参与柑橘果皮多甲氧基黄酮生物合成的氧甲基转移酶、编码基因及扩增引物对和应用
文献发布时间:2023-06-19 19:28:50
技术领域
本发明属于分子生物技术领域,具体涉及参与柑橘果皮多甲氧基黄酮生物合成的氧甲基转移酶、编码基因及扩增引物对和应用。
背景技术
类黄酮是最大的一类植物多酚,占8000多种天然酚类化合物的一半以上,常以游离态或与糖苷形式存在,对植物的生长、发育、开花、结果以及抵御异物的入侵起着重要作用。柑橘中类黄酮含量丰富,以黄酮和黄烷酮两类为主,其中在柑橘果皮中特异性存在一类甲基化程度较高的黄酮苷元,其特点是苯色原酮结构上具有4个及以上的甲氧基,因此也被称为多甲氧基黄酮(PMFs)。近年来,因其在抗氧化、抗肿瘤、抗炎、降血糖等方面的显著作用而受到广泛关注,然而其在柑橘中的生物合成途径尚未明晰。
PMFs合成的最后一步由类黄酮甲基转移酶(OMT)催化,OMT可催化S-腺苷-L-甲硫氨酸中的-CH
发明内容
有鉴于此,本发明的目的在于提供参与柑橘果皮多甲氧基黄酮生物合成的氧甲基转移酶、编码基因及扩增引物对和应用,在柑橘营养品质改良及体外生物合成多甲氧基黄酮方面具有广泛的应用前景。
为了实现上述发明目的,本发明提供以下技术方案:
本发明提供了一个参与柑橘果皮多甲氧基黄酮生物合成的氧甲基转移酶CsCCoAOMT1,所述氧甲基转移酶CsCCoAOMT1的氨基酸序列如SEQ ID NO.1所示。
本发明还提供了编码上述氧甲基转移酶CsCCoAOMT1的基因,所述基因的核苷酸序列如SEQ ID NO.2所示。
本发明还提供了一种扩增上述基因的引物对,所述引物包括上游引物和下游引物,所述上游引物的核苷酸序列如SEQ ID NO.3所示,所述下游引物的核苷酸序列如SEQ IDNO.4所示。
本发明还提供了上述氧甲基转移酶CsCCoAOMT1在将邻羟基环境的 6,7,8,3'位羟基同时催化为甲氧基中的应用。
优选的,所述催化的底物包括黄酮类化合物。
优选的,所述催化发生在柑橘体内或柑橘体外。
本发明还提供了利用上述氧甲基转移酶CsCCoAOMT1或上述基因在改良柑橘营养品质中的应用。
优选的,所述应用包括在柑橘果皮中过表达上述基因。
优选的,所述改良柑橘营养品质包括提升果皮中多甲氧基黄酮的含量。
本发明还提供了一种提升果皮中多甲氧基黄酮含量的方法,包括在柑橘的果皮中过表达上述基因。
有益效果:本发明公开了一种参与柑橘多甲氧基黄酮生物合成的氧甲基转移酶CsCCoAOMT1,并具体公开了其氨基酸序列和编码基因序列。本发明以冰糖橙果实为材料,克隆了CsCCoAOMT1基因,并通过原核表达得到 CsCCoAOMT1重组蛋白。该重组蛋白在体外高效催化多种黄酮类化合物,将处于邻羟基环境的6,7,8,3'位羟基催化为甲氧基;通过农杆菌介导的瞬时转化,在冰糖橙果皮中过表达CsCCoAOMT1基因可有效提升果皮中多甲氧基黄酮的含量。本发明在柑橘营养品质改良及体外生物合成多甲氧基黄酮方面具有广泛的应用前景。
附图说明
图1为CsCCoAOMT1在冰糖橙发育阶段的表达模式分析结果;
图2为重组CsCCoAOMT1蛋白的SDS-PAGE电泳结果;
图3为不同底物与CsCCoAOMT1蛋白的体外催化产物;
图4为CsCCoAOMT1蛋白在不同条件下的催化效率;
图5为CsCCoAOMT1基因在冰糖橙果实上瞬时过表达显著提高四种主要PMFs的含量;
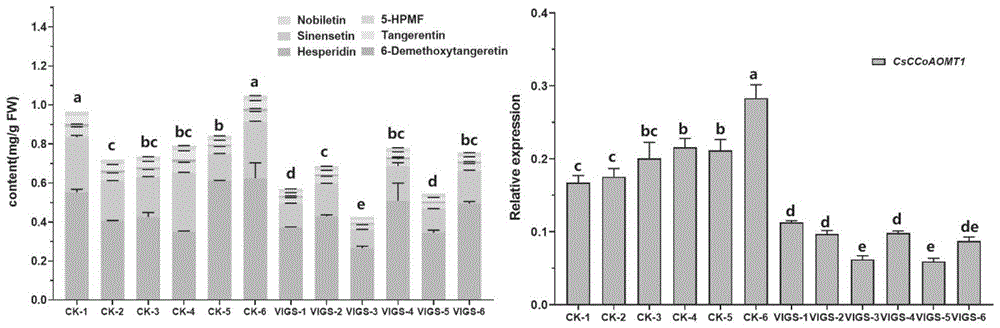
图6为VIGS后CsCCoAOMT1的表达量和总黄酮量的关系。
具体实施方式
本发明提供了一个参与柑橘果皮多甲氧基黄酮生物合成的氧甲基转移酶CsCCoAOMT1,所述氧甲基转移酶CsCCoAOMT1的氨基酸序列如SEQ ID NO.1所示。
本发明所述氧甲基转移酶CsCCoAOMT1参与柑橘果皮多甲氧基黄酮 (PMFs)的生物合成,该酶属于Ⅱ类氧甲基转移酶,且催化过程不依赖金属离子;并且所述氧甲基转移酶CsCCoAOMT1在体外高效催化多种黄酮类化合物,将处于邻羟基环境的6,7,8,3'位羟基催化为甲氧基。
本发明还提供了编码上述氧甲基转移酶CsCCoAOMT1的基因,所述基因的核苷酸序列如SEQ ID NO.2所示。
本发明所述基因的获得,优选以拟南芥中已报道OMT基因为参考,与柑橘基因组数据库(http://www.citrusgenomedb.org/)进行BLAST分析获得 CsCCoAOMT1,核苷酸序列为SEQ ID NO.2。
本发明优选根据SEQ ID NO.2所示的CDS序列和pET32a表达载体,选择EcoRI和XhoI为酶切位点,设计特异引物对SEQ ID NO.3和SEQ ID NO.4,以冰糖橙果实油胞层cDNA为模板,使用Phanta Max Super-Fidelity DNA Polymerase(诺唯赞,中国)进行PCR扩增,经琼脂糖凝胶电泳分析后回收产物,通过同源重组法将其连接到pET32a载体上进行测序分析。本发明所述PCR扩增的反应体系以30μL计,优选包括2×Buffer 15μL,dNTPs 0.6 μL,上下游引物(10mM)各1.2μL,酶0.6μL,模板DNA 1μL,DEPC 水10.4μL。本发明所述PCR扩增的反应程序,优选为:95℃预变性3min; 95℃15sec,58℃15sec,72℃45sec,35个循环;72℃彻底延伸5min。经过测序验证,得到与柑橘基因组数据库相匹配的CsCCoAOMT1序列SEQ IDNO.2,并使用构建了插入CsCCoAOMT1全长的CsCCoAOMT1-pET32a载体.
本发明还提供了一种扩增上述基因的引物对,所述引物包括上游引物和下游引物,所述上游引物的核苷酸序列如SEQ ID NO.3所示,所述下游引物的核苷酸序列如SEQ IDNO.4所示。
本发明所述引物对优选与上述相同,在此不再赘述。
本发明还提供了上述氧甲基转移酶CsCCoAOMT1在将邻羟基环境的 6,7,8,3'位羟基催化为甲氧基中的应用。
本发明所述催化的底物优选包括黄酮类化合物,更优选包括木犀草素、槲皮素、圣草酚、圣草次苷、新圣草次苷、黄芩素、7,8-二羟基黄酮、3',4'- 二羟基黄酮和咖啡酰辅酶A中的一种或多种。在本发明实施例中, CsCCoAOMT1蛋白能体外催化木犀草素、槲皮素、圣草酚、圣草次苷、新圣草次苷、3',4'-二羟基黄酮的3'位羟基甲基化生成对应的3'甲酯化合物(金圣草素、异鼠李素、高圣草酚、高圣草酚-7-O-鼠李糖苷、高圣草酚-7-O-新橙皮糖苷和4'-羟基-3'-甲氧基黄酮);催化黄芩素生成6-甲氧基黄芩(千层纸素A)和7-甲氧基黄芩;催化7,8-二羟基黄酮生成7-甲氧基-8羟基黄酮和 8-甲氧基-7-羟基黄酮;催化咖啡酰辅酶A生成了阿魏酰辅酶A。本发明所述 CsCCoAOMT1对不同底物的催化效率有所不同,以槲皮素为底物时,催化效率最高,其K
本发明所述催化可发生在柑橘体内或柑橘体外,当在体外催化时,针对不同的底物,其最适的催化体系并不相同,如以槲皮素为底物时, CsCCoAOMT1蛋白在Tris-HCl(pH8.0)和45℃时的催化效率最高。在柑橘体内,所述CsCCoAOMT1蛋白在植物体内也表现出对黄酮类化合物高效的甲基化催化活性,促进柑橘PMFs积累,从而达到改良柑橘营养品质的作用。
本发明还提供了利用上述氧甲基转移酶CsCCoAOMT1或上述基因在改良柑橘营养品质中的应用。
本发明所述应用,优选包括在柑橘果皮中过表达上述基因。本发明对所述过表达的方法并没有特殊限定,利用本领域的常规基因过表达方法即可。在本发明实施例中,将所述CsCCoAOMT1基因在冰糖橙果实上瞬时过表达实,结果显示过表达组的CsCCoAOMT1基因相对表达量显著上调,其六种主要PMFs:异甜橙黄酮、甜橙黄酮、川陈皮素、6-去甲橘皮素、七甲氧基黄酮和5-去甲川陈皮素的含量均出现显著上升,证明CsCCoAOMT1的高表达与PMFs积累有显著相关性,CsCCoAOMT1在柑橘果皮PMFs生物合成过程中起到重要的促进作用。
本发明还提供了一种提升果皮中多甲氧基黄酮含量的方法,包括在柑橘的果皮中过表达上述基因。本发明所述方法优选与上述相同,在此不再赘述。
下面结合实施例对本发明提供的参与柑橘果皮多甲氧基黄酮生物合成的氧甲基转移酶、编码基因及扩增引物对和应用进行详细的说明,但是不能把它们理解为对本发明保护范围的限定。
实施例1 CsCCoAOMT1基因的克隆
以拟南芥中已报道OMT基因为参考,与柑橘基因组数据库(http://www.citrusgenomedb.org/)进行BLAST分析获得CsCCoAOMT1,核苷酸序列为SEQ ID NO.2。根据CDS序列和pET32a表达载体,选择EcoRI 和XhoI为酶切位点,设计特异引物对SEQ IDNO.3和SEQ ID NO.4,以冰糖橙果实油胞层cDNA为模板,使用Phanta Max Super-FidelityDNA Polymerase(诺唯赞,中国)进行PCR扩增,经琼脂糖凝胶电泳分析后回收产物,通过同源重组法将其连接到pET32a载体上进行测序分析。
PCR反应体系为30μL,包括2×Buffer 15μL,dNTPs 0.6μL,上下游引物(10mM)各1.2μL,酶0.6μL,模板DNA 1μL,DEPC水10.4μL。 PCR反应程序为95℃预变性3min;95℃15sec,58℃15sec,72℃45sec, 35个循环;72℃彻底延伸5min。
经过测序验证,得到与柑橘基因组数据库相匹配的CsCCoAOMT1序列 SEQ IDNO.2,并使用构建了插入CsCCoAOMT1全长的 CsCCoAOMT1-pET32a载体。
实施例2 CsCCoAOMT1在冰糖橙发育阶段的表达模式分析
1、柑橘果实发育阶段:共收集10个发育时期的冰糖橙果实,分别于盛花后10(S0)、30(S1)、60(S2)、80(S3)、100(S4)、120(S5)、150 (S6)、180(S7)、210(S8)、240(S9)天采摘。每个发育时期设置3个生物学重复,每个重复4个果实;将冰糖橙果皮分为油胞层和白皮层两个部分,用液氮速冻后存放于-80℃待用。
2、HPLC分析:将油胞层和白皮层样品磨成粉末,称取0.1g粉末加入 1ml80%乙醇,超声30min后4000rpm离心10min,收集上清液。重复一次,合并上清液(2ml),用于HPLC检测。HPLC检测体系:以色谱乙腈(A) 和超纯水(B)为流动相,进样体积为10μL,流速1ml/min,0-5min,20% A;5-10min,20%-27%A;10-15min,27%A;15-25min,27%-40%A,25-35min,40%-60%A;35-40min,60%-80%A;40-42min,80%-100%A;42-45min, 100%-20%A;45-50min,20%A。
3、RNA提取:使用CTAB法提取油胞层和白皮层样品的总RNA。称取0.3g样品粉末,加入到4ml65℃预热的CTAB/β-巯基乙醇溶液中,充分涡旋后在65℃裂解5min;加入4ml三氯甲烷:异戊醇(24:1/V:V),充分涡旋后15℃,10000rpm离心10min;吸取上清液到新离心管中,再次加入4ml 三氯甲烷:异戊醇(24:1/V:V)并充分涡旋,离心后吸取上清液到新的离心管中;向上清液中加入1/4体积的10M氯化锂溶液,4℃过夜静置。第二天, 4℃,10000rpm离心30min,小心弃去上清保留沉淀,向沉淀中加入400μL 65℃预热的SSTE,轻轻拍打溶解沉淀后加入400μL三氯甲烷:异戊醇 (24:1/V:V),充分涡旋,转入到1.5ml离心管中,于15℃,10000rpm离心 10min;小心吸取上清液到新的1.5ml离心管,加入2倍体积-20℃预冷的无水乙醇,颠倒混匀后于-80℃静置30min;4℃,10000rpm离心20min,弃上清后将沉淀在通风橱中吹干剩余液体,加入20μL DEPC水溶解沉淀得到总 RNA样品。
4、cDNA合成:以CTAB法提取的总RNA为模板,使用逆转录试剂盒 PrimeScriptTMRT reagent Kit with gDNA Eraser(Takara,日本),参照说明书将1μg总RNA进行逆转录合成cDNA。
5、基因表达分析:以柑橘β-actin为内参基因(SEQ ID NO.5),引物为SEQ ID NO.6和SEQ ID NO.7,CsCCoAOMT1 qPCR引物为SEQ ID NO.8 和SEQ ID NO.9。
qPCR反应体系为25μL,包括12.5μL TB Green(Takara,日本),上下游引物(10μM)各0.5μL,稀释后的cDNA 2μL,DEPC水9.5μL;反应程序为95℃3min;95℃10s,60℃30s,循环45次;95℃10s,溶解曲线从65℃到95℃,每5s上升0.5℃。QPCR仪器为CFX96实时荧光定量PCR 仪(Bio-Rad,美国),每次检测都包括不含cDNA的阴性对照。
黄酮组分分析结果表明,冰糖橙果皮的PMFs特异性积累在油胞层中,且随着发育时期先上升后下降,在S2时期达到最高。CsCCoAOMT1基因表达趋势在S1时期表达量最高,随后下降,到S4时期再次开始上升;这种趋势跟PMFs含量趋势一致,但基因表达提前一个时期出现高峰,推测可能是先发生CsCCoAOMT1基因表达的上调,随后影响了果皮PMFs的积累(图1)。
实施例3 CsCCoAOMT1重组蛋白表达及纯化
1、目的基因重组细胞构建:将测序正确的CsCCoAOMT1-pET32a载体通过热激法转入大肠杆菌表达菌株BL21(DE3)pLysS(唯地,中国),在含氨苄青霉素(Amp)的培养基上培养14小时,筛选阳性单菌落,菌液保存于50%甘油中,放于-80℃待用。
2、原核表达与蛋白纯化:将上述菌液在Amp抗性培养基上划线培养,挑取单菌落于20ml含Amp的LB液体培养基中过夜培养(37℃,200rpm);取10ml菌液加到500ml含Amp的液体LB培养基中,37℃,200rpm培养至 OD
SDS-PAGE电泳结果显示,CsCCoAOMT1-pET32a重组蛋白分子量约45kDa(图2),与理论计算值46.7kDa一致,且条带单一,浓度较高,可用于下一步的酶活分析。
实施例4 CsCCoAOMT1体外酶活功能鉴定
1、底物区域选择性测定:选取木犀草素、槲皮素、圣草酚、圣草次苷、新圣草次苷、黄芩素、7,8-二羟基黄酮、3',4'-二羟基黄酮、咖啡酰辅酶A九种底物验证CsCCoAOMT1重组蛋白的O-甲基化催化功能;反应体系200μ L,包括S-腺苷甲硫氨酸(SAM)1mM、底物200μM、重组蛋白25μL和 Tris-HCl缓冲液;反应液于37℃孵育2h后加入2倍体积甲醇终止反应,室温12000rpm离心15min,吸取上清进行HPLC及质谱分析。
2、酶反应最适pH和最适温度测定:选取槲皮素为底物,测定不同pH 和温度对CsCCoAOMT1重组蛋白反应效率的影响。最适pH测定:选择磷酸钾缓冲液和Tris-HCl缓冲液作为测试,其中磷酸缓冲液的pH梯度为5.5、 6.0、6.5、7.0、7.5,Tris-HCl缓冲液的pH梯度为7.0、7.5、8.0、8.5、9.0;反应体系同上,37℃孵育30min后终止反应。最适温度测定:设置25℃、 30℃、35℃、40℃、45℃、50℃、55℃、60℃、65℃、70℃10个温度梯度,在Tris-HCl(pH8.0)缓冲液中反应30min后终止反应。样品室温12000rpm 离心15min,吸取上清进行HPLC分析,HPLC分析条件与实施例2条件一致。
3、酶反应动力学参数测定:使用MTase-GloTM Methyltransferase Assay 试剂盒(Promaga,美国)测定CsCCoAOMT1重组蛋白对不同底物的亲和力和催化效率;反应体系20μL,包括2mM SAM,10μL稀释的 CsCCoAOMT1重组蛋白,反应缓冲液和不同浓度的底物(0~200μM);37℃孵育30min后,用0.5%三氟乙酸(TFA)终止反应,参照试剂盒说明书加入检测试剂,通过酶标仪测定RLU变化值来检测酶对底物活力。使用GraphPad Prism软件作图,用米氏方程(Michaelics-Menten)拟合计算k
六种底物反应后均能产生新的物质峰,根据标准品图谱和质谱分析确定了反应产物。分析结果表明,CsCCoAOMT1蛋白能体外催化木犀草素、槲皮素、圣草酚、圣草次苷、新圣草次苷、3',4'-二羟基黄酮的3'位羟基甲基化生成对应的3'甲酯化合物(金圣草素、异鼠李素、高圣草酚、高圣草酚-7-O- 鼠李糖苷、高圣草酚-7-O-新橙皮糖苷、4'-羟基-3'-甲氧基黄酮);催化黄芩素生成6-甲氧基黄芩和7-甲氧基黄芩;催化7,8-二羟基黄酮生成7-甲氧基-8羟基黄酮和8-甲氧基-7-羟基黄酮。(图3)。
4、最适pH和最适温度测定实验表明,以槲皮素为底物时, CsCCoAOMT1蛋白在Tris-HCl(pH8.0)和45℃时的催化效率最高(图4)。
酶催化反应动力学试验测定了CsCCoAOMT1重组蛋白对七种黄酮类化合物催化常数,其中对槲皮素的催化效率最高,其次是木犀草素、7,8-二羟基黄酮、3',4'-二羟基黄酮和黄芩等黄酮类化合物,对咖啡酰辅酶A的催化效率低于黄酮类化合物,对圣草酚的催化效率最低(表1);但总体而言, CsCCoAOMT1重组蛋白对这些化合物都表现出较强的的催化活性,最低的圣草酚k
表1酶催化反应动力学评价结果
实施例5冰糖橙果实瞬时过表达CsCCoAOMT1
根据pBI121载体和CsCCoAOMT1目的基因的序列设计特异性引物对 SEQ ID NO.10和SEQ ID NO.11,将CsCCoAOMT1的完整CDS序列(SEQ ID NO.2)构建在pBI121载体上。以空pBI121载体作为对照,将 CrOMT1-pBI121重组载体用电激法转入到农杆菌EHA105中。选择颜色和大小一致、无机械损伤、无病虫害的冰糖橙果实,每个果实为一个重复,选取果实赤道面的对立两侧用于注射,其中一侧注射含有 CsCCoAOMT1-pBI121的农杆菌悬浮液,另一侧注射pBI121空载作为对照。注射后,果实在黑暗中放置12小时,5天后对注射区域的果皮进行取样分析。
CsCCoAOMT1基因在冰糖橙果实上瞬时过表达实验结果表明,与对照组(pBI121)相比,过表达组的CsCCoAOMT1基因相对表达量显著上调,其六种主要PMFs:异甜橙黄酮、甜橙黄酮、川陈皮素、6-去甲橘皮素、七甲氧基黄酮和5-去甲川陈皮素的含量均出现显著上升(图5)。瞬时过表达结果证明CsCCoAOMT1的高表达与PMFs积累有显著相关性,CsCCoAOMT1在柑橘果皮PMFs生物合成过程中起到重要的促进作用。
实施例6 TRV介导的病毒诱导基因沉默(VIGS)
以锦橙材料进行柑橘VIGS试验,选取CsCCoAOMT1基因CDS区300bp 片段构建TRV2载体,引物对序列为SEQ ID NO.12和SEQ ID NO.13;将构建好的CsCCoAOMT1-TRV2质粒电激法转化至农杆菌EHA105菌株中,检测得到阳性克隆。将携带TRV1和TRV2重组体的农杆菌细胞在LB液体培养基中生长到OD
VIGS实验表明,与对照组植株(TRV2)相比,6个VIGS阳性 (CsCCoAOMT1-TRV2)植株中CsCCoAOMT1的相对表达水平均出现不同程度的下降,最低组(VIGS-5)仅为CK组均值的28.2%。阳性植株的总黄酮含量也表现出显著的下降,最低组(VIGS-3)为0.427mg/g FW,为CK 组均值(0.850mg/g FW)的50.2%;其他阳性株系的总黄酮含量也均表现出不同程度的下降(图6)。VIGS结果进一步证明了CsCCoAOMT1的表达水平与柑橘总黄酮积累相关。
以上所述仅是本发明的优选实施方式,应当指出,对于本技术领域的普通技术人员来说,在不脱离本发明原理的前提下,还可以做出若干改进和润饰,这些改进和润饰也应视为本发明的保护范围。