一种快速检测基因SNP的方法及试剂盒
文献发布时间:2024-04-18 20:00:50
技术领域
本发明涉及一种快速检测基因SNP的方法及试剂盒,属于分子生物学技术领域。
背景技术
如何鉴定植物基因型,现有技术中的主流方法是采用DNA分子标记检测。其中,DNA限制性片段长度多态性(RFLP)首次用于构建人类连锁图谱,开创了以DNA多态性作为分子标记的先河,也被称为第一代分子标记;随着PCR技术的应用,RFLP标记逐渐被以PCR为基础的分子标记所取代,开发出第二代分子标记微卫星标记或简单重复多样性(SSR);近年来广泛采用的分子标记是单核苷酸多态性(SNP),SNP是2个DNA序列之间的单个核苷酸差异,由核苷酸的替代、插入或缺失造成。核苷酸作为遗传的最小单元,SNP标记提供了分子标记的最基本形式。SNP也可以包括由于多个核苷酸的插入和缺失造成的In Del。目前,SNP已经取代其他分子标记,成为主流的分子标记,广泛用于水稻育种研究。
近年来,SNP检测技术领域的发展十分迅速,出现大量的SNP检测方法;
(1)测序法。Sanger测序是DNA序列分析的经典方法,可直接获取核酸序列信息,是SNP检测的“金标准”。Sanger测序可发现未知的SNP位点,确定SNP的突变类型和突变位置,是一种无法替代的最直接、最准确的SNP检测方法。通量低、耗时长、配套仪器及试剂昂贵也是其不可避免的缺点。
(2)CASP法。酶切扩增多态性序列(Cleaved Amplified Polymorphic Sequence,CASP),又称限制性片段长度多态性聚合酶链反应(PCR-RFLP),是PCR技术与RFLP技术相结合的方法。该方法基于DNA片段在酶切位点的碱基变异,采用相应的限制性内切酶,对该DNA片段的PCR扩增产物进行酶切,从而产生不同的电泳图谱,由此确定SNP位点的剪辑类型。CAPS仅能检测位于酶切位点的SNP,对于酶切位点之外的SNP则无法检测,应用具有局限性。
(3)TaqMan探针法与分子信标法。TaqMan探针是一种双标记、自淬灭的水解探针,其5’和3’末端分别标记荧光基团和淬灭基团,在探针结构完整时,二者距离较近,荧光基团的信号可被淬灭。若TaqMan探针与靶标序列完全匹配,探针结合DNA模板,Taq酶延伸至探针位置时,其外切酶活性将水解探针切割,释放荧光基团,增强荧光信号。分子信标法与TaqMan探针法原理类似,均是通过探针中单个碱基的识别能力区分SNP。分子信标法采用茎环结构,TaqMan探针使用MGB修饰,以提高探针对SNP的识别特异性。
(4)高分辨率溶解曲线法。高分辨溶解曲线分析技术(high-resolution meltinganalysis,HRM)是通过特定染料与扩增后产物结合后形成特定的熔解峰进行分析检测的方法,使用更强DNA结合能力的饱和荧光染料,具有更高分辨率的溶解曲线,不影响PCR扩增,在DNA解链过程中也不会发生重排。但是饱和染料价格昂贵,增加实验成本。
(5)KASP法。KASP(Kompetitive Allele-Specific PCR)是基于引物末端碱基的特异性匹配对SNP进行检测的方法,该方法针对SNP的等位基因设计两条对应的上游引物,引物3’末端分别位于各自的SNP位点上,5’末端各自带有一段独特的标签序列,并设计一条共用的下游引物;除此之外,针对两条上游引物的标签序列,设计出荧光探针,5’末端标记有荧光基团,同时对应于荧光探针,设计出互补配对的淬灭探针,在其3’端标记淬灭基团。反应未开始时,荧光探针与淬灭探针相结合,无法发出荧光信号。当上游引物中的一条或两条与模板完全匹配,启动扩增,扩增产物可结合下游引物,即形成带标签的DNA片段;该DNA片段可与荧光探针结合,最后形成带有荧光标记的扩增产物。可对SNP位点进行检测。
(6)引物延伸法。引物延伸法是基于PCR反应的一种检测方法,针对SNP位点的两个等位基因分别设计两条不同的特异性引物,3’末端碱基恰好位于SNP处,在其下游设计一条共用引物,与两条特异性引物分别进行PCR反应,当SNP位点与特异性引物的3’末端互补配对时,引物顺利延伸,得到扩增产物。根据扩增结果,可以判断出两个等位基因的SNP位点的突变情况。该方法使用常规PCR,简单易操作,应用广泛,无需昂贵的仪器及设备,成本低廉。
除此之外,还有一些先进的SNP检测技术,例如基因芯片法,将大量针对SNP的寡核苷酸探针高密度地固定在芯片上,从而形成多重寡核苷酸微阵列,而经荧光染料标记后的待测DNA片段,与芯片上固定好的探针阵列进行杂交反应,固定目标序列,清洗去除非目标片段,最后扫描检测芯片上的荧光信号,由此确定SNP位点。检测通量高,可实现自动化。质谱法是基于基质辅助激光解吸电离飞行时间质谱技术的检测方法,其检测过程中,需通过激光激发DNA分子片段,再利用飞行时间判断DNA片段的分子量大小,从而确定SNP位点。
综上所述,早期的SNP检测方法,例如测序法,虽然准确,但是耗时长,通量低;近些年发展的新的SNP检测技术,如芯片法,质谱法,变性高效液相色谱法等。与传统方法相比,检测通量大,准确度高,检测速度快,但是依托于先进的仪器与技术,检测成本相对较高,无法广泛应用。
上述方法中,最具应用价值、应用广泛且低成本的方法是引物延伸法。基于3条引物,两个常规的PCR反应即可检测和分型SNP;但是,在具体实践应用中,也往往产生大量的假阳性,大大限制了引物延伸法的准确度。本领域的研究者们也对此方法进行一系列的改造,例如:人为掺入错配碱基、增加外围引物进行巢式PCR,以此期望加大区分度、提高准确度等。但是值得注意的是,假阳性的产生是由这种方法的原理决定的,因此这些诸多的改进,无论怎样改进,都不能100%避免假阳性的产生。故而,一种简便易行、可操作性强,而又具有相当准确度、高效经济的通过PCR反应检测SNP的方法对于核酸分子诊断、遗传学分析等领域是迫切需要的。
胶体金免疫层析技术是将特异性的抗原或抗体以条带状固定在NC膜上形成检测线(T线)和内参线(R线),胶体金标记试剂(抗体或单克隆抗体)吸附在结合垫上。待测样本经提取后,通过采用5’末端带有特定标记的引物进行PCR扩增,并将扩增产物直接添加到试纸条一端的样本垫上,扩增产物通过毛细作用向前移动,在结合垫上与胶体金标记试剂相互作用,当移动至固定的抗原或抗体的区域时,待检物与金标试剂的结合物有与之发生特异性结合而被截留,聚集在检测带上,形成肉眼可见的观察的显色结果。该检测方法广泛用于抗原抗体及核酸检测领域,但在植物基因SNP检测领域还鲜有使用。
本领域技术人员希望能够开发针对植物基因的SNP检测方法,该方法简便易行、可操作性强,对设备仪器要求低,而又具有相当准确度。
发明内容
为解决上述技术问题,本发明人设计了一种基于引物3’末端双脱氧修饰及5’末端标记并结合胶体金免疫层析的新型检测方法,保留了引物延伸法快速、高效、灵敏度高、可操作性强的特点,又从根本上解决了假阳性的产生,显著缩短结果可视化时间,为植物基因的SNP检测提供了一种便捷、灵敏、低廉而又准确的方法。
本发明第一方面提供了一种快速检测基因SNP的方法,其特征在于,所述方法包括;
(1)获得待测基因组DNA样品;
(2)使用引物对和高保真DNA聚合酶与非高保真DNA聚合酶的混合酶体系对所述DNA样品中的靶基因进行PCR扩增,其中所述高保真聚合酶与非保真聚合酶的比例为(1-3):1;
其中所述引物对包含正向引物和反向引物,其中正向引物和反向引物之一被指定为辨别性引物,所述辨别性引物自3’末端核苷酸起的第1-6个核苷酸中的任一个或多个核苷酸对应于待测SNP位点,并且与突变型靶基因的序列互补,所述辨别性引物的其余核苷酸与野生型靶基因的序列互补配对,并且所述辨别性引物的3’末端核苷酸是经双脱氧修饰的,其中正向引物和反向引物中的另一引物被指定为非辨别性引物,其位于待测SNP位点上游或下游并且其序列与野生型靶基因的序列互补配对;
(3)分析PCR扩增产物,若获得目的扩增产物,则表明待测靶基因中不存在突变;若未获得目的扩增产物,则表明待测靶基因中存在突变。
在本具体的实施方案中,分析PCR扩增产物可以采用琼脂糖凝胶电泳(或聚丙烯酰胺凝胶电泳)分离扩增产物,并根据电泳分离后的结果,判断待测DNA样品的SNP基因型:若分离后无条带,则待测DNA样品(等位基因)的基因型为突变型,即待测靶基因中存在突变;若分离后有条带,则待测DNA样品(等位基因)的基因型为野生型,即待测靶基因中不存在突变。
在优选的实施方案中,分析PCR扩增产物可以采用胶体金试纸检测分析扩增产物。这可以包括对所述辨别性引物的5’末端进行DIG或TAMRA标记,并对所述非辨别性引物的5’末端进行生物素标记,然后将PCR扩增产物用于胶体金试纸检测。进一步地,步骤(2)中所述引物对还包含用于扩增内参基因的引物对。进一步地,所述用于扩增内参基因的引物对中的第一引物的5’末端进行了TAMRA或DIG标记,并且所述内参基因第一引物的5’末端的标记与所述辨别性引物的5’末端的标记不同,以便区分,并且所述用于扩增内参基因的引物对中的第二引物的5’末端进行了生物素标记。
如本文所用,内参基因不受限制并且可以由本领域技术人员常规地选择。例如内参基因的例子可以包括但不限于GAPDH或β-肌动蛋白基因。所述用于扩增内参基因的引物对
因此,在具体的实施方案中,本发明提供了一种快速检测基因SNP的方法,其特征在于,所述方法包括;
(1)获得待测基因组DNA样品;
(2)使用引物对和高保真DNA聚合酶与非高保真DNA聚合酶的混合酶体系对所述DNA样品中的靶基因进行PCR扩增,其中所述高保真聚合酶与非保真聚合酶的比例为(1-3):1;
其中所述引物对包含正向引物和反向引物,其中正向引物和反向引物之一被指定为辨别性引物,所述辨别性引物自3’末端核苷酸起的第1-6个核苷酸中的任一个或多个核苷酸对应于待测SNP位点,并且与突变型靶基因的序列互补,所述辨别性引物的其余核苷酸与野生型靶基因的序列互补配对,并且所述辨别性引物的3’末端核苷酸是经双脱氧修饰的,其中正向引物和反向引物中的另一引物被指定为非辨别性引物,其位于待测SNP位点上游或下游并且其序列与野生型靶基因的序列互补配对;
以及对所述辨别性引物的5’末端进行DIG或TAMRA标记,并对所述非辨别性引物的5’末端进行生物素标记,
并且所述引物对还包含用于扩增内参基因的引物对,所述用于扩增内参基因的引物对中的第一引物的5’末端进行了TAMRA或DIG标记,条件是其标记与所述辨别性引物的5’末端的标记不同,并且所述用于扩增内参基因的引物对中的第二引物的5’末端进行了生物素标记;和
(3)将步骤(2)所得扩增产物用于胶体金试纸检测。
在具体的实施方案中,所述胶体金试纸检测包括:用样品稀释液对扩增产物进行稀释,吸取稀释后的样品缓慢滴加在试纸条加样孔上,根据试纸检测结果判断突变的存在或不存在:当待测基因为野生型时,内参带与检测带都出现红色条带;当待测基因为突变型时,仅内参带出现红色条带。
在具体的实施方案中,正向引物和反向引物中的另一引物(即非辨别性引物)位于待测SNP位点上游或下游可以由本领域技术人员判定,例如,当指定正向引物和反向引物中的正向引物为辨别性引物时,则反向引物位于待测SNP位点下游;当指定正向引物和反向引物中的反向引物为辨别性引物时,则正向引物位于待测SNP位点上游。
在具体的实施方案中,所述SNP包括由点突变、插入突变或缺失突变引起的SNP。
在具体的实施方案中,3’末端核苷酸双脱氧修饰包括:ddC、ddG、ddA、ddT。本发明以ddC为例进行研究说明。
在具体的实施方案中,所述正向引物和反向引物的长度为20-40个核苷酸。
在具体的实施方案中,所述高保真DNA聚合酶是具有3’-5’外切酶活性的DNA聚合酶,所述非高保真DNA聚合酶是不具有3’-5’外切酶活性的DNA聚合酶。
在具体的实施方案中,所述高保真聚合酶与非保真聚合酶的比例为(1-3):1,更佳地为(1.5-2.5):1,最优选为2:1。
在具体的实施方案中,所述胶体金试纸检测是胶体金免疫层析试纸条检测。
在具体的实施方案中,所述待测基因是植物基因。
在具体的实施方案中,所述待测基因包含如SEQ ID NO:5所示的序列,并且所述SNP位于SEQ ID NO:5中第28位的核苷酸处。
在具体的实施方案中,所述辨别性引物的序列如SEQ ID NO:3所示,以及所述非辨别性引物的序列如SEQ ID NO:2所示。
在具体的实施方案中,使用所述辨别性引物和非辨别性引物进行PCR扩增的退火温度为65.2℃。
本发明另一方面提供了一种快速检测待测植物基因SNP的方法的试剂盒,所述试剂盒包括;
正向引物和反向引物,其中正向引物和反向引物之一被指定为辨别性引物,所述辨别性引物自3’末端核苷酸起的第1-6个核苷酸中的任一个或多个核苷酸对应于待测SNP位点,并且与突变型靶基因的序列互补,所述辨别性引物的其余核苷酸与野生型靶基因的序列互补配对,并且所述辨别性引物的3’末端核苷酸是经双脱氧修饰的,其中正向引物和反向引物中的另一引物被指定为非辨别性引物,其位于待测SNP位点上游或下游并且其序列与野生型靶基因的序列互补配对;和
高保真DNA聚合酶与非高保真DNA聚合酶的混合酶体系,其中所述高保真聚合酶与非保真聚合酶的比例为(1-3):1。
在具体的实施方案中,所述辨别性引物的5’末端还进行了DIG或TAMRA标记,并且所述非辨别性引物的5’末端还进行了生物素标记,并且所述试剂盒还包含用于胶体金试纸检测的层析试纸条以及用于扩增内参基因的引物对,;
在具体的实施方案中,所述层析试纸条包被有抗生素抗体的胶体金颗粒以及抗TAMRA抗体或抗DIG抗体。
在具体的实施方案中,所述待测基因是植物基因。
在具体的实施方案中,所述待测基因包含如SEQ ID NO:5所示的序列,并且所述SNP位于SEQ ID NO:5中第28位的核苷酸处,所述辨别性引物的序列如SEQ ID NO:3所示,以及所述非辨别性引物的序列如SEQ ID NO:2所示,并且使用所述辨别性引物和非辨别性引物进行PCR扩增的退火温度为65.2℃。
可以理解的是,在本发明范围内,本发明的上述各项技术特征和在下文(如实施例中)具体描述的各项技术特征之间可以相互组合,从而构成新的或优选的技术方案。限于篇幅,在此不一一赘述。
附图说明
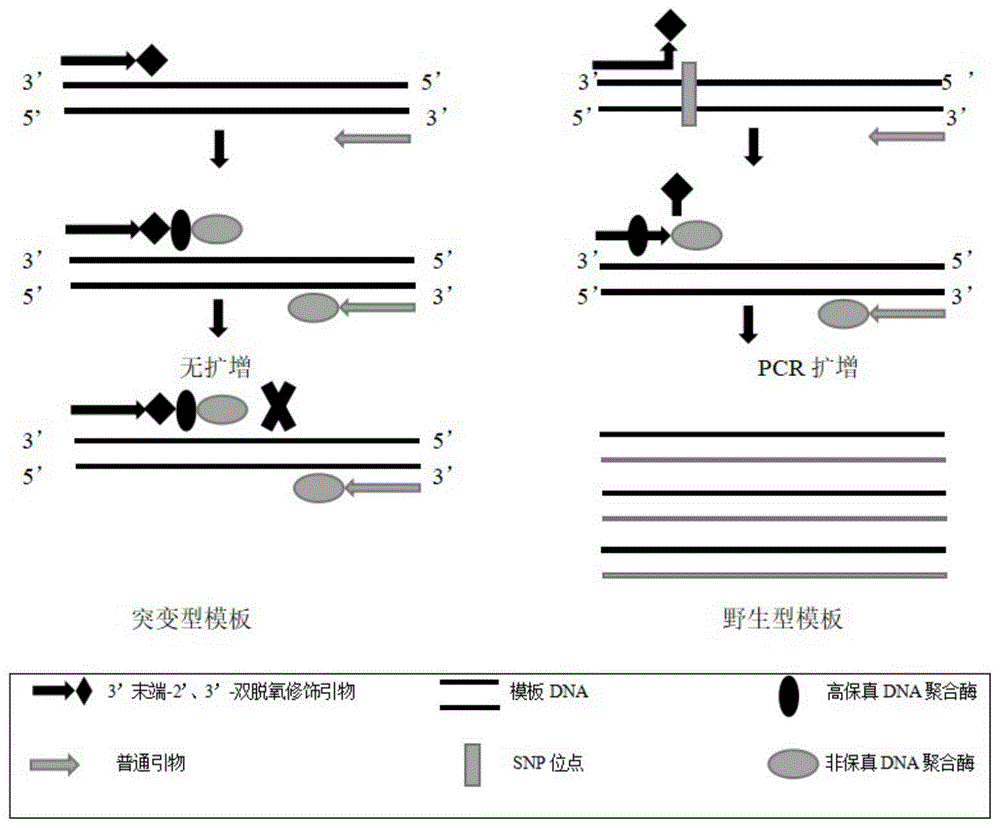
图1为基于PCR引物3’末端双脱氧修饰的待测基因的SNP检测方法原理图(以正向引物作为3’-双脱氧修饰引物为例)。
图2为基于PCR引物3’末端核苷酸不同修饰方式的待测基因的SNP检测琼脂糖凝胶电泳结果图。
图3为基于PCR引物3’末端核苷酸双脱氧修饰的待测基因的SNP检测方法,不同的高保真酶与非保真酶混合体系的实验琼脂糖凝胶电泳结果图。
图4为基于PCR引物3’末端核苷酸双脱氧修饰的待测基因的SNP检测方法,不同退火温度实验的琼脂糖凝胶电泳结果图。
图5为基于PCR引物3’末端核苷酸双脱氧修饰的待测基因的SNP检测方法灵敏度分析。
图6为基于PCR引物3’末端核苷酸双脱氧修饰的待测基因的SNP检测方法特异性分析。
图7为胶体金免疫层析试纸条原理示意图,分为内参线(C线),检测线(T线)及加样区。
图8为基于引物3’末端核苷酸双脱氧修饰结合5’末端标记的不同模板的扩增产物的胶体金免疫层析结果。
注:以上图中,“WT”代表野生型(Wild Type);“MT”代表突变型(Mutunt Type);“NC”代表阴性对照(Negative Control);“M”为DL2000DNAMarker(TaKaRa)。
具体实施方式
以水稻株高相关基因为例,针对于该基因的单核苷酸多态性(SNP),本发明人设计了一种基于引物3’末端双脱氧修饰结合胶体金免疫层析的新型检测方法,保留了引物延伸法快速、高效、灵敏度高、可操作性强的特点,又从根本上解决了假阳性的产生,为植物基因以及其它相关基因的SNP检测提供了一种便捷、灵敏、低廉而又准确的方法。
本发明人通过针对待测基因的SNP设计引物,并对正向引物中的3’末端核苷酸进行-双脱氧修饰从而造成“封闭”效果,再结合反应体系中高保真DNA聚合酶的3’,5’-核酸外切酶活性,非高保真DNA聚合酶的聚合作用,从而达到有效检测SNP的目的。本发明的方法只需针对待检测的SNP位点设计一对引物,通过一个常规PCR反应,外加凝胶电泳(琼脂糖凝胶或聚丙烯酰胺凝胶)分离,即可达到检测SNP基因型的目的。经验证,本发明的方法操作简单,无需依赖昂贵试剂、精密仪器,且灵敏度高、准确性强。且在引物3’末端双脱氧修饰后,对引物5’末端进行DIG、TAMRA和Biotin标记,结合胶体金免疫层析检测,替换掉琼脂糖凝胶电泳过程,降低检测时间。适用于植物基因的SNP的检测以及其它相关基因SNP的检测。
本发明通过合成双脱氧引物进行PCR反应检测SNP的原理为:首先设计一条3’末端核苷酸经双脱氧修饰的引物,该引物自3’末端碱基完全覆盖待测的SNP位点,且引物序列和基因型为突变型的等位基因序列完全互补配对。再设计一条下游(或上游)引物,与待测DNA样品一同进行PCR反应,当待测DNA样品SNP位点处的基因型为突变型时,由于3’末端核苷酸是经过双脱氧修饰的,因此下一个dNTP原料由于无法与该3’末端核苷酸正常形成3’,5’-磷酸二酯键,因而无法添加上去,即该条引物被“封闭”,无法顺利进行延伸,因此最终结果是无法扩增出包含SNP位点的目的片段。凝胶电泳检测结果为无目的扩增条带;而当待测的DNA样品基因型为野生型时,由于在SNP位点处,碱基序列发生了改变(置换、增添或缺失),导致该双脱氧修饰引物的3’末端与此模板无法互补配对,即形成了错配,这样在PCR反应体系中高保真DNA聚合酶3’,5’-核酸外切酶活性的作用下,该3’末端包括双脱氧修饰的核苷酸在内的错配的几个核苷酸就会被逐个切除下来,直至引物剩余的若干寡核苷酸可以和模板序列完全互补配对为止。由此,切除错配碱基后,解除了双脱氧修饰的“封闭”作用,因此在DNA聚合酶的作用下可以顺利延伸,最终与另一条引物一起,经过PCR反应可以扩增得到一条覆盖SNP位点的大小约106bp的目的片段。经凝胶电泳分离后,可得到一条清晰的条带,与之前描述的基因型为突变型的DNA样品PCR反应后的凝胶电泳分离结果形成鲜明对比。由此达到检测SNP位点的目的。3’末端可被修饰的核苷酸包括A、T、C、G,本发明以引物末端C为例进行实验研究。
本发明在双脱氧引物修饰的基础之上,充分利用引物5’末端可修饰的优势,在引物5’末端进行DIG、TAMRA和Biotin标记。在解决假阳性干扰的基础之上,缩短检测时间,由常规的琼脂糖凝胶电泳进化为胶体金层析试纸条,检测时间由30-50min缩短为2-10min,大大降低检测时间,节约检测成本。结果可视化,操作简单,对操作人员、仪器的要求极低。
本发明所述的PCR反应体系包括;PCR反应缓冲液(高保真DNA聚合酶2XTaqMasterMix II(Dye Plus),非保真DNA聚合酶2X TaqMasterMix(Dye Plus)),一对上/下游引物,待测DNA样品,灭菌去离子水。上游引物为辨别性引物,SNP位点碱基与双脱氧修饰的3’末端碱基序列相同。对于高保真DNA聚合酶2X Taq MasterMix II(Dye Plus),主要利用其3’,5’-核酸外切酶活性(校对功能),对于非保真DNA聚合酶2X TaqMasterMix(DyePlus),主要利用其DNA聚合活性(引物延伸活性)。使用常规PCR仪进行反应。
作为本发明的一种优选方式,本发明的方法操作步骤如下:
1.确定待测基因的SNP位点,明确SNP位点突变前后序列。
2.根据待测基因的SNP位点,比对野生型与突变体的序列,设计两条特异性引物,其中一条引物3’末端碱基恰好位于SNP处,另一条引物在其下游(或上游),一对引物扩增片段为106bp左右,设计的引物序列如表1。对于突变类型为数个碱基的置换、增添或缺失时,可当作单碱基处理。
表1
3.引物合成及修饰。对特异性引物即覆盖SNP位点的引物3’末端核苷酸进行双脱氧修饰或5’末端标记,交予上海铂尚生物技术有限公司进行合成并纯化。
4.待测水稻DNA提取。水稻的样本可采用种子、幼苗、叶片、穗子等组织或器官,优选种子或叶片。具体在本实施例中,将水稻嫩叶剪直径约2-3厘米的部分,置于96深孔板中,每孔加入2粒5mm钢珠及400μL TPS溶液,60HZ,1min进行破碎;65℃烘箱加热20min,期间轻微颠倒混匀;在4000rpm离心20min,吸取上清液200μL转移至新的离心管中,加入等体积预冷的异丙醇,轻轻颠倒混匀,4000rpm离心20min;弃上清液,加入400μL75%乙醇,洗涤2遍,获得全基因组DNA,过夜至晾干为止,加入200μL无菌水溶解备用。
5.PCR反应。配制PCR反应体系20μL,其中20ng的水稻基因组模板,其中高保真DNA聚合酶(2X Taq MasterMix II(Dye Plus))用量为5μL-7.5μL(20μL体系),其中非保真DNA聚合酶(2X Taq MasterMix(Dye Plus))用量为2.5μL-5μL(20μL体系)。上下游引物各1μL,补充灭菌去离子水至20μL。PCR反应条件如下:盖子加热至105℃;95℃预热3min;95℃变性30s;65℃退火30S;65℃延伸30S;重复35个循环;72℃保持10min。退火及延伸温度可根据设计的引物情况自行摸索、设定。或根据所用的Taq DNA聚合酶反应所需的环境条件自行优化反应体系与反应条件。反应在常规PCR仪中进行即可。
6.琼脂糖凝胶电泳(或聚丙烯酰胺凝胶电泳)分离。
7.根据电泳分离后的结果,即可判断待测DNA样品的SNP基因型:若分离后无条带,则待测DNA样品(等位基因)的基因型为突变型,即该基因待研究的SNP位点处序列发生了突变;若分离后有一条大小为106的条带,则待测DNA样品(等位基因)的基因型为野生型。
8.使用5’末端标记引物对,进行PCR反应,反应条件与程序与步骤5一致。取扩增产物用于检测,用样品稀释液对扩增产物稀释10倍待用,稀释倍数可根据目标物浓度进行调整。取出层析试纸条放于水平桌面上(尽快使用),注意不要触碰NC膜;用移液枪或滴管吸取80μL待检样品缓慢滴加在试纸条加样孔上,2-10分钟判读结果。
9.胶体金试纸条结果分析
本发明所述的方法适用于检测突变类型少数碱基核苷酸的置换、增添或缺失的SNP位点,尤其适用于单碱基核苷酸的置换、增添或缺失的SNP检测。
以下将结合具体实施方式对本发明进行详细描述。但这些实施方式并不限制本发明,本领域的普通技术人员根据这些实施方式所做出的结构、方法、或功能上的变换均包含在本发明的保护范围内。
下面实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到(例如,常规生化试剂商店购买得到的)。
待测水稻基因组DNA:
以下实施例中所用到的水稻基因组DNA样品,均为上海中科荃银分子育种技术有限公司提供。
试剂及仪器:
以下实施例中所用到的高保真DNA聚合酶(2X Taq MasterMix II(Dye Plus),非保真聚合酶(2X Taq Master Mix(Dye Plus)均购自购自南京诺唯赞生物科技股份有限公司;PCR反应所用引物均由上海铂尚生物技术有限公司合成并纯化;DIG抗体,Biotin抗体,TAMRA抗体均购自珠海博美生物科技有限公司。层析试纸条购自西安金磁纳米生物技术有限公司。
基因序列信息:
以下实施例中,设计引物所参考的水稻株高相关基因全长(参考序列)下载自国家水稻数据中心(China Rice Data Ceter),其登录号为:Os05g0514600。
实施例1:引物3’末端不同修饰方式对PCR反应的封闭效果
本发明人选取待测基因序列片段,分别为野生型WT(SEQ ID NO:5)和突变体MT(SEQ ID NO:6)。在这两条序列中存在一个单核苷酸突变位点,由野生型的T突变为C。据此,设计正向和反向引物,反向引物位于突变点下游与相应位置野生型和突变型模板完全匹配;正向引物与突变型模板完全匹配,其3’末端碱基位于单核苷酸突变位点处(如表1所示),对正向引物3’末端碱基分别进行磷酸化修饰及双脱氧修饰ddC。修饰的正向引物分别命名为正向引物-ddC、正向引物-P。
表2
引物序列如下:
正向引物:5’-TGGCGCTCTCGTGAGGTCGCTGCATAC-3’(SEQ ID NO:1)
反向引物:5’-ACTACTATTTCACGTTTACTTGACGA-3’(SEQ ID NO:2)
正向引物-ddC:5’-TGGCGCTCTCGTGAGGTCGCTGCATA-ddC-3’(SEQ ID NO:3)
正向引物-P:5’-TGGCGCTCTCGTGAGGTCGCTGCATAC-P-3’(SEQ ID NO:4)
反应体系如下:
反应条件如下:
95℃3min
95℃30S
65℃30S
65℃30S
72℃10min
10℃+∞
按上述反应体系及反应条件在常规PCR仪上进行反应,获得扩增产物进行琼脂糖凝胶电泳,所用凝胶浓度为2%,分子量标记为购自TaKaRa公司的DL2000 DNAMarker,电泳结果如图2所示,从图中可看到,在非保真酶体系下,无修饰的引物在野生型及突变体模板中均有显著条带;磷酸化修饰下,野生型和突变体出现微弱条带;ddc修饰下,野生型和突变体均无条带出现;与磷酸化修饰相比ddC修饰引物的封闭效果最佳,可完全抑制PCR反应。
实施例2:高保真酶与非保真酶混合体系介导的3’末端核苷酸双脱氧修饰(ddC)待测基因的SNP检测方法。
本发明人在研究中发现,高保真酶的3’→5'外切酶活性不是完全特异性的,不仅可以切除并校正与模板不匹配的引物,对与模板完全匹配的引物也有一定的外切酶活性。因此如果在反应体系中只单独使用高保真酶的话不能达到选择性扩增的效果,即对于突变型和野生型模板都能扩增,容易造成假阳性结果。因此,本发明人在反复研究后发现,在PCR反应中,将高保真DNA聚合酶与非高保真DNA聚合酶以适当比例混合使用,将大大提高检测的灵敏度及准确性。加入少量高保真DNA聚合酶,主要发挥校对功能:切除3’羟基被封闭的核苷,暴露出正常3’羟基;至于非高保真DNA聚合酶,主要发挥聚合酶作用进行PCR扩增反应。
使用与实施例1中完全相同的野生型模板序列(SEQ ID NO:6)突变型模板序列MT(SEQ ID NO:7),使用实施例1中的正向引物-ddC(SEQ ID NO:3)和反向引物(SEQ ID NO:2)为上下游引物,本发明人研究发现高保真聚合酶与非保真聚合酶的比例为(1-3):1,较佳地为(1.5-2):1,配制PCR反应体系20μL,其中高保真DNA聚合酶(2X Taq MasterMix II(DyePlus))用量为5μL-7.5μL(20μL体系),其中非保真DNA聚合酶(2XTaq MasterMix(DyePlus))用量为2.5μL-5μL(20μL体系)。按下列条件进行PCR反应。
反应体系如下:
反应条件如下:
95℃3min
95℃30S
65℃30S
65℃30S
72℃10min
10℃+∞
按上述反应体系及反应条件在常规PCR仪上进行反应,获得扩增产物进行琼脂糖凝胶电泳,所用凝胶浓度为2%,分子量标记为购自TaKaRa公司的DL2000 DNAMarker,电泳结果如图3所示,可看出高保真聚合酶与非保真聚合酶的比例为1:1时,野生型和突变体均无条带,推测是高保真酶的量偏少,无法发挥出校对功能;1.5:1时,仅野生型出现微弱条带;2:1时,突变体出现微弱条带,野生型条带明显亮突变体;2.5:1及3:1野生型和突变体条带均有目的较亮条带,无法区分。所以可在高保真聚合酶与非保真聚合酶的比例为2:1的体系中通过优化退火温度来达到理想效果,即突变型模板无条带而野生型模板有条带。
实施列3:退火温度影响的3’末端核苷酸双脱氧修饰(ddC)待测基因的SNP检测方法。
使用与实施例1中完全相同的野生型模板序列(SEQ ID NO:6)突变型模板序列MT(SEQ ID NO:7),使用实施例1中的正向引物-ddC(SEQ ID NO:3)和反向引物(SEQ ID NO:2)为上下游引物,本发明人研究发现当高保真聚合酶与非保真聚合酶的比例为为2:1时,做温度PCR反应,退火温度分别为64.8℃、65℃、65.2℃、65.4℃、65.6℃。按下列条件进行PCR反应。
反应条件如下:
95℃3min
95℃30S
64.6-65.6℃30S
64.6-65.6℃30S
72℃10min
10℃+∞
按上述反应体系及反应条件在常规PCR仪上进行反应,获得扩增产物进行琼脂糖凝胶电泳,所用凝胶浓度为2%,分子量标记为购自TaKaRa公司的DL2000 DNAMarker,电泳结果如图4所示,当温度<65.2℃时,野生型及突变体均有目的条带,突变体较野生型条带亮度稍弱,当温度≥65.2℃时,突变体无条带,只有野生型均有目的条带,但是随着温度逐渐伸高,野生型条带亮度逐渐减弱,故反应的最适温度为65.2℃。
实施列4:基于PCR引物3’末端核苷酸双脱氧修饰的待测基因的SNP检测方法灵敏度分析。
使用与实施例1中完全相同的野生型模板序列(SEQ ID NO:6)突变型模板序列MT(SEQ ID NO:7),将这两个DNA序列分别连接到pMD S,把重组质粒转化入DH5a感受态细胞中,挑取单菌落扩大培养,经菌液PCR筛选出阳性克隆后,送至上海铂尚生物技术有限公司测序,最终挑选出序列完全正确的重组质粒。用限制性内切酶SacI(大连TaKaRa公司)分别对这两种重组质粒进行单酶切,并对酶切后得到的线性质粒进行纯化,之后用超微量紫外分光光度计分别测定其OD值,并换算为拷贝/ml。用1xTE以10倍作为梯度标准对两种线性质粒进行逐步稀释,最终制备成10
反应条件如下:
95℃3min
95℃30S
65.2℃30S
65.2℃30S
72℃10min
10℃+∞
按上述反应体系及反应条件在常规PCR仪上进行反应,获得扩增产物进行琼脂糖凝胶电泳,所用凝胶浓度为2%,分子量标记为购自TaKaRa公司的DL2000 DNAMarker,电泳结果如图5所示,从电泳图中可以明显看出,当反应体系(20μL)中模板含量分别为1x10
实施列5、基于PCR引物3’末端核苷酸双脱氧修饰待测基因的SNP检测方法特异性分析。
使用与实施例1中完全相同的野生型模板序列(SEQ ID NO:6)突变型模板序列MT(SEQ ID NO:7),反应体系(20μL)中加入的野生型(MT)序列模板所占的比例分别按100%、50%、10%、1%、0.1%、0%依次递减,此外其余反应体系组分及反应条件等均与实施例4中描述的相同,经PCR反应之后,对反应物进行琼脂糖凝胶电泳鉴定(所用凝胶浓度为2%。电泳结果如图6所示。从电泳图中可以明显看出,当反应体系(20μL)中突变型(MT)模板量占总模板量的比例分别为100%、50%、10%、时,均有目的条带,且目的条带亮度随体系中突变型模板比例的降低而减弱。当野生型模板质粒所占比例为0%,即所加模板均为突变体模板时,则无目的条带。
实施列6、基于PCR引物3’末端核苷酸双脱氧修饰结合胶体金免疫层析的待测基因的SNP检测方法。
使用与实施例1中完全相同的野生型模板序列(SEQ ID NO:6)突变型模板序列MT(SEQ ID NO:7),使用表1的5’修饰特异引物对SEQ ID NO:8与SEQ ID NO:9,内参引物对SEQID NO:10与SEQ ID NO:11,进行PCR反应,获得扩增产物。取扩增产物用于检测,用样品稀释液对扩增产物稀释10倍。取出层析试纸条放于水平桌面上(尽快使用),注意不要触碰NC膜;用移液枪或滴管吸取80μL待检样品缓慢滴加在试纸条加样孔上,2-10分钟判读结果。结果如图8所示,当模板为野生型时,内参带(R线)与检测带(T线)都出现红色条带;当模板为突变型,仅内参带(R线)出现红色条带。
综上所述,本发明对引物3’末端核苷酸进行双脱氧修饰(ddC),使用高保真DNA聚合酶与非高保真DNA聚合酶混合体系,结合利用高保真酶的3’→5'外切酶活性,非保真酶的DNA聚合活性,将大大提高检测的灵敏度及准确性。并在3’末端双脱氧修饰(ddC)的基础上对引物5’末端进行标记,采用胶体金免疫层析试纸检测,在排除假阳性干扰的情况下,大大缩短结果可视化时间。对仪器设备要求低,低成本、易操作、适合广大的育种工作者。
应当理解,上文所列出的一系列的详细说明仅仅是针对本发明的可行性实施方式的具体说明,它们并非用以限制本发明的保护范围,凡未脱离本发明技艺精神所作的等效实施方式或变更均应包含在本发明的保护范围之内。
- 一种地下停车场双井式钢结构车辆输送架
- 基于预制地下连续墙结构的装配式支护结构施工工艺
- 软基自沉式地下空间结构构造及施工工艺
- 一种结构简单通用性强的双井式地下水循环施工工艺
- 一种结构简单通用性强的双井式地下水循环施工工艺