嘧啶核苷磷酸化酶突变体及2'-氟代核苷的制备方法
文献发布时间:2024-04-18 19:58:21
技术领域
本发明涉及酶催化领域,具体而言,涉及一种嘧啶核苷磷酸化酶突变体及2'-氟代核苷的制备方法。
背景技术
小核酸类药物包括反义核酸(ASO),小干扰RNA(siRNA)和适配体(aptamer)等,其活性成分主要是RNA和DNA,可以根据碱基互补配对原则特异性地作用于靶点。目前,已有超过10多种小核酸类药物获批上市,应用于眼科领域和心血管和代谢等遗传病领域。临床实验表明,小核酸类药物还可以应用于癌症,神经疾病,呼吸系统和传染病等诸多领域,因而具有广阔的应用前景。小核酸类药物往往需要特定的化学修饰以提高其稳定性并延长其在体内的半衰期,如对核糖的2'位置进行氟代修饰。因而,随着小核酸类药物市场规模的不断扩大,市场对此类药物的重要合成前体2'-氟修饰的核苷单体的需求也而持续增长。
目前,市场上2'-氟代核苷单体是以化学法生产,而化学合成过程比较繁琐,会涉及多个步骤,需要对多个官能团进行保护和脱保护,并且需要用到有机试剂对环境造成污染等,导致生产成本较高。此外,2'-氟修饰的核苷单体还可以用生物法进行合成,如可以利用嘧啶核苷磷酸化酶(PyNP)和嘌呤核苷磷酸化酶(PNP)将2'-氟-2'-脱氧尿苷和腺嘌呤一锅法来合成2'-氟-2'-脱氧腺苷。但是,由于嘧啶核苷磷酸化酶(PyNP)催化的2‘-氟-2'-脱氧尿苷生成2'-氟-2'-脱氧核糖-1-磷酸和尿嘧啶的反应由于酶的活性较差,因而限制了利用生物法来合成2'-氟修饰核苷单体的商业化生产。Szeker等人利用热葡糖苷地芽孢杆菌(Geobacillus thermoglucosidasius)来源的野生型嘧啶核苷磷酸化酶(GsPyNP)在0.5 h内仅能催化0.004 mM的2'-氟-2'-脱氧尿苷,而嗜热栖热菌(Thermus thermophilus)来源的野生型嘧啶核苷磷酸化酶(TtPyNP)可水解0.156 mM的2'-氟-2'-脱氧尿苷。在最优条件下,利用野生型TtPyNP反应17 h也仅能催化0.65 mM的2'-氟-2'-脱氧尿苷(Comparativeinvestigations on thermostable pyrimidine nucleosidephosphorylases fromGeobacillus thermoglucosidasius and Thermus thermophilus)。
发明内容
本发明的主要目的在于提供一种嘧啶核苷磷酸化酶突变体及2'-氟代核苷的制备方法,以解决现有技术中的酶催化合成2'-氟代核苷产量低的问题。
为了实现上述目的,根据本发明的第一个方面,提供了一种嘧啶核苷磷酸化酶突变体,包括:(a)以SEQ ID NO:1所示的嘧啶核苷磷酸化酶野生型酶TtPyNP为基础发生突变的蛋白,所述突变选自如下任意一种或多种突变:K80、V4、I7、R8、R17、E19、G27、R30、P34、L47、G49、L50、L58、D61、S91、F105、G112、T119、E135、A151、A157、V173、I179、I183、A189、A190、F206、M207、L215、G224、Q225、A227、V231、E251、A271、L274、R280、L281、L284、G285、R303、D329、V337、G355、K360;或(b)与(a)中限定的氨基酸序列具有70%以上同源性且具有嘧啶核苷磷酸化酶活性的蛋白质。
进一步地,(a)中,各位点取代的氨基酸的类型各自独立地选自如下:
K80Q、K80Y、V4P、V4G、I7G、R8N、R8F、R17E、E19P、G27S、R30K、P34K、P34L、P34G、L47H、G49T、G49V、G49S、L50A、L50D、L50G、L50K、L50N、L50R、L50S、L58T、L58Q、D61T、S91A、S91V、S91C、F105I、F105T、G112S、G112T、G112A、G112L、G112C、T119N、E135K、E135A、A151S、A157D、A157N、A157T、V173S、V173T、I179V、I183V、A189V、A189T、A190T、A190V、A190I、F206M、M207L、M207R、L215I、G224D、Q225N、A227D、V231D、E251V、A271S、L274K、L274I、R280V、L281R、L284S、L284G、G285D、R303K、R303A、D329F、V337T、G355S、K360L、K360I;
其中,数字前字母代表原始氨基酸,数字后字母代表突变氨基酸。
进一步地,突变包括如下任意一种氨基酸突变:
K80Q+G112A、K80Q+G112C、K80Q+G112S、K80Q+G112T、L50A+K80Q+G112T、L50D+K80Q+G112T、L50G+K80Q+G112T、L50K+K80Q+G112T、L50N+K80Q+G112T、L50S+K80Q+G112T、K80Q+G112T+A157T、D61T+K80Q+G112T+A157T、E19P+K80Q+G112T+A157T、G27S+K80Q+G112T+A157T+L281R、G49S+K80Q+G112T+A157T、G49T+K80Q+G112T+A157T、G49V+K80Q+G112T+A157T、I7G+K80Q+G112T+A157T、K80Q+G112T+A157T+A190T+L281R、K80Q+G112T+A157T+A190V+L281R、K80Q+G112T+A157T+A271S+L281R、K80Q+G112T+A157T+L215I+L281R、K80Q+G112T+A157T+L274I+L281R、K80Q+G112T+A157T+L274K、K80Q+G112T+A157T+L281R、K80Q+G112T+A157T+L281R+G355S、K80Q+G112T+A157T+L281R+K360I、K80Q+G112T+A157T+L281R+V337T、K80Q+G112T+A157T+L284G、K80Q+G112T+A157T+L284S、K80Q+G112T+A157T+Q225N+L281R、K80Q+G112T+A157T+R280V、K80Q+G112T+A157T+R303K、K80Q+G112T+A157T+V173S、K80Q+G112T+A157T+V173T+A189V+L281R、K80Q+G112T+A157T+V173T+L281R、K80Q+G112T+A189V、K80Q+G112T+A190I、K80Q+G112T+A190T、K80Q+G112T+E135A+A157T+L281R、K80Q+G112T+E135K、K80Q+G112T+K360I、K80Q+G112T+V173T、L47H+K80Q+G112T+A157T、L50A+K80Q+G112T+A157T+V173T+L281R、L50K+K80Q+G112T+A157T+L281R、L50N+K80Q+G112T+A157T+L281R、L50R+K80Q+G112T+A157T、L58Q+K80Q+G112T+A157T、L58T+K80Q+G112T+A157T、P34G+K80Q+G112T+A157T、P34K+K80Q+G112T+A157T、P34L+K80Q+G112T+A157T+L281R、R17E+K80Q+G112T+A157T、V4G+K80Q+G112T+A157T+L281R、V4P+K80Q+G112T+A157T、V4P+L50R+G112C+I183V+A190T+L215I+L281R+K360L、E19P+P34K+F105I+A157N+V173S+F206M+V231D+L281R、K80Q+G112T+A157T+V173T+A227D+L281R、K80Q+G112T+A157T+V173T+G224D+L281R、K80Q+G112T+A157T+V173T+L281R+D329F、K80Q+G112T+A157T+V173T+L281R+G285D、K80Q+G112T+A157T+V173T+L281R+R303A、L47H+L58Q+S91V+T119N+A157T+M207R+G224D+A271S、R17E+R30K+G112L+E135K+A157D+I183V+L284G+K360I、R30K+K80Q+G112T+A157T+V173T+L281R、R8F+G27S+S91A+G112T+A151S+V173T+M207L+V337T、V4G+R8N+P34L+G49V+L50D+V173T+L215I+G355S、K80Q+G112S+I179V+A227D+L274K+R280V+R303A+K360L、K80Q+G112T+A157T+V173T+A189T+A227D+L281R、K80Q+G112T+A157T+V173T+A189V+A227D+L281R、K80Q+G112T+A157T+V173T+A190V+A227D+L281R、K80Q+G112T+A157T+V173T+F206M+A227D+L281R、K80Q+G112T+A157T+V173T+I179V+A227D+L281R、K80Q+G112T+A157T+V173T+I183V+A227D+L281R、K80Q+G112T+A157T+V173T+M207R+A227D+L281R、K80Q+G112T+T119N+A157T+V173T+A227D+L281R、K80Y+F105T+G112A+A157T+V173T+A189T+Q225N+L274I、L50A+K80Q+V173T+I183V+A189T+Q225N+L274I+G285D、L50G+K80Y+T119N+V173T+L215I+L284S+R303K+D329F、K80Q+G112T+T119N+A157T+V173T+I179V+A227D+E251V、K80Q+G112T+T119N+A157T+V173T+I179V+A227D+L281R、K80Q+G112T+T119N+A157T+V173T+I183V+A227D+L281R、K80Q+G112T+T119N+A157T+V173T+A189T+A227D+L281R、K80Q+G112T+T119N+A157T+V173T+A189V+A227D+L281R、K80Q+G112T+T119N+A157T+V173T+A190T+A227D+L281R、K80Q+G112T+T119N+A157T+V173T+A190V+A227D+L281R 、 K80Q+G112T+T119N+A157T+V173T+F206M+A227D+L281R、K80Q+G112T+T119N+A157T+V173T+M207L+A227D+L281R。
进一步地,嘧啶核苷磷酸化酶突变体包括与(a)中限定的氨基酸序列具有75%以上、80%以上、85%以上,更优选95%以上,进一步优选99%以上同源性且具有嘧啶核苷磷酸化酶突变体的蛋白质。
为了实现上述目的,根据本发明的第二个方面,提供了一种DNA分子,该DNA分子编码上述嘧啶核苷磷酸化酶突变体。
为了实现上述目的,根据本发明的第三个方面,提供了一种重组质粒,该重组质粒连接有上述DNA分子。
为了实现上述目的,根据本发明的第四个方面,提供了一种宿主细胞,该宿主细胞内含有上述DNA分子或重组质粒。
进一步地,宿主细胞包括原核细胞;优选地,原核细胞包括大肠杆菌。
为了实现上述目的,根据本发明的第五个方面,提供了一种2'-氟代核苷的制备方法,该制备方法包括:a)利用上述嘧啶核苷磷酸化酶突变体,催化底物核苷裂解为磷酸化2'-氟代戊糖和游离碱基,底物核苷为2'-氟代戊糖结构;b)利用嘌呤核苷磷酸化酶,催化磷酸化2'-氟代戊糖和底物碱基结合,制备获得2'-氟代核苷。
进一步地,底物核苷包括2'-氟-2'-脱氧尿苷、2'-氟-2'-脱氧胞苷或2'-脱氧-2'-氟胸苷。
进一步地,底物碱基包括含有修饰或未修饰的腺嘌呤、鸟嘌呤、胸腺嘧啶或胞嘧啶,或含有修饰的尿嘧啶。
进一步地,2'-氟代核苷包括含有2'-氟-2'-脱氧戊糖结构的核苷,优选地,2'-氟代核苷包括2'-氟-2'-脱氧腺苷、2'-氟-2'-脱氧-2-氨基腺苷、2'-氟-2'-脱氧鸟苷或2'-氟-2'-脱氧胞苷;或者,2'-氟代核苷选自碱基结构上具有取代基的2'-氟代-2'-脱氧核苷。
为了实现上述目的,根据本发明的第六个方面,提供了一种磷酸化2'-氟代戊糖的制备方法,该制备方法包括:利用上述嘧啶核苷磷酸化酶突变体,催化底物核苷裂解为磷酸化2'-氟代戊糖和游离碱基,底物核苷为2'-氟代戊糖结构。
进一步地,底物核苷包括2'-氟-2'-脱氧尿苷、2'-氟-2'-脱氧胞苷或2'-脱氧-2'-氟胸苷。
应用本发明的技术方案,利用上述嘧啶核苷磷酸化酶突变体,能够催化含有2'-氟代戊糖结构的底物核苷裂解为磷酸化2'-氟代戊糖和游离碱基,进而制备获得目标产物2'-氟代核苷。上述嘧啶核苷磷酸化酶突变体相较于野生型酶,酶活较好,制备2'-氟代核苷产物的转化率和效率较高,能够大幅减少酶的使用量,从而降低生产成本,适用于工业化放大生产。
附图说明
构成本申请的一部分的说明书附图用来提供对本发明的进一步理解,本发明的示意性实施例及其说明用于解释本发明,并不构成对本发明的不当限定。在附图中:
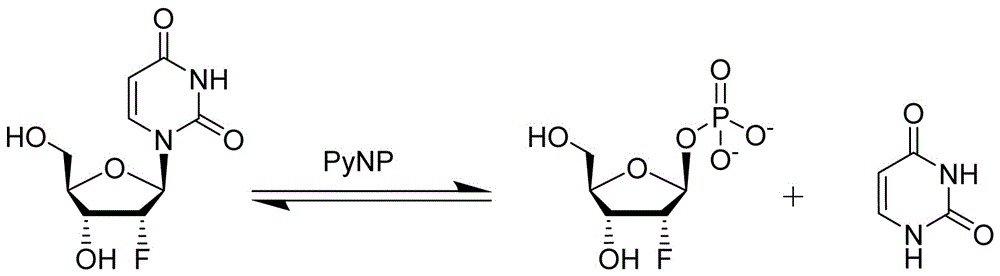
图1示出了根据本发明实施例中利用嘧啶核苷磷酸化酶,催化具有2'-氟代戊糖的底物核苷裂解为磷酸化2'-氟代戊糖和游离碱基的化学反应示意图。
图2示出了根据本发明实施例中利用嘌呤核苷磷酸化酶,催化磷酸化2'-氟代戊糖和底物碱基结合,制备获得2'-氟代核苷的化学反应示意图。
图3示出了根据本发明实施例6的以2'-氟- 2'-脱氧尿苷和腺嘌呤为底物制备2'-氟- 2'-脱氧腺苷的HPLC图谱。
具体实施方式
需要说明的是,在不冲突的情况下,本申请中的实施例及实施例中的特征可以相互组合。下面将结合实施例来详细说明本发明。
术语解释:
2'-氟代核苷:戊糖2位碳原子上羟基被氟原子取代的核苷。
如背景技术所提到的,在现有技术中,利用酶催化制备2'-氟代核苷的产量和产量均较低,难以满足工业生产的要求因而,在本申请中发明人尝试开发一种酶活更高的嘧啶核苷磷酸化酶突变体,能够高效制备目标产物2'-氟代核苷,因而提出了本申请的一系列保护方案。
在本申请第一种典型的实施方式中,提供了一种嘧啶核苷磷酸化酶突变体,包括:(a)以SEQ ID NO:1所示的嘧啶核苷磷酸化酶野生型酶TtPyNP为基础发生突变的蛋白,突变选自如下任意一种或多种突变:K80、V4、I7、R8、R17、E19、G27、R30、P34、L47、G49、L50、L58、D61、S91、F105、G112、T119、E135、A151、A157、V173、I179、I183、A189、A190、F206、M207、L215、G224、Q225、A227、V231、E251、A271、L274、R280、L281、L284、G285、R303、D329、V337、G355、K360;或
(b)与(a)中限定的氨基酸序列具有70%以上同源性且具有嘧啶核苷磷酸化酶活性的蛋白质。
在一种优选的实施例中,上述(a)中,各位点取代的氨基酸的类型各自独立地选自如下:
K80Q、K80Y、V4P、V4G、I7G、R8N、R8F、R17E、E19P、G27S、R30K、P34K、P34L、P34G、L47H、G49T、G49V、G49S、L50A、L50D、L50G、L50K、L50N、L50R、L50S、L58T、L58Q、D61T、S91A、S91V、S91C、F105I、F105T、G112S、G112T、G112A、G112L、G112C、T119N、E135K、E135A、A151S、A157D、A157N、A157T、V173S、V173T、I179V、I183V、A189V、A189T、A190T、A190V、A190I、F206M、M207L、M207R、L215I、G224D、Q225N、A227D、V231D、E251V、A271S、L274K、L274I、R280V、L281R、L284S、L284G、G285D、R303K、R303A、D329F、V337T、G355S、K360L、K360I;其中,数字前字母代表原始氨基酸,数字后字母代表突变氨基酸。
在一种优选的实施例中,突变包括如下任意一种氨基酸突变:
K80Q+G112A、K80Q+G112C、K80Q+G112S、K80Q+G112T、L50A+K80Q+G112T、L50D+K80Q+G112T、L50G+K80Q+G112T、L50K+K80Q+G112T、L50N+K80Q+G112T、L50S+K80Q+G112T、K80Q+G112T+A157T、D61T+K80Q+G112T+A157T、E19P+K80Q+G112T+A157T、G27S+K80Q+G112T+A157T+L281R、G49S+K80Q+G112T+A157T、G49T+K80Q+G112T+A157T、G49V+K80Q+G112T+A157T、I7G+K80Q+G112T+A157T、K80Q+G112T+A157T+A190T+L281R、K80Q+G112T+A157T+A190V+L281R、K80Q+G112T+A157T+A271S+L281R、K80Q+G112T+A157T+L215I+L281R、K80Q+G112T+A157T+L274I+L281R、K80Q+G112T+A157T+L274K、K80Q+G112T+A157T+L281R、K80Q+G112T+A157T+L281R+G355S、K80Q+G112T+A157T+L281R+K360I、K80Q+G112T+A157T+L281R+V337T、K80Q+G112T+A157T+L284G、K80Q+G112T+A157T+L284S、K80Q+G112T+A157T+Q225N+L281R、K80Q+G112T+A157T+R280V、K80Q+G112T+A157T+R303K、K80Q+G112T+A157T+V173S、K80Q+G112T+A157T+V173T+A189V+L281R、K80Q+G112T+A157T+V173T+L281R、K80Q+G112T+A189V、K80Q+G112T+A190I、K80Q+G112T+A190T、K80Q+G112T+E135A+A157T+L281R、K80Q+G112T+E135K、K80Q+G112T+K360I、K80Q+G112T+V173T、L47H+K80Q+G112T+A157T、L50A+K80Q+G112T+A157T+V173T+L281R、L50K+K80Q+G112T+A157T+L281R、L50N+K80Q+G112T+A157T+L281R、L50R+K80Q+G112T+A157T、L58Q+K80Q+G112T+A157T、L58T+K80Q+G112T+A157T、P34G+K80Q+G112T+A157T、P34K+K80Q+G112T+A157T、P34L+K80Q+G112T+A157T+L281R、R17E+K80Q+G112T+A157T、V4G+K80Q+G112T+A157T+L281R、V4P+K80Q+G112T+A157T、V4P+L50R+G112C+I183V+A190T+L215I+L281R+K360L、E19P+P34K+F105I+A157N+V173S+F206M+V231D+L281R、K80Q+G112T+A157T+V173T+A227D+L281R、K80Q+G112T+A157T+V173T+G224D+L281R、K80Q+G112T+A157T+V173T+L281R+D329F、K80Q+G112T+A157T+V173T+L281R+G285D、K80Q+G112T+A157T+V173T+L281R+R303A、L47H+L58Q+S91V+T119N+A157T+M207R+G224D+A271S、R17E+R30K+G112L+E135K+A157D+I183V+L284G+K360I、R30K+K80Q+G112T+A157T+V173T+L281R、R8F+G27S+S91A+G112T+A151S+V173T+M207L+V337T、V4G+R8N+P34L+G49V+L50D+V173T+L215I+G355S、K80Q+G112S+I179V+A227D+L274K+R280V+R303A+K360L、K80Q+G112T+A157T+V173T+A189T+A227D+L281R、K80Q+G112T+A157T+V173T+A189V+A227D+L281R、K80Q+G112T+A157T+V173T+A190V+A227D+L281R、K80Q+G112T+A157T+V173T+F206M+A227D+L281R、K80Q+G112T+A157T+V173T+I179V+A227D+L281R、K80Q+G112T+A157T+V173T+I183V+A227D+L281R、K80Q+G112T+A157T+V173T+M207R+A227D+L281R、K80Q+G112T+T119N+A157T+V173T+A227D+L281R、K80Y+F105T+G112A+A157T+V173T+A189T+Q225N+L274I、L50A+K80Q+V173T+I183V+A189T+Q225N+L274I+G285D、L50G+K80Y+T119N+V173T+L215I+L284S+R303K+D329F、K80Q+G112T+T119N+A157T+V173T+I179V+A227D+E251V、K80Q+G112T+T119N+A157T+V173T+I179V+A227D+L281R、K80Q+G112T+T119N+A157T+V173T+I183V+A227D+L281R、K80Q+G112T+T119N+A157T+V173T+A189T+A227D+L281R、K80Q+G112T+T119N+A157T+V173T+A189V+A227D+L281R、K80Q+G112T+T119N+A157T+V173T+A190T+A227D+L281R、K80Q+G112T+T119N+A157T+V173T+A190V+A227D+L281R 、 K80Q+G112T+T119N+A157T+V173T+F206M+A227D+L281R、K80Q+G112T+T119N+A157T+V173T+M207L+A227D+L281R。
或与上述氨基酸序列具有70%、75%、80%、85%、90%、95%、96%、97%、98%、99%、99.5%或99.9%以上同源性的蛋白质。
在一种优选的实施例中,嘧啶核苷磷酸化酶突变体包括与(a)中限定的氨基酸序列具有75%、80%、85%、90%、95%、96%、97%、98%、99%、99.5%或99.9%以上同源性且具有嘧啶核苷磷酸化酶突变体的蛋白质。
上述氨基酸突变均在本申请实施例中进行试验探究,相较于具有SEQ ID NO:1所示的氨基酸序列的母本,均具有催化含有2'-氟代戊糖结构的底物核苷裂解为磷酸化2'-氟代戊糖和游离碱基的活性。上述突变位点均为在氨基酸活性位点周围进行的突变,此种突变能够改善突变体与底物的结合能力和/ 或催化能力。而对于远离活性位点的突变,对酶的催化能力影响较小,因此能够获得与上述氨基酸序列具有70%、75%、80%、85%、90%、95%、96%、97%、98%、99%、99.5%或99.9%或以上同源性、且催化活性相同的蛋白质。
本说明书中的同源性(Identity)是指氨基酸序列之间的“同源性”,即氨基酸序列中的种类相同的氨基酸残基的比率的总计。氨基酸序列的同源性可以利用BLAST(BasicLocal Alignment Search Tool)、FASTA等比对程序来确定。
70%、75%、80%、85%、90%、95%、99%以上(比如85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、98.5%、99%、99.5%、99.6%、99.7%、99.8%以上,甚至99.9%以上)同源性且具有相同功能的蛋白质,其活性位点、活性口袋、活性机制、蛋白结构等均和(a)序列提供的蛋白质大概率相同,为通过氨基酸突变获得的同源蛋白。
如本文所用,氨基酸残基缩写如下:丙氨酸(Ala;A)、天冬酰胺(Asn;N)、天冬氨酸(Asp;D)、精氨酸(Arg;R)、半胱氨酸(Cys;C)、谷氨酸(Glu;E)、谷氨酰胺(Gln;Q)、甘氨酸(Gly;G)、组氨酸(His;H)、异亮氨酸(Ile;I)、亮氨酸(Leu;L)、赖氨酸(Lys;K)、蛋氨酸(Met;M)、苯丙氨酸(Phe;F)、脯氨酸(Pro;P),丝氨酸(Ser;S)、苏氨酸(Thr;T)、色氨酸(Trp;W)、酪氨酸(Tyr;Y)和缬氨酸(Val;V)。
取代、替换等规则,一般情况下,性质类似的氨基酸之间相互替换后的效果也类似。例如,在上述同源蛋白中,可发生保守的氨基酸替换。“保守的氨基酸替换”包括但不限于:
疏水性氨基酸(Ala、Cys、Gly、Pro、Met、Val、Ile、Leu)被其他疏水性氨基酸取代;
侧链粗大的疏水性氨基酸(Phe、Tyr、Trp)被其他侧链粗大的疏水性氨基酸取代;
侧链带正电的氨基酸(Arg、His、Lys)被其他侧链带正电的氨基酸取代;
侧链有极性不带电的氨基酸(Ser、Thr、Asn、Gln)被其他侧链有极性不带电的氨基酸取代。
本领域技术人员也可以根据现有技术中的“blosum62评分矩阵”等本领域技术人员熟知的氨基酸替换规则对氨基酸进行保守替换。
本申请中所用的“AlphaFold2-Multimer”,是一种公开的能够预测蛋白复合体构象的人工智能模型,对于蛋白质立体结构的预测能够十分接近在真实试验中利用冷冻电子显微镜等设备所观测的水平。能够获得较为真实的蛋白结构,从而指导对于蛋白结构和蛋白活性的探究。
在本申请第二种典型的实施方式中,提供了一种DNA分子,该DNA分子编码上述嘧啶核苷磷酸化酶突变体。在本申请第三种典型的实施方式中,提供了一种重组质粒,该重组质粒连接有上述DNA分子。
上述DNA能够编码上述嘧啶核苷磷酸化酶突变体,并能够连接在重组质粒上形成环状DNA。上述DNA和重组质粒均能在RNA聚合酶、核糖体、tRNA等的作用下,进行转录、翻译,获得上述嘧啶核苷磷酸化酶突变体。
在本申请第四种典型的实施方式中,提供了一种宿主细胞,该宿主细胞内含有上述DNA分子或重组质粒。
在一种优选的实施例中,宿主细胞包括原核细胞;优选地,原核细胞包括大肠杆菌。
利用上述宿主细胞,能够在宿主细胞中进行重组质粒的复制,也能够将重组质粒上携带的DNA分子进行转录、翻译,获得大量嘧啶核苷磷酸化酶突变体。利用现有技术,对宿主细胞进行破碎蛋白纯化、破碎后粗酶催化或其他方式,能够获得嘧啶核苷磷酸化酶突变体,并进行后续对于底物核苷的催化。该宿主细胞为非植物来源的宿主细胞。
在本申请第五种典型的实施方式中,提供了一种2'-氟代核苷的制备方法,该制备方法包括:a)利用上述嘧啶核苷磷酸化酶突变体,催化底物核苷裂解为磷酸化2'-氟代戊糖和游离碱基,底物核苷2'-氟代戊糖结构;b)利用嘌呤核苷磷酸化酶,催化磷酸化2'-氟代戊糖和底物碱基结合,制备获得2'-氟代核苷。
在上述制备方法中,嘌呤核苷磷酸化酶包括但不限于如SEQ ID NO:3所示的、来源于嗜热脂肪地芽孢杆菌Geobacillus stearothermophilus蛋白质或其突变体(同源性80%及以上),也包括其他来源的嘌呤核苷磷酸化酶(PNP)。
利用上述制备方法,能够利用上述嘧啶核苷磷酸化酶突变体(PyNP),首先进行如下催化,反应示意图如图1所示。将底物核苷制备为磷酸化2'-氟代戊糖和游离碱基。磷酸化2'-氟代戊糖中的磷酸基团来源于催化体系中的磷酸盐。具有底物核苷的结构包括但不限于图1中的结构。
再利用现有技术中的嘌呤核苷磷酸化酶(PNP),利用底物碱基取代磷酸化2'-氟代戊糖上的磷酸基团,从而获得结合有底物碱基的2'-氟代核苷。此步骤中所用的PNP,可以灵活选自现有技术中的嘌呤核苷磷酸化酶,均能够催化反应的进行。反应示意图如图2所示,底物碱基和磷酸化2'-氟代戊糖的结构包括但不限于图2中的结构。
且上述a)和b)的两步骤,能够在同一时间、同一容器、同一反应条件下同步进行,因此上述制备方法能够实现一锅法制备2'-氟代核苷。上述制备方法的转化率和效率较高,能够大幅减少酶的使用量,从而降低生产成本,适用于工业化放大生产。
在一种优选的实施例中,底物核苷包括2'-氟-2'-脱氧尿苷、2'-氟-2'-脱氧胞苷或2'-脱氧-2'-氟胸苷。
在一种优选的实施例中,底物碱基包括含有修饰或未修饰的腺嘌呤、鸟嘌呤、胸腺嘧啶或胞嘧啶,或含有修饰的尿嘧啶。
在一种优选的实施例中,2'-氟代核苷包括含有2'-氟-2'-脱氧戊糖结构的核苷,优选地,2'-氟代核苷包括2'-氟-2'-脱氧腺苷、2'-氟-2'-脱氧-2-氨基腺苷、2'-氟-2'-脱氧鸟苷或2'-氟-2'-脱氧胞苷;或者,2'-氟代核苷选自碱基结构上具有取代基的2'-氟代-2'-脱氧核苷。
上述碱基结构上的取代基或上述底物碱基的修饰,包括但不限于在碱基结构上引入甲基(-CH
甲基化(Methylation):核苷酸碱基经甲基基团的添加而发生的修饰。例如,在DNA中,胞嘧啶(Cytosine)碱基可以发生甲基化,形成5-甲基胞嘧啶(5-methylcytosine)。
氨基甲酰化(Aminomethylation):核苷酸碱基经氨基甲酰基团的添加而发生的修饰。这种修饰可以在胞嘧啶和腺嘌呤(Adenine)碱基上发生。
糖基化(Glycosylation):核苷酸碱基经糖基(sugar moiety)的添加而发生的修饰。例如,腺苷(Adenosine)核苷酸上的腺苷可以发生糖基化。
磷酸化(Phosphorylation):核苷酸碱基经磷酸基团的添加而发生的修饰。磷酸化修饰可以发生在胞嘧啶、腺嘌呤和鸟嘌呤(Guanine)碱基上。
硫代(Sulfuration):核苷酸碱基经硫基团的添加而发生的修饰。硫代修饰可以在胞嘧啶、腺嘌呤和鸟嘌呤碱基上发生。
硝化(Nitration):核苷酸碱基经硝酸酯基团的添加而发生的修饰。这种修饰可以发生在胞嘧啶和腺嘌呤碱基上。
氧化(Oxidation):核苷酸碱基经氧基团的添加而发生的修饰。例如,在DNA中,鸟嘌呤碱基经氧化修饰形成8-氧鸟嘌呤(8-oxoG)。
脱氨(Deamination):核苷酸碱基经氨基团丧失而发生的修饰。这种修饰可以发生在胞嘧啶、腺嘌呤和鸟嘌呤碱基上。
在本申请第六种典型的实施方式中,提供了一种磷酸化2'-氟代戊糖的制备方法,该制备方法包括:利用上述嘧啶核苷磷酸化酶突变体,催化底物核苷裂解为磷酸化2'-氟代戊糖和游离碱基,底物核苷为2'-氟代戊糖结构。
在一种优选的实施例中,底物核苷包括2'-氟-2'-脱氧尿苷、2'-氟-2'-脱氧胞苷或2'-脱氧-2'-氟胸苷。
下面将结合具体的实施例来进一步详细解释本申请的有益效果。
实施例1 TtPyNP和GsPNP表达菌株的构建
由NCBI获得嗜热栖热菌Thermus thermophilus来源的嘧啶核苷磷酸化酶(TtPyNP)的蛋白质序列(Accession No. BAD71594),如SEQ ID NO:1所示,密码子优化后获得编码酶的DNA序列,如SEQ ID NO:2所示,并将其克隆到表达载体pET28a(+)上,得到质粒pET28a-TtPyNP。将pET28a-TtPyNP质粒化转至大肠杆菌BL21(DE3)感受态中,获得单克隆菌株。
使用同样的方法得到pET28a-GsPNP(GsPNP,NCBIAccession No. BAA13510)单克隆菌株,GsPNP蛋白序列如SEQ ID NO:3所示。
SEQ ID NO:1:
MNPVAFIREKREGKKHRREDLEAFLLGYLHDEVPDYQVSAWLMAAFLRGLDPEETLWLTETMARSGKVLDLSGLPHPVDQHSSGGVGDKVSLVVGPILAASGCTFAKMSGRTLAHTGGNIDKLESVPGWRGEMTEAEFLERARRVGLVIAAQSPDLTPLDGKLYALRDVTATTESVPLIASSIMSKKLAVGARSIVLDVKVGRGAFMKTLEEARLLAKTMVAIGQGDGRRVRALLTSMEAPLGRAVGNAIEVREAIEALKGEGPGDLLEVALALAEEALRREGLDPALARKALEGGAALEKFRAFLEAQGGDPRAVEDFSLLPLAEEHPLRAEREGVVREVDAYKVGLAVLALGGGRKRKGEPIDHGVGVYLLKKPGDRVERGEALALVYHRRRGLEEALGHLREAYALGEEAHPAPLVLEAI。
SEQ ID NO:2:
atgaacccggtggcgtttattcgcgaaaaacgcgaaggcaagaaacatcgccgcgaggatctggaagcgtttctgctgggctatctgcacgatgaagtgccggattatcaggtgagcgcgtggctgatggcggcgtttttacgcggcttagatccggaagaaaccctgtggctgaccgaaacgatggcccgcagcggtaaagtgctggatctgagcggtctgccgcatccggttgatcagcatagcagcggcggcgtgggcgataaagtgagcttagtggtgggcccgattctggcggcgagcggttgtacctttgcgaaaatgtcgggccgcaccctggcgcataccggtggtaacattgataaactggaaagcgtgccgggctggcgcggcgaaatgaccgaagcagaatttctggaacgcgcccgtcgcgttggtctggttattgcggcgcaaagcccggatttaacgccgttagatggcaaactgtatgcgctgcgcgatgtgaccgcgaccactgaatcagttccgctgattgcgagcagcattatgagcaagaaactggcggtgggcgcccgtagcattgtgttagatgttaaagtgggccgcggcgcgtttatgaaaaccctggaagaagcgcgtctgctggcgaaaacgatggtggcgattggccagggcgatggccgtcgtgttcgtgcgttattaaccagcatggaagcgcctctgggtcgtgcggttggcaatgcgattgaagtgcgtgaagcgattgaagcgctgaaaggcgaaggcccgggcgatttattagaagtggcgctggcgttagcggaagaagcgctgcgtcgtgaaggcttagatccggcgctggcgcgtaaagcgttagaaggtggcgcggcgctggaaaaatttcgcgcgtttctggaagcgcagggcggcgatccgcgtgcagttgaagattttagcctgctgccgctggcggaagaacatccgttacgtgcggaacgcgaaggcgtggttcgcgaagttgatgcgtataaagtgggcctggcggtgctggcgttaggcggtggtcgtaaacgtaaaggtgaaccgattgatcatggcgtgggcgtgtatctgctgaagaaaccgggcgatcgcgtggaacgcggcgaagcgttagcgttagtgtatcatcgccgccgcggcttagaagaagcgctgggtcatttacgcgaagcgtatgcgctgggcgaagaagcgcatccggcgcctctggttttagaagcgatttaa。
SEQ ID NO:3:
MSVHIGAKEHEIADKILLPGDPLRAKYIAETFLEGATCYNQVRGMLGFTGTYKGHRISVQGTGMGVPSISIYITELMQSYNVQTLIRVGTCGAIQKDVKVRDVILAMTSSTDSQMNRMTFGGIDYAPTANFDLLKTAYEIGKEKGLQLKVGSVFTADMFYNENAQFEKLARYGVLAVEMETTALYTLAAKFGRKALSVLTVSDHILTGEETTAEERQTTFNEMIEVALETAIRQ。
实施例2 TtPyNP突变体的构建
通过全质粒PCR进行饱和突变或定点突变。第一轮突变以实施例1中得到的pET28a-TtPyNP为模板,之后的突变以上一轮筛选出的最佳突变体为模板。PCR反应条件为:94℃预变性10 min;25个扩增循环(94℃变性30 s,68℃退火延伸6.5 min),68℃终延伸10min。得到的PCR产物用DpnI酶消化后转入大肠杆菌BL21(DE3)感受态细胞,37℃过夜培养得到单克隆。
实施例3 TtPyNP突变体的表达及酶液制备
突变体的表达:将突变体接种于含5mL Luria-Bertani液体培养基(含50μg/ mL卡那霉素)的试管中,37℃培养16 h后以1%的接种量接种于含500 mL Luria-Bertani液体培养基的2 L摇瓶中,37℃培养至OD
酶液的制备:称取0.1 g上述制备的菌泥,用1 mL pH为7.5的100 mM磷酸钾缓冲液重悬,震荡混匀后用超声破碎仪将菌体混悬液破碎(功率:30%,时间:5min)。
GsPNP的表达和酶液制备同上述操作。
实施例4 HPLC检测方法
HPLC检测方法:色谱柱AtlantiseT3 Column, 4.6 mmx150 mm,流动相为0.1%的TFA/ 甲醇,流速为1 mL/ min, 40℃,UV检测器,检测波长254 nm,检测时长为15 min。利用此种HPLC条件能够检测2'-氟-2'-脱氧尿苷的转化率。
实施例5 TtPyNP突变体的反应验证
以TtPyNP为模版,构建单点突变和多点突变,并进行反应验证。反应条件为:20 mM2'-氟-2'-脱氧尿苷,20 mM PBS(pH7.0),49 μL 酶液,总体积 1 mL,60℃,反应1 h。取样送HPLC检测,2'-氟-2'-脱氧尿苷的转化率参见表1。结果显示,含K80突变位点的活性提高较多,最终突变体K80Q+G112T+T119N+A157T+V173T+M207L+A227D+L281R转化率可达到30%以上。
表1
。/>
续表1
。
注:上述表中,+代表转化率1-10%(不包括端点值10%),++代表转化率10-15%(不包括端点值15%),+++代表转化率15-20%(不包括端点值20%),++++代表转化率20-25%(不包括端点值25%),+++++代表转化率25-30%,++++++代表转化率>30%。
实施例6 TtPyNP突变体用于2'-氟- 2'-脱氧腺苷的合成
在20 mM 磷酸缓冲液(pH 7.5)中配制10 mL反应体系,其中包括两种底物:100 mM2'-氟- 2'-脱氧尿苷(2'-Deoxy-2'-fluorouridine)和250 mM腺嘌呤(Adenine),两种酶液:1.07 mL突变体K80Q+G112T+T119N+A157T+V173T+M207L+A227D+L281R酶液,4.3 mL的GsPNP酶液,60℃反应137 h。反应结束后,取1 mL反应液加入等体积的DMSO溶解,稀释25倍后送HPLC检测,HPLC谱图参见图3。2'-氟- 2'-脱氧尿苷转化率为>80%(转化生成尿嘧啶(Uracil)),腺嘌呤转化率>30%,2'-氟- 2'-脱氧腺苷(2'-Deoxy-2'-fluoroadenosine)产量>20g/ L。
从以上的描述中,可以看出,本发明上述的实施例实现了如下技术效果:利用上述嘧啶核苷磷酸化酶突变体,能够高效催化含有2'-氟代戊糖结构的底物核苷裂解为磷酸化2'-氟代戊糖和游离碱基。进一步地,利用上述嘧啶核苷磷酸化酶突变体和现有技术中的嘌呤核苷磷酸化酶,能够一锅法制备获得2'-氟代核苷。上述嘧啶核苷磷酸化酶突变体相较于野生型酶,酶活较好,制备2'-氟代核苷产物的转化率和效率较高,能够大幅减少酶的使用量,从而降低生产成本,适用于工业化放大生产。
以上所述仅为本发明的优选实施例而已,并不用于限制本发明,对于本领域的技术人员来说,本发明可以有各种更改和变化。凡在本发明的精神和原则之内,所作的任何修改、等同替换、改进等,均应包含在本发明的保护范围之内。
- 1-(2-脱氧-2-氟-4-硫代-β-D-阿拉伯呋喃糖基)胞嘧啶的合成中间体和硫代核苷的合成中间体以及它们的制造方法
- 1-(2-脱氧-2-氟-4-硫代-β-D-阿拉伯呋喃糖基)胞嘧啶的合成中间体和硫代核苷的合成中间体以及它们的制造方法