一种蓝光介导调节质粒及其构建方法和应用
文献发布时间:2023-06-19 12:04:09
技术领域
本发明涉及微生物技术领域,具体涉及一种蓝光介导调节质粒及其构建方法和应用。
背景技术
基因表达过程的传统调节方法多为外源性的化学诱导剂结合可溶性转录因子来实现对基因表达的人工控制,该方式是上个世纪推动生物技术、合成生物学及医学研究快速发展的主要动力之一。虽然这些传统调控方式可以初步实现基因表达的时间和表达量控制,但是其有限的可逆性及其以诱导剂为基础的运输调节机制使得其存在着种种局限性和缺陷,例如:其潜在的脱靶效应、转运过程延迟及毒性。在理想情况下,能够快速、精确地随意“开启”或“关闭”的生物调控元件,将有效提高分析和调控复杂生物基因网络的能力,光调节方式的时间调节精度可达到毫秒的范围。
来源于山葡萄红杆菌(Erythrobacter litoralis)的EL222光敏蛋白可有效实现光调控操作,EL222由222个氨基酸组成,N端为光氧化调控域(Light-Oxygen-Voltage,LOV),C端为螺旋-转角-螺旋(helix-turn-helix,HTH)DNA结合域,LOV的C末端是连接LOV和HTH的Jα螺旋。LOV的核心是Per-Arnt-SIM(PAS)结构域,可以与黄素单核苷酸(FMN)结合。属于光诱导同源二聚化的系统。在蓝光照射下,FMN和LOV的结合导致Jα螺旋从PAS摇摆,从而释放HTH,使EL222二聚并与DNA结合。在黑暗中,N端LOV结构域抑制DNA结合的C端HTH结构域时,EL222自发地逆转,从而迅速使EL222失活。目前,EL222蛋白已初步在大肠杆菌及酿酒酵母中得到验证,但尚未在毕赤酵母中报道其适用性。
毕赤酵母作为真核生物,具有真核表达系统的许多优点,如蛋白质加工、折叠、翻译后修饰等,而且更容易操作。与哺乳动物细胞培养和杆状病毒等真核表达系统相比,毕赤酵母表达系统具有更快捷、更简单、更廉价、表达水平更高等优点。毕赤酵母作为一种酵母,具有与酿酒酵母相似的分子和遗传操作优势,其外源蛋白表达量是酿酒酵母的十倍甚至上百倍。上述诸多优势使得毕赤酵母成为真核蛋白表达系统的首选。然而毕赤酵母的诱导调节方式仍处于外源性化学诱导剂的阶段,其最常使用的诱导剂甲醇毒性强且易燃,不利于毕赤酵母在食品、医药、农产品加工等领域的应用。因此,开发一种新型的毕赤酵母的光调节方式,可有效解决传统化学诱导剂的毒性、不稳定性及转运延迟性等缺陷,拓展毕赤酵母表达调控的方式方法,成为本领域迫切需要解决的一个技术问题。
发明内容
为了解决现有技术中存在的问题,本发明的目的在于提供一种蓝光调节启动子PC120,可以快速、精准的实现对目的基因的蓝光诱导表达。
本发明的目的还在于提供一种蓝光介导调节质粒及其构建方法和应用,可用于毕赤酵母蓝光诱导表达,该调节方式无毒无害、响应快速、操作简便、可逆性好、时空分辨率高。
为了实现上述发明目的,本发明提供以下技术方案:
本发明提供了一种蓝光调节启动子PC120,所述启动子的核酸序列如SEQ ID NO.1所示。
优选的,所述启动子可被蓝光蛋白SV40-VP-EL识别并结合。
优选的,所述蓝光蛋白SV40-VP-EL的氨基酸序列如SEQ ID NO.2所示。
优选的,所述蓝光蛋白SV40-VP-EL的核酸序列如SEQ ID NO.3所示。
本发明还提供了一种蓝光介导调节质粒,所述质粒包括上述的蓝光调节启动子PC120和转录翻译蓝光蛋白的融合基因PGAP-SV40-VP-EL。
优选的,所述融合基因PGAP-SV40-VP-EL的核酸序列如SEQ ID NO.4所示。
本发明还提供了上述蓝光介导调节质粒的构建方法,所述构建步骤包括:
以pAO815质粒为骨架,将融合基因PGAP-SV40-VP-EL替换pAO815质粒AOX1promoter以及AOX1 promoter和AOX1 terminator之间的序列,得到质粒pGSVEA;再将包含启动子PC120的另一融合基因替换质粒pGSVEA的CGTTCGTTTGTGC序列即可。
优选的,所述包含启动子PC120的融合基因还包括报道基因和/或目的基因。
优选的,所述融合基因通过GibsonAssembly的方式定点组装到pAO815质粒上。
本发明还提供了上述蓝光介导调节质粒和/或上述蓝光介导调节质粒的构建方法在毕赤酵母诱导表达中的应用。
与现有技术相比,本发明的技术方案的有益效果如下:
本发明为毕赤酵母的诱导表达方法提供了一个新型的蓝光诱导调节方式。本发明采用光调节方式对毕赤酵母进行诱导表达,无毒无害、响应快速、操作简便、可逆性好、时空分辨率高,推动了毕赤酵母在食品、医药、农产品加工等领域的广泛应用。
本发明仅需一个质粒即可实现对目的基因的蓝光诱导表达,拓展了毕赤酵母表达调控的方式方法。
附图说明
图1为蓝光调节质粒的机制示意图。
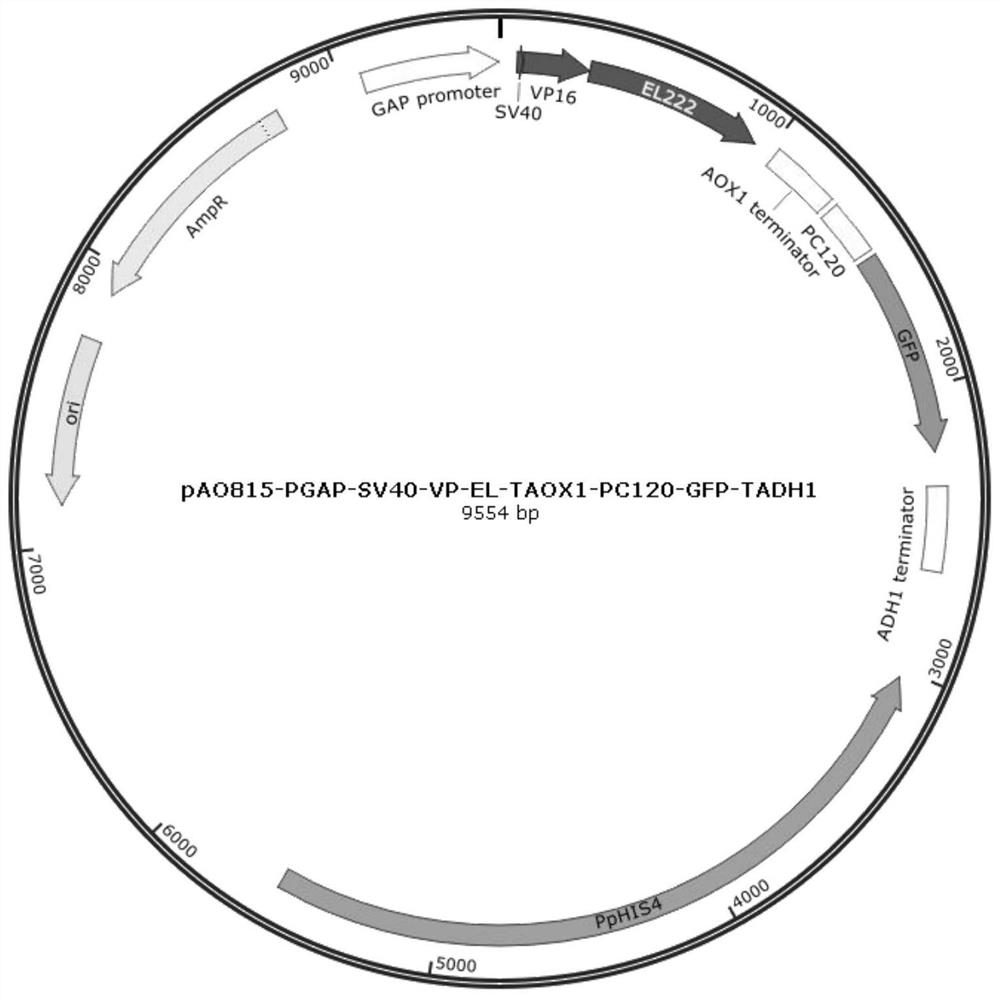
图2为pGSVEACA载体图,图中PpHIS4是毕赤酵母整合位点及筛选标记,AmpR是抗生素抗性,ori是复制起始位点,GAP promoter是毕赤酵母GAP启动子,SV40是核定位信号,VP16是转录激活域,EL222是蓝光响应蛋白,AOX1 terminator是酵母AOX1终止子,PC120是蓝光调节启动子,GFP是绿色荧光蛋白,ADH1 terminator是酿酒酵母ADH1终止子。
图3为pGSVEACA载体转化毕赤酵母在MD平板上的克隆。
图4为蓝光调节工程菌基因组PCR验证阳性克隆。
图5为蓝光调节工程菌分别在蓝光条件和黑暗条件下诱导12h的MD平板,左为黑暗条件,右为蓝光条件。
图6为蓝光调节工程菌分别在蓝光条件和黑暗条件下诱导的MD摇瓶培养,(A)为蓝光诱导1h后实验组Full Light与对照组Full Dark的荧光强度;(B)为蓝光诱导4h后实验组Full Light与对照组Full Dark的荧光强度;(C)为蓝光诱导7h后实验组Full Light与对照组Full Dark的荧光强度;(D)为蓝光诱导10h后实验组Full Light与对照组Full Dark的荧光强度;(E)为实验组Full Light与对照组Full Dark在蓝光诱导期间每小时所取样品的荧光强度平均值的趋势图。
具体实施方式
本发明提供了一种蓝光调节启动子PC120,所述启动子的核酸序列如SEQ ID NO.1所示。本发明启动子PC120可被蓝光蛋白SV40-VP-EL识别并结合,从而快速、精准的实现对目的基因的蓝光诱导表达。
本发明蓝光蛋白SV40-VP-EL的氨基酸序列如SEQ ID NO.2所示,核酸序列如SEQID NO.3所示。本发明中蓝光蛋白SV40-VP-EL的核酸序列也可以为编码蛋白SV40-VP-EL的其余核酸序列变种。
本发明提供了一种蓝光介导调节质粒,包括上述蓝光调节启动子PC120和转录翻译蓝光蛋白的融合基因PGAP-SV40-VP-EL。本发明融合基因PGAP-SV40-VP-EL中的PGAP表示毕赤酵母GAP启动子GAP promoter,SV40表示核定位信号SV40,VP表示转录激活域VP16、EL表示蓝光响应蛋白EL222。
在本发明中,PGAP的核酸序列为毕赤酵母GS115 ATCC:20864的GAP1启动子序列(GenBank AccessionNo:CP014716 REGION:809514..809990)。
本发明融合基因PGAP-SV40-VP-EL的核酸序列如SEQ ID NO.4所示。
本发明还提供了上述蓝光介导调节质粒的构建方法,包括:以pAO815质粒为骨架,将融合基因PGAP-SV40-VP-EL替换pAO815质粒AOX1 promoter以及AOX1 promoter和AOX1terminator之间的序列,得到质粒pGSVEA;再将包含启动子PC120的另一融合基因替换质粒pGSVEA的CGTTCGTTTGTGC序列,得到蓝光调节质粒pGSVEACA。本发明构建得到的pGSVEACA质粒,见图2。
本发明得到的质粒pGSVEA导入毕赤酵母后可以转录翻译得到蓝光蛋白SV40-VP-EL,优选的,所述毕赤酵母为毕赤酵母GS115。
本发明得到的蓝光调节质粒pGSVEACA可以转录翻译得到蓝光蛋白SV40-VP-EL,在蓝光照射下,蓝光蛋白SV40-VP-EL中的光敏蛋白EL222的N端(光氧化调控域)与C端(螺旋-转角-螺旋DNA结合域)相互作用,使EL222二聚并与启动子PC120结合,开启含启动子PC120的另一融合基因的转录。
在本发明中,包含启动子PC120的另一融合基因为PC120-GFP-TADH1,其中PC120表示蓝光调节启动子PC120,GFP表示绿色荧光蛋白GFP,TADH1表示酵母ADH1终止子ADH1terminator,TADH1的核酸序列为酿酒酵母Y169的ADH1终止子序列(GenBank AccessionNo:CP033484 REGION:156786..157080)。本发明中绿色荧光蛋白GFP可以替换为其他报告基因,也可以替换为其他目的基因。
本发明融合基因PC120-GFP-TADH1的核酸序列如SEQ ID NO.5所示。
本发明中,两段融合基因可以通过Gibson Assembly的方式定点组装到pAO815质粒上。本发明对具体的组装方式不作限定。
本发明还提供了上述蓝光介导调节质粒和/或蓝光介导调节质粒的构建方法在毕赤酵母诱导表达中的应用。本发明将蓝光调节质粒pGSVEACA转化导入毕赤酵母中,即可实现对目的基因的蓝光诱导表达,拓展了毕赤酵母表达调控的方式方法,推动了毕赤酵母在食品、医药、农产品加工等领域的广泛应用。
下面将结合本发明中的实施例,对本发明中的技术方案进行清楚、完整地描述。显然,所描述的实施例仅仅是本发明一部分实施例,而不是全部的实施例。基于本发明中的实施例,本领域普通技术人员在没有做出创造性劳动前提下所获得的所有其他实施例,都属于本发明保护的范围。
实施例1
本实施例对基因序列进行了优化。
将NCBI中检索到的VP16及EL222基因序列和蛋白序列,按照毕赤酵母密码子偏好性进行优化,重新合成DNA序列。
具体的优化方法为:根据基因合成公司金斯瑞密码子优化软件OptimumGene
实施例2
本实施例以毕赤酵母的常用质粒pAO815为骨架设计并构建了蓝光调节质粒pGSVEACA。
具体的构建方法为:
将实施例1中优化获得的PGAP-SV40-VP-EL基因序列进行全合成,然后使用Gibsonassembly技术替换pAO815质粒AOX1 promoter以及AOX1 promoter和AOX1 terminator之间的序列,得到质粒pGSVEA;
将实施例1中优化获得的PC120-GFP-TADH1基因序列进行全合成,然后使用Gibsonassembly技术替换质粒pGSVEA的CGTTCGTTTGTGC序列,得到蓝光调节质粒pGSVEACA。
本实施例中,PGAP-SV40-VP-EL基因序列中并未包含终止子序列,所以需要借用pAO815质粒骨架上的AOX1 terminator,从而构建完整的SV40-VP-EL开放阅读框,实现SV40-VP-EL的组成型表达。
本实施例中,为了让单个质粒实现蓝光诱导的效果,需要在构建生成的质粒pGSVEA基础上,将PC120-GFP-TADH1基因序列插入到PGAP-SV40-VP-EL的下游,故采用PC120-GFP-TADH1基因序列替换质粒pGSVEA的CGTTCGTTTGTGC序列。
实施例3
本实施例对蓝光调节质粒pGSVEACA进行转化、诱导表达与分析。
(1)质粒的提取与定量检测
1)取10mL菌体过夜培养物,13400g离心1min弃上清液收沉淀。
2)依次加入500μL溶液Ⅰ,溶液Ⅱ及溶液Ⅲ,立即温和地上下翻转6~8次后静置5min,13400g离心10min。
其中溶液Ⅰ的组成为25mM Tris-HCl(pH=8.0),10mM EDTA,50mM葡萄糖;
溶液Ⅱ的组成为250mM NaOH,1%(W/V)SDS;
溶液Ⅲ的组成为3M醋酸钾,5M醋酸。
3)将上一步收集的上清液加入过滤柱中,13400g离心1min后加入450μL异丙醇混合均匀。而后加入吸附柱中,13400g离心1min后弃去废液。
4)将700μL漂洗液加入吸附柱中,13400g离心1min后弃去废液。
5)吸附柱除净漂洗液后,向吸附膜中央滴加150μL的去离子水,静置2min后13400g离心2min。
6)收集管内液体并测定质粒pGSVEACA浓度,-20℃保存备用。
(2)质粒pGSVEACA的线性化和回收
1)向样品中加入1/10体积的3M乙酸钠溶液和2.5倍体积的无水乙醇混匀,于-20℃放置1h。而后12000rpm离心10min,弃上清。
2)2.5倍体积的预冷75%乙醇重悬沉淀,12000rpm离心10min,弃上清。并置于超净工作台吹15min。
3)向管底滴加25μL的去离子水重悬,并测定质粒浓度,-20℃保存备用。
(3)毕赤酵母GS115的电转化
1)在制备好的毕赤酵母GS115感受态细胞中加入5~10μg线性化质粒pGSVEACA并混匀,转移到预冷的新电转杯(0.2cm规格)中,冰浴5min。
2)快速取出电转杯并擦干,在电压为1.5kV、电容为25μF、电阻为200Ω的参数下电击,立即加入1mL预冷的1M山梨醇,吸取液体并转移至新的1.5mL无菌离心管,于30℃静置孵育1.5h。
3)12000rpm离心20s后弃去大部分上清,重悬菌体后涂布到MD筛选平板,28℃恒温培养2d至出现肉眼可见的单菌落。酵母转化结果见图3。
(4)毕赤酵母转化子的诱导表达与荧光强度定性分析
随机挑取转化出的酵母转化子至接种于5mL MD液体培养基中28℃摇床培养19h。保菌并抽取基因组DNA,设计基因阳性检测引物进行PCR扩增,获得阳性菌株。本发明PCR所用引物见表1。
表1 PCR引物
表1中GAP-F序列如SEQ ID NO.6所示,ADH-R序列如SEQ ID NO.7所示,AOX-R序列如SEQ ID NO.8所示,AOX-F序列如SEQ ID NO.9所示。
PCR扩增结果见图4,表明挑取培养的酵母转化子中含有目的质粒。
根据PCR检测结果,选取3~5个阳性克隆划线于MD培养基上,先于28℃黑暗环境下培养24h后对实验组进行蓝光诱导孵育。
蓝光诱导24h后,将实验组与对照组置于475nm的激发光下进行荧光强度的定性比较,对比效果见图5,可见,蓝光诱导后的菌株在475nm的激发光下发绿色荧光,表明通过蓝光诱导后的毕赤酵母细胞内转录翻译得到了绿色荧光蛋白,PC120启动子可以成功识别并结合蓝光蛋白SV40-VP-EL,精准开启报道基因的转录翻译。
(5)毕赤酵母转化子的流式细胞术分析
根据PCR检测结果,选取3~4个阳性克隆,各取50-100μL培养液转接至50mLMD液体培养基中,28℃黑暗环境下,摇床培养至菌体OD 600nm达到0.8~1.2后进行蓝光诱导表达。蓝光诱导每隔1h对实验组和对照组进行取样,共诱导10h。
对所取样品进行预处理以便进行流式细胞术分析,具体操作为:样品菌液在4℃下以5000rpm离心1min,弃上清后用PBS重悬并稀释至肉眼不可见的状态,取适当体积的样品稀释液通过300目的滤网过滤后存于流式分析管中以备进行流式细胞术分析。
对实验组及对照组的样品进行流式细胞术分析后所得出的荧光强度与诱导时间的关系见表2和图6。
表2菌体在蓝光诱导和黑暗环境下不同时间的荧光强度
由表2和图6可知,蓝光持续照射诱导8小时后GFP的荧光强度达到最高,其强度是同时期对照组的16.4倍。直方图中呈现出单一且均匀的峰,说明蓝光可对携带有pGSVEACA质粒的GS115细胞实现均匀的诱导效应。并且实验组和对照组直方图的交叉重叠部分并不多,说明诱导强度较为良好。整体而言,该流式细胞术结果说明pGSVEACA质粒可有效实现GS115细胞对蓝光高效均一的响应。
以上所述仅是本发明的优选实施方式,应当指出,对于本技术领域的普通技术人员来说,在不脱离本发明原理的前提下,还可以做出若干改进和润饰,这些改进和润饰也应视为本发明的保护范围。
序列表
<110> 华中科技大学
<120> 一种蓝光介导调节质粒及其构建方法和应用
<160> 9
<170> SIPOSequenceListing 1.0
<210> 1
<211> 190
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 1
taggtagcct ttagtccatg cgttataggt agcctttagt ccatgcgtta taggtagcct 60
ttagtccatg cgttataggt agcctttagt ccatgcgtta taggtagcct ttagtccatg 120
aagcttagac actagagggt atataatgga agctcgactt ccagcttggc aatccggtac 180
tgttggtaaa 190
<210> 2
<211> 294
<212> PRT
<213> 人工序列(Artificial Sequence)
<400> 2
Pro Leu Leu Leu Ala Leu Val Ala Pro Pro Thr Ala Val Ser Leu Gly
1 5 10 15
Ala Gly Leu His Leu Ala Gly Gly Ala Val Ala Met Ala His Ala Ala
20 25 30
Ala Leu Ala Ala Pro Ala Leu Ala Met Leu Gly Ala Gly Ala Ser Pro
35 40 45
Gly Pro Gly Pro Thr Pro His Ala Ser Ala Pro Thr Gly Ala Leu Ala
50 55 60
Met Ala Ala Pro Gly Pro Gly Gly Met Pro Thr Ala Ala Leu Gly Ile
65 70 75 80
Ala Gly Thr Gly Gly Gly Ala Ala Ala Thr Ala Val Gly Val Gly Pro
85 90 95
Pro Ala Gly Thr Val Leu Ala Leu Ile Gly Ala Ser Pro Ile Ala Ser
100 105 110
Val Val Ser Ala Pro Ala Leu Ala Ala Ala Pro Leu Ile Ala Ile Ala
115 120 125
Gly Ala Pro Thr Ala Leu Thr Gly Thr Ser Gly Gly Gly Cys Val Gly
130 135 140
Ala Ala Cys Ala Pro Leu Ala Gly Ser Gly Thr Gly Pro Thr Leu Thr
145 150 155 160
Ala Leu Ile Ala Gly Gly Val Ala Gly His Leu Pro Val Leu Val Gly
165 170 175
Ile Leu Ala Thr Leu Leu Ala Gly Thr Pro Pro Ala Ala Ala Val Leu
180 185 190
Val Ala Pro Ile Thr Ala Ala Ala Ala Gly Leu Leu Thr Pro Leu Gly
195 200 205
Ser Gly Val Gly Val Ala Ala Ala Gly Pro Ala Met Gly Met Ala Ala
210 215 220
Ala Gly Ala Ala Ala Gly Met Leu Ala Thr Leu Ser Pro Ala Gly Leu
225 230 235 240
Gly Val Thr Thr Leu Val Ala Ser Gly Leu Ala Ala Leu Gly Val Ala
245 250 255
Ala Ala Leu Gly Leu Ser Gly Leu Thr Val Leu Met His Ala Gly Leu
260 265 270
Val Met Gly Leu Leu Ala Leu Leu Thr Ser Ala Ala Leu Val Ala Ile
275 280 285
Ala Val Gly Ala Gly Ile
290
<210> 3
<211> 882
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 3
ccaaagaaaa agagaaaagt tgctccacct actgatgttt ctttgggaga tgagttgcat 60
ttggatggtg aagatgttgc tatggctcac gctgatgctt tggatgattt tgatttggat 120
atgttgggag atggagattc tccaggtcct ggtttcactc cacatgattc tgctccttac 180
ggtgctttgg atatggctga ttttgagttc gaacaaatgt ttactgatgc tttgggtatt 240
gatgagtatg gtggtggtgc tgatgatact agagttgaag ttcaaccacc tgctcaatgg 300
gttttggatt tgattgaggc ttctcctatt gcttctgttg tttctgatcc aagattggct 360
gataaccctt tgatcgctat taatcaagct tttactgatt tgactggtta ctctgaagag 420
gaatgtgttg gtagaaactg tagattcttg gctggttctg gtactgaacc atggttgact 480
gataaaatta gacaaggtgt tagagagcat aagcctgttt tggttgaaat cttgaactac 540
aaaaaggatg gtactccatt cagaaatgct gttttggttg ctcctatcta cgatgatgat 600
gatgagttgt tgtatttctt gggttctcaa gttgaagttg atgatgatca accaaatatg 660
ggtatggcta gaagagagag agctgctgaa atgttgagaa ctttgtctcc tagacaattg 720
gaagttacta ctttggttgc ttctggtttg agaaacaagg aggttgctgc tagattgggt 780
ttgtctgaaa agactgttaa gatgcacaga ggtttggtta tggagaaatt gaatttgaag 840
acttctgctg atttggttag aattgctgtt gaagctggta tc 882
<210> 4
<211> 1754
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 4
tttttgtaga aatgtcttgg tgtcctcgtc caatcaggta gccatctctg aaatatctgg 60
ctccgttgca actccgaacg acctgctggc aacgtaaaat tctccggggt aaaacttaaa 120
tgtggagtaa tggaaccaga aacgtctctt cccttctctc tccttccacc gcccgttacc 180
gtccctagga aattttactc tgctggagag cttcttctac ggcccccttg cagcaatgct 240
cttcccagca ttacgttgcg ggtaaaacgg aggtcgtgta cccgacctag cagcccaggg 300
atggaaaagt cccggccgtc gctggcaata atagcgggcg gacgcatgtc atgagattat 360
tggaaaccac cagaatcgaa tataaaaggc gaacaccttt cccaattttg gtttctcctg 420
acccaaagac tttaaattta atttatttgt ccctatttca atcaattgaa caactatagg 480
aattcgcctt agacagactg ttcctcagtt caagttgggc acttacgaga agaccggtat 540
gccaaagaaa aagagaaaag ttgctccacc tactgatgtt tctttgggag atgagttgca 600
tttggatggt gaagatgttg ctatggctca cgctgatgct ttggatgatt ttgatttgga 660
tatgttggga gatggagatt ctccaggtcc tggtttcact ccacatgatt ctgctcctta 720
cggtgctttg gatatggctg attttgagtt cgaacaaatg tttactgatg ctttgggtat 780
tgatgagtat ggtggtggtg ctgatgatac tagagttgaa gttcaaccac ctgctcaatg 840
ggttttggat ttgattgagg cttctcctat tgcttctgtt gtttctgatc caagattggc 900
tgataaccct ttgatcgcta ttaatcaagc ttttactgat ttgactggtt actctgaaga 960
ggaatgtgtt ggtagaaact gtagattctt ggctggttct ggtactgaac catggttgac 1020
tgataaaatt agacaaggtg ttagagagca taagcctgtt ttggttgaaa tcttgaacta 1080
caaaaaggat ggtactccat tcagaaatgc tgttttggtt gctcctatct acgatgatga 1140
tgatgagttg ttgtatttct tgggttctca agttgaagtt gatgatgatc aaccaaatat 1200
gggtatggct agaagagaga gagctgctga aatgttgaga actttgtctc ctagacaatt 1260
ggaagttact actttggttg cttctggttt gagaaacaag gaggttgctg ctagattggg 1320
tttgtctgaa aagactgtta agatgcacag aggtttggtt atggagaaat tgaatttgaa 1380
gacttctgct gatttggtta gaattgctgt tgaagctggt atctaagagc ttgggcccgt 1440
tttagcctta gacatgactg ttcctcagtt caagttgggc acttacgaga agcttgctag 1500
attctaatca agaggatgtc agaatgccat ttgcctgaga gatgcaggct tcatttttga 1560
tactttttta tttgtaacct atatagtata ggattttttt tgtcattttg tttcttctcg 1620
tacgagcttg ctcctgatca gcctatctcg cagctgatga atatcttgtg gtaggggttt 1680
gggaaaatca ttcgagtttg atgtttttct tggtatttcc cactcctctt cagagtacag 1740
aagattaagt gaga 1754
<210> 5
<211> 1367
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 5
cgggagatct tcgctagcct cgagtaggta gcctttagtc catgcgttat aggtagcctt 60
tagtccatgc gttataggta gcctttagtc catgcgttat aggtagcctt tagtccatgc 120
gttataggta gcctttagtc catgaagctt agacactaga gggtatataa tggaagctcg 180
acttccagct tggcaatccg gtactgttgg taaattcagt gtggtggact taaggagatg 240
agaaaaggtg aagagttgtt tactggtgtt gttcctattt tggttgaatt ggatggagat 300
gttaatggtc ataagttctc tgtttctggt gaaggagagg gagatgctac ttacggtaaa 360
ttgactttga agtttatttg tactactggt aaattgccag ttccttggcc aactttggtt 420
actacttttg gttacggtgt tcaatgtttc gctagatatc ctgatcatat gaagcaacac 480
gatttcttta agtctgctat gccagaaggt tacgttcaag agagaactat tttctttaag 540
gatgatggta actacaagac tagagctgag gttaagtttg agggagatac tttggttaac 600
agaatcgaat tgaagggtat tgatttcaag gaggatggta acatcttggg tcataagttg 660
gaatacaact acaactctca caacgtttac attatggctg ataagcaaaa gaacggtatt 720
aaggttaact tcaagatcag acataacatc gaggatggtt ctgttcaatt ggctgatcac 780
taccaacaaa acactcctat tggagatggt cctgttttgt tgccagataa tcactatttg 840
tctactcaat ctgctttgtc taaagatcca aacgaaaaga gagatcatat ggttttgttg 900
gagtttgtta ctgctgctgg tattactcac ggtatggatg aattgtataa ataaggtacc 960
gaacaaaaac tcatctcaga agaggatctg aatagcggcg gccgccatca tcatcatcat 1020
cattgagttt tagccttaga catgactgtt caagttgggc acttacgaga agtaaataag 1080
ttataaaaaa aataagtgta tacaaatttt aaagtgactc ttaggtttta aaacgaaaat 1140
tcttgttctt gagtaactct ttcctgtagg tcaggttgct ttctcaggta tagcatgagg 1200
tcgctcttat tgaccacacc tctaccggca tgccgagcaa atgcctgcaa atcgctcccc 1260
atttcaccca attgtagata tgctaactcc agcaatgagt tgatgaatct cggtgtgtat 1320
tttatgtcct cagaggacaa cacctgttgt aatcgttctt ccacacg 1367
<210> 6
<211> 29
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 6
gatttttgta gaaatgtctt ggtgtcctc 29
<210> 7
<211> 24
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 7
cgtgtggaag aacgattaca acag 24
<210> 8
<211> 19
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 8
ggctagcgaa gatctcccg 19
<210> 9
<211> 19
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 9
cgggagatct tcgctagcc 19
- 一种蓝光调节启动子、蓝光调节启动子的融合基因、蓝光介导调节质粒及构建方法和应用
- 一种蓝光介导调节质粒及其构建方法和应用