Fab改造诱导形成的双特异性抗体及其制备方法和用途
文献发布时间:2023-06-19 11:54:11
技术领域
本发明属于抗体工程领域,具体涉及一种Fab改造诱导形成的双特异性抗体及其制备方法和用途。
背景技术
双特异性抗体有多种构建方式,其中IgG型双特异性抗体具有和普通抗体相似的结构、理化性质和Fc段功能。通常IgG型双特异性抗体由两条氨基酸序列不同的重链(即抗抗原A的重链HC_A和抗抗原B的重链HC_B)和两条氨基酸序列不同的轻链(即抗抗原A的轻链LC_A和抗抗原B的轻链LC_B)组成。当4条多肽链组合时,两条重链之间会形成同源二聚体和异源二聚体,轻重链之间也会形成错配,因此会产生8种不同的组合方式,其中只有一种为所需要的目标抗体分子。而从8种分子中分离纯化得到目标分子效率极低且非常困难。
抗体工程领域的进展使得IgG型双特异性抗体的制备取得了重大的进展。大多数IgG型双特异性抗体通过改造Fc段促使抗体的两条重链形成异源二聚体(Ridgway,Prestaet al.1996;Carter 2001,US2010286374A1,CN106883297A,US20150307628A1),然而,由于轻重链之间相互作用更加复杂,使得轻重链的特异性配对更加困难。具体来说,理想的状态是,构成双特异性抗体的两条重链和两条轻链中,轻链LC_A只特异性的和重链HC_A配对,而不会和重链HC_B配对,同时轻链LC_B只特异性的和重链HC_B配对,而不会和重链HC_A配对。CN104968677A;WO2016172485A2;WO2014082179A;Nat Biotechnol.2014Feb;32(2):191-8.;Protein Sci.2017Oct;26(10):2021-2038.;Protein Eng Des Sel.2017Sep 1;30(9):685-696.;公开了如何使轻重链正确配对的方法,然而本领域仍需要寻找合适的优化以进一步提高轻链配对的特异性、减少错配的副产物以及双特异性抗体的产率。
发明内容
本发明的目的在于克服现有技术中的不足,提供一种Fab改造诱导形成的双特异性抗体及其制备方法和用途。本发明通过综合考虑界面氨基酸之间的各种相互作用,如静电作用、空间作用等,在抗体轻重链相互作用界面特定位置引入了氨基酸修饰,并且在重链、轻链中引入连接肽进行优化,使得轻重链正确配对比例提高到99%以上,而且显著的降低了氨基酸修饰对突变体表达量的影响。
本发明的第一个方面是提供一种双特异性抗体,所述双特异性抗体包含:可与某一特异性抗原结合的重链A及与所述重链A配对的轻链a,以及可与另一特异性抗原结合的重链B及与所述重链B配对的轻链b;重链A和重链B均具有抗体重链可变区VH结构域、抗体重链恒定区CH1结构域、CH2结构域、CH3结构域,轻链a和轻链b均具有抗体轻链可变区VL结构域和轻链恒定区CL结构域;重链A的VH结构域和CH1结构域之间插入连接肽、和/或重链B的VH结构域和CH1结构域之间插入连接肽、和/或轻链a的VL结构域和CL结构域之间插入连接肽、和/或轻链b的VL结构域和CL结构域之间插入连接肽;与野生型的人抗体相比,所述重链A和轻链a、重链B和轻链b具有选自下列的突变中的一种或多种:(a)VH结构域的Q39发生突变,且VL结构域的Q38发生突变;(b)CH1结构域的L145和/或L128发生突变,且CL结构域的V133发生突变;以上所述的氨基酸的位置根据KABAT编号的EU索引确定。
应当理解的是,如果存在,重链A的VH结构域和CH1结构域之间插入的连接肽、重链B的VH结构域和CH1结构域之间插入的连接肽、轻链a的VL结构域和CL结构域之间插入的连接肽、轻链b的VL结构域和CL结构域之间插入的连接肽可以是全部相同的、或部分相同的、或各不相同的。
优选地,所述连接肽长度为1-4个氨基酸。即如果存在,重链A的VH结构域和CH1结构域之间插入的连接肽长度为1-4个氨基酸;如果存在,重链B的VH结构域和CH1结构域之间插入的连接肽长度为1-4个氨基酸;如果存在,轻链a的VL结构域和CL结构域之间插入的连接肽长度为1-4个氨基酸;如果存在,轻链b的VL结构域和CL结构域之间插入的连接肽长度为1-4个氨基酸。
优选地,所述连接肽选自:G,GG,GS,SG,SS,GGG,GGS,GSG,SGG,GSS,SGS,SSG,SSS,GGGG,GGGS,GGSG,GSGG,SGGG,GGSS,SSGG,GSSG,GSGS,SGSG,SGGS,GSSS,SGSS,SSGS,SSSG,A,AA,AS,SA,SS,AAA,AAS,ASA,SAA,ASS,SAS,SSA,SSS,AAAA,AAAS,AASA,ASAA,SAAA,AASS,SSAA,ASSA,ASAS,SASA,SAAS,ASSS,SASS,SSAS,SSSA,GA,AG,GGA,GAG,AGG,GAA,AGA,AAG,GGGA,GGAG,GAGG,AGGG,GGAA,AAGG,GAAG,GAGA,AGAG,AGGA,GAAA,AGAA,AAGA,AAAG,或其他任意氨基酸的组合。
优选地,本发明的双特异性抗体中的所述重链A和重链B、轻链a和轻链b含有选自以下一组的突变:(1)重链A的L145突变为带正电荷氨基酸,轻链a的V133突变为带负电荷氨基酸,且重链B的L145突变为带负电荷氨基酸,轻链b的V133突变为带正电荷氨基酸;(2)重链A的L128突变为带正电荷氨基酸,轻链a的V133突变为带负电荷氨基酸,且重链B的L128突变为带负电荷氨基酸,轻链b的V133突变为带正电荷氨基酸;(3)重链A的L145突变为带正电荷氨基酸,轻链a的V133突变为带负电荷氨基酸,且重链B的L128突变为带负电荷氨基酸,轻链b的V133突变为带正电荷氨基酸;(4)重链A的L128突变为带正电荷氨基酸,轻链a的V133突变为带负电荷氨基酸,且重链B的L145突变为带负电荷氨基酸,轻链b的V133突变为带正电荷氨基酸;(5)重链A的Q39、L145突变为带正电荷氨基酸,轻链a的Q38、V133突变为带负电荷氨基酸,且重链B的Q39、L145突变为带负电荷氨基酸,轻链b的Q38、V133突变为带正电荷氨基酸;(6)重链A的Q39、L128突变为带正电荷氨基酸,轻链a的Q38、V133突变为带负电荷氨基酸,且重链B的Q39、L128突变为带负电荷氨基酸,轻链b的Q38、V133突变为带正电荷氨基酸;(7)重链A的Q39、L145突变为带正电荷氨基酸,轻链a的Q38、V133突变为带负电荷氨基酸,且重链B的Q39、L128突变为带负电荷氨基酸,轻链b的Q38、V133突变为带正电荷氨基酸;(8)重链A的Q39、L128突变为带正电荷氨基酸,轻链a的Q38、V133突变为带负电荷氨基酸,且重链B的Q39、L145突变为带负电荷氨基酸,轻链b的Q38、V133突变为带正电荷氨基酸。
进一步优选地,所述带正电荷氨基酸指K(赖氨酸)或R(精氨酸),所述带负电荷氨基酸指D(天冬氨酸)或E(谷氨酸),所述重链A和重链B、轻链a和轻链b含有选自以下一组的突变:1)重链A:L145K或L145R,轻链a:V133D或V133E,且重链B:L145D或L145E,轻链b:V133K或V133R;2)重链A:L128K或L128R,轻链a:V133D或V133E,且重链B:L128D或L128E,轻链b:V133K或V133R;3)重链A:L145K或L145R,轻链a:V133D或V133E,且重链B:L128D或L128E,轻链b:V133K或V133R;4)重链A:L128K或L128R,轻链a:V133D或V133E,且重链B:L145D或L145E,轻链b:V133K或V133R;5)重链A:(Q39K或Q39R)+(L145K或L145R),轻链a:(Q38D或Q38E)+(V133D或V133E),且重链B:(Q39D或Q39E)+(L145D或L145E),轻链b:(Q38K或Q38R)+(V133K或V133R);6)重链A:(Q39K或Q39R)+(L128K或L128R),轻链a:(Q38D或Q38E)+(V133D或V133E),且重链B:(Q39D或Q39E)+(L128D或L128E),轻链b:(Q38K或Q38R)+(V133K或V133R);7)重链A:(Q39K或Q39R)+(L145K或L145R),轻链a:(Q38D或Q38E)+(V133D或V133E),且重链B:(Q39D或Q39E)+(L128D或L128E),轻链b:(Q38K或Q38R)+(V133K或V133R);8)重链A:(Q39K或Q39R)+(L128K或L128R),轻链a:(Q38D或Q38E)+(V133D或V133E),且重链B:(Q39D或Q39E)+(L145D或L145E),轻链b:(Q38K或Q38R)+(V133K或V133R)。
其中,Q39R是指Gln39被替换为精氨酸(R)、Q39K是指Gln39被替换为赖氨酸(K)、Q39E是指Gln39被替换为谷氨酸(E)、Q39D是指Gln39被替换为天冬氨酸(D)、Q38R是指Gln38被替换为精氨酸(R)、Q38K是指Gln38被替换为赖氨酸(K)、Q38E是指Gln38被替换为谷氨酸(E)、Q38D是指Gln38被替换为天冬氨酸(D)、L145R是指亮氨酸145被替换为精氨酸(R)、L145K是指亮氨酸145被替换为赖氨酸(K)、L145E是指亮氨酸145被替换为谷氨酸(E)、L145D是指亮氨酸145被替换天冬氨酸(D)、L128R是指亮氨酸128被替换为精氨酸(R)、L128K是指亮氨酸128被替换为赖氨酸(K)、L128E是指亮氨酸128被替换为谷氨酸(E)、L128D是指亮氨酸128被替换为天冬氨酸(D)、V133R是指缬氨酸133被替换为精氨酸(R)、V133K是指缬氨酸133被替换为赖氨酸(K)、V133E是指缬氨酸133被替换为谷氨酸(E)、V133D是指缬氨酸133被替换为天冬氨酸(D)。
优选地,重链A和重链B的CH3结构域分别命名为CH3_A结构域和CH3_B结构域,与野生型的人抗体重链恒定区CH3结构域相比,所述CH3_A和CH3_B结构域含有有利于形成双特异性抗体的突变,不局限于例如WO9627011、CN101198698B、CN102459346B、CN105051069A、US2016177364A1、US2010286374A1、CN106883297A、US20150307628A1、CN104968677A、NatBiotechnol.2014Feb;32(2):191-8。本发明的实施方案中,涉及如下所示位置氨基酸的突变中的一种从而有利于形成双特异性抗体:(a1)CH3_A结构域:F405E+K409F+K370D,CH3_B结构域:S364R+E357S;(a2)CH3_A结构域:F405E+K409F+K370D+S354C,CH3_B结构域:S364R+E357S+Y349C;(a3)CH3_A结构域:F405E+K409F+K370D+Y349C,CH3_B结构域:S364R+E357S+S354C(b1)CH3_A结构域:F405E+K409F+K392D,CH3_B结构域:D399K;(b2)CH3_A结构域:F405E+K409F+K392D+S354C,CH3_B结构域:D399K+Y349C;(b3)CH3_A结构域:F405E+K409F+K392D+Y349C,CH3_B结构域:D399K+S354C;(c1)CH3_A结构域:F405E+K409F+K439D,CH3_B结构域:E356K+E357K;(c2)CH3_A结构域:F405E+K409F+K439D+S354C,CH3_B结构域:E356K+E357K+Y349C;(c3)CH3_A结构域:F405E+K409F+K439D+Y349C,CH3_B结构域:E356K+E357K+S354C;(d1)CH3_A结构域:F405E+K409F+L368D,CH3_B结构域:S364R;(d2)CH3_A结构域:F405E+K409F+L368D+S354C,CH3_B结构域:S364R+Y349C;(d3)CH3_A结构域:F405E+K409F+L368D+Y349C,CH3_B结构域:S364R+S354C;(e1)CH3_A结构域:F405E+K409F+L368D,CH3_B结构域:S364K;(e2)CH3_A结构域:F405E+K409F+L368D+S354C,CH3_B结构域:S364K+Y349C;(e3)CH3_A结构域:F405E+K409F+L368D+Y349C,CH3_B结构域:S364K+S354C;(f1)CH3_A结构域:F405E+K409F+K360E,CH3_B结构域:Q347R;(f2)CH3_A结构域:F405E+K409F+K360E+S354C,CH3_B结构域:Q347R+Y349C;(f3)CH3_A结构域:F405E+K409F+K360E+Y349C,CH3_B结构域:Q347R+S354C;(g1)CH3_A结构域:F405E+K409F+K370D+K360E,CH3_B结构域:S364R+E357S+Q347R;(g2)CH3_A结构域:F405E+K409F+K370D+K360E+S354C,CH3_B结构域:S364R+E357S+Q347R+Y349C;(g3)CH3_A结构域:F405E+K409F+K370D+K360E+Y349C,CH3_B结构域:S364R+E357S+Q347R+S354C。
Q347R是指谷氨酰胺Gln347被替换为精氨酸(R)、Y349C是指酪氨酸Tyr349被替换为半胱氨酸(C)、S354C是指丝氨酸Ser354被替换为半胱氨酸(C)、E356K是指谷氨酸Glu356被替换为赖氨酸(K)、E357K是指谷氨酸Glu357被替换为赖氨酸(K)、E357S是指谷氨酸Glu357被替换为丝氨酸(S)、K360E是指赖氨酸Lys360被替换为赖氨酸(K)、S364R是指丝氨酸Ser364被替换为精氨酸(R)、S364K是指丝氨酸Ser364被替换为赖氨酸(K)、L368D是指亮氨酸Leu368被替换为天冬氨酸(D)、K370D是指赖氨酸Lys370被替换为天冬氨酸(D)、K392D是指赖氨酸Lys392被替换为天冬氨酸(D)、D399K是指天冬氨酸Asp399被替换为赖氨酸(K)、K439E是指赖氨酸Lys439被替换为谷氨酸(E)、F405E是指苯丙氨酸Phe405被替换为谷氨酸(E)、K409F是指赖氨酸Lys409被替换为苯丙氨酸(F)。
本发明的第二个方面是提供一种组合物,其含有:(1)本发明第一个方面所述的异源二聚体,以及(2)药学上可接受的载体和/或稀释剂和/或赋形剂。
本发明的第三个方面是提供一种多核苷酸,所述多核苷酸包含:编码本发明第一个方面所述的双特异性抗体的重链A的核苷酸分子A,编码本发明第一个方面所述的双特异性抗体的轻链a的核苷酸分子a,编码本发明第一个方面所述的双特异性抗体的重链B的核苷酸分子B,编码本发明第一个方面所述的双特异性抗体的轻链b的核苷酸分子b。
本发明的第四个方面是提供一种载体组合,所述载体组合包括:选自含有核苷酸分子A的重组载体、含有核苷酸分子a的重组载体、同时含有所述核苷酸分子A和核苷酸分子a的重组载体、含有所述核苷酸分子B的重组载体、含有核苷酸分子b的重组载体、同时含有所述核苷酸分子B和核苷酸分子b的重组载体中的两种或以上,且所述载体组合同时含有核苷酸分子A、核苷酸分子a、核苷酸分子B和核苷酸分子b。
其中,上述各重组载体所使用的表达载体为本领域常规的表达载体,是指包含适当的调控序列,例如启动子序列、终止子序列、多腺苷酰化序列、增强子序列、标记基因和/或序列以及其他适当的序列的表达载体。所述表达载体可以是病毒或质粒,如适当的噬菌体或者噬菌粒,更多技术细节请参见例如Sambrook等,Molecular Cloning:A LaboratoryManual,第二版,Cold Spring Harbor Laboratory Press,1989。许多用于核酸操作的已知技术和方案请参见Current Protocols in Molecular Biology,第二版,Ausubel等编著。本发明所述表达载体较佳地为pDR1,pcDNA3.1(+),pcDNA3.1/ZEO(+),pDHFR,pTT5,pDHFF,pGM-CSF或pCHO 1.0,更佳地为pTT5。
本发明的第五个方面是提供一种重组宿主细胞,所述重组宿主细胞含有所述载体组合。
本发明所述的重组宿主细胞的原始宿主细胞可以为本领域常规的各种宿主细胞,只要能满足使上述重组载体稳定地自行复制,且所携带所述的核苷酸可被有效表达即可。其中所述原始宿主细胞可以原核表达细胞或真核表达细胞,所述宿主细胞较佳地包括:COS、CHO(中国仓鼠卵巢,Chinese H amster Ovary)、NS0、sf9、sf21、DH5α、BL21(DE3)或TG1,更佳地为E.coli TG1、BL21(DE3)细胞(表达单链抗体或Fab抗体)或者CHO-K1细胞(表达全长IgG抗体)。将前述表达载体转化至宿主细胞中,即可得本发明优选的重组宿主细胞。其中所述转化方法为本领域常规转化方法,较佳地为化学转化法,热激法或电转法。
作为优选的方案,所述重组宿主细胞的原始宿主细胞优选为真核细胞,进一步优选为CHO细胞或293E细胞。
本发明的第六个方面是提供本发明第一个方面所述的双特异性抗体,本发明第二个方面所述的组合物,本发明第三个方面所述的多核苷酸、本发明第四个方面所述的载体组合、或本发明第五个方面所述的重组宿主细胞在制备双特异性抗体、双特异性融合蛋白和抗体-融合蛋白嵌合体中的用途。
本发明的第七个方面是提供一种制备本发明第一个方面所述的双特异性抗体的方法,其特征在于,使用本发明第五个方面所述的重组宿主细胞表达所述双特异性抗体。
在本发明中,所述重组宿主细胞同时含有编码本发明第一个方面所述的双特异性抗体的重链A的核苷酸分子A、编码本发明第一个方面所述的双特异性抗体的轻链a的核苷酸分子a、编码本发明第一个方面所述的双特异性抗体的重链B的核苷酸分子B、编码本发明第一个方面所述的双特异性抗体的轻链b的核苷酸分子b,利用该重组宿主细胞表达,回收,得到双特异性抗体。
其中,所述双特异性抗体可以用标准的实验手段从重组宿主细胞中纯化。例如,当异二聚体蛋白包含抗体Fc片段,可以用蛋白A来纯化。纯化方法包括但不限于色谱技术如体积排阻法、离子交换法、亲和色谱法及超滤法,或者上述各种方法的适当组合。
在本发明中,所述重组宿主细胞中核苷酸分子A、核苷酸分子a、核苷酸分子B和核苷酸分子b的摩尔比例为(1-3):(1-3):(1-3):(1-3),例如1:1:1:1、1:1:1.5:1.5、1:1:2:2、1:1:2.5:2.5、1:1:3:3、3:3:1:1、2.5:2.5:1:1、2:2:1:1、或1.5:1.5:1:1。
在本发明的实施方案中,其中所述的轻链选自κ链或λ链,其中所述的恒定区来源于IgG(例如IgG1、IgG2、IgG3、IgG4)、IgA(例如IgA1、IgA2)、IgD、IgE或IgM。
在本发明中,所述CH1和CL来源于抗体Fab片段,优选来源于人的抗体Fab片段。在一般情况下,人抗体Fab片段的CH1和CL结构域来源于野生型的人抗体Fab片段。本发明中人抗体Fab片段也包括对于野生型人抗体Fab序列的个别氨基酸的改变,例如包括某些在糖基化位点突变的氨基酸,或者其它无义的突变。除了本发明中提到的突变外,还可能含有其它不影响抗体Fab段功能的突变。
在本发明中,所述CH3来源于抗体Fc片段,优选来源于人的抗体Fc片段。在一般情况下,人抗体Fc片段的CH3结构域来源于野生型的人抗体Fc片段。野生型的人抗体Fc是指存在于人群中的氨基酸序列,当然Fc片段在个体中会有一些细微的差异。本发明中人抗体Fc片段也包括对于野生型人抗体Fc序列的个别氨基酸的改变,例如包括某些在糖基化位点突变的氨基酸,或者其它无义的突变。对于CH3以及CH2结构域,除了本发明中提到的突变外,还可能含有其它不影响抗体特别是Fc段功能的突变。
在本发明中,所述氨基酸位置的编号均根据Kabat EU编号索引的位置确定。本领域技术人员知晓,即使上述区域中由于氨基酸的插入或缺失或其它突变导致氨基酸序列的改变,根据Kabat EU编号索引确定的与标准序列对应的各氨基酸的位置编号仍然不变。所述EU索引描述于Kabat等,Sequences of Proteins of Immunological Interest,第5版Public Health Service,National Institutes of Health,Bethesda,MD.(1991)。
本发明的有益效果:
本发明通过综合考虑界面氨基酸之间的各种相互作用,如静电作用、空间作用等,在抗体轻重链相互作用界面特定位置引入了氨基酸修饰,且对重链VH/CH1之间连接肽和/或轻链VL/CL之间连接肽进行了优化,使得轻重链正确配对比例提高到99%以上,并显著的降低了氨基酸修饰对突变体表达量的影响,从而提高抗体的产量,降低生产成本。
附图说明
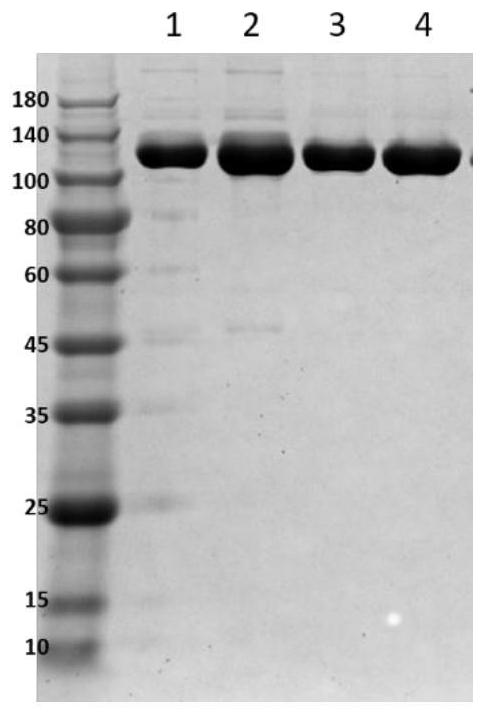
图1为轻重链配对的电泳分析结果。4-12%SDS-PAGE蛋白凝胶电泳。泳道从左到右依次为:蛋白分子量标准、A
图2为ELISA检测EGFR×HER2双特异性抗体与抗原EGFR-ECD-Fc结合活性。
图3为ELISA检测EGFR×HER2双特异性抗体与抗原HER2-ECD-Fc结合活性。
图4为ELISA检测EGFR×HER2双特异性抗体同时与抗原HER2-ECD-Fc和EGFR-ECD-Fc结合活性。
图5为ELISA检测EGFR×cMet双特异性抗体与抗原EGFR-ECD-Fc结合活性。
图6为ELISA检测EGFR×cMet双特异性抗体与抗原cMet-ECD-Fc结合活性。
图7为ELISA检测EGFR×cMet双特异性抗体同时与抗原cMet-ECD-Fc和EGFR-ECD-Fc结合活性。
具体实施方式
以下实施例、实验例是对本发明进行进一步的说明,不应理解为是对本发明的限制。实施例不包括对传统方法的详细描述,如那些用于构建载体和质粒的方法,将编码蛋白的基因插入到这样的载体和质粒的方法或将质粒引入宿主细胞的方法.这样的方法对本领域中具有普通技术的人员是众所周知的,并且在许多出版物中都有所描述,包括Sambrook,J.,Fritsch,E.F.and Maniais,T.(1989)Molecular Cloning:A Laboratory Manual,2
以下实施例中使用的实验材料和来源以及实验试剂的配制方法具体说明如下。
1、实验材料:
293E细胞:来自NRC biotechnology Research Institute。
2、实验试剂:
PBS:购自生工生物工程(上海)股份有限公司,货号B548117。
柠檬酸:购自国药集团化学试剂有限公司。
Prime star HS DNA polymerase:购自Takara公司,货号R010A。
无内毒素质粒大提试剂盒:购自TIANGEN公司,货号DP117。
3、实验仪器:
HiTrap MabSelectSuRe柱:购自GE公司。
AKTA-FPLC快速蛋白液相色谱系统:购自GE公司。
C1000 Touch Thermal Cycler PCR仪:购自Bio-Rad公司。
Chemidoc MP凝胶成像仪:购自Bio-Rad公司。
离心机:购自Eppendorf公司。
G1600AX毛细管电泳仪:购自安捷伦公司。
MicroCal PEAQ-DSC微量热差式扫描量热仪:购自马尔文公司。
Octet分子相互作用系统:购自ForteBio公司。
Xevo G2-XS Tof飞行时间质谱:购自Waters公司。
实施例1.CH1/CL点突变设计
当在一种细胞内共表达两种不同抗体分子时,4条多肽链重链A、轻链a、重链B和轻链b会随机配对产生双特异性抗体和错配抗体。为了减少错配的发生,我们希望构成双特异性抗体的两条重链和两条轻链中,轻链a只特异性的和重链A配对,而不会和重链B配对,同时轻链b只特异性的和重链B配对,而不会和重链A配对。因此需要对抗体轻重链进行工程改造。表1列举了抗体CH1/CL界面上相互作用的氨基酸,通过对这些氨基酸进行突变,有可能促进双特异性抗体轻重链的正确配对。抗体轻链选自κ链和λ链,轻链恒定区Cκ和Cλ上位于相互作用界面上的氨基酸高度保守。因此虽然本发明中抗体轻链上的所有突变均是在κ链上完成,但同样适用于λ链。
表1.CH1/CL界面上相互作用的氨基酸
1.1构建带有CH1/Cκ突变的重组抗体
将抗HER2抗体Trastuzumab的重链HC(SEQ ID NO:1)和轻链LC(SEQ ID NO:2)亚克隆至哺乳动物细胞表达载体pTT5获得用于哺乳动物细胞表达Trastuzumab的重组表达载体。将抗EGFR抗体Cetuximab的重链(SEQ ID NO:3)和轻链(SEQ ID NO:4)亚克隆至哺乳动物细胞表达载体pTT5获得用于哺乳动物细胞表达Cetuximab的重组表达载体。将抗IL17抗体anti-IL17Ab的重链(SEQ ID NO:5)和轻链(SEQ ID NO:6)亚克隆至哺乳动物细胞表达载体pTT5获得用于哺乳动物细胞表达anti-IL17Ab的重组表达载体。根据实施例1的表1所示,抗体轻链的EU编号133位的Val(或叫做V)和抗体重链上EU编号为128位的Leu(或叫做L)和145位的Leu之间存在相互作用。利用重叠PCR法对Trastuzumab、Cetuximab和anti-IL17Ab的重链L128或L145以及轻链的V133进行突变,最终分别得到表2所示突变体对映的用于在哺乳动物细胞中表达的突变载体。
1.2瞬时表达Trastuzumab、Cetuximab、anti-IL17Ab野生型和突变体,并检测不同突变组合对抗体表达量的影响
将步骤1的突变组合相应的表达载体用PEI转染至悬浮培养的293E细胞,且重链和轻链的重组表达载体共转比例为1:1。培养5~6天后,收集瞬时表达培养上清液,利用Fc捕获法通过Fortebio检测抗体的表达量。结果如表2所示,1)所有抗体的野生型和突变体都能正常表达,表明重链(L128或L145)和轻链V133引入带相反电荷的氨基酸不影响抗体轻重链的配对。2)Trastuzumab的突变体的表达量比野生型有明显的下降,这和WO2016172485A2中Figure 1A和1B展示了相似的解果。具体来说,WO2016172485A2报道了将抗VEGF抗体Ranibizumab的重链L128或L145突变为带负电荷氨基酸如D或E,同时将轻链V133突变为带正电荷氨基酸如R或K时,突变体蛋白表达量明显降低。3)然而本实施例发现,在Cetuximab和anti-IL17Ab的重链(L128或L145)和轻链V133引入带相反电荷的氨基酸后,突变体的表达量并没有降低。因此,在抗体重链(L128或L145)和轻链V133引入相反电荷的氨基酸并不一定影响抗体的表达,WO2016172485A2中Figure 1A和1B展示的结果并不具有普遍规律。
表2CH1/CL突变对抗体表达水平的影响
实施例2轻重链配对实验
2.1VH/VL和CH1/CL突变组合
在抗体VH/VL界面氨基酸上引入突变可以促进双特异性抗体轻重链的正确配对。CN104968677A、WO2016172485A2公开了在抗体重链的Q39和轻链的Q38引入带相反电荷的氨基酸,有利于双特异性抗体轻重链的正确配对。在实施例1的基础上,本实施例选择抗EGFR抗体Panitumumab、抗HER2抗体Trastuzumab和抗cMet抗体Onartuzumab作为模板,进一步设计了表3所示的突变组合。将Trastuzumab的重链HC(SEQ ID NO:1)和轻链LC(SEQ ID NO:2)、Onartuzumab的重链HC(SEQ ID NO:7)和轻链LC(SEQ ID NO:8)、Panitumumab的重链HC(SEQ ID NO:9)和轻链LC(SEQ ID NO:10)亚克隆至哺乳动物细胞表达载体pTT5获得用于哺乳动物细胞表达的重组表达载体。利用重叠PCR法对Trastuzumab、Panitumumab和Onartuzumab的重链HC和轻链LC编码基因进行组合突变,最终分别得到用于在哺乳动物细胞中表达突变体的重组表达载体。
表3
2.2在抗体VH/CH1之间和或VL/CL之间引入连接肽,有利于提高突变体的表达水平
实施例1表明抗体重链(L128或L145)和轻链V133突变引入带相反电荷的氨基酸,并不一定影响抗体的表达。本发明进一步创造性的发现,当抗体重链(L128或L145)和轻链V133突变引入带相反电荷的氨基酸导致表达量明显下降时,通过在抗体VH/CH1之间和或VL/CL之间引入连接肽有利于提高突变体的表达水平。具体来说,在抗体重链EU编号118位的Ala前插入连接肽linker1,在抗体轻链EU编号108位的Arg前插入连接肽linker2,linker1和linker2为长度为0-4个氨基酸的连接肽,2个氨基酸和4个氨基酸的连接肽序列,优选GG和GGGS。其中尤其以linker1为2个氨基酸的连接肽如“GG”效果最为明显。据此,构建如表4所示的突变体。将突变组合相应的表达载体用PEI转染至悬浮培养的293E细胞,且重链和轻链的重组表达载体共转比例为1:1。培养5~6天后,收集瞬时表达培养上清液,利用Fc捕获法通过Fortebio检测抗体的表达量,通过Protein A亲和层析法纯化蛋白,使用微量热差式扫描量热仪MicroCal PEAQ-DSC测量Tm值。结果如表4所示,1)组合P0和组合P1表达水平无明显区别,表明连接肽不影响抗体的表达;2)在抗体VH/CH1之间和/或VL/CL之间插入连接肽,尤其是在VH/CH1之间插入连接肽GG,能显著提高突变体的表达量;3)点突变不影响Fab的热稳定性,插入连接肽仅会影响Fab和CH3的熔解峰耦合,但不会影响Fab的热稳定性。
表4VH/VL和CH1/CL突变组合对表达水平、热稳定性的影响
注:*Fab和CH3的熔解峰耦合
2.3正交实验
在实施例2的表3中,在抗体VH/CH1之间引入连接肽“GG”的突变体均能正常表达,表明轻重链能正确配对。因此进一步选取组合P2、组合P6和组合P12、组合P16的轻重链进行正交实验。具体来说,组合P2、组合P6的重链通过突变带有正电荷,轻链带有负电荷,相反的组合P12、组合P16的重链通过突变带有负电荷,轻链带有正电荷,因此,理论上组合P2、组合P6的轻链和组合P12、组合P16的重链之间存在相同电荷的排斥不能配对,组合12、组合P16的轻链和组合P2、组合P6的重链之间也存在相同电荷的排斥不能配对。
如表5所示,将P2、P6的重链A命名为重链A
表5
实施例3.EGFR×HER2双特异性抗体的制备
3.1轻重链配对实验
为了构建EGFR×HER2双特异性抗体,随机选择实施例2的表4中抗EGFR抗体组合P2、P3、P5,抗HER2抗体组合T4进行轻重链配对实验。为了验证EGFR×HER2双特异性抗体的轻重链是否完全正确配对,本实施例如表7所示将EGFR抗体和HER2抗体的轻链和EGFR抗体的重链共转染,观察HER2抗体的轻链是否会干扰EGFR抗体的正确配对。另外将EGFR抗体和HER2抗体的轻链和HER2抗体的重链共转染,观察EGFR抗体的轻链是否会干扰HER2抗体的正确配对。表达载体用PEI转染至悬浮培养的293E细胞,转染比例为A:a:b=1:1:1。培养5~6天后,收集瞬时表达培养上清液,通过Protein A亲和层析法纯化蛋白。利用还原毛细管凝胶电泳分析两条轻链的比例。结果如表7所示,HER2抗体T4的轻链能完全和T4的重链正确配对,并且不受EGFR抗体P2、P3、P5轻链的干扰。EGFR抗体P2、P3、P5轻链能和EGFR抗体P2、P3、P5重链正确配对,其中,EGFR抗体P2受影响最小,当HER2抗体轻链和P2抗体轻链等比例转染时,P2抗体的配对完全不受干扰。
表7.EGFR×HER2双特异性抗体轻重链配对实验
3.2.EGFR×HER2双特异性抗体分子构建,表达和纯化
选择EGFR抗体P2和HER2抗体T4构建EGFR×HER2双特异性抗体。在P2的重链CH3结构域引入S364R+E357S+Y349C+I253N点突变,在T4的重链CH3结构域引入F405E+K409F+K370D+S354C点突变,促使P2和T4的重链优先形成异源二聚体,如表8所示。表达载体用PEI转染至悬浮培养的293E细胞,转染比例为重链A:轻链a:重链B:轻链b=3:3:1:1,培养5~6天后,收集瞬时表达培养上清液,通过Protein A亲和层析法纯化蛋白。LC-MS分析结果显示,未检测到轻重链错配和重链同源二聚体,双特异性抗体纯度约100%。
表8.EGFR×HER2双特异性抗体分子构建方法
3.3.ELISA法检测EGFR×HER2双特异性抗体结合抗原活性
将重组EGFR-ECD-Fc蛋白用包被液稀释至3μg/ml,以50μl/孔加入酶标板,4℃过夜。PBST洗板3次,加入200μl/孔封闭液,37℃放置1小时后PBST洗板1次待用。用稀释液稀释抗EGFR×HER2双特异性抗体和对照抗体(EGFR抗体P2)至100μg/ml,4倍比稀释形成12个浓度梯度(最高浓度为100000ng/ml,最低浓度为0.02ng/ml),依次加入封闭后的酶标板,100μl/孔,37℃放置1小时。PBST洗板3次,加入HRP标记的鼠抗人Fab抗体,37℃放置30分钟。PBST洗板3次后,在吸水纸上尽量拍干残留液滴,每孔加入100μl的TMB,室温(20±5℃)避光放置5分钟;每孔加入终止液终止底物反应,酶标仪450nm处读取OD值,GraphPad Prism6进行数据分析,作图并计算EC
为了检测抗EGFR×HER2双特异性抗体与HER2的结合能力,将重组HER2-ECD-Fc蛋白用包被液稀释至0.4μg/ml,50μl/孔加入酶标版,4℃过夜。PBST洗板3次,加入200μl/孔封闭液,37℃放置1小时后PBST洗板1次待用。用稀释液稀释抗EGFR×HER2双特异性抗体和对照抗体(HER2抗体T4)至100μg/ml,4倍比稀释形成12个浓度梯度(最高浓度为100000ng/ml,最低浓度为0.02ng/ml),依次加入封闭后的酶标板,100μl/孔,37℃放置1小时。PBST洗板3次,加入HRP标记的鼠抗人Fab抗体,37℃放置30分钟。PBST洗板3次后,在吸水纸上尽量拍干残留液滴,每孔加入100μl的TMB,室温(20±5℃)避光放置5分钟;每孔加入50μl 2M H
为了检测抗EGFR×HER2双特异性抗体同时结合HER2和EGFR的能力,将重组HER2-ECD-Fc蛋白用包被液稀释至0.4μg/ml,50μl/孔加入酶标版,4℃过夜。PBST洗板3次,加入200μl/孔封闭液,37℃放置1小时后PBST洗板1次待用。用稀释液稀释抗EGFR×HER2双特异性抗体至100μg/ml,4倍比稀释形成12个浓度梯度(最高浓度为100000ng/ml,最低浓度为0.02ng/ml),依次加入封闭后的酶标板,100μl/孔,37℃放置1小时。PBST洗板3次,按150ng/孔加入EGFR-ECD-Fc-biotin,37℃放置1小时。PBST洗板3次后加入HRP标记的Streptavidin,37℃放置30分钟。PBST洗板3次后,在吸水纸上尽量拍干残留液滴,每孔加入100μl的TMB,室温(20±5℃)避光放置5分钟;每孔加入50μl 2M H
4.1轻重链配对实验
为了构建EGFR×cMet双特异性抗体,随机选择实施例2中抗EGFR抗体组合P2、P3,抗cMet抗体组合O2、O3进行筛选。为了验证EGFR×cMet双特异性抗体的轻重链是否完全正确配对,本实施例如表9所示将EGFR抗体和cMet抗体的轻链和EGFR抗体的重链共转染,观察cMet抗体的轻链是否会干扰EGFR抗体的正确配对。另外将EGFR抗体和cMet抗体的轻链和cMet抗体的重链共转染,观察EGFR抗体的轻链是否会干扰cMet抗体的正确配对。表达载体用PEI转染至悬浮培养的293E细胞,转染比例为A:a:b=1:1:1。培养5~6天后,收集瞬时表达培养上清液,通过Protein A亲和层析法纯化蛋白。利用还原毛细管凝胶电泳分析两条轻链的比例。结果如表9所示,cMet抗体O2、O3的轻链能完全和O2、O3的重链正确配对,并且不受EGFR抗体P2、P3轻链表达的干扰。EGFR抗体P2、P3轻链能和EGFR抗体P2、P3重链正确配对,其中,EGFR抗体P2受影响最小,当cMet抗体轻链和P2抗体轻链等比例转染时,P2抗体的配对完全不受干扰。
表9.EGFR×cMet双特异性抗体轻重链配对实验
4.2.EGFR×cMet双特异性抗体分子构建,表达和纯化
选择EGFR抗体P2和cMet抗体O2构建EGFR×cMet双特异性抗体。在P2的重链CH3结构域引入S364R+E357S+Y349C+I253N点突变,在O2的重链CH3结构域引入F405E+K409F+K370D+S354C点突变,促使P2和O2的重链优先形成异源二聚体,如表10所示。表达载体用PEI转染至悬浮培养的293E细胞,转染比例为重链A:轻链a:重链B:轻链b=2:2:1:1,培养5~6天后,收集瞬时表达培养上清液,通过Protein A亲和层析法纯化蛋白。LC-MS分析结果显示,未检测到轻重链错配和重链同源二聚体,双特异性抗体纯度约100%。
表10.EGFR×cMet双特异性抗体分子构建方法
4.3.ELISA法检测EGFR×cMet双特异性抗体结合抗原活性
将重组EGFR-ECD-Fc蛋白用包被液稀释至3μg/ml,以50μl/孔加入酶标板,4℃过夜。PBST洗板3次,加入200μl/孔封闭液,37℃放置1小时后PBST洗板1次待用。用稀释液稀释抗EGFR×cMet双特异性抗体和对照抗体(EGFR抗体P2)至100μg/ml,4倍比稀释形成12个浓度梯度(最高浓度为100000ng/ml,最低浓度为0.02ng/ml),依次加入封闭后的酶标板,100μl/孔,37℃放置1小时。PBST洗板3次,加入HRP标记的鼠抗人Fab抗体,37℃放置30分钟。PBST洗板3次后,在吸水纸上尽量拍干残留液滴,每孔加入100μl的TMB,室温(20±5℃)避光放置5分钟;每孔加入终止液终止底物反应,酶标仪450nm处读取OD值,GraphPad Prism6进行数据分析,作图并计算EC
为了检测抗EGFR×cMet双特异性抗体与cMet的结合能力,将重组cMet-ECD-Fc蛋白用包被液稀释至0.4μg/ml,50μl/孔加入酶标版,4℃过夜。PBST洗板3次,加入200μl/孔封闭液,37℃放置1小时后PBST洗板1次待用。用稀释液稀释抗EGFR×cMet双特异性抗体和对照抗体(cMet抗体O2)至100μg/ml,4倍比稀释形成12个浓度梯度(最高浓度为100000ng/ml,最低浓度为0.02ng/ml),依次加入封闭后的酶标板,100μl/孔,37℃放置1小时。PBST洗板3次,加入HRP标记的鼠抗人Fab抗体,37℃放置30分钟。PBST洗板3次后,在吸水纸上尽量拍干残留液滴,每孔加入100μl的TMB,室温(20±5℃)避光放置5分钟;每孔加入50μl 2M H
为了检测抗EGFR×cMet双特异性抗体同时结合cMet和EGFR的能力,将重组cMet-ECD-Fc蛋白用包被液稀释至0.4μg/ml,50μl/孔加入酶标版,4℃过夜。PBST洗板3次,加入200μl/孔封闭液,37℃放置1小时后PBST洗板1次待用。用稀释液稀释抗EGFR×cMet双特异性抗体至100μg/ml,4倍比稀释形成12个浓度梯度(最高浓度为100000ng/ml,最低浓度为0.02ng/ml),依次加入封闭后的酶标板,100μl/孔,37℃放置1小时。PBST洗板3次,按150ng/孔加入EGFR-ECD-Fc-biotin,37℃放置1小时。PBST洗板3次后加入HRP标记的Streptavidin,37℃放置30分钟。PBST洗板3次后,在吸水纸上尽量拍干残留液滴,每孔加入100μl的TMB,室温(20±5℃)避光放置5分钟;每孔加入50μl 2M H
以上对本发明的具体实施例进行了详细描述,但其只是作为范例,本发明并不限制于以上描述的具体实施例。对于本领域技术人员而言,任何对本发明进行的等同修改和替代也都在本发明的范畴之中。因此,在不脱离本发明的精神和范围下所作的均等变换和修改,都应涵盖在本发明的范围内。
序列表
<110> 周易
<120> Fab改造诱导形成的双特异性抗体及其制备方法和用途
<160> 10
<170> SIPOSequenceListing 1.0
<210> 1
<211> 450
<212> PRT
<213> Artificial
<400> 1
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 2
<211> 214
<212> PRT
<213> Artificial
<400> 2
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 3
<211> 449
<212> PRT
<213> Artificial
<400> 3
Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Asn Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Ser Gly Gly Asn Thr Asp Tyr Asn Thr Pro Phe Thr
50 55 60
Ser Arg Leu Ser Ile Asn Lys Asp Asn Ser Lys Ser Gln Val Phe Phe
65 70 75 80
Lys Met Asn Ser Leu Gln Ser Asn Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95
Arg Ala Leu Thr Tyr Tyr Asp Tyr Glu Phe Ala Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 4
<211> 214
<212> PRT
<213> Artificial
<400> 4
Asp Ile Leu Leu Thr Gln Ser Pro Val Ile Leu Ser Val Ser Pro Gly
1 5 10 15
Glu Arg Val Ser Phe Ser Cys Arg Ala Ser Gln Ser Ile Gly Thr Asn
20 25 30
Ile His Trp Tyr Gln Gln Arg Thr Asn Gly Ser Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Glu Ser Ile Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Ser Ile Asn Ser Val Glu Ser
65 70 75 80
Glu Asp Ile Ala Asp Tyr Tyr Cys Gln Gln Asn Asn Asn Trp Pro Thr
85 90 95
Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 5
<211> 445
<212> PRT
<213> Artificial
<400> 5
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Asp Met Ser Trp Gly Arg Gln Ala Pro Gly Lys Arg Leu Glu Trp Val
35 40 45
Ala Phe Met Ser Ser Gly Gly Ser Thr Tyr Tyr Pro Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Asp Arg Phe Gly Ser Tyr Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro
115 120 125
Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val
130 135 140
Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala
145 150 155 160
Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly
165 170 175
Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly
180 185 190
Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys
195 200 205
Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys
210 215 220
Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu
225 230 235 240
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
245 250 255
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
260 265 270
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
275 280 285
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
290 295 300
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
305 310 315 320
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
325 330 335
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser
340 345 350
Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys
355 360 365
Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
370 375 380
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly
385 390 395 400
Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
405 410 415
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
420 425 430
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 6
<211> 214
<212> PRT
<213> Artificial
<400> 6
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Asp His Ile Asn Asn Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Ser Gly Ala Thr Ser Leu Glu Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Phe Trp Ser Thr Pro Phe
85 90 95
Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Arg
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 7
<211> 448
<212> PRT
<213> Artificial
<400> 7
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Leu His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Met Ile Asp Pro Ser Asn Ser Asp Thr Arg Phe Asn Pro Asn Phe
50 55 60
Lys Asp Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Tyr Arg Ser Tyr Val Thr Pro Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 11
<211> 220
<212> PRT
<213> Artificial
<400> 11
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ser Ser Gln Ser Leu Leu Tyr Thr
20 25 30
Ser Ser Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys
35 40 45
Ala Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Ala Tyr Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile
100 105 110
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
115 120 125
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
130 135 140
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
145 150 155 160
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
165 170 175
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
180 185 190
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
195 200 205
Ser Pro Val Thr Lys Arg Phe Asn Arg Gly Glu Cys
210 215 220
<210> 9
<211> 449
<212> PRT
<213> Artificial
<400> 9
Gln Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Gly Ser Val Ser Ser Gly
20 25 30
Asp Tyr Tyr Trp Thr Trp Ile Arg Gln Ser Pro Gly Lys Gly Leu Glu
35 40 45
Trp Ile Gly His Ile Tyr Tyr Ser Gly Asn Thr Asn Tyr Asn Pro Ser
50 55 60
Leu Lys Ser Arg Leu Thr Ile Ser Ile Asp Thr Ser Lys Thr Gln Phe
65 70 75 80
Ser Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Ile Tyr Tyr
85 90 95
Cys Val Arg Asp Arg Val Thr Gly Ala Phe Asp Ile Trp Gly Gln Gly
100 105 110
Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 10
<211> 214
<212> PRT
<213> Artificial
<400> 10
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Leu Glu Thr Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Phe Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Ile Ala Thr Tyr Phe Cys Gln His Phe Asp His Leu Pro Leu
85 90 95
Ala Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Arg
195 200 205
Phe Asn Arg Gly Glu Cys
210
- Fab改造诱导形成的双特异性抗体及其制备方法和用途
- 高效诱导抗体重链恒定区的异源二聚体形成的CH3域变体对,其制备方法及用途