“种系突变”)。从本文公开的重链和轻链可变区序列开始,本领域普通技术人员可以容易地产生包括一个或多个单独种系突变或其组合的许多抗体和抗原结合片段。在某些实施例中,V
本发明还包含包括本文公开的HCVR、LCVR和/或CDR氨基酸序列中的任何一个的变体的抗CD28抗体和/或抗CD22抗体和抗原结合分子。包含在本发明的此方面内的示例性变体包含具有一个或多个保守取代的本文公开的HCVR、LCVR和/或CDR氨基酸序列中的任何一个的变体。例如,本发明包含具有HCVR、LCVR和/或CDR氨基酸序列的抗CD28抗体和抗原结合分子,相对于如本文表6中所示的HCVR、LCVR和/或CDR氨基酸序列中的任何一个,所述HCVR、LCVR和/或CDR氨基酸序列具有例如10个或更少、8个或更少、6个或更少、4个或更少等的保守氨基酸取代。
术语“表位”是指与称为互补位的抗体分子的可变区中的特定抗原结合位点相互作用的抗原决定簇。单个抗原可以具有多于一个表位。因此,不同的抗体可以与抗原上的不同区域结合并且可以具有不同的生物学效应。表位可以是构象的或线性的。构象表位通过来自线性多肽链的不同区段的空间上并列的氨基酸产生。线性表位是由多肽链中的相邻氨基酸残基产生的表位。在某些情况下,表位可以包含抗原上的糖、磷酰基或磺酰基的部分。
当提及核酸或其片段时,术语“基本同一性”或“基本上相同”指示,当通过适当的核苷酸插入或缺失与另一个核酸(或其互补链)最佳比对时,核苷酸序列同一性为核苷酸碱基的至少约95%,并且更优选地,至少约96%、97%、98%或99%,如通过如FASTA、BLAST或Gap等任何众所周知的序列同一性算法测量的,如下文所讨论的。在某些情况下,与参考核酸分子具有基本同一性的核酸分子可以对具有与由参考核酸分子编码的多肽相同或基本上类似的氨基酸序列的多肽进行编码。
当应用于多肽时,术语“基本相似性”或“基本上类似”意味着当如通过使用默认空位权重的程序GAP或BESTFIT最佳比对时,两个肽序列共享至少95%的序列同一性,甚至更优选地至少98%或99%的序列同一性。优选地,不相同的残基位置因保守氨基酸取代而不同。“保守氨基酸取代”是氨基酸残基被具有化学性质(例如,电荷或疏水性)类似的侧链(R基团)的另一个氨基酸残基取代的氨基酸取代。通常,保守氨基酸取代不会实质上改变蛋白质的功能性质。在两个或更多个氨基酸序列因保守取代而彼此不同的情况下,可以向上调整序列同一性百分比或相似性程度,以校正取代的保守本质。用于作出此调整的方法是本领域技术人员所熟知的。参见例如,Pearson(1994)《分子生物学方法(MethodsMol.BioI.)》24:307-331。具有化学性质类似的侧链的氨基酸基团的实例包含(1)脂肪族侧链:甘氨酸、丙氨酸、缬氨酸、亮氨酸和异亮氨酸;(2)脂肪族羟基侧链:丝氨酸和苏氨酸;(3)含酰胺的侧链:天冬酰胺和谷氨酰胺;(4)芳香族侧链:苯丙氨酸、酪氨酸和色氨酸;(5)碱性侧链:赖氨酸、精氨酸和组氨酸;(6)酸性侧链:天冬氨酸和谷氨酸;以及(7)含硫侧链是半胱氨酸和蛋氨酸。优选的保守氨基酸取代基是:缬氨酸-亮氨酸-异亮氨酸、苯丙氨酸-酪氨酸、赖氨酸-精氨酸、丙氨酸-缬氨酸、谷氨酸-天冬氨酸和天冬酰胺-谷氨酰胺。可替代地,保守替代是在Gonnet等人(1992),《科学(Science)》256:1443-1445中公开的PAM250对数似然矩阵中具有正值的任何变化。“适度保守”替代是在PAM250对数似然矩阵中具有非负值的任何变化。
通常使用序列分析软件测量也称为序列同一性的多肽的序列相似性。蛋白质分析软件使用分配给包含保守氨基酸取代在内的各种取代、缺失和其它修饰的相似性度量来匹配类似的序列。例如,GCG软件含有如Gap和Bestfit等程序,所述程序可以与默认参数一起使用,以确定紧密相关的多肽如来自不同生物体物种的同源多肽之间或野生型蛋白质与其突变蛋白之间的序列同源性或序列同一性。参见例如6.1版GCG。还可以使用使用默认或推荐的参数的FASTA(6.1版GCG中的程序)来比较多肽序列。FASTA(例如,FASTA2和FASTA3)提供了询问序列与搜索序列之间的最佳重叠区的比对和序列同一性百分比(Pearson(2000)同上)。当比较本发明的序列与含有来自不同生物体的大量序列的数据库时,另一个优选的算法是使用默认参数的计算机程序BLAST,尤其是BLASTP或TBLASTN。参见例如,Altschul等人(1990)《分子生物学杂志(J.Mol.BioI.)》215:403-410以及Altschul等人(1997)《核酸研究》25:3389-402。
术语“细胞增殖性病症”和“增殖性病症”是指与从利用本发明的抗CD28/抗CD22双特异性抗原结合分子或方法的治疗中受益的一定程度的异常细胞增殖相关的病症。这包含慢性和急性病症,包含使哺乳动物易患所讨论病症的那些病理病状。在一个实施例中,细胞增殖性病症是在哺乳动物中通常以不受调节的细胞生长/增殖为特征的生理病状癌症。
如本文所使用的,“肿瘤”是指所有赘生性细胞生长和增殖(无论是恶性的还是良性的)以及所有癌前和癌性细胞和组织。术语“癌症”、“癌性”、“细胞增殖性病症”、“增殖性病症”和“肿瘤”如本文所引用的并不相互排斥。
“B细胞增殖性病症”包含霍奇金淋巴瘤(Hodgkin's lymphoma)、非霍奇金淋巴瘤(NHL),如侵袭性NHL、复发侵袭性NHL、低度/滤泡性NHL、小淋巴细胞(SL)NHL、中度/滤泡性NHL、中度弥漫性NHL、高度免疫母细胞性NHL、高度淋巴母细胞性NHL、高度小型非切割细胞NHL、大体积疾病NHL、包含复发性惰性NHL和利妥昔单抗难治性惰性NHL的惰性NHL;难治性NH、难治性惰性NHL、套细胞淋巴瘤、艾滋病相关淋巴瘤和华氏巨球蛋白血症(Waldenstrom's Macroglobulinemia)、淋巴细胞显性霍奇金疾病(LPHD)、小淋巴细胞淋巴瘤(SLL)、慢性淋巴细胞白血病(CLL);白血病,包含急性淋巴母细胞白血病(ALL)、慢性淋巴细胞白血病(CLL)、毛细胞白血病、慢性成髓细胞白血病;以及其它血液系统恶性肿瘤。
如本文所使用的,术语“非霍奇金淋巴瘤”或“NHL”是指淋巴系统的癌症而不是霍奇金淋巴瘤。霍奇金淋巴瘤通常可以与非霍奇金淋巴瘤区别开来:霍奇金淋巴瘤中存在里德-斯泰伯格氏细胞(Reed-Sternberg cell),而非霍奇金淋巴瘤中不存在所述细胞。如本文所使用的术语所涵盖的非霍奇金淋巴瘤的实例包含根据本领域已知的分类方案由本领域的技术人员(例如,肿瘤学家或病理学家)如此识别的任何淋巴瘤,如修订的欧美淋巴瘤(Revised European-American Lymphoma,REAL)方案,如《临床血液学彩色图谱(ColorAtlas of Clinical Hematology)》(第3版),A.Victor Hoffbrand和John E.Pettit(编)(哈考特出版社公司(Harcourt Publishers Ltd.),2000)所描述的。具体地,参见图11.57、11.58和11.59中的列表。更具体的实例包含但不限于复发性或难治性NHL、前线低度NHL、III/IV期NHL、化疗耐药NHL、前体B淋巴母细胞白血病和/或淋巴瘤、小淋巴细胞淋巴瘤、B细胞慢性淋巴细胞白血病和/或幼淋巴细胞白血病和/或小淋巴细胞淋巴瘤、B细胞幼淋巴细胞淋巴瘤、免疫细胞瘤和/或淋巴浆细胞性淋巴瘤、淋巴浆细胞性淋巴瘤、边缘区B细胞淋巴瘤、脾边缘区淋巴瘤、结外边缘区-MALT淋巴瘤、结节边缘区淋巴瘤、毛细胞白血病、浆细胞瘤和/或浆细胞骨髓瘤、低度/滤泡性淋巴瘤、中度/滤泡性NHL、套细胞淋巴瘤、滤泡中心淋巴瘤(滤泡性)、中度弥漫性NHL、弥漫性大B细胞淋巴瘤、侵袭性NHL(包含侵袭性前线NHL和侵袭性复发NHL)、自体干细胞植入后复发或难治的NHL、原发性纵隔大B细胞淋巴瘤、原发性渗出性淋巴瘤、高度免疫母细胞性NHL、高度淋巴母细胞性NHL、高度小型非切割细胞NHL、大体积疾病NHL、伯基特淋巴瘤(Burkitt's lymphoma)、前体(外周)大颗粒淋巴细胞白血病、蕈样肉芽肿病和/或塞扎里综合征(Sezary syndrome)、皮肤(表皮)淋巴瘤、间变性大细胞淋巴瘤、血管中心性淋巴瘤。
双特异性抗原结合分子
本发明的抗体可以是单特异性的、双特异性的或多特异性的。多特异性抗体可以对一个靶多肽的不同表位具有特异性或者可以含有对多于一个靶多肽具有特异性的抗原结合结构域。参见例如,Tutt等人,1991,《免疫学期刊(J.Immunol.)》147:60-69;Kufer等人,2004,《生物技术趋势(Trends Biotechnol.)》22:238-244。本发明的抗CD28抗体和/或抗CD22抗体可以与另一种功能分子例如另一种肽或蛋白质连接或共表达。例如,抗体或其片段可以与一个或多个其它分子实体如另一个抗体或抗体片段功能性地连接(例如,通过化学偶联、遗传融合、非共价缔合或以其它方式),以产生具有第二结合特异性的双特异性或多特异性抗体。
本文使用的表述“抗CD28抗体”和/或“抗CD22抗体”旨在包含单特异性抗CD28抗体和/或单特异性抗CD22抗体两者以及包括CD28结合臂或CD22结合臂和结合靶抗原的臂的双特异性抗体。因此,本发明包含双特异性抗体,其中免疫球蛋白的一个臂结合人CD28或CD22,并且免疫球蛋白的另一个臂对靶抗原具有特异性。CD28或CD22双特异性抗体的另一个臂所结合的靶抗原可以是在细胞、组织、器官、微生物或病毒上或附近表达的任何抗原,期望针对所述抗原的靶向性免疫应答。CD28结合臂可以包括如本文的表1中所示的HCVR/LCVR或CDR氨基酸序列中的任何一个。CD22结合臂可以包括如本文表1中所示的HCVR/LCVR或CDR氨基酸序列中的任何一个。在某些实施例中,CD28结合臂结合人CD28并诱导人T细胞增殖。
在本发明的其中抗体的一个臂结合CD28并且另一个臂结合靶抗原的双特异性抗体的背景下,靶抗原可以是肿瘤相关抗原,如CD22。
根据某些示例性实施例,本发明包含特异性地结合CD28和CD22的双特异性抗原结合分子。此类分子在本文中可以被称为例如“抗CD28/抗CD22”或“抗CD28xCD22”或“CD28xCD22”或“抗CD22/抗CD28”或“抗CD22xCD28”或“CD22xCD28”双特异性分子或“αCD22x αCD28”或“αCD28 x αCD22”或其它类似的术语。
根据某些示例性实施例,双特异性抗原结合分子(例如,双特异性抗体)可以具有效应臂和靶向臂。效应臂可以是与效应细胞(例如,T细胞)上的抗原结合的第一抗原结合结构域(例如,抗CD28抗体)。靶向臂可以是与靶细胞(例如,肿瘤细胞)上的抗原结合的第二抗原结合结构域(例如,抗CD22抗体)。根据某些示例性实施例,效应臂与CD28结合,并且靶向臂与CD22结合。双特异性抗CD28/CD22可以向效应细胞(例如,T细胞)提供共刺激信号。效应臂在没有聚类的情况下没有刺激T细胞的作用。单独的效应臂几乎没有刺激T细胞的作用,除非与靶向臂组合。肿瘤靶向臂可以具有不完善的肿瘤特异性。作为靶向臂的靶标的抗原(例如,CD22)可以在肿瘤细胞的一部分上表达。可以通过与同抗CD3双特异性抗原结合分子(例如,抗CD3/CD20双特异性抗体)的组合重叠来增加肿瘤靶向臂的特异性。
如本文所使用的,表述“抗原结合分子”意指包括至少一个互补决定区(CDR)或由其组成的蛋白质、多肽或分子复合物,所述互补决定区单独地或与一个或多个另外的CDR和/或框架区(FR)组合地与特定抗原特异性地结合。在某些实施例中,抗原结合分子是抗体或抗体的片段,如在本文其它地方定义的那些术语。
如本文所使用的,表述“双特异性抗原结合分子”意指包括至少第一抗原结合结构域和第二抗原结合结构域的蛋白质、多肽或分子复合物。双特异性抗原结合分子内的每个抗原结合结构域包括至少一个CDR,所述至少一个CDR单独地或与一个或多个另外的CDR和/或FR组合地与特定抗原特异性地结合。在本发明的背景下,第一抗原结合结构域特异性地结合第一抗原(例如,CD28),并且第二抗原结合结构域特异性地结合第二不同抗原(例如,CD22)。
在本发明的某些示例性实施例中,双特异性抗原结合分子是双特异性抗体。双特异性抗体的每个抗原结合结构域包括重链可变结构域(HCVR)和轻链可变结构域(LCVR)。在包括第一抗原结合结构域和第二抗原结合结构域的双特异性抗原结合分子(例如,双特异性抗体)的背景下,第一抗原结合结构域的CDR可以用前缀“D1”表示,并且第二抗原结合结构域的CDR可以用前缀“D2”表示。因此,第一抗原结合结构域的CDR在本文中可以被称为D1-HCDR1、D1-HCDR2和D1-HCDR3;并且第二抗原结合结构域的CDR在本文中可以被称为D2-HCDR1、D2-HCDR2和D2-HCDR3。
第一抗原结合结构域和第二抗原结合结构域可以彼此直接或间接连接以形成本发明的双特异性抗原结合分子。可替代地,第一抗原结合结构域和第二抗原结合结构域可以各自与单独的多聚化结构域连接。一个多聚化结构域与另一个多聚化结构域的缔合促进了两个抗原结合结构域之间的缔合,由此形成双特异性抗原结合分子。如本文所使用的,“多聚化结构域”是能够与具有相同或类似结构或构成的第二多聚化结构域缔合的任何大分子、蛋白质、多肽、肽或氨基酸。例如,多聚化结构域可以是包括免疫球蛋白C
本发明的双特异性抗原结合分子通常包括两个多聚化结构域,例如,各自独立地是单独的抗体重链的一部分的两个Fc结构域。第一多聚化结构域和第二多聚化结构域可以是同一IgG同种型,例如IgG1/lgG1、IgG2/lgG2、IgG4/lgG4。可替代地,第一多聚化结构域和第二多聚化结构域可以是不同的IgG同种型,例如IgG1/lgG2、IgG1/lgG4、IgG2/lgG4等。
在某些实施例中,多聚化结构域是Fc片段或含有至少一个半胱氨酸残基的长度为1到约200个氨基酸的氨基酸序列。在其它实施例中,多聚化结构域是半胱氨酸残基或含半胱氨酸的短肽。其它多聚化结构域包含包括亮氨酸拉链、螺旋环基序或卷曲螺旋基序或者由其组成的肽或多肽。
可以使用任何双特异性抗体形式或技术来制备本发明的双特异性抗原结合分子。例如,可以将具有第一抗原结合特异性的抗体或其片段与一个或多个其它分子实体如具有第二抗原结合特异性的另一种抗体或抗体片段等功能性地连接(例如,通过化学偶联、遗传融合、非共价缔合或以其它方式),以产生双特异性抗原结合分子。可以在本发明的背景下使用的具体示例性双特异性形式包含但不限于例如,基于scFv的或双抗体双特异性形式、IgG-scFv融合、双可变结构域(OVO)-Ig、四杂交瘤(Quadroma)、结节入孔(knobs-into-holes)、共同轻链(例如,具有结节入孔的共同轻链等)、CrossMab、CrossFab、(SEED)body、亮氨酸拉链、Ouobody、IgG1/lgG2、双作用Fab(OAF)-IgG以及Mab
在本发明的双特异性抗原结合分子的背景下,与野生型、天然存在形式的Fc结构域相比,多聚化结构域例如Fc结构域可以包括一个或多个氨基酸变化(例如,插入、缺失或取代)。例如,本发明包含双特异性抗原结合分子,其在Fc结构域中包括一个或多个修饰,从而产生在Fc与FcRn之间具有经修饰的结合相互作用(例如,增强或减弱)的经修饰的Fc结构域。在一个实施例中,双特异性抗原结合分子包括C
本发明还包含双特异性抗原结合分子,其包括第一C
在某些实施例中,Fc结构域可以是嵌合的,从而组合衍生自多于一种免疫球蛋白同种型的Fc序列。例如,嵌合Fc结构域可以包括衍生自人IgG1、人IgG2或人IgG4 C
序列变体
如与衍生出单独抗原结合结构域的对应种系序列相比,本发明的抗体和双特异性抗原结合分子可以在重链和轻链可变结构域的框架和/或CDR区中包括一个或多个氨基酸取代、插入和/或缺失。通过将本文公开的氨基酸序列与可从例如公共抗体序列数据库获得的种系序列进行比较,可以容易地确定此类突变。本发明的抗原结合分子可以包括衍生自本文公开的示例性氨基酸序列中的任何一个的抗原结合片段,其中一个或多个框架和/或CDR区内的一个或多个氨基酸突变为衍生出抗体的种系序列的一个或多个对应的残基或突变为另一个人种系序列的一个或多个对应的残基或突变为一个或多个对应的种系残基的保守氨基酸取代(此类序列变化在本文中统称为“种系突变”)。从本文公开的重链和轻链可变区序列开始,本领域普通技术人员可以容易地产生包括一个或多个单独种系突变或其组合的许多抗体和抗原结合片段。在某些实施例中,V
本发明还包含抗原结合分子,其中一个或两个抗原结合结构域包括具有一个或多个保守取代的本文公开的HCVR、LCVR和/或CDR氨基酸序列中的任何一个的变体。例如,本发明包含抗原结合分子,其包括具有HCVR、LCVR和/或CDR氨基酸序列的抗原结合结构域,相对于本文公开的HCVR、LCVR和/或CDR氨基酸序列中的任何一个,所述HCVR、LCVR和/或CDR氨基酸序列具有例如10个或更少、8个或更少、6个或更少、4个或更少等的保守氨基酸取代。“保守氨基酸取代”是氨基酸残基被具有化学性质(例如,电荷或疏水性)类似的侧链(R基团)的另一个氨基酸残基取代的氨基酸取代。通常,保守氨基酸取代不会实质上改变蛋白质的功能性质。具有化学性质类似的侧链的氨基酸基团的实例包含(1)脂肪族侧链:甘氨酸、丙氨酸、缬氨酸、亮氨酸和异亮氨酸;(2)脂肪族羟基侧链:丝氨酸和苏氨酸;(3)含酰胺的侧链:天冬酰胺和谷氨酰胺;(4)芳香族侧链:苯丙氨酸、酪氨酸和色氨酸;(5)碱性侧链:赖氨酸、精氨酸和组氨酸;(6)酸性侧链:天冬氨酸和谷氨酸;以及(7)含硫侧链是半胱氨酸和蛋氨酸。优选的保守氨基酸取代基是:缬氨酸-亮氨酸-异亮氨酸、苯丙氨酸-酪氨酸、赖氨酸-精氨酸、丙氨酸-缬氨酸、谷氨酸-天冬氨酸和天冬酰胺-谷氨酰胺。可替代地,保守替代是在以下文献中公开的PAM250对数似然矩阵中具有正值的任何变化:Gonnet等人(1992)《科学》,256:1443-1445。“适度保守”替代是在PAM250对数似然矩阵中具有非负值的任何变化。
本发明还包含抗原结合分子,其包括具有与本文公开的HCVR、LCVR和/或CDR氨基酸序列中的任何一个基本相同的HCVR、LCVR和/或CDR氨基酸序列的抗原结合结构域。当提及氨基酸序列时,术语“基本同一性”或“基本上相同”意味着当如通过使用默认空位权重的程序GAP或BESTFIT进行最佳比对时,两个氨基酸序列共享至少95%的序列同一性,甚至更优选地,至少98%或99%的序列同一性。优选地,不相同的残基位置因保守氨基酸取代而不同。在两个或更多个氨基酸序列因保守取代而彼此不同的情况下,可以向上调整序列同一性百分比或相似性程度,以校正取代的保守本质。用于作出此调整的方法是本领域技术人员所熟知的。参见例如,Pearson(1994)《分子生物学方法)》24:307-331。
通常使用序列分析软件测量也称为序列同一性的多肽的序列相似性。蛋白质分析软件使用分配给包含保守氨基酸取代在内的各种取代、缺失和其它修饰的相似性度量来匹配类似的序列。例如,GCG软件含有如Gap和Bestfit等程序,所述程序可以与默认参数一起使用,以确定紧密相关的多肽如来自不同生物体物种的同源多肽之间或野生型蛋白质与其突变蛋白之间的序列同源性或序列同一性。参见例如6.1版GCG。还可以使用使用默认或推荐的参数的FASTA(6.1版GCG中的程序)来比较多肽序列。FASTA(例如,FASTA2和FASTA3)提供了询问序列与搜索序列之间的最佳重叠区的比对和序列同一性百分比(Pearson(2000)同上)。当比较本发明的序列与含有来自不同生物体的大量序列的数据库时,另一个优选的算法是使用默认参数的计算机程序BLAST,尤其是BLASTP或TBLASTN。参见例如,Altschul等人(1990)《分子生物学杂志》215:403-410以及Altschul等人(1997)《核酸研究》25:3389-402。
pH依赖性结合
本发明包含具有pH依赖性结合特性的抗CD28/抗CD22双特异性抗原结合分子。例如,如与中性pH相比,本发明的抗CD28抗体在酸性pH下可以表现出与CD28的结合降低。可替代地,如与中性pH相比,本发明的抗CD22抗体在酸性pH下可以表现出与CD22的结合增强。表述“酸性pH”包含小于约6.2例如约6.0、5.95、5.9、5.85、5.8、5.75、5.7、5.65、5.6、5.55、5.5、5.45、5.4、5.35、5.3、5.25、5.2、5.15、5.1、5.05、5.0或更小的pH值。如本文所使用的,表述“中性pH”意指pH为约7.0到约7.4。表述“中性pH”包含约7.0、7.05、7.1、7.15、7.2、7.25、7.3、7.35和7.4的pH值。
在某些情况下,“如与中性pH相比,在酸性pH下……结合降低”是就在酸性pH下抗体与其抗原结合的K
具有pH依赖性结合特性的抗体可以例如通过筛选如与中性pH相比在酸性pH下与特定抗原的结合降低(或增强)的抗体群来获得。另外,氨基酸水平上的抗原结合结构域的修饰可以产生具有pH依赖性特性的抗体。例如,通过用组氨酸残基取代抗原结合结构域(例如,在CDR内)的一个或多个氨基酸,可以获得在相对于中性pH的酸性pH下与抗原的结合降低的抗体。
包括Fc变体的抗体
根据本发明的某些实施例,提供了包括Fc结构域的抗CD28/抗CD22双特异性抗原结合分子,所述Fc结构域包括例如如与中性pH相比在酸性pH下增强或减弱抗体与FcRn受体的结合的一个或多个突变。例如,本发明包含在Fc结构域的C
例如,本发明包含包括Fc结构域的抗CD28/抗CD22双特异性抗原结合分子,所述Fc结构域包括选自由以下组成的组的一个或多个突变对或突变组:250Q和248L(例如,T250Q和M248L);252Y、254T和256E(例如,M252Y、S254T和T256E);428L和434S(例如,M428L和N434S);以及433K和434F(例如,H433K和N434F)。在本发明的范围内设想了前述Fc结构域突变的所有可能的组合和本文公开的抗体可变结构域内的其它突变。
抗体和抗原结合分子的生物学特性
本发明包含以高亲和力结合人CD28和/或CD22的抗体和其抗原结合片段。本发明还包含以中等或低亲和力结合人CD28和/或CD22的抗体和其抗原结合片段,这取决于治疗背景和所期望的特定靶向性质。例如,在其中一个臂结合CD28并且另一个臂结合靶抗原(例如,CD22)的双特异性抗原结合分子的背景下,可能令人期望的是,靶抗原结合臂以高亲和力结合靶抗原,而抗CD28臂仅以中等或低亲和力结合CD28。以这种方式,可以实现抗原结合分子对表达靶抗原的细胞的优先靶向,同时避免一般/非靶向性CD28结合以及随之产生的与之相关的不良副作用。
根据某些实施例,本发明包含抗体和抗体的抗原结合片段,其以小于约15nM的K
根据某些实施例,本发明包含抗体和抗体的抗原结合片段,其以小于约60μM的K
根据某些实施例,本发明包含抗体和抗体的抗原结合片段,其以小于约45μM的K
本发明还包含结合人CD22的抗体和其抗原结合片段,其中解离半衰期(t
本发明还包含结合猴CD22的抗体和其抗原结合片段,其中解离半衰期(t
本发明还包含结合人CD28的抗体和其抗原结合片段,其中解离半衰期(t
本发明还包含能够与人CD28以及人和猴CD22结合的双特异性抗原结合分子(例如,双特异性抗体)。根据某些实施例,本发明的双特异性抗原结合分子与表达CD28和/或CD22的细胞特异性地相互作用。可以通过荧光活化细胞分选(FACS)来评估双特异性抗原结合分子结合表达CD28和/或CD22的细胞的程度,如本文实例6中所展示的。例如,本发明包含特异性地结合表达CD28但不表达CD22(例如,T细胞)的人细胞系或猕猴细胞以及表达CD22但不表达CD28(例如,B细胞或Nalm6细胞)的人细胞系或猕猴细胞的双特异性抗原结合分子。本发明包含结合上述细胞和细胞系中的任何一个的双特异性抗原结合分子,其中EC
本发明包含能够与人CD28和/或人CD22结合的双特异性抗原结合分子(例如,双特异性抗体)。根据某些实施例,本发明的双特异性抗原结合分子与表达CD28和/或CD22的细胞特异性地相互作用。可以通过流式细胞术来评估双特异性抗原结合分子结合表达CD28和/或CD22的细胞的程度,如本文实例7中所展示的。例如,本发明包含特异性地结合表达CD28但不表达CD22(例如,T细胞)的人细胞以及表达CD22但不表达CD28(例如,用人CD22转导的HEK293细胞以及被基因修饰以缺失CD80和CD86的Raji B细胞)的人细胞系的双特异性抗原结合分子。本发明包含结合上述细胞和细胞系中的任何一个的双特异性抗原结合分子,其中EC
本发明还提供了诱导或增强对肿瘤细胞进行CD20xCD3 T细胞介导的杀伤的效力的抗CD28/抗CD22双特异性抗原结合分子。例如,本发明包含诱导或增加对肿瘤细胞进行CD20xCD3 T细胞介导的杀伤的效力的抗CD28xCD22抗体,其中EC
本发明还包含抗CD28/抗CD22双特异性抗原结合分子,其表现出选自由以下组成的组的一个或多个特性:活化T细胞、诱导IL-2释放、诱导人PBMC中的CD25+上调以及增加人T细胞介导的对表达CD22的细胞系的细胞毒性(参见例如本文实例9)。本发明还包含当与结合CD20和CD3的双特异性抗体(如但不限于REGN1979)组合时增强对表达CD22的肿瘤细胞的杀伤的抗CD28/抗CD22双特异性抗原结合分子。本发明还包含当与结合PD-1的抗体(如但不限于西米普利单抗)组合时增强对表达CD22的肿瘤细胞的杀伤的抗CD28/抗CD22双特异性抗原结合分子。(参见实例10-15)。
表位作图和相关技术
本发明的抗原结合分子所结合的CD28或CD22上的表位可以由CD28蛋白或CD22蛋白的3个或更多个(例如,3个、4个、5个、6个、7个、8个、9个、10个、11个、12个、13个、14个、15个、16个、17个、18个、19个、20个或更多个)氨基酸的单个连续序列组成。可替代地,表位可以由CD28或CD22的多个非连续氨基酸(或氨基酸序列)组成。本发明的抗体可以与CD28单体内含有的氨基酸相互作用或者可以与CD28二聚体的两条不同CD28链上的氨基酸相互作用。如本文所使用的,术语“表位”是指与被称为互补位的抗体分子的可变区中的特定抗原结合位点相互作用的抗原决定簇。单个抗原可以具有多于一个表位。因此,不同的抗体可以与抗原上的不同区域结合并且可以具有不同的生物学效应。表位可以是构象的或线性的。构象表位通过来自线性多肽链的不同区段的空间上并列的氨基酸产生。线性表位是由多肽链中的相邻氨基酸残基产生的表位。在某些情况下,表位可以包含抗原上的糖、磷酰基或磺酰基的部分。
可以使用本领域普通技术人员已知的各种技术来确定抗体的抗原结合结构域是否在多肽或蛋白质内“与一个或多个氨基酸相互作用”。可以用于确定特定抗体或抗原结合结构域的表位或结合结构域的示例性技术包含例如:常规交叉阻断测定,如在以下文献中描述的常规交叉阻断测定:
本发明进一步包含抗CD28和抗CD22抗体,其与同本文所述的特定示例性抗体(例如,包括本文表6中所示的氨基酸序列中的任何一个的抗体)中的任何一个相同的表位结合。
根据某些实施例,本发明提供了结合人CD22上的表位的抗体和抗体的抗原结合片段,所述表位包括SEQ ID NO:34、SEQ ID NO:35和/或SEQ ID NO:36的一个或多个氨基酸,如通过实例3和4中所示的质谱法检测到的氢氘交换确定的。
同样,本发明还包含抗CD28和/或抗CD22抗体,其与本文所述的特定示例性抗体(例如,包括本文表6中所示的氨基酸序列中的任何一个的抗体)中的任何一个竞争与CD28和/CD22结合。
本发明还包含双特异性抗原结合分子,其包括特异性地结合人CD28的第一抗原结合结构域和特异性地结合人CD22的第二抗原结合片段,其中第一抗原结合结构域与CD28上的与本文所描述的特定示例性CD28特异性抗原结合结构域中的任何一个相同的表位结合,和/或其中第二抗原结合结构域与CD22上的与本文所描述的特定示例性CD22特异性抗原结合结构域中的任何一个相同的表位结合。
同样,本发明还包含双特异性抗原结合分子,其包括特异性地结合人CD28的第一抗原结合结构域和特异性地结合人CD22的第二抗原结合片段,其中第一抗原结合结构域与本文所描述的特定示例性CD28特异性抗原结合结构域中的任何一个竞争与CD28结合,和/或其中第二抗原结合结构域与本文所描述的特定示例性CD22特异性抗原结合结构域中的任何一个竞争与CD22结合.
通过使用本领域中已知的常规方法,可以容易地确定特定的抗原结合分子(例如,抗体)或其抗原结合结构域是否与同本发明的参考抗原结合分子相同的表位结合或竞争与本发明的参考抗原结合分子结合。例如,为了确定测试抗体是否与CD28(或CD22)上的同本发明的参考双特异性抗原结合分子相同的表位结合,首先允许参考双特异性分子与CD28蛋白(或CD22蛋白)结合。接下来,评估测试抗体与CD28(或CD22)分子结合的能力。如果测试抗体在与参考双特异性抗原结合分子饱和结合后能够与CD28(或CD22)结合,则可以得出结论,测试抗体与CD28(或CD22)中的与参考双特异性抗原结合分子不同的表位结合。另一方面,如果测试抗体在与参考双特异性抗原结合分子饱和结合后无法与CD28(或CD22)分子结合,那么测试抗体可以与CD28(或CD22)中的与本发明的参考双特异性抗原结合分子所结合的表位相同的表位结合。然后可以进行另外的常规实验(例如,肽突变和结合分析)以确认所观察到的测试抗体结合缺乏是否实际上是由于与同参考双特异性抗原结合分子相同的表位结合或者空间阻断(或另一种现象)是否是所观察到的结合的缺乏的原因。可以使用ELISA、RIA、Biacore、流式细胞术或本领域可用的任何其它定量或定性抗体结合测定执行这种实验。根据本发明的某些实施例,如果一种抗原结合蛋白的例如1倍、5倍、10倍、20倍或100倍过量抑制另一种抗原结合蛋白的至少50%,但优选地75%、90%或甚至99%的结合(如在竞争性结合测定中所测量的),则两种抗原结合蛋白与同一(或重叠)表位结合(参见例如,Junghans等人,《癌症研究(Cancer Res.)》1990:50:1495-1502)。可替代地,如果抗原中的减少或消除一种抗原结合蛋白的结合的基本上所有氨基酸突变均减少或消除另一种抗原结合蛋白的结合,则认为两种抗原结合蛋白与同一表位结合。如果减少或消除一种抗原结合蛋白的结合的氨基酸突变的仅子集减少或消除另一种抗原结合蛋白的结合,则认为两种抗原结合蛋白具有“重叠表位”。
为了确定抗体或其抗原结合结构域是否与参考抗原结合分子竞争结合,以两种朝向执行上述结合方法:在第一种朝向中,允许参考抗原结合分子在饱和条件下与CD28蛋白(或CD22蛋白)结合,然后评估测试抗体与CD28(或CD22)分子的结合。在第二种朝向中,允许测试抗体在饱和条件下与CD28(或CD22)分子结合,然后评估参考抗原结合分子与CD28(或CD22)分子的结合。如果在两种朝向上仅第一(饱和的)抗原结合分子能够与CD28(或CD22)分子结合,那么可以得出结论,测试抗体和参考抗原结合分子竞争与CD28(或CD22)结合。如本领域普通技术人员所理解的,与参考抗原结合分子竞争结合的抗体可能不一定与同参考抗体相同的表位结合,但是可以通过结合重叠或相邻表位来空间上阻断参考抗体的结合。
抗原结合结构域的制备和双特异性分子的构建
对特定抗原具有特异性的抗原结合结构域可以通过本领域已知的任何抗体生成技术来制备。一旦获得,对两种不同抗原(例如,CD28和CD22)具有特异性的两种不同的抗原结合结构域就可以相对于彼此适当地布置,以使用常规方法产生本发明的双特异性抗原结合分子。(本文其它地方提供了对可以用于构建本发明的双特异性抗原结合分子的示例性双特异性抗体形式的讨论)。在某些实施例中,本发明的多特异性抗原结合分子的单独组分(例如,重链和轻链)中的一个或多个衍生自嵌合抗体、人源化抗体或完全人抗体。用于制备此类抗体的方法是本领域熟知的。例如,可以使用VELOCIMMUNE
可以使用经基因工程化的动物来制备人双特异性抗原结合分子。例如,可以使用无法重新布置和表达内源性小鼠免疫球蛋白轻链可变序列的经基因修饰的小鼠,其中小鼠仅表达由人免疫球蛋白序列编码的一个或两个人轻链可变结构域,所述人免疫球蛋白序列与内源性小鼠κ基因座处的小鼠κ恒定基因可操作地连接。这种经遗传修饰的小鼠可以用于产生包括两条不同的重链的完全人双特异性抗原结合分子,所述重链与包括衍生自两个不同人轻链可变区基因区段中的一个的可变结构域的相同轻链缔合。(关于此工程化小鼠和其用于产生双特异性抗原结合分子的用途的详细讨论,参见例如US2011/0195454,所述文献的全部内容通过引用并入本文)。
生物等效性
本发明涵盖具有不同于所描述抗体的氨基酸序列但保留结合CD28和/或CD22的能力的氨基酸序列的抗原结合分子。与亲本序列相比,此类变体分子包括氨基酸的一个或多个添加、缺失或取代,但表现出与所描述抗原结合分子的生物活性基本上等效的生物活性。同样,本发明的对抗原结合分子进行编码的DNA序列涵盖这样的序列:与所公开的序列相比,其包括核苷酸的一个或多个添加、缺失或取代,但对与本发明的所描述抗原结合分子基本上生物等效的抗原结合分子进行编码。上文讨论了此类变体氨基酸和DNA序列的实例。
本发明包含与本文所示的示例性抗原结合分子中的任何一个生物等效的抗原结合分子。如果例如两种抗原结合蛋白或抗体是在类似的实验条件下以相同的摩尔剂量(单剂量或多剂量)施用时吸收速率和吸收程度未示出显著差异的药物等效物或药物替代物,则认为所述两种抗原结合蛋白或抗体是生物等效的。如果一些抗体在其吸收程度上等效但在其吸收速率上不等效,则认为所述抗体是等效物或药物替代物,并且可以认为所述抗体是生物等效的,因为有意的并且反映在标记上的这种吸收速率差异在例如长期使用时对于达到有效的身体药物浓度并非是必需的并且对于所研究的特定药物产品而言被认为是医学上无关紧要的。
在一个实施例中,如果两种抗原结合蛋白的安全性、纯度和效力不存在临床上有意义的差异,则所述两种抗原结合蛋白是生物等效的。
在一个实施例中,与不存在参考产品与生物产品之间的转换的持续疗法相比,如果患者可以在参考产品与生物产品之间转换一次或多次而没有预期的副作用风险增加(包含免疫原性的临床显著变化或有效性降低),则两种抗原结合蛋白是生物等效的。
在一个实施例中,如果两种抗原结合蛋白均针对一个或多个使用条件通过一种或多种共同作用机制起作用(只要这些机制是已知的),则所述两种抗原结合蛋白是生物等效的。
生物等效性可以通过体内和体外方法来证明。生物等效性度量包含例如:(a)在人或其它哺乳动物中的体内测试,其中在血液、血浆、血清或其它生物流体中测量抗体或其代谢物的浓度随时间的变化;(b)与人体内生物利用度数据相关并且可合理地预测的体外测试;(c)在人或其它哺乳动物中的体内测试,其中测量抗体(或其靶标)的适当急性药理作用随时间的变化;以及(d)在建立抗体的安全性、功效或生物利用度或生物等效性的控制良好的临床试验中。
可以通过例如对残基或序列进行各种取代或缺失非生物活性所需的末端或内部残基或序列来构建本文所示的示例性双特异性抗原结合分子的生物等效变体。例如,可以缺失非生物活性所必需的半胱氨酸残基或用其它氨基酸替代,以防止在复性时形成不必要的或不正确的分子内二硫桥。在其它背景下,生物等效抗体可以包含本文所示的示例性双特异性抗原结合分子,所述示例性双特异性抗原结合分子包括修饰抗体的糖基化特性的氨基酸变化,例如消除或去除糖基化的突变。
物种选择性和物种交叉反应性
根据某些实施例,本发明提供了与人CD28结合但不与来自其它物种的CD28结合的抗原结合分子。本发明还提供了与人CD22结合但不与来自其它物种的CD22结合的抗原结合分子。本发明还包含与人CD28结合并且与来自一个或多个非人物种的CD28结合的抗原结合分子;和/或与人CD22结合并且与来自一个或多个非人物种的CD22结合的抗原结合分子。
根据本发明的某些示例性实施例,提供了抗原结合分子,所述抗原结合分子与人CD28和/或人CD22结合并且视情况而定可以与小鼠、大鼠、豚鼠、仓鼠、沙鼠、猪、猫、狗、兔子、山羊、绵羊、牛、马、骆驼、猕猴、狨猴、恒河猴或黑猩猩CD28和或CD22中的一种或多种结合或不与其结合。例如,在本发明的特定示例性实施例中,提供了双特异性抗原结合分子,其包括结合人CD28和猕猴CD28的第一抗原结合结构域以及特异性地结合人CD22的第二抗原结合结构域。
免疫缀合物
本发明涵盖与治疗部分缀合的抗原结合分子(“免疫缀合物”),如细胞毒素、化学治疗药物、免疫抑制剂或放射性同位素。细胞毒性剂包含对细胞有害的任何药剂。用于形成免疫缀合物的合适的细胞毒性剂和化学治疗剂的实例是本领域已知的(参见例如,WO 05/103081,所述文献的全部内容通过引用并入本文)。
治疗性调配物和施用
本发明提供了包括本发明的抗原结合分子的药物组合物。本发明的药物组合物与合适的载体、赋形剂和提供经改善的转移、递送、耐受性等的其它药剂一起调配。在所有药物化学家已知的处方集中可以找到许多合适的调配物:《雷明顿氏药物科学(Remington'sPharmaceutical Sciences)》,宾夕法尼亚州伊斯顿的马克出版公司(Mack PublishingCompany,Easton,PA)。这些调配物包含例如粉末、糊剂、软膏、凝胶剂、蜡、油、脂质、含有囊泡的脂质(阳离子或阴离子)(如LIPOFECTIN
施用于患者的抗原结合分子的剂量可以根据患者的年龄和体型、目标疾病、病状、施用途径等而变化。通常根据体重或体表面积计算优选的剂量。当本发明的双特异性抗原结合分子出于治疗目的用于成年患者中时,通常以约0.01到约20mg/kg体重、更优选地约0.02到约7mg/kg体重、约0.03到约5mg/kg体重或约0.05到约3mg/kg体重的单剂量静脉内施用本发明的双特异性抗原结合分子可以是有利的。根据病状的严重程度,可以调整治疗的频率和持续时间。可以凭经验确定用于施用双特异性抗原结合分子的有效剂量和时间表;例如,可以通过定期评估来监测患者进展,并且相应地调整剂量。此外,可以使用本领域熟知的方法执行剂量的种间类推(例如,Mordenti等人,1991,《药学研究(Pharmaceut.Res.)》,8:1351)。
各种递送系统是已知的并且可以用于施用本发明的药物组合物,例如,包封在脂质体、微颗粒、微胶囊、能够表达突变病毒的重组细胞、受体介导的胞吞作用中(参见例如,Wu等人,1987,《生物化学杂志(J.Biol.Chem.)》262:4429-4432)。引入的方法包含但不限于真皮内、肌肉内、腹膜内、静脉内、皮下、鼻内、硬膜外和口服途径。组合物可以通过任何方便的途径例如通过输注或快速浓注、通过经由上皮或皮肤粘膜内层(例如,口腔粘膜、直肠和肠道粘膜等)吸收来施用并且可以与其它生物活性剂一起施用。施用可以是全身性的或局部的。
可以用标准针头和注射器皮下或静脉内递送本发明的药物组合物。另外,就皮下递送而言,笔递送装置易于应用于递送本发明的药物组合物。这种笔递送装置可以是可重复使用的或一次性的。可重复使用的笔递送装置通常利用含有药物组合物的可更换药筒。一旦已经施用了药筒内的全部药物组合物,并且药筒是空的,就可以容易地丢弃空药筒并且用含有药物组合物的新药筒更换。然后可以重复使用笔递送装置。在一次性笔递送装置中,没有可更换的药筒。相反,一次性笔递送装置预填充有固持在装置内的贮存器中的药物组合物。一旦贮存器中的药物组合物清空,就丢弃整个装置。
许多可重复使用的笔递送装置和自动注射器递送装置应用于皮下递送本发明的药物组合物。实例包含但不限于AUTOPEN
在某些情况下,药物组合物可以在受控释放系统中递送。在一个实施例中,可以使用泵(参见Langer,同上;Sefton,1987,《CRC:生物工程关键评论(CRC Crit.RefBiomed.Eng.)》14:201)。在另一个实施例中,可以使用聚合物材料;参见《控释医学应用(Medical Applications of Controlled Release)》,Langer和Wise(编辑),1974,佛罗里达州波卡拉顿d CRC出版社(CRC Pres.,Boca Raton,Florida)。在又另一个实施例中,控释系统可以放置在组合物的靶标附近,因此仅需要全身剂量的一部分(参见例如,Goodson,1984,《控释医学应用(Medical Applications of Controlled Release)》同上,第2卷,第115-138页)。其它控释系统在Langer,1990,《科学》249:1527-1533的评论中讨论。
可注射制剂可以包含用于静脉内、皮下、皮内和肌内注射、滴注等的剂型。可以通过公开已知的方法来制备这些可注射制剂。例如,可以例如通过将上述抗体或其盐溶解、悬浮或乳化在无菌水性介质或常规用于注射的油性介质中来制备可注射制剂。作为用于注射的水性介质,存在例如生理盐水、含有葡萄糖的等渗溶液以及其它助剂等,其可以与如醇(例如,乙醇)、多元醇(例如,丙二醇、聚乙二醇)等合适的增溶剂、非离子表面活性剂[例如,聚山梨醇酯80、HCO-50(氢化蓖麻油的聚氧乙烯(50mol)加合物)]等组合地使用。作为油性介质,采用例如芝麻油、大豆油等,其可以与如苯甲酸苄酯、苯甲醇等增溶剂等组合地使用。如此制备的注射剂优选地填充在合适的安瓿中。
有利地是,将上述用于口服或肠胃外使用的药物组合物制备成适合于配合一定剂量的活性成分的单位剂量剂型。这种单位剂量剂型包含例如片剂、丸剂、胶囊剂、注射剂(安瓿)、栓剂等。所含的上述抗体的量通常为每单位剂量剂型约5mg到约500mg;特别是在注射形式下,优选的是,上述抗体的含量为约5mg到约100mg并且对于其它剂型为约10mg到约250mg。
抗原结合分子的治疗性用途
本发明包含包括向有需要的受试者施用治疗性组合物的方法,所述治疗性组合物包括抗CD28抗体或特异性地结合CD28和靶抗原(例如,CD22)的双特异性抗原结合分子。治疗性组合物可以包括本文所公开的抗体或双特异性抗原结合分子中的任何一个以及药学上可接受的载体或稀释剂。如本文所使用的,表述“有需要的受试者”意指表现出一种或多种癌症症状或征兆(例如,表现出肿瘤或患有本文在下文提及的癌症中的任何一种癌症的受试者)或者以其它方式受益于CD22活性的抑制或降低或者CD22+细胞的耗竭的人或非人动物。
本发明的抗体和双特异性抗原结合分子(以及包括其的治疗性组合物)尤其可用于治疗其中刺激、活化和/或靶向免疫应答将会有益的任何疾病或病症。具体地,本发明的抗CD28/抗CD22双特异性抗原结合分子可以用于治疗、预防和/或改善与CD22表达或活性或者CD22+细胞增殖相关或由其介导的任何疾病或病症。实现本发明的治疗性方法的作用机制包含在效应细胞例如T细胞存在的情况下杀伤表达CD22的细胞。可以使用本发明的双特异性抗原结合分子抑制或杀伤的表达CD22的细胞包含例如癌性B细胞。
本发明的抗原结合分子可以用于治疗例如血液、骨髓、淋巴结(例如,胸腺、脾脏)、结肠、肝脏、肺、乳腺、肾癌、中枢神经系统和膀胱癌中出现的原发性和/或转移性肿瘤。根据某些示例性实施例,本发明的双特异性抗原结合分子用于治疗B细胞增殖性病症。
本发明还包含用于治疗受试者的残留癌症的方法。如本文所使用的,术语“残留癌症”意指在用抗癌疗法治疗后受试者的一个或多个癌细胞的存在或持续存在。
根据某些方面,本发明提供了用于治疗与CD22表达(例如,B细胞增殖性病症)相关的疾病或病症的方法,所述方法包括在示出受试者对其它类型的抗癌疗法无应答后向受试者施用本文其它地方描述的双特异性抗原结合分子中的一种或多种。例如,本发明包含用于治疗B细胞增殖性病症的方法,所述方法包括在受试者已经接受针对患有癌症例如B细胞增殖性病症的患者的标准护理后1天、2天、3天、4天、5天、6天、1周、2周、3周或4周、2个月、4个月、6个月、8个月、1年或更长时间向患者施用抗CD28/抗CD22双特异性抗原结合分子。在其它方面,最初在一个或多个时间点处向受试者施用包括IgG4 Fc结构域的本发明的双特异性抗原结合分子(抗CD28/抗CD22双特异性抗原结合分子)(例如,以提供前列腺癌细胞的稳健初始耗竭),之后在随后的时间点处施用包括不同IgG结构域如IgG1 Fc结构域的等效双特异性抗原结合分子。可以设想的是,可以结合其它双特异性抗原结合分子如结合抗CD20/抗CD3双特异性抗体使用本发明的抗CD28/抗CD22抗体。还可以设想的是,结合检查点抑制剂例如靶向PD-1和CTLA-4以及其它靶标的检查点抑制剂使用本发明的双特异性抗体。将靶向同一肿瘤抗原(例如,CD22)的两种双特异性抗体组合可以是有利的,但是其中一种双特异性抗体靶向T细胞上的CD3,而另一种双特异性抗体靶向如CD28等共刺激分子。此组合可以单独用于增强肿瘤细胞杀伤或者可以与检查点抑制剂组合地使用。
组合疗法和调配物
本发明包含包括本文所述的示例性抗体和双特异性抗原结合分子中的任何一种与一种或多种另外的治疗活性组分的组合的组合物和治疗性调配物以及包括向有需要的受试者施用此类组合的治疗方法。
可以与本发明的抗原结合分子组合或组合施用的另外的示例性治疗剂包含例如化学疗法、放射疗法、靶向PD-1的检查点抑制剂(例如,如派姆单抗、纳武单抗或西米普利单抗等抗PD-1抗体,参见US9,987,500,SEQ ID NO 162/170的HCVR/LCVR)、CTLA-4、LAG3、TIM3等、如GITR、OX40、4-1BB等靶向分子的共刺激激动剂二价抗体、CD3x双特异性抗体(参见例如US9,657,102(REGN1979)、WO2017/053856A1、WO2014/047231A1、WO2018/067331A1和WO2018/058001A1)、靶向CD22 X CD3、CD22 X CD28或靶向CD20 X CD3的其它抗体以及其它共刺激CD28x双特异性抗体。
可以与本发明的抗体有益地组合施用的其它药剂包含例如他莫昔芬(tamoxifen)、芳香酶抑制剂和细胞因子抑制剂,包含小分子细胞因子抑制剂和与如IL-1、IL-2、IL-3、IL-4、IL-5、IL-6、IL-8、IL-9、IL-11、IL-12、IL-13、IL-17、IL-18等细胞因子或其各自的受体结合的抗体。本发明的药物组合物(例如,包括如本文所公开的抗CD28/抗CD22双特异性抗原结合分子的药物组合物)还可以作为治疗性方案的一部分施用,所述治疗性方案包括选自以下的一种或多种治疗组合:“ICE”:异环磷酰胺(例如,
本发明还包含治疗组合,所述治疗组合包括本文提及的抗原结合分子中的任何一种和VEGF、Ang2、DLL4、EGFR、ErbB2、ErbB3、ErbB4、EGFRvlll、cMet、IGF1 R、B-raf、PDGFR-o、PDGFR-I3、FOLH1、PRLR、STEAP1、STEAP2、TMPRSS2、MSLN、CA9、尿溶蛋白(uroplakin)中的一个或多个或上述细胞因子中的任何一个的抑制剂,其中所述抑制剂是适体、反义分子、核酶、siRNA、肽体、纳米抗体或抗体片段(例如,Fab片段;F(ab')
本发明包含包括本文所述的抗原结合分子中的任何一种与一种或多种化学治疗剂的组合的组合物和治疗性调配物。化学治疗剂的实例包含:烷化剂,如噻替派(thiotepa)和环磷酰胺(Cytoxan
可以在施用本发明的抗原结合分子之前、同时或之后立刻施用一种或多种另外的治疗活性组分;(出于本公开的目的,此类施用方案被认为是抗原结合分子“与另外的治疗活性组分组合”施用)。
本发明包含药物组合物,其中本发明的抗原结合分子与如本文其它地方所描述的一种或多种另外的治疗活性组分一起共同调配。
施用方案
根据本发明的某些实施例,可以在限定的时间过程内向受试者施用多剂量的抗原结合分子(例如,抗CD28抗体或特异性地结合CD22和CD28的双特异性抗原结合分子)。根据本发明的此方面的方法包括向受试者依次施用多剂量的本发明的抗原结合分子。如本文所使用的,“依次施用”意味着在不同时间点例如在隔开预定间隔(例如,数小时、数天、数周或数月)的不同日期向受试者施用每个剂量的抗原结合分子。本发明包含包括以下的方法:向患者依次施用单一初始剂量的抗原结合分子、然后是一个或多个第二剂量的抗原结合分子并且任选地然后是一个或多个第三剂量的抗原结合分子。
术语“初始剂量”、“第二剂量”和“第三剂量”是指施用本发明的抗原结合分子的时间序列。因此,“初始剂量”是在治疗方案开始时施用的剂量(也被称为“基线剂量”);“第二剂量”是在初始剂量之后施用的剂量;并且“第三剂量”是在第二剂量之后施用的剂量。初始剂量、第二剂量和第三剂量可以均含有相同量的抗原结合分子,但是在施用频率方面通常可能彼此不同。然而,在某些实施例中,在治疗过程期间,初始剂量、第二剂量和/或第三剂量中含有的抗原结合分子的量彼此不同(例如,被适当地调高或调低)。在某些实施例中,在治疗方案开始时施用两个或更多个(例如,2个、3个、4个或5个)剂量作为“负荷剂量”,然后是以较低频率为基础施用的后续剂量(例如,“维持剂量”)。
在本发明的一个示例性实施例中,在前一剂量后1到26(例如,1、1
根据本发明的此方面的方法可以包括向患者施用任何数量的第二剂量和/或第三剂量的抗原结合分子(例如,抗CD28抗体或特异性地结合CD22和CD28的双特异性抗原结合分子)。例如,在某些实施例中,仅向患者施用单个第二剂量。在其它实施例中,向患者施用两个或更多个(例如,2个、3个、4个、5个、6个、7个、8个或更多个)第二剂量。同样,在某些实施例中,仅向患者施用单个第三剂量。在其它实施例中,向患者施用两个或更多个(例如,2个、3个、4个、5个、6个、7个、8个或更多个)第三剂量。
在涉及多个第二剂量的实施例中,每个第二剂量可以按与其它第二剂量相同的频率施用。例如,可以在前一剂量后1到2周向患者施用每个第二剂量。类似地,在涉及多个第三剂量的实施例中,每个第三剂量可以按与其它第三剂量相同的频率施用。例如,可以在前一剂量后2到4周向患者施用每个第三剂量。可替代地,向患者施用第二剂量和/或第三剂量的频率可以在治疗方案的过程中变化。施用频率也可以在由医师治疗的过程期间根据临床检查后个体患者的需要进行调整。
在一个实施例中,将抗原结合分子(例如,特异性地结合CD22和CD28的双特异性抗原结合分子)以基于体重的剂量施用于受试者。“基于体重的剂量”(例如,以mg/kg为单位的剂量)是根据受试者的体重改变的抗体或其抗原结合片段或双特异性抗原结合分子的剂量。
在另一个实施例中,将抗体或其抗原结合片段或双特异性抗原结合分子以固定剂量施用于受试者。“固定剂量”(例如,以mg为单位的剂量)意味着将一个剂量的抗体或其抗原结合片段或双特异性抗原结合分子用于所有受试者,而不论任何特定受试者相关因素如体重如何。在一个特定实施例中,本发明的抗体或其抗原结合片段或双特异性抗原结合分子的固定剂量是基于预定的体重或年龄。
通常,抗原结合分子本发明的合适剂量的范围可以为约0.001到约200.0毫克每千克接受者体重,通常范围为约1到50mg每千克体重。例如,抗体或其抗原结合片段或双特异性抗原结合分子可以按每单剂量约0.1mg/kg、约0.2mg/kg、约0.5mg/kg、约1mg/kg、约1.5mg/kg、约2mg/kg、约3mg/kg、约5mg/kg、约10mg/kg、约15mg/kg、约20mg/kg、约25mg/kg、约30mg/kg、约40mg/kg、约50mg/kg施用。所陈述值的中间值和范围也旨在是本发明的一部分。
在一些实施例中,将本发明的抗原结合分子按约25mg到约2500mg之间的固定剂量施用。在一些实施例中,本发明的抗原结合分子按以下固定剂量施用:约25mg、约30mg、约50mg、约75mg、约100mg、约125mg、约150mg、约175mg、200mg、约225mg、约250mg、约275mg、约300mg、约325mg、约350mg、约375mg、约400mg、约425mg、约450mg、约475mg、约500mg、约525mg、约550mg、约575mg、约600mg、约625mg、约650mg、约675mg、约700mg、约725mg、约750mg、约775mg、约800mg、约825mg、约850mg、约875mg、约900mg、约925mg、约950mg、约975mg、约1000mg、约1500mg、约2000mg或约2500mg。所陈述值的中间值和范围也旨在是本发明的一部分。
抗体的诊断用途
也可以例如出于诊断目的使用本发明的双特异性抗体来检测和/或测量样品中的CD28或CD22或者CD28表达性或CD22表达性细胞。例如,可以使用抗抗CD28 x抗CD22抗体或其片段来诊断以CD28或CD22的异常表达(例如,过度表达、表达不足、缺乏表达等)为特征的病状或疾病。用于CD28或CD22的示例性诊断测定可以包括例如使从患者获得的样品与本发明的抗体接触,其中用可检测的标记或报告分子来标记所述抗体。可替代地,未标记的抗体可以与本身被可检测地标记的第二抗体组合用于诊断应用。可检测的标记或报告分子可以是:放射性同位素,如
实例
提出以下实例,以向本领域普通技术人员提供关于如何制备和使用本发明的方法和组合物的完整公开和描述,并且以下实例不旨在限制发明人认为是其发明的范围。已经努力确保关于所使用的数字(例如,量、温度等)的准确性,但是应当考虑一些实验误差和偏差。除非另外指示,否则份数是重量份数,分子量是平均分子量,温度是摄氏度,并且压力是或接近大气压。
实例1:抗CD22xCD28抗体的构建
抗CD28抗体的生成
通过用与小鼠IgG2a的Fc部分融合的人CD28蛋白或用表达CD28的细胞或用对CD28进行编码的DNA对
根据此实例的方法生成的示例性抗CD28抗体的某些生物学性质在下文所示的实例中详细描述。
抗CD22抗体的生成
通过用人CD22抗原(例如,参见hCD22 ecto(D20-R687).hFc,R&D系统公司,目录号1968-SL-050;登录号CAA42006)(还参见图3)对经基因修饰的小鼠(
免疫后,从每只小鼠收获脾细胞,并且将其(1)与小鼠骨髓瘤细胞融合以保持其活力并形成杂交瘤细胞并筛选其CD22特异性或(2)使用人CD22片段作为结合和鉴定反应性抗体(抗原阳性B细胞)的分选试剂进行B细胞分选(如US 2007/0280945A1中所描述的)。
最初分离出具有人可变区和小鼠恒定区的针对CD22的嵌合抗体。对抗体进行表征和选择以获得包含亲和力、选择性等期望的特性。如果需要,则用期望的人恒定区例如野生型或经修饰的IgG1或IgG4恒定区来替代小鼠恒定区,以生成完全人抗CD22抗体。虽然所选择的恒定区可以根据具体用途而变化,但是高亲和力抗原结合和靶标特异性特性存在于可变区中。
结合CD28和CD22的双特异性抗体的生成
使用标准方法构建包括抗CD22特异性结合结构域和抗CD28特异性结合结构域的双特异性抗体,其中抗CD22抗原结合结构域和抗CD28抗原结合结构域各自包括与共同LCVR配对的不同的独特HCVR。在一些情况下,利用来自抗CD28抗体的重链、来自抗CD22抗体的重链和共同轻链构建双特异性抗体(参见表1)。
根据本实例产生的双特异性抗体包括两个单独的抗原结合结构域(即,结合臂)。第一抗原结合结构域包括衍生自抗CD28抗体的重链可变区(“CD28-VH”),并且第二抗原结合结构域包括衍生自抗CD22抗体的重链可变区(“CD22-VH”)。抗CD22抗CD28两者共享共同的轻链。CD28-VH/CD22-VH配对产生了特异性地识别T细胞上的CD28和肿瘤细胞上的CD22的抗原结合结构域。
实例2:重链和轻链可变区氨基酸和核酸序列
表1列示了本发明的所选抗CD22抗体的重链和轻链可变区以及CDR的氨基酸序列标识符。对应的核酸序列标识符列示于表2中。
表1:CD22抗体的氨基酸序列标识符
表2:CD22抗体的核酸序列标识符
表3列示了本发明的所选抗CD28抗体的重链和轻链可变区(HCVR和LCVR)、CDR的氨基酸序列标识符。对应的核酸序列标识符列示于表4中。
表3:CD28抗体的氨基酸序列标识符
表4:CD28抗体的核酸序列标识符
表5中列示了构建的各种抗CD22x抗CD28双特异性抗体的组分部分汇总。表6和7列示了双特异性抗体的HCVR、LCVR、CDR以及重链和轻链序列标识符。
表5:抗CD22 x抗CD28双特异性抗体的组分部分汇总
表6示出了本文例证的用于双特异性抗CD22 x抗CD28抗体的氨基酸序列标识符。对应的核酸序列标识符列示于表7中。
表6:抗CD22 x抗CD28双特异性抗体的氨基酸序列
表7:抗CD22 x抗CD28双特异性抗体的核酸序列
实例3:通过氢氘交换对REGN5837与CD22的结合的表位作图
执行质谱法H/D交换表位作图(HDX-MS)以便确定CD22(重组人CD22,SEQ ID NO:50)中的与H4sH33037P2(参见表1,SEQ ID NO:2/10的HCVR/LCVR对)(抗hCD22单克隆抗体;REGN5837的亲本抗hCD22抗体)相互作用的氨基酸残基。在例如以下文献中列示了H/D交换方法的一般说明:Ehring(1999)《分析生物化学》267(2):252-259;以及Engen和Smith(2001)《分析化学》73:256A-265A。
在集成的HDX/MS平台上执行HDX-MS实验,所述平台由用于氘标记和淬火的Leaptec HDX PAL系统、用于样品消解和上样的Waters Acquity M-Class(辅助溶剂管理器)、用于解析梯度的Waters Acquity M-Class(μ二元溶剂管理器)和用于肽质量测量的Thermo Q Exactive HF质谱仪组成。
将标记溶液制备成pD 7.0的在D
使用Byonic搜索引擎(蛋白质度量公司(Protein Metrics))针对包含CD22和其随机化序列的数据库搜索未氘化的CD22样品的LC-MS/MS数据。使用非特异性酶消化和人糖基化作为常见的变量修饰,将搜索参数(在ELN中)设置为默认值。然后将鉴定出的肽的列表导入HDX工作台软件(版本3.3)中,以计算通过LC-MS从所有氘化样品中检测到的每种肽的氘摄取量。对于给定的肽,使用每个时间点处的质心质量(强度加权平均质量)来计算氘摄取量(D)和氘摄取量百分比(%D)(参见下文)。
从单独的hCD22.mmH和与H4sH33037P2(SEQ ID NO:2/10的HCVR/LCVR对)样品复合的hCD22.mmH两者中鉴定出总共427种来自hCD22.mmH的肽(SEQ ID NO:50),这表示hCD22的序列覆盖率为92.0%。表现出高于5%的D摄取量值差异百分比的任何肽均被定义为受到显著保护(表8)。对于hCD22.mmH,对应于氨基酸481-505(NVQYAPRDVRVRKIKPLSEIHSGNS;SEQID NO:57)和523-537(FWEKNGRLLGKESQLNF;SEQ ID NO:58)的肽受H4sH33037P2显著保护。
表8:与H4sH33037P2结合时具有显著保护的所选CD22.mmH肽
实例4:通过氢氘交换对H4sH33041P2与CD22的结合的表位作图
执行质谱法H/D交换肽表位作图(HDX-MS)以便确定与H4sH33041P2(具有SEQ IDNO:18/10的HCVR/LCVR对的抗hCD22单克隆抗体)(REGN5838的亲本抗hCD22)相互作用的CD22(重组人CD22,SEQ ID NO:50)的氨基酸残基。在例如以下文献中列示了H/D交换方法的一般说明:Ehring(1999)《分析生物化学》267(2):252-259;以及Engen和Smith(2001)《分析化学》73:256A-265A。
在集成的HDX/MS平台上执行HDX-MS实验,所述平台由用于氘标记和淬火的Leaptec HDX PAL系统、用于样品消解和上样的Waters Acquity M-Class(辅助溶剂管理器)、用于解析梯度的Waters Acquity M-Class(μ二元溶剂管理器)和用于肽质量测量的Thermo Q Exactive HF质谱仪组成。
将标记溶液制备成pD 7.0的在D
使用Byonic搜索引擎(蛋白质度量公司)针对包含CD22和其随机化序列的数据库搜索未氘化的CD22样品的LC-MS/MS数据。使用非特异性酶消化和人糖基化作为常见的变量修饰,将搜索参数(在ELN中)设置为默认值。然后将鉴定出的肽的列表导入HDX工作台软件(版本3.3)中,以计算通过LC-MS从所有氘化样品中检测到的每种肽的氘摄取量。对于给定的肽,使用每个时间点处的质心质量(强度加权平均质量)来计算氘摄取量(D)和氘摄取量百分比(D%),如下文所示。
从单独的hCD22.mmH和与H4sH33041P2样品复合的hCD22.mmH两者中鉴定出总共454种来自hCD22.mmH(SEQ ID NO:50)的肽,这表示hCD22的序列覆盖率为90.5%。表现出高于5%的D摄取量值差异百分比的任何肽均被定义为受到显著保护。对于hCD22.mmH,对应于氨基酸246-277(CEVSSSNPEYTTVSWLKDGTSLKKQNTFTLNL;SEQ ID NO:59)的肽受H4sH33041P2显著保护。表9提供了来自与H4sH33041P2结合时具有显著保护的所选肽的结果。
表9:与H4sH33041P2结合时具有显著保护的所选CD22.mmH肽
实例5:CD22 xCD28双特异性抗体的表面等离子体共振得到的结合亲和力和动力学常数
使用利用Biacore T-200或Biacore 4000仪器的实时表面等离子体共振生物传感器确定hCD22.mmH(SEQ ID NO:50)和mfCD22.mmH(SEQ ID NO:51)与纯化的抗CD22xCD28双特异性mAb或抗CD22双价亲本mAb(参见表1,mAB33037P2;HCVR/LCVR:SEQ ID NO:2/10)和mAb33041P2HCVR/LCVR:SEQ ID NO:18/10)结合的平衡解离常数(K
K
在25℃下人和cyno CD22与纯化的mAb结合的结合动力学参数在下文示出在表10-12中。
使用利用Biacore T-200仪器的实时表面等离子体共振生物传感器确定hCD28.mmH(SEQ ID NO:54)纯化的抗CD22xCD28双特异性mAb或抗CD28双价亲本mAb(关于mAb14226P2,参见表3和4)的平衡解离常数(K
K
在25℃下人CD28与纯化的mAb结合的结合动力学参数在下文示出在表13中。
表10:在25℃下人CD22.mmH与抗CD22xCD28双特异性mAb的结合动力学
表11:在25℃下猴CD22.mmH(XP_005588899.1)与抗CD22xCD28双特异性mAb的结合动力学
表12:在25℃下猴CD22.mmH(EHH59463.1)与抗CD22xCD28双特异性mAb的结合动力学
表13:在25℃下人CD28.mmH与抗CD22xCD28双特异性mAb的结合动力学
实例6:使用流式细胞术的抗CD28和抗CD22xCD28双特异性抗体与靶细胞系(Nalm6)、效应细胞系(Jurkat)以及食蟹猴T细胞和B细胞的结合特异性
利用流式细胞分析来确定CD22xCD28双特异性抗体与表达Nalm6细胞的人CD22和表达Jurkat细胞的人CD28以及与食蟹猴T(CD28+)细胞和B(CD22+)细胞的结合。简而言之,将Jurkat细胞和Nalm6细胞的1x10
在与第二抗体一起温育之后,洗涤细胞、将其重悬于200μL含有1%过滤FBS的冷PBS中并在BD LSR_Fortessa上通过流式细胞术进行分析。将食蟹猴T细胞鉴定为CD2+/CD16-并且将B细胞鉴定为CD20+。使用4参数非线性回归分析在Prism软件中计算FACS结合的EC
表14提供了CD22xCD28双特异性抗体与表达CD22的细胞系表面的结合数据,如通过流式细胞术确定的。表14还提供了CD22xCD28双特异性抗体与表达人CD28的细胞系表面的结合数据,如通过流式细胞术确定的。
REGN5837与Nalm6细胞以1.3E-08M的EC
REGN5837与Jurkat细胞以2.1E-08M的EC
表15提供了CD22xCD28双特异性抗体与食蟹猴(柬埔寨来源)T细胞和B细胞的表面的数据的结合,如通过流式细胞术确定的。
REGN5837与所测试的12只食蟹猴中的12只的B细胞以及所测试的12只食蟹猴中的11只的T细胞结合。与CD20+B细胞的结合的范围为第二抗体的12.6-30.3倍,其中中位数是15.7倍。与CD2+/CD16-T细胞的结合的范围为第二抗体的1.2-5.2倍,其中中位数是3.5倍。将阳性结合定义为比二级抗体大1.2倍。REGN5838与所测试的12只食蟹猴中的12只的B细胞以及所测试的12只食蟹猴中的11只的T细胞结合。与CD20+B细胞的结合的范围为第二抗体的6.5-13.5倍,其中中位数是9.3倍。与CD2+/CD16-T细胞的结合的范围为第二抗体的1.2-4.7倍,其中中位数是3.8倍。将阳性结合定义为比第二抗体大1.2倍。同种型对照抗体未表现出与猕猴T或B细胞的任何结合。
表14:工程化靶细胞和效应细胞上的流式细胞术实验的结合和结合倍数结果。
表15:猕猴(柬埔寨来源)T和B细胞上的流式细胞术实验的结合倍数结果。
实例7:使用流式细胞术的抗CD28和抗CD22xCD28双特异性抗体与人CD4+T细胞和工程化的靶细胞的结合特异性
使用流式细胞术分析来研究CD22 x CD28双特异性(REGN5837;REGN5838)和对照抗体与表达人CD28(人CD4
从自健康供体白细胞包中获得的人外周血单核细胞(PBMC)分离出人CD4
如下制备包含HEK293细胞系和人Raji B细胞的靶细胞。
用人CD22(登录号NP_001762.2的氨基酸M1到A847)转导表达人CD20(登录号NP_068769.2的氨基酸M1到P297)的稳定HEK293细胞系(ATCC,#CRL-1573)。将人CD22阳性细胞通过荧光活化细胞分选(FACS)进行分离并进行单克隆。将所得的克隆细胞系(HEK293/hCD20/hCD22克隆E4)维持在补充有500g/mL G418的DMEM+10%+P/S/G+NEAA中。
使用CRIPSR技术通过缺失CD80和CD86来基因修饰人Raji B细胞(ATCC#CCL-86),所述细胞在细胞表面上内源性地表达CD20、CD22、Fcγ受体(FcγR)、CD80和CD86。CD80和CD86是已知的CD28配体。将工程化的Raji/CD80和CD86阴性细胞维持在补充有HEPES和丙酮酸钠的RPMI+10%FBS+青霉素+链霉素+谷氨酰胺中。
如下对细胞进行染色。
简而言之,将人CD4+T细胞、HEK293/hCD20、HEK293/hCD20/hCD22以及Raji/CD80和CD86阴性细胞重悬于含有D-PBS+2%FBS的染色缓冲液中。将Raji细胞与小鼠IgG(最终浓度为625mg/mL)一起温育,以阻断内源性Fcγ受体。简而言之,在96孔平板中,将2x10
通过流式细胞术在原代人CD4
图1和表16中汇总了EC
表16:人CD4
缩写:NC=无法计算(针对其中结合没有达到饱和的曲线表示);ND=未确定
表16.抗体与人CD4
如所预期的,CD28抗体(亲本(REGN5705;HCVR/LCVR SEQ ID NO:35/36)或其双特异性形式(REGN5837、REGN5838和单臂CD28对照(SEQ ID NO:48))均未与阴性HEK293/hCD20细胞结合。由于非特异性结合,因此利用同种型对照抗体在最高浓度下检测到大约1.8倍的弱结合(图1和表16)。
在HEK293/hCD20/hCD22(REGN5837为16.44倍并且REGN5838为34.9倍,其中EC
在原代人CD4
实例8:通过抗CD20xCD3双特异性抗体的抗CD22xCD28双特异性抗体共刺激增强靶向的细胞毒性、T细胞活化和细胞因子释放
在靶向被工程化以缺乏CD80和CD86表达的Raji细胞(Raji-80/86DKO)的96小时细胞毒性测定中评估CD20xCD3靶向的杀伤的CD22xCD28增强。简而言之,将人PBMC以1×10
通过将细胞与同CD2、CD4、CD8和CD25直接缀合的抗体一起温育来评估T细胞活化。将表达CD25的CD8+细胞的百分比报告为T细胞活化度量。另外,随着T细胞的增殖,对CellTraceViolet进行稀释,这导致如FACS所测量的MFI较低。因此将T细胞增殖报告为CD8+T细胞上CellTraceViolet的MFI的减少。使用4参数非线性回归分析在Prism软件中计算缺乏CD80和CD86表达和结合的靶Raji细胞的EC
收集来自此测定的上清液以用于细胞因子水平的分析。按照制造商的说明书使用流式细胞珠阵列(CBA)试剂盒对IL 17a、IFNγ、TNFα、IL-10、IL-6、IL-4和IL-2的浓度进行分析。从试剂盒标准品所生成的曲线中插入细胞因子水平并将细胞因子水平报告为pg/mL。在Prism软件中使用4参数非线性回归分析计算最大细胞因子水平。
测试用于评估抗CD20xCD3双特异性抗体REGN1979(参见上文)与共刺激CD22xCD28抗体或单臂CD28或同种型对照抗体组合地诱导未刺激的人T细胞杀伤表达人CD20和CD22的靶细胞的能力的测定的结果。
REGN1979活化并指导人T细胞以剂量依赖性方式耗竭缺乏CD80和CD86表达的Raji细胞。与REGN1979和单臂CD28或同种型对照抗体相比,将固定浓度的CD22xCD28双特异性抗体添加到REGN1979使REGN1979的细胞毒性功效(EC
如分别通过CD8+细胞上的CD25上调或CellTrace Violet稀释测量的,观察到的由REGN1979介导的靶细胞裂解与T细胞活化和增殖相关。与REGN1979和单臂CD28或同种型对照抗体相比,将固定浓度的CD22xCD28双特异性抗体添加到REGN1979使REGN1979诱导T细胞活化和增殖的效力分别增强2.1到2.6倍和7.4-8.4倍(表17)。
REGN1979诱导人细胞因子的释放。利用REGN1979与CD22xCD28双特异性抗体组合地观察到的释放的细胞因子在固定浓度的CD22xCD28共刺激分子与固定浓度的单臂CD28或同种型对照抗体存在的情况下有所增强(表18)。
总之,与利用CD20xCD3同对照抗体组合地观察到的相比,共刺激增加了靶向的细胞毒性、T细胞活化和细胞因子释放的效力。
表17:细胞毒性和T细胞活化的EC
表18:细胞因子释放(pg/ml)
实例9:CD22双特异性抗体的生物测定
通过刺激T细胞受体(TCR)来实现T细胞活化,所述TCR识别在抗原呈递细胞(APC)上由I类或II类主要组织相容性复合物(MHCI或MHCII)蛋白呈递的特定肽(Goldrath等人1999)。活化的TCR进而发起一系列信号传导事件,所述信号传导事件可以由各种转录因子所驱动的报告基因监测,所述各种转录因子如活化蛋白1(AP-1)、活化的T细胞的核因子(NFAT)或活化的B细胞的核因子κ-轻链-增强子(NFκB)。然后,通过在T细胞上组成型地或可诱导地表达的共受体(如CD28、CTLA-4(细胞毒性T淋巴细胞相关蛋白4)、PD-1(程序性细胞死亡蛋白1)、LAG-3(淋巴细胞活化基因3)或其它分子)的接合来进一步细化T细胞应答(Sharpe等人2002)。共刺激分子CD28由其在APC上表达的内源性配体CD80或CD86活化。CD28在TCR活化之后加强细胞信号,如由NFκB转录因子控制的通路。CD28共信号对于有效的T细胞活化如T细胞分化、增殖、细胞因子释放和细胞死亡具有重要性(Smeets等人2012)。
使用原代人CD4
开发T细胞/APC(抗原呈递细胞)的基于荧光素酶的报告基因测定,以在评估与抗CD28 x抗CD22双特异性抗体接合时CD28活化对NFκB活性的影响。
依据制造商的说明书通过用NFκB-荧光素酶(NFκB-Luc)慢病毒报告基因(来自凯杰公司(Qiagen))转导永生人Jurkat T细胞(ATCC#TIB-152)来工程化克隆的报告T细胞系。将克隆的报告基因系(Jurkat/NFκB-Luc克隆1C11)维持在补充有1μg/mL嘌呤霉素的RPMI+10%FBS+青霉素+链霉素+谷氨酰胺中。
用人CD22(登录号NP_001762.2的氨基酸M1到A847)转导表达人CD20(登录号NP_068769.2的氨基酸M1到P297)的稳定HEK293细胞系(ATCC,#CRL-1573)。将人CD22阳性细胞通过荧光活化细胞分选(FACS)进行分离并进行单克隆。将所得的克隆细胞系(HEK293/hCD20/hCD22克隆E4)维持在补充有500μg/mL G418的DMEM+10%+P/S/G+NEAA中。
使用CRIPSR技术通过缺失CD80和CD86来基因修饰人Raji B细胞(ATCC#CCL-86),所述细胞在细胞表面上内源性地表达CD20、CD22、Fcγ受体(FcγR)、CD80和CD86。CD80和CD86是已知的CD28配体。将工程化的Raji/CD80和CD86阴性细胞维持在补充有HEPES和丙酮酸钠的RPMI+10%FBS+青霉素+链霉素+谷氨酰胺中。
在此实验中,通过两种双特异性抗体来刺激工程化的报告T细胞。通过靶向工程化的报告T细胞上的CD3分子以及HEK293上或Raji/CD80和CD86阴性B细胞上的CD20的T细胞活化双特异性抗体REGN2281(具有包含EQ ID NO:39的一个重链臂、包含SEQ ID NO:40的一个重链臂和SEQ ID NO:41的轻链臂的抗CD20 X抗CD3抗体)来递送第一刺激。第一刺激绕过了对TCR通过其天然配体来进行活化的需要,所述天然配体是MHC分子上显示的特定肽。第二刺激由CD28双特异性抗体(即,抗CD28 x抗CD22双特异性抗体)驱动。此抗体通过其在APC上表达的配体CD80/CD86来模仿T细胞上的CD28活化。所述抗体与T细胞上的CD28和HEK293细胞上或Raji/CD80和CD86阴性B细胞上的CD22互相作用并驱动工程化的报告T细胞上的CD28的活化。TCR和CD28的同时活化导致NFκB的转录活性增强,这增加了报告基因荧光素酶的产生。
将补充有10%FBS和P/S/G的RPMI1640作为测定介质用于制备用于筛选抗CD22 x抗CD28双特异性抗体的细胞悬浮液和抗体稀释液。
筛选前一天,在细胞培养基中将工程化报告T细胞培养至0.5×10
按以下顺序添加试剂:1)将固定浓度的最终200pM REGN2281(抗CD20 x抗CD3,参见上文)或REGN1945(hIgG4同种型对照,参见上文)添加到96孔白色平底平板的每个孔中;2)将重悬至4×10
通过使用GraphPad Prism
开发原代CD4
从健康供体白细胞包中分离出人外周血单核细胞(PBMC)。按照制造商的推荐方案使用50mL SepMate
在此测定中,使用抗CD20 x抗CD3双特异性抗体(REGN2281,参见上文)与工程化以表达人CD20的HEK293细胞或表达Raji细胞的内源性CD20组合地通过在原代CD4
在测定当天,在刺激培养基(补充有10%FBS、HEPES、NaPyr、NEAA和含有50U/ml全能核酸酶的0.01mM BME的X-VIVO 15细胞培养基)中解冻先前分离并冷冻的人CD4
通过使用GraphPad Prism
结果汇总和结论:
在使用荧光素酶活性作为读数的基于报告细胞的生物测定中,评估抗CD22 x抗CD28双特异性抗体在CD22靶标表达不存在或存在的情况下通过T细胞上的CD28提供共刺激的能力。
图2中(A和B)中示出了活化曲线,表19和20中汇总了除了200pM恒定REGN1945(hIgG4同种型对照)或REGN2281(抗CD20 x抗CD3)之外还与HEK293/hCD20或HEK293/hCD20/hCD22细胞共温育的工程化报告T细胞的EC
当用200pM REGN1945处理报告T细胞和HEK293衍生的APC时,在不存在TCR刺激的情况下,CD28双特异性抗体均未示出荧光素酶活性增加,而不管测定中使用的HEK293系如何。观察到仅HEK293/hCD20细胞(2.18倍)和HEK293/hCD20/hCD22细胞(2.05倍)的亲本CD28抗体(REGN5705)的萤光素酶活化增加。单臂CD28和同种型对照抗体在这种情况下没有产生萤光素酶应答(表19和图2)。
当报告T细胞和HEK293衍生的APC用200pM REGN2281处理时,抗CD22 x抗CD28双特异性抗体(REGN5837和REGN5838)均与CD22表达性HEK293诱导强荧光素酶活性,这通过EC
当报告T细胞和Raji/CD80和CD86阴性用200pM REGN1945处理时,抗CD22 x抗CD28双特异性抗体(REGN5837和REGN5838)和亲本CD28抗体均诱导荧光素酶活性,而单臂CD28对照抗体和同种型对照没有示出活性(表19和图2)。
当报告T细胞和Raji/CD80和CD86阴性用200pM REGN2281处理时,所有CD28双特异性抗体(REGN5837和REGN5838)和单臂CD28对照抗体(包含亲本CD28抗体)诱导荧光素酶活性。仅抗CD22 x抗CD28和亲本CD28抗体的EC
在测量IL-2细胞因子产生的功能性原代CD4
图3(A和B)中示出了活化曲线,表21中汇总了在2nM恒定REGN1945(hIgG4同种型对照)或REGN2281(抗CD20 x抗CD3)存在的情况下与HEK293/hCD20、HEK293/hCD20/hCD22或Raji/CD80和CD86阴性细胞共温育的CD4
在含有具有恒定量的REGN1945的HEK293/hCD20或HEK293/hCD20/CD22细胞的孔中,没有观察到可测量的IL-2释放,这是由于不存在足够的同种异体初级T细胞刺激(图3)。然而,在含有具有恒定量的REGN1945的Raji/CD80和CD86阴性细胞的孔中,检测到IL-2释放,这是由于显著的同种异体应答提供了足够的初级刺激,即使在不存在抗体介导的CD3聚类的情况下也是如此(图3和表21)。
当添加了恒定2nM浓度的REGN2281和亲本CD28 mab(REGN5705,参见上文)时,在含有HEK293/hCD20或HEK293/hCD20/CD22细胞的样品中检测到可测量的IL-2水平。与双价CD28 mAb形成对比,当抗CD22 x抗CD28双特异性mAb添加到含有HEK293/hCD20细胞和REGN2281的孔中时,IL2释放没有显著增强。仅在HEK293/hCD20/CD22细胞和REGN2281存在的情况下,抗CD22 x抗CD28双特异性mAb显著增强IL-2释放(图3和表22)。
下文列示了表19-22。
表19呈现了与HEK293/hCD20、HEK293/hCD20/hCD22或RAJI/CD80和CD86阴性细胞以及200pM恒定REGN1945(同种型对照)共温育的工程化T细胞中的荧光素酶活性的表列EC
表20呈现了与HEK293/hCD20、HEK293/hCD20/hCD22或Raji/CD80和CD86阴性细胞和200pM恒定REGN2281(抗CD20 x抗CD3)共温育的工程化T细胞中的荧光素酶活性的表列EC
表21呈现了来自与HEK293/hCD20、HEK293/hCD20/hCD22或RAJI/CD80和CD86阴性细胞以及2nM恒定REGN1945(同种型对照)共温育的CD4
表22呈现了来自与HEK293/hCD20、HEK293/hCD20/hCD22或Raji/CD80和CD86阴性细胞和2nM恒定REGN2281(CD20 x抗CD3)共温育的CD4
表19:在利用200pM REGN1945(同种型对照)不存在TCR刺激的情况下工程化报告T细胞中的荧光素酶活性的EC
缩写:ND=未确定
表20:在利用200pM REGN2281(αCD20 x αCD3)存在TCR刺激的情况下工程化报告T细胞中的荧光素酶活性的EC
缩写:NC=无法计算(针对其中应答没有达到饱和的曲线表示);ND=未确定。
表21:在2nM REGN1945(同种型对照)存在的情况下来自原代人CD4
缩写:NC=无法计算(针对其中应答没有达到饱和的曲线表示);ND=未确定。
表22:在2nM REGN2281(αCD20 x αCD3)存在的情况下来自原代人CD4
缩写:NC=无法计算(针对其中应答没有达到饱和的曲线表示);ND=未确定。
实例10:抗CD22 X抗CD28抗体的组合加上西米普利单抗对来自工程化以表达PD-L1的细胞的IL-2释放的效果
材料与方法
RAJI细胞
RAJI是从11岁男性(
RAJI/CD80和CD86阴性
使用CRISPR/Cas9系统消除RAJI细胞中的CD80和CD86的表达。
NALM6克隆G5
NALM6克隆是从19岁男性分离的急性淋巴母细胞白血病(ALL)细胞系[NALM6克隆G5(ATCC,#CRL-3273)]。将NALM6细胞维持在RPMI+10%FBS+P/S/G中。
WSU-DLCL2
WSU-DLCL2是从41岁白人男性的胸腔积液分离出来的人DLBCL细胞系(莱布尼兹研究所-DSMZ(Leibnitz Institute-DSMZ),目录号ACC 575)。
PD-L1工程化细胞系
将NALM-6、RAJI CD80和CD86阴性(RAJI/CD80-CD86-)和WSU-DLCL2细胞系进行基因工程化以稳定地表达人PD-L1(登录号NP_054862.1的氨基酸M1-T290)。将所得的细胞系NALM6/PD-L1、RAJI/CD80-CD86-/PD-L1和WSU-DLCL2/PD-L1维持在它们各自的培养基中,所述培养基补充有用于RAJI/CD80-CD86-的0.5μg/mL嘌呤霉素以及用于NALM-6/PD-L1和WSU-DLCL2/PD-L1细胞的1μg/mL嘌呤霉素。
用于T细胞增殖和IL-2释放的T细胞活化测定
使用人原代T细胞和同种异体B细胞淋巴瘤细胞系([NALM-6、NALM-6/PD-L1、RAJI/CD80-CD86-、RAJI/CD80-CD86-/PD-L1、WSU-DLCL2、WSU-DLCL2/PD-L1])在固定浓度的西米普利单抗存在的情况下评估REGN5837对IL-2释放的影响。原代白细胞与基因上不同的细胞的共培养造成同种异体决定簇的不相容并且可能导致T细胞活化。关于使用NALM-6和RAJI/CD80-CD86-(+/-PD-L1)细胞的测定,利用来自3个供体的富集的人原代T细胞执行T细胞活化测定,而利用WSU-DLCL2(+/-PD-L1)细胞的测定使用来自1个供体的T细胞。
在用于测试REGN5837+REGN2810组合处理的T细胞活化测定中使用的T细胞的分离。
关于利用NALM-6和RAJI/CD80-CD86-细胞的实验,CD3+T细胞从3个供体PBMC(555109、555130和555131)中分离出来,而来自一个供体(555175)的PBMC用于利用WSU-DLCL2细胞的测定。关于供体555109,使用密度梯度离心将PBMC从外周血中分离出来。简而言之,将15ml Ficoll-Paque PLUS添加到50ml圆锥管中,并且随后将与含有2%FBS的PBS以1:1稀释的30ml血在顶部分层。在关掉制动器的情况下以400x g离心30分钟之后,将单核细胞层转移到新鲜管中、利用含有2%FBS的PBS稀释5倍并以300x g离心8分钟。关于供体555130、555131和555175,按照制造商方案使用来自干细胞技术公司(Stem CellTechnologies)的EasySep直接人PBMC分离试剂盒(EasySep Direct Human PBMCIsolation Kit)将PBMC从来自健康供体的外周血中分离出来。将分离的PBMC冷冻在含有10%DMSO的FBS中。关于CD3+T细胞分离,将PBMC冷冻小瓶在37℃水浴中解冻并在含有50U/ml全能核酸酶的刺激培养基(补充有10%FBS、HEPES、NaPyr、NEAA和0.01mM BME的X-VIVO15细胞培养基)中稀释。按照制造商方案,将细胞以1200rpm离心10分钟、重悬于EasySep缓冲液中并使用干细胞技术公司EasySep T细胞分离试剂盒进行分离。
利用人OVCAR-3、PEO1、NALM-6、RAJI/CD80-CD86-和WSU-DLCL2细胞(+/-PD-L1)的T细胞活化测定
将重悬于刺激培养基(补充有10%FBS、HEPES、NaPyr、NEAA和0.01mM BME的X-VIVO15细胞培养基)中的CD3+T细胞以1x10
关于IL-2释放,根据制造商的方案,使用人IL-2αLISA试剂盒对5μL上清液进行测试。在珀金埃尔默公司的多标记平板读取器Envision上获取IL-2测量结果。已知的IL-2浓度的标准曲线包含在内并且用于衍生pg/ml值。
一式三份地对所有连续稀释液的IL-2释放进行测试。使用GraphPad Prism
结果汇总和结论:
在测量IL-2细胞因子产生的功能性原代CD3+T细胞测定中,评估抗CD22 x抗CD28双特异性抗体在内源性地表达CD22的B细胞淋巴细胞细胞系存在的情况下通过T细胞上CD28提供共刺激的能力。
图5A和5B中示出了与NALM-6(+/-PD-L1)(图5A)或RAJI/CD80-CD86-(+/-PD-L1)(图5B)温育的T细胞的活化曲线。在20nM恒定hIgG4
在人原代T细胞和同种异体B细胞淋巴细胞细胞系RAJI/CD80-CD86-和NALM-6存在的情况下,REGN5837介导了IL-2释放的浓度依赖性增加。非TAAxCD28对照抗体以高抗体浓度略微增加IL-2。在RAJI/CD80-CD86-或NALM-6细胞上不存在PD-L1的情况下,添加20nM西米普利单抗对IL-2释放没有影响。与同非PD-L1表达性细胞温育的T细胞相比,在表达PD-L1的RAJI/CD80-CD86-或NALM-6细胞存在的情况下,响应于单独利用REGN5837的处理释放的最大IL-2有所减少。在具有NALM-6/PD-L1细胞的条件下,添加西米普利单抗最低限度地增强了REGN5837介导的IL-2释放,然而在RAJI/CD80-CD86-/PD-L1细胞存在的情况下,这将IL-2释放显著地增强到在具有RAJI/CD80-CD86-细胞(缺乏PD-L1)的条件下所观察到的水平。
在人原代T细胞和同种异体B细胞淋巴细胞细胞系WSU-DLCL2存在的情况下,没有观察到IL-2释放的浓度依赖性增加。相反地,观察到非TAAxCD28对照抗体以浓度依赖性方式降低IL-2释放。与很少到不表达CD28配体的NALM-6和RAJI/CD80-CD86-细胞不同,已知WSU-DLCL2细胞系表达CD28配体。在已知REGN5837的CD28结合臂以及因此非TAAxCD28抗体的CD28臂与CD28配体竞争与CD28结合时,非TAAxCD28对照抗体通过在WSU-DLCL2细胞上表达的CD28配体阻断CD28活化,这导致IL-2释放降低。与非TAAxCD28对照不同,IL-2没有被REGN5837降低,因为其通过其CD22结合臂锚定到WSU-DLCL2细胞的能力允许其表现得类似于CD28配体,从而基本上替代所述配体。与不表达PD-L1的WSU-DLCL2细胞相比,在WSU-DLCL2/PD-L1细胞存在的情况下,基础IL-2释放有所降低。在西米普利单抗不存在的情况下单独添加REGN5837导致IL-2释放略微增强。在添加20nM西米普利单抗时,基础活性有所增强并且可以进一步利用剂量滴定REGN5837来略微增强,从而使REGN5837和西米普利单抗的组合的最大IL-2释放比单独的处理更高。如利用WSU-DLCL2细胞所观察到的,在非TAAxCD28存在的情况下WSU-DLCL2/PD-L1细胞的温育导致IL-2水平降低,而不论西米普利单抗或hIgG4
表23A:在工程化以表达PD-L1的NALM-6细胞中,REGN5837与西米普利单抗的组合使IL-2释放增强超过单独的REGN5837处理
ND:未确定,因为没有观察到浓度依赖性应答。
表23B:在工程化以表达PD-L1的RAJI/CD80
NC:未计算,因为数据未拟合4参数逻辑方程。
表24:在表达PD-L1的WSU-DLCL2细胞中,REGN5837与西米普利单抗的组合使IL-2释放增强超过单独的REGN5837处理
ND:未确定,因为没有观察到浓度依赖性应答。
NC:未计算,因为数据未拟合4参数逻辑方程。
实例11:在REGN1979存在或不存在的情况下施用REGN5837的抗肿瘤功效
引言
REGN5837是被设计成用于通过桥接CD22
下文所描述的研究评估了在次有效剂量的CD20xCD3 bsAb(REGN1979)存在或不存在的情况下向携带已确立B细胞白血病肿瘤8天的免疫缺陷NSG小鼠施用CD22xCD28 bsAbREGN5837的抗肿瘤功效。
简而言之,向小鼠(每组n=6到9)腹膜内(IP)移植人外周血单核细胞(PBMC)并在12天之后静脉内(IV)植入人NALM-6B细胞白血病细胞,所述人NALM-6 B细胞白血病细胞被工程化以表达荧光素酶从而实现生物发光成像(NALM-6-luc)。将0.04mg/kg、0.4mg/kg和4mg/kg REGN5837与0.04mg/kg固定剂量的REGN1979组合的抗肿瘤功效与REGN5837和REGN1979单一疗法并且与非桥接IgG4
材料与方法
人衍生的细胞系
NALM-6-luc:NALM-6细胞系是从19岁男性患者(DSMZ,目录号ACC 128)分离的急性淋巴母细胞白血病细胞系;用EF1a-荧光素酶-2A-GFP-Puro慢性病(基因靶标公司(GenTarget))来修饰此系以便于体内肿瘤细胞生长成像。
PBMC:从ReachBio公司目录号0500-401供体号0180905(肿瘤生长实验)和0180621(血清抗体试验)获得人PBMC
实验设计
测试系统
在所有实验中使用雌性NSG小鼠(年龄为8-9周龄)。向所有小鼠IP移入人PBMC并且然后在移植后12天IV植入NALM-6-luc B细胞白血病细胞。实验设计在表25中详述。在整个研究期间,通过一周两次生物发光成像来监测肿瘤生长。对于所有的实验,将小鼠收容在标准条件下的再生元公司(Regeneron)动物设施中。所有实验根据再生元公司的机构动物护理和使用委员会(the Institutional Animal Care and Use Committee)的准则执行。
NSG小鼠的移植
向雌性免疫缺陷NSG小鼠IP移植4x10
NALM-6-Luc培养条件和肿瘤植入
用EF1a-荧光素酶-2A-GFP-Puro慢病毒(基因靶标公司)来修饰NALM-6细胞系以便于体内肿瘤细胞生长成像。将细胞系维持在于嘌呤霉素选择下补充有PSG(青霉素、链霉素和谷氨酰胺)的10%FBS的RPMI中。
通过离心收集NALM-6-luc细胞并以2.5x10
用于肿瘤测量的抗体给药
在用测试物品或对照给药之前,将小鼠分配到根据肿瘤负荷和T细胞移植水平分层的组。按照表25所陈述的剂量在植入(为了体内功效)后第8天、第15天和第22天通过IP注射作为单一疗法或组合地施用抗体(REGN5837、REGN1979、REGN5671[非TAAxCD28非桥接对照bsAb]或H4sH17664D[非TAAxCD3非桥接对照bsAb])。
表25:用于评估肿瘤生长的实验设计
肿瘤测量和所指定终点
使用IVIS光谱仪器每周两次对植入有NALM-6-luc肿瘤的小鼠进行成像,并且使用Living Image软件分析数据。成像之前,向小鼠IP注射萤光素底物。十分钟之后,用异氟烷和经过定量的生物发光(总通量,表示为光子每秒[p/s])将小鼠麻醉。根据IACUC标准,当小鼠开始表现出移植物抗宿主疾病(GVHD)的迹象时(评估为体重减轻≥20%),实验结束。
肿瘤生长的统计分析
使用双向方差分析(ANOVA)、然后使用图基事后检验对肿瘤体积随时间推移的结果进行分析以用于多次比较。当p<0.05时,差异被视为在统计学上具有显著性。使用GraphPad Prism软件(版本8)执行统计分析。
结果
在REGN1979存在或不存在的情况下施用REGN5837的抗肿瘤功效
携带NALM-6-luc肿瘤的免疫缺陷NSG小鼠接受如本文所描述的抗体或非桥接对照的IP注射。
在携带肿瘤的小鼠中,与非桥接对照bsAb(非TAAxCD28和非TAAxCD3 bsAb)相比,在植入后第23天在0.04mg/kg REGN1979存在的情况下用0.04mg/kg、0.4mg/kg和4mg/kgREGN5837治疗导致了对肿瘤生长的统计学显著抑制(分别为p<0.05、p<0.01和p<0.001)(图6)。在植入后第20天,针对0.4mg/kg和4mg/kg组(两个组均为p<0.05)观察到对肿瘤生长的显著抑制。与非桥接对照bsAb相比,REGN5837(4mg/kg)和REGN1979(0.04mg/kg)单一疗法均未显著减少肿瘤生长。任何REGN5837+REGN1979组合剂量与bsAb单一疗法之间的差异未达到统计学显著性。在整个给药期期间,在用非桥接对照bsAb给药时观察到肿瘤快速生长,并且在第23天,将所有小鼠安乐死。在所有组中,在实验结束时在至少一只小鼠中观察到GVHD(评估为体重减少≥20%)。
在使用不同组小鼠的独立实验中,在以下时间点采集血液以确定血清抗体浓度:在第7天给药后1小时、4小时、在第14天和第21天给药前1小时和给药后4小时以及在第29天一次。在第14天和第21天给药前1小时确定给药期期间血清中抗体的低谷浓度。在存在0.04mg/kg REGN1979的情况下施用0.04mg/kg、0.4mg/kg和4mg/kg REGN5837剂量与范围分别为定量限值以下(BLQ)到0.1μg/mL、1.6μg/mL到2.3μg/mL以及16.5μg/mL到21.1μg/mL的在血清中的REGN5837的低谷浓度相关。在所有情况下,血清中REGN1979的低谷浓度是BLQ(数据未示出)。
结论
在存在0.04mg/kg REGN1979的情况下0.04mg/kg、0.4mg/kg和4mg/kg REGN5837剂量有效抑制小鼠中的NALM-6B细胞白血病肿瘤生长。相对于对照,利用4mg/kg REGN5837或0.04mg/kg REGN1979单一疗法未观察到显著抑制肿瘤。
实例12:CD22细胞+人PBMC+/-CD22xCD28共刺激双特异性抗体(固定CD22xCD28、滴定CD20xCD3)上的基于FACS的细胞毒性
材料与方法
在靶向Nalm6或WSU-DLCL2细胞的96小时细胞毒性测定中评估CD20xCD3靶向的杀伤的CD22xCD28增强。简而言之,将人PBMC以1×10
通过将细胞与同CD2、CD4、CD8和CD25直接缀合的抗体一起温育来评估T细胞活化。将表达CD25的CD8+细胞的百分比报告为T细胞活化度量。另外,随着T细胞的增殖,对CellTraceViolet进行稀释,这导致如FACS所测量的MFI较低。因此T,将细胞增殖报告为CD8+T细胞上的CellTraceViolet MFI的减少或已减少CellTraceViolet MFI的CD8+细胞的百分比。
收集来自此测定的上清液以用于细胞因子水平的分析。按照制造商的说明书使用流式细胞珠阵列(CBA)试剂盒对IL 17a、IFNγ、TNFα、IL-10、IL-6、IL-4和IL-2的浓度进行分析。从试剂盒标准品所生成的曲线中插入细胞因子水平并将细胞因子水平报告为pg/mL。在Prism软件中使用4参数非线性回归分析计算靶细胞杀伤、T细胞活化、增殖和细胞因子水平以及最大细胞因子水平的EC50值。
结果、汇总和结论:
测试抗CD20xCD3双特异性抗体REGN1979与共刺激CD22xCD28抗体或者单臂CD28或同种型对照抗体组合地诱导原初人T细胞杀伤表达人CD20和CD22的靶细胞的能力。
REGN1979活化并指导人T细胞以剂量依赖性方式杀伤Nalm6(图7)或WSU-DLCL2(图8)细胞。将固定浓度的CD22xCD28双特异性抗体添加到REGN1979使REGN1979针对Nalm6细胞的细胞毒性功效(EC50)与利用单臂CD28或同种型对照抗体的REGN1979相比增强4.7-5.2倍(表26)或针对WSU-DLCL2细胞与单独的REGN1979相比增强17.5倍(表27)。
如分别通过CD8+细胞上的CD25上调或CellTrace Violet稀释测量的,观察到的由REGN1979介导的靶细胞裂解与T细胞活化和增殖相关(图7、图8)。将固定浓度的CD22xCD28双特异性抗体添加到REGN1979使REGN1979诱导的T细胞活化和增殖的效力在Nalm6细胞存在的情况下与利用单臂CD28或同种型对照抗体的REGN1979相比分别增强2.3-2.6倍和5.4-7.1倍(表26)并且在WSU-DLCL2细胞存在的情况下与单独的REGN1979相比分别增强8.2倍和16.1倍(表27)。
在利用人PBMC和WSU-DLCL2细胞的测定中,REGN1979诱导人细胞因子的释放。与由仅REGN1979诱导的细胞因子释放相比,在固定浓度的CD22xCD28存在的情况下利用REGN1979观察到的释放的细胞因子有所增强(表28、图9)。
总之,与单独地或与对照抗体组合地利用CD20xCD3所观察到的相比,共刺激增加了靶向的细胞毒性、T细胞活化和细胞因子释放的效力。
表列数据汇总:
表26:利用Nalm6靶标的细胞毒性和T细胞活化的EC50值(1个实验)
表27:利用WSU-DLCL2靶标的细胞毒性和T细胞活化的EC50值(2次实验的平均值)
表28:来自WSU-DLCL2细胞毒性测定的细胞因子释放(2个实验的平均值)
实例13:NHL+人PBMC+/-CD22xCD28刺激(固定CD22xCD28、滴定CD20xCD3)上的基于FACS的细胞毒性
实验程序
在靶向在人基质细胞(HS-5)存在的情况下从具有自体PBMC的原代NHL患者活检中分离出来的NHL细胞的96小时细胞毒性测定中评估CD20xCD3靶向杀伤的CD22xCD28增强。简而言之,将HS-5细胞以每孔5000个细胞平板接种在平底96孔平板中并温育过夜。第二天,用1uM荧光跟踪染料CellTrace Violet对来自NHL患者的PBMC进行标记。将骨髓和标记的PBMC(效应细胞/靶细胞比为10:1)平板接种在具有基质细胞的孔中并在37℃下与CD20xCD3双特异性抗体REGN1979的连续稀释液(浓度范围:6.7nM到10.7pM)和2.5ug/ml(16.7nM)固定浓度的CD22xCD28共刺激分子REGN5837或单臂对照CD28双特异性(REGN5678)一起温育96个小时。从平板收获细胞并在FACS BD LSRFortessa-X20上通过FACS进行分析。关于FACS分析,利用抗体混合物(CD45、CD19、CD4、CD8、CD25)和可固定活/死近IR反应性染料(英杰公司)对细胞进行染色。紧接在FACS分析之前将20,000个计数珠添加到每个孔中,并且针对每个样品收集10,000个珠。为了评估杀伤的特异性,在活CD45+Violet阴性CD19+群体上对细胞进行门控。基于相对于未处理孔中的靶细胞数量归一化的经过处理的孔中的靶细胞数量来计算存活率。
将T细胞门控为活CD45+Violet标记的CD4+或CD8+群体。将表达CD25的CD8+和CD4+细胞的百分比报告为T细胞活化的度量。另外,随着T细胞的增殖,对CellTraceViolet进行稀释,这导致如FACS所测量的MFI较低。因此,将T细胞增殖报告为CD8+和CD4+T细胞上的CellTraceViolet的MFI的减少。
在Prism软件中使用4参数非线性回归分析计算靶向杀伤以及T细胞活化和增殖的EC50值。
结果汇总和结论:
测试抗CD20xCD3双特异性抗体REGN1979与共刺激CD22xCD28抗体或单臂CD28对照抗体组合地诱导原初自体T细胞杀伤来自患者骨髓的NHL细胞的能力。
REGN1979活化并指导人T细胞以剂量依赖性方式耗竭NHL。与利用单臂CD28对照抗体或没有共刺激对照的REGN1979相比,将固定浓度的CD22xCD28双特异性抗体添加到REGN1979使REGN1979的细胞毒性功效(EC50)提高了2.3和3.5倍(表29)。
如分别通过CD8+和CD4+细胞上的CD25上调或CellTrace Violet稀释测量的,观察到的由REGN1979介导的靶细胞裂解与T细胞活化和增殖相关。与利用单臂CD28或没有共刺激对照的REGN1979相比,将固定浓度的CD22xCD28双特异性抗体添加到REGN1979使REGN1979诱导的T细胞活化和增殖的效力分别增强2.8到4.2倍和2.8-4.8倍(表29和图10)。
总之,与利用CD20xCD3与对照抗体组合地观察到的相比,共刺激增加了靶向的细胞毒性和T细胞活化的效力。
表列数据汇总:
表29:细胞毒性和T细胞活化的EC50值
实例14:在弥漫性大B细胞淋巴瘤(DLBCL)模型中单独的和与REGN1979组合的REGN5837的抗肿瘤功效的体外表征和体内评估
研究引言和结果汇总的材料与方法
进行体外和体内研究以评估:
(1)REGN5837通过桥接CD28
(2)在存在或不存在0.4mg/kg或4mg/kg REGN1979的情况下向携带DLBCL肿瘤的免疫缺陷NSG小鼠施用的CD22xCD28 bsAb REGN5837的抗肿瘤功效。
在一定的浓度范围内组合地测试REGN5837和REGN1979,以评估REGN5837对REGN1979介导的针对人DLBCL细胞系(WSU-DLCL2)的T细胞细胞毒性、后期T细胞活化标志物(CD25)的上调、T细胞增殖和来自原代人T细胞的细胞因子释放的影响。REGN5837以浓度依赖性方式增强REGN1979介导T细胞细胞毒性、CD4
向免疫缺陷NSG小鼠(每组n=6到7)皮下(SC)植入比率为1:1的WSU-DLCL2细胞和人PBMC。将1mg/kg REGN5837与0.4mg/kg或4mg/kg剂量的REGN1979组合的抗肿瘤功效与REGN5837和REGN1979单一疗法并且与非桥接IgG4
总之,当在体外在一定的浓度范围内组合地测试REGN5837和REGN1979时,REGN5837增强了REGN1979在CD22
表30:REGN5837使用人PBMC对REGN1979介导的T细胞活化(通过针对靶细胞的细胞毒性、CD25表达和T细胞增殖测量)的影响汇总
实例15:REGN5837对REGN1979介导的人T细胞活化的影响的评估以及在WSU-DLCL2细胞存在的情况下在一定的浓度范围内组合地测试REGN5837和REGN1979。
在体内和体外都进行了研究以通过测量以下参数来评估人CD22xCD28 bsAbREGN5837在次有效剂量的人CD20xCD3 bsAb(REGN1979)存在或不存在的情况下在植入人PBMC和WSU-DLCL2细胞后的NSG小鼠中的抗肿瘤功效:a)针对CD22
材料与方法
细胞系
WSU-DLCL2:WSU-DLCL2是从41岁白人男性的胸腔积液分离出来的人DLBCL细胞系(莱布尼兹研究所-DSMZ,目录号ACC 575)。
人PBMC
关于细胞毒性、T细胞活化、T细胞增殖和细胞因子释放,从纽约血液中心(the NewYork Blood Center)获得来自人供体的leukopak细胞(供体号1500A)。
关于体内小鼠实验,从ReachBio公司获得人PBMC(目录号0500-401)。
实验设计
在一定的浓度范围内组合地测试REGN5837和REGN1979以评估REGN5837对REGN1979介导的针对WSU-DLCL2细胞的T细胞细胞毒性、T细胞增殖、后期T细胞活化标志物(CD25)的细胞表面表达以及来自人T细胞的细胞因子释放的影响。如本文所描述的,确定靶细胞杀伤、T细胞活化和T细胞增殖的百分比。
如本文所描述的,评估使用WSU-DLCL2细胞和PBMC在DLBCL模型中单独的以及与REGN1979组合的REGN5837的抗肿瘤功效。参见表31。
REGN5837对REGN1979介导T细胞活化的效力的影响的体外评估
按照制造商推荐的方案使用50mL SepMate
使用PBMC的基于流式细胞术的T细胞活化测定
使用WSU-DLCL2靶细胞和人PBMC作为效应细胞,计算REGN5837增强通过同种异体初次刺激或利用通过REGN1979提供的“信号1”介导的T细胞活化的能力。如本文所描述的,对PBMC进行淋巴细胞富集。如本文所描述的,将靶细胞和效应细胞与测试物品和对照抗体温育。如本文所描述的,执行流式细胞术以评估T细胞细胞毒性、增殖和T细胞活化标志物的上调。另外,如本文所描述,评估REGN5837对REGN1979介导的细胞因子释放的影响。测试非TAAxCD28 bsAb(含有与REGN5837相同的CD28结合臂以及非结合臂)作为REGN5837的非桥接对照。
PBMC的淋巴细胞富集
以1x10
PBMC和靶细胞与测试物品的温育
收获淋巴细胞富集的PBMC并用1μM Violet Cell Tracker荧光跟踪染料进行标记。用1μM荧光染料Vybrant CFDA-SE对WSU-DLCL2靶细胞进行标记。
随后,将标记染料的PBMC与标记染料的靶细胞以5:1的比率平板接种在在圆底96孔平板中(WSU-DLCL2为5x10
将平板接种的PBMC和靶细胞与测试物品或其范围为12.9pM到100nM(REGN5837或非TAAxCD28)和4.8fM到10nM(REGN1979或非TAAxCD3)的最终浓度下的对照在37℃下温育72小时。
流式细胞术分析
在与测试物品和对照温育之后,用活/死染色剂并且用针对CD2、CD4、CD8和CD25的荧光团标记的抗体的混合物对染料标记的细胞进行染色。添加计数珠(20μL每孔),紧接着在BD Celesta流式细胞仪上进行样品分析。使用流式细胞术数据来确定靶细胞存活率、T细胞增殖和T细胞活化标志物的上调。使用GraphPad Prism软件从9点剂量-应答曲线上的四参数逻辑方程确定EC
靶细胞存活率
将每个实验条件下的活靶细胞的百分比计算为相对于收集的珠的数量/孔归一化的活的CFDA-SE标记的靶细胞数量/孔。将靶细胞存活率百分比确定为任何实验条件下的活靶细胞的数量与无抗体对照条件下的活靶细胞(在仅PBMC存在的情况下的靶细胞)的数量之比。
在以此方式报告的情况下,将每个实验条件下的针对靶细胞的细胞毒性百分比确定为100减去存活率百分比(如上文所描述的那样计算)。
CD4
通过在活CD2
CD4
通过计算分裂的细胞占CD4
细胞因子释放分析
根据制造商的说明书使用BD流式细胞珠阵列人Th1/Th2/Th17细胞因子试剂盒对细胞培养上清液内的细胞因子(IL-4、IL-2、IL-6、IL-10、TNF-α、IFN-γ和IL-17Α)的水平进行定量。
使用WSU-DLCL2细胞异种移植物的DLBCL体内模型
在所有实验中使用雌性NSG小鼠。向所有小鼠SC植入WSU-DLCL2 B细胞淋巴瘤细胞并用抗体进行IP给药。在整个研究期间,使用卡尺每周测量肿瘤体积若干次。对于所有的实验,将小鼠收容在标准条件下的再生元公司动物设施中。所有实验根据再生元公司的机构动物护理和使用委员会(IACUC)的准则执行。
WSU-DLCL2细胞培养条件和肿瘤植入
从莱布尼兹研究所-DSMZ获得WSU-DLCL2细胞系,并将其维持在具有补充有盘尼西林、链霉素、谷氨酰胺和1mM HEPES的10%FBS的RPMI-1640培养基中。
收集WSU-DLCL2细胞(3x10
用于肿瘤测量的抗体给药
在用测试物品或对照给药之前,将小鼠分配到根据肿瘤负荷分层的组。
在植入后第1天、第8天和第15天以表31所陈述的剂量通过IP注射以单一疗法或组合地施用抗体(REGN5837、REGN1979、REGN5671[非TAAxCD28非桥接对照bsAb]或H4sH17664D[非TAAxCD3非桥接对照bsAb])。
表31:用于评估肿瘤生长和存活率的实验设计
肿瘤测量和所指定终点
使用肿瘤X和Y直径的卡尺测量结果(长度和宽度垂直测量结果)随时间推移监测肿瘤生长。计算肿瘤体积(X*Y*[X/2],其中X是较短直径)。当肿瘤达到指定的肿瘤终点(肿瘤直径>20mm或肿瘤溃疡)时,将小鼠安乐死。此终点符合IACUC标准。
肿瘤生长和存活率的统计分析
使用双向方差分析(ANOVA)、然后使用图基事后检验对肿瘤体积随时间推移的结果进行分析以用于多次比较。跨所有组使用Mantel-Cox测试分析随时间推移的存活率结果,并且运行Mantel-Cox测试以进行逐个单独组的比较。当p<0.05时,差异被视为在统计学上具有显著性。使用GraphPad Prism 8软件执行统计分析。
结果
REGN5837对REGN1979介导针对WSU-DLCL2靶细胞的T细胞细胞毒性和来自人PBMC的细胞因子释放的能力的影响
在一定的浓度范围内组合地测试REGN5837和REGN1979,以评估REGN5837对REGN1979介导的针对WSU-DLCL2细胞的细胞毒性、T细胞活化、T细胞增殖以及来自人PBMC的T细胞的细胞因子释放的影响,如先前所描述的。表30示出了REGN5837对REGN1979介导的针对WSU-DLCL2细胞的细胞毒性、T细胞活化和T细胞增殖的影响。表32中汇总了展现出REGN5837对细胞因子释放的影响的来自2个人供体的数值结果。
REGN5837对REGN1979介导的细胞毒性和人T细胞增殖的影响
通过评估REGN1979介导的针对WSU-DLCL2靶细胞的细胞毒性、REGN1979介导的T细胞活化以及REGN1979介导的对来自人PBMC的人CD4
在REGN5837存在的情况下REGN1979介导的来自人PBMC的细胞因子释放
评估增加浓度的REGN5837对REGN1979介导的来自人PBMC的细胞因子释放的效力和最大应答的影响。在人PBMC和WSU-DLCL2细胞存在的情况下,增加浓度的REGN5837以浓度依赖性方式增强REGN1979介导的IL-2、IL-4、IL-6、IL-10、TNF-α、IFN-γ和IL-17Α释放的最大水平(图12)。此外,增加浓度的REGN5837示出了增强REGN1979介导细胞因子释放的效力的趋势。图12和表32中汇总了EC
表32
在次有效剂量的REGN1979存在或不存在的情况下施用REGN5837的抗肿瘤功效
携带WSU-DLCL2肿瘤的免疫缺陷NSG小鼠接受抗体或非桥接对照的IP注射,如本文先前所描述的。
在携带肿瘤的小鼠中,在第28天(最后抗体剂量后6天),与非桥接对照bsAb(非TAAxCD28和非TAAxCD3 bsAb)相比,在0.4mg/kg或4mg/kg REGN1979存在的情况下用1mg/kgREGN5837治疗导致统对肿瘤生长的计学上显著抑制(图13A和13B)。在第46天,相对于REGN1979单一疗法,1mg/kg REGN5837和0.4mg/kg REGN1979的组合导致肿瘤体积显著减小。
在第28天,相对于非桥接对照,0.4mg/kg和4mg/kg REGN1979单一疗法导致适度肿瘤抑制,然而REGN5837单一疗法无效果。在整个给药期期间,在用非桥接对照bsAb给药时观察到肿瘤快速生长,并且在第125天,将所有小鼠安乐死。
Mantel-Cox测试跨所有组检测存活率的统计学上显著差异(p=0.0001),并且执行另外的Mantel-Cox测试以进行逐个组的比较。与用非桥接对照抗体给药的小鼠(无存活率)(图14)相比,观察到用1mg/kg REGN5837与0.4mg/kg或4mg/kg REGN1979组合地给药的小鼠的存活率显著增加(分别为85%和70%的存活率)。
此外,与用REGN5837或REGN1979单一疗法给药的小鼠相比,观察到用1mg/kgREGN5837与0.4mg/kg或4mg/kg REGN1979组合地给药的小鼠的存活率显著增加。
本发明不应限于本文所述的具体实施例的范围。事实上,除了本文所述的那些修改之外,本发明的各种修改对于本领域的技术人员来说也将从前面的描述和附图中变得显而易见。此类修改旨在落入所附权利要求的范围内。
- 双特异性抗CD28 X抗CD22抗体以及其用途
- 双特异性抗PSMA X抗CD28抗体及其用途