一种李树和多花黄精的套栽方法
文献发布时间:2024-04-18 19:58:21
技术领域
本发明涉及中药材种植技术领域,特别是一种李树和多花黄精的套栽方法,属于现代化农业技术和中药材高品质培育技术融合创新领域,属于种植基础技术中的现代化科学分析技术相结合的研究创新成果。
背景技术
李树,又名李子树,是蔷薇科李属木本植物,是中国李的古老良种。喜光,抗寒,适应性强,以温暖湿润的气候环境和排水良好的砂质壤土生长为佳。李树小枝无毛,叶片为矩圆状倒卵形或椭圆状倒卵形,李树枝叶生长较为致密,林下光照度适中。
黄精,为百合科植物滇黄精、黄精或多花黄精的燥根茎。按形状不同,民间习称“鸡头黄精”、“姜形黄精”等,黄精具有补气养阴,健脾,润肺,益肾之功效。其中,又以多花黄精最为著名,多花黄精产于我国四川、贵州、湖南、湖北、安徽、浙江、福建等地,生林下、灌丛或山坡阴面或海拔500-2100米。具有喜阴、耐寒、怕干旱的特性,在干燥地区生长不良,在湿润荫蔽的环境下植株生长良好。在土层较深厚、疏松肥沃、排水和保水性能较好的壤土中生长良好;在贫瘠干旱及粘重的地块不适宜植株生长。
现有技术没有将李树和多花黄精进行套栽的种植方法,也缺少相关研究,经过检索查询发现部分黄精和其他植物套种的案例,但大多以促进林木生长为目的,尚未见通过黄精与其他作物套种改良种植品质的案例。
例如中国发明专利申请CN 105918047 A公开一种山核桃与黄精套种的方法,涉及山区植物种植技术领域,包括栽植核桃树、撒施栽植肥、栽植黄精、撒施肥料、种植管理。本发明方法合理、操作方便、实施简单。该专利主要是针对特定林木品质(金寨县大别山山核桃,Carya Dabieshanensis)在大别山区广泛分布,在安徽金寨县天堂寨、关庙、吴家店及燕子河等十几个乡镇海拔500m以上的阴坡和半阴坡,以600~800m较集中。为了增加大别山地区的农业产值,提出了山核桃与黄精套种的方法,种植效果主要是实现山核桃和黄精的共同种植生长目的。
另外一些专利针对黄精市场需求量不断扩大,价格逐年上涨,供不应求的问题,提出人工栽培黄精的技术,重点解决人工大量挖掘野生黄精,造成生态环境恶化,水土大量流失的问题。如中国发明专利申请CN 114271167 A公开的一种黄精人工种植方法,该专利主要针对黄精的培育问题,属于黄精种植的基础培育技术研究。又如如中国发明专利申请CN112189522 A公开一种林下仿生栽培丹参、黄精和芍药的种植方法,提出一种完善的利用林地原生资源进行丹参和芍药栽培的技术方案,提高土地使用率,可改善生态环境。
中国发明专利申请CN 108040724 A公开了一种梨树下套种黄精的种植方法,涉及种植技术领域,本发明包括选地整松土,梨树移栽、黄精的处理及栽植以及除草施肥等步骤,采用本发明提供的方法能够避免了黄精遭到曝晒,提高了黄精的成活率,从而提高了黄精的产量;梨树为黄精提供适宜的生长环境,同时黄精生长抑制杂草生长,有效的减少了除草用药,提高梨树和黄精的产量。
袁应菊的中国硕士学位论文《不同复合模式对黄精生长特性及林地土壤酶活性的影响研究》公开了将黄精和多种植物进行套种套栽的园艺,眼界结果表明在林下复合种植中草药,可以为中草药提供仿野生的生长环境,提升中草药品质,并改善林地土壤的酶活性。该研究重点关注了林下种植过程中,林内小环境、光照强度、枯落物量等的不同,导致林下中草药的生长量和品质发生变化。该论文重点比较了乔木+草本(C+P)、乔木+灌木+草本(C+E+P)、灌木+草本(E+P)的林下生态因子、中草药黄精的光合生理特性、根茎生长量、可溶性糖含量及黄精林地土壤酶的特性影响作用。研究比较了不同复合模式下种植中草药,由于林内小环境、光照强度、枯落物量等的不同,导致林下中草药的生长量、品质及林地土壤酶的活性不同。对比3种典型的林药复合模式:乔木+草本(C+P)、乔木+灌木+草本(C+E+P)、灌木+草本(E+P),比较不同套栽方法,林下生态因子、中草药黄精的光合生理特性、根茎生长量、可溶性糖含量及黄精林地土壤酶的特性,最终研究结果显示:乔木+灌木+草本套栽的方法中药材栽种效果最佳。然而,该研究主要还是在于林下中草药生长过程中光合生理环境和特性的影响,仅在一定程度改善生长速度、中药材产量的影响,并不能够改变套栽中药材的品质,不具有选择性。
如果能够更进一步的优化种植方式方法,提高黄精套栽协同效果,可以在现有提升产量的基础上,实现黄精品质进一步增强,更好的发挥黄精在特定产品的应用价值。
发明内容
本发明的目的是针对现有技术对于植物套种研究较少,特别是对于多花黄精大量种植以后,选择适宜耕地较为困难的问题,虽有一些关于黄精和其他作物套栽的园艺技术,但主要是提升产量,并未实现黄精品质的特异性选择优化改善,为此本发明提供一种李树和多花黄精的套栽方法。在种植李树的地里种植喜阴的多花黄精药材,增大了土地的利用率,同时实现多花黄精和李树种植过程中的相互协同促进,改善多花黄精的种植品质,特别是针对性提高黄精根茎某些特别的多糖成分含量,提升特定方面应用价值。最后,采用本发明的方法可以实现新农村建设过程中,中药材和传统果树园林的高效率高价值套栽,经济效益成倍增加。
为了实现上述发明目的,本发明采用以下技术方案:
一种李树和多花黄精的套栽方法,包括以下步骤:
S1、挖定植穴,在底部铺上一层基肥,然后将李子树放入定植穴,回填土壤将李树苗埋入。
其中,每一行内相邻定植穴的间隔为5-6米,定植穴的行间距为7-9米。
如果是已有李树苗圃或园林,满足上述李树栽种间隔的,可以在栽有李树的苗圃或园林基础上作畦开沟,调整李树种植状态,达到上述要求的作畦开沟规范要求。
S2、整地、除草、浇水,维护李树生长至2-4米高,树冠宽度达到2-5米。优选,维护李树生长至3米高,树冠宽度达到4米,即李树的高度和宽度可以大于前述控制数值。或者,优选培育李树苗圃/园林,直至李树高度≥3米,树冠宽度≥4米。
S3、春季3-4月份,在相邻李树之间,作畦开沟,畦宽1.1-1.5米,畦长5-9米,畦高30-60厘米,相邻畦间横沟的宽度为0.2-0.4米,沟深为低于李树根部和地面接触位置的高度30-40厘米;横沟距离李树的距离至少0.4米。
将黄精根状茎芽头朝上放置在沟内,横竖间隔25-40厘米放一段;然后覆盖厚度6-10厘米的泥土(优选作畦开沟时挖出的细土),压实,浇透水。例如,根据李树行间距为6米,按照李树行间距作畦开沟,设置畦宽1.2米,畦长7米,横沟宽0.3米,沟深0.3米;在相邻行的李树之间开3条沟,相邻沟间距1.2米,沟宽0.3米,沟深0.3米。
S4、每隔1-15天对种植地进行一次除草;除草完成后,浇透水,使得土壤湿润。
黄精追肥:等黄精地上部分出苗高度达到40-60厘米时,对黄精追肥,每亩施加80-800公斤的农家肥,共计施肥1-2次;每年施农家肥总计1000-1080公斤。
李树追肥:每年在3月、李树开花后、5月、10月,对前述生长期李子树各追施1次;对李树追肥,前述各次追肥为每亩分别施加30-60公斤、30-60公斤、40-80公斤、500-800公斤农家肥,共计施肥4次。优选的,前述四次追肥分别为40公斤、40公斤、50公斤、500-800公斤的农家肥,每次追肥用量可以在±30%范围内变化,优选±20%范围内变化,优选10%范围内变化。
S5、修剪处理:
在每年在11月-次年1月,对李子树修剪,长势较差的枝条剪去,提高坐果率。
本发明李树和多花黄精的套栽方法优化了李树和黄精套作过程中的水肥管理和病虫害防治,提高栽种效率,减少病虫害。将黄精和李树栽种的作畦开沟的宽度、间隔距离和高度进行综合优化,使得黄精的根系和李树的根系能够在土壤中产生相互作用,实现了两者生长促进作用,使得关键酶的含量发生一定的调整变化,黄精生长品质更好,能够获得更具有药用价值的优质黄精。即黄精和李树根茎相互协同促进,使得黄精根茎中关键酶含量显著提高,能够栽种得到品质更加优秀的黄精,相关根本原理还可以进一步深入研究,提升套栽园艺技术的效率。
而且,每次采挖黄精时相当于翻土一次,有利于雨水浸入土地,为李树植株和下一轮的黄精植株提供水分。种植过程更加节省人力,且黄精和李树的产量都较高,也提高了土壤的利用率,节省了土地资源,经济效益成倍增加。最为重要的是收获的黄精品质更好,其中富含的黄精多糖更具有选择性,能够更好的发挥保健养生开发价值。
本发明黄精和李树套栽方法并不严格限定各个步骤的顺序,本领域技术人员可以根据作物生长情况灵活调整各个步骤的实施时机,当李树生长达到要求以后,即可开始套栽黄精。除非植物生长情况限制,不应认为本发明描述的李树和黄精套栽方法严格限定了各个步骤的先后顺序。
进一步,所述李树是茵红李、杏李、毛梗李、南华李、潘园李、红美人李、白美人李、迟蜜李、醉李。
进一步,所述黄精是滇黄精、黄精或多花黄精。优选的,所述黄精是多花黄精。
进一步,所述基肥是农家肥。优选的,所述基肥是腐熟农家肥。
优选的,所述农家肥包含以下重量份原料经过7-30天腐熟而成:土壤100份、粪便5-12份、木屑10-30份、氮肥0.01-0.1份、磷肥0.01-0.1份,其中动物粪便以干重计算。氮肥和磷肥以氮元素、磷元素的重量计算。
农家肥腐熟过程中,利用机械或人工将土壤破碎,与粪便、木屑混合均匀,进行腐熟处理,然后撒入氮肥和磷肥,搅拌混合均匀,即得经过腐熟的农家肥。也可以将氮肥、磷肥和其他原料直接混合均匀,然后再进行腐熟处理。腐熟过程为堆肥腐熟,注意监控农家肥堆肥腐熟的温度,如果堆肥温度超过60℃,则需要翻堆降低堆肥的温度。
优选的,每一个定植穴铺基肥的用量为0.56-0.80公斤。
进一步,步骤S3中,在李树种植2-8年后,种植黄精。李树种植两年以后,植株长势较好,植株茂盛,阳光不能照射进地里,多花黄精为喜阴植物,李树植株的间隙处可以种植多花黄精。根据李树生长情况确定是否可以种植黄精,如果李树生长速度较为缓慢,或者原始移栽李树苗较小的,则需要多培育一段时间,确保李树达到足够高度。优选的,李树生长高度≥3米,树冠宽度≥4米,安排黄精种植。
进一步,步骤S3中,选择根状茎直径4-6厘米的黄精,切割长度3-5厘米的截断,确保每段上有2-3芽节;将黄精截断;然后将截断捞起,在截断切口处涂抹草木灰,直至伤口全部被覆盖,阴凉处晾干,备用;切段备用的黄精根状茎
进一步,在黄精种下之后的年份,6-7月采摘李子果以后,修剪树枝,提高树下采光率。李子的采摘季节为6-7月,黄精的种植季节为9-10月,多花黄精为多年生植物,在冬末春初采摘较好。
优选的,黄精种植3-5年后,在9-10月收割黄精。
进一步,收割黄精后,间隔7-9个月时间后进行土壤肥力养护。待第2年后,再次栽种黄精。可以根据黄精种植情况,选择性进行休耕,养护地力,提升黄精种植效果。休耕间隔为1年、2年、3年、4年或5年,根据土地地力适当调整休耕间隔时间。
进一步,步骤S5中,修剪后的枝条、枝叶就地处理,剪断回土增肥。
采用上述发明方案,可以取得的有益效果如下:
1、本发明李树和多花黄精的套栽方法,通过优化两种植株套作过程中的水肥管理和病虫害防治,提高栽种效率,减少病虫害。对于黄精栽种过程中作畦开沟的宽度、间隔距离和高度进行综合优化,特别是挖沟深度控制水分扩散流动,结合已经生长中的李树根部的土壤高度作为基准,控制沟深度和李树根系生长的深度影响。利用沟深度调整水肥,使得李树根系较深,较发散。然后,对于黄精栽种的时候,深度控制到6-10里面,并进行压实,使得黄精截断成活以后,根系生长的深度适宜,能够和李树根系在土壤中相互作用,实现了两者生长促进作用,使得关键酶的含量发生一定的调整变化,进而实现黄精品质的选择性控制。
2、本发明李树和多花黄精的套栽方法,根据种植的深度和作畦开沟的深度,控制两种作物的根系发展,实现了李树和黄精的协同生长促进作用,黄精根茎生长过程中,关键酶基因sacA(β-果糖苷酶)、PMM(磷酸甘露糖异构酶)、SPS(蔗糖磷酸合成酶)的相对表达量更高,表明黄精多糖的品质因为套种的缘故有提升,具有更高的药用价值,品质得到提升。
3、本发明李树和多花黄精的套栽方法,当栽种多花黄精的时候,每次采挖黄精时相当于翻土一次,有利于雨水浸入土地,为李树植株和下一轮的黄精植株提供水分。因此,套栽种植更加节省人力,且黄精和李树的产量都较高,也提高了土壤的利用率,节省了土地资源,经济效益显著增加。
4、本发明李树和黄精套栽方法还可以在修剪李树枝条的时候,调整黄精枝叶和李树枝叶的相互作用,使得黄精的生长与李树遮荫作用平衡协同,利于黄精的产量和品质的控制。
附图说明:
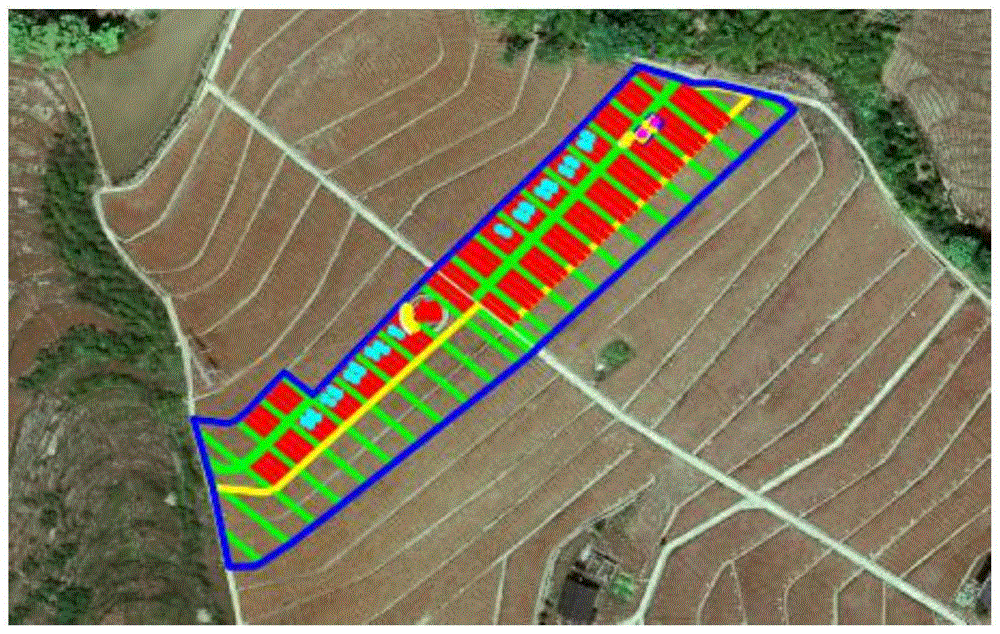
图1是本发明实施例2李树和多花黄精的套栽的场地航拍照片。
图2是图1中标记的李树和多花黄精的套栽规划区块划分图。
图3是种植李树苗圃管理后,李树生长至3米高在相邻李树之间,作畦开沟的照片。其中,标牌为准备套栽黄精的苗圃。
图4是黄精幼苗套栽在李树林间的照片。
图5是李树下套栽黄精幼苗长高达到45厘米以上的照片。
图6是黄精单种的照片,采用泡沫箱作为栽培容器。
图7是黄精和李树套栽第3年的照片,黄精高度达到48厘米。
图中标记:11-黄精和李树套栽区域B、12-黄精和李树套在区域A、21-黄精单种区域B、22-黄精单种区域A。
具体实施方式
为了更好的说明本发明方案,以下结合具体实施例以及相关测试例进行说明,应当理解下列实施例只是本发明的较优实施方案,并非对于本发明技术方案的限定。
实施例1
一种李树和多花黄精的套栽方法,具体套栽园艺如下:
种植李树幼苗植株:选择适合当地抗病性强且产量高的李子树品种,挖定植穴,往定植穴底部铺上一层基肥,将李子树苗埋入其中,回填土壤固定李子树,确保李子树全部根系埋入土壤中。在李子树种植好后,浇透水。
李子树管理:每年李子树生长期要追施2-3次的有机肥,确保李子树的生长健康状态,特别是李子树根系和植株的高度、树冠宽度达到良好状态。同时,在李子树管理过程中,还需要注意修剪处理,把李子树长势较差的枝条剪去,提高坐果率。
栽种多花黄精:在李树植株种植两年后,李树植株长势较好,植株茂盛,阳光不能照射进地里,多花黄精为喜阴植物,李树植株的间隙处种植多花黄精。
李子采摘和黄精收割:李子的采摘季节为6-7月,黄精的种植季节为9-10月,多花黄精为多年生植物,在冬末春初采摘较好。
本实施例概述本发明李树和多花黄精的套栽方法,本发明将李子树的栽种管理和多花黄精的栽种管理有机结合,利用李子树提供阴凉环境,同时利用多花黄精栽种加强李子树林下管理,抑制杂草生长,两者的生长相辅相成。从农艺栽种的角度提供了互相配合协同的基础栽种条件,有利于两种作物的生长状态改善提升。
实施例2
为了更进一步详细说明李树和多花黄精的套栽方法,以下结合2016-2023年在某苗圃进行黄精和李子树套栽的园艺管理情况,进行详细李树和多花黄精套栽、管理、养护和收割工作说明。李树和黄精套栽种植地区的航拍照片如图1所示,栽种场地环境:气候较为阴湿的半高山,种植区域土层湿润肥沃、且深厚疏松,整体环境肥力良好。栽种场地划分如图2所示:划分成黄精和李树套栽区域A和B,对应图2中标记12和11区域,相应的黄精记为黄精L(林下黄精);黄精单种区域A和B,对应图2中标记22和21区域,相应的黄精记为黄精CK(黄精单作)。具体而言,林下黄精为间隔在李树之间区域种植;黄精单作为图中分为多个标记的泡沫箱子中分布。
黄精和李树套栽区域A和B(标记12和11区域)的种植方法如下。
农家肥准备:农家肥可以用其他同等肥料代替,本发明使用以下腐熟农家肥原料。农家肥重量份原料组成如下:土壤100份、粪便10份、木屑20份、氮肥0.05份、磷肥0.05份,其中动物粪便以干重计算,氮肥和磷肥以氮元素、磷元素的重量计算。
使用机械或人工将土壤破碎,加入粪便、木屑,翻料混合均匀。堆积成0.8米至1.6米高的物料堆,进行腐熟处理,经过20天腐熟而成。腐熟完成后,撒入氮肥和磷肥,搅拌混合均匀,即得经过腐熟的农家肥。腐熟过程为堆肥腐熟,注意监控农家肥堆肥腐熟的温度,如果堆肥温度超过60℃,则需要翻堆降低堆肥的温度。
腐熟完成后,得到的农家肥含水率控制在45%-65%之间,如果堆肥含水率过低,则需要人工洒水增湿;如果与降雨过多,导致堆肥含水率过高,则需要注意防雨,并增加翻堆次数,以减少腐熟堆肥农家肥的含水率。
以上腐熟农家肥仅仅是本发明具体实施方案中的一个具体案例,本领域技术人员还可以采用其他方式进行农家肥堆肥腐熟处理,在确保农家肥肥力适宜的情况下,采用其他方法准备农家肥并不影响李子树和黄精的种植效果。
李子树种植:挖定植穴,每一行内相邻定植穴的间隔为6米,定植穴的行间距为8米。在底部铺上约0.7公斤农家肥作为基肥。将高度80-120厘米的茵红李树幼苗放入定植穴,回填土壤将李树苗根系充分埋入土壤,并固定李子树呈竖直状。以上茵红李还可以选择其他品种的李树,定植穴的间距控制在上述适宜间隔距离。更具体的本实施例李树的种植选择现有种植好的李树果园,李树为16年种植的李树,李树栽种区域如图2所示,图中标记11和12的区域种植茵红李,李树高度及树冠宽度符合规划种植黄精的条件。如果是新建李树果园则应当注意定植穴间隔距离,栽种李树管理培育达到设计高度和树冠宽度(树高≥3米高,树冠宽度≥4米),再进行黄精套栽工作。
李子树苗圃管理:整地、除草、浇水,维护李树生长至3米高,树冠宽度达到4米。李树种植两年以后,植株长势较好,植株茂盛,阳光不能照射进地里,满足种植多花黄精的条件。本实施例种植李子树生长态势良好,如果土壤肥力较差,李子树更为缓慢,则需增加李子树培养时间,以确保李树达到前述生长状态(高度≥3米,树冠宽度≥4米)。
开挖黄精种植沟:李子树达到前述高度,树冠宽度达到前述宽度,次年春季3-4月份,在相邻李树之间,作畦开沟,畦宽1.2米,畦长5-6米(因为土地缘故,李树行距略有波动变化),畦高35厘米;即,相邻李树之间按照行距1.5米开横沟,沟宽0.3米,沟深0.3米,相邻沟之间间距1.2米,沟深为低于李树根部地面高度约35厘米,如图3所示。需要注意沟深是以李树根茎和里面接触的部分作为基础点,水平高度下挖0.35米,不宜对李树根部进行堆土处理,尽量保持李树根部土壤平整。沟深度影响李树根系发育,控制黄精栽种以后生根的空间大小,以及两者的相互影响。
准备黄精种植根状茎:选择根状茎直径约5厘米的多花黄精,切割成长度3-5厘米的截断,确保每段上有2-3芽节;黄精截断后,在截断切口处涂抹草木灰,将黄精伤口全部被覆盖草木灰,置于阴凉处晾干,得到切段黄精根状茎,备用。
黄精初次种植:将前述准备的黄精根状茎芽头朝上放置在挖好的沟内,每间隔30-30厘米放一段(横向、纵向各间隔30厘米);然后将作畦开沟时挖出的细土回填黄精栽种沟,覆盖厚度6-10厘米(平均约8厘米),压实,浇透水,记为林间黄精L或黄精L。多花黄精的栽种深度约8厘米左右,当黄精成活以后,生长的根系向下发展,和李树是根系存在较大的作用间隔,两者相互作用更为理想。黄精种下以后,注意压实土壤,避免黄精倒下,也保持黄精成活以后生根的位置的精确控制。
多花黄精为喜阴植物,李树植株的间隙处可以种植多花黄精,实现两者套栽,相互促进生长状态,达到高品质的栽种目的,黄精L栽种的初始状态如图4所示。
黄精种植管理:每隔10天对种植地进行一次除草;除草完成后,浇透水,使得土壤湿润。
黄精追肥:等黄精地上部分出苗高度达到45厘米时,如图5所示,对黄精追肥,每亩施加80-800公斤的农家肥,追肥施加于黄精幼苗侧面5-10厘米范围内,每年施肥2次,每年施农家肥总计约1000公斤/每亩。记录每亩栽种黄精数量,计算每棵黄精使用农家肥重量。
李树追肥:每年在3月施加40公斤农家肥,开花后施加40公斤农家肥,5月施加50公斤农家肥,10月施加600公斤农家肥,共计施肥4次,以上施用农家肥均为每亩施用农家肥的重量。
李子树采摘:每年6-7月采摘,收获李子果。在黄精种下之后的年份,采摘李子果后,还应修剪树枝,提高树下采光率。
黄精收割及复种:多花黄精为多年生植物,黄精种植3-5年后,在9-10月收割黄精。如果观察到土壤肥力下降,则改在冬末春初收割,收割黄精后,间隔7-9个月时间后进行土壤肥力养护,待次年9-10月再次栽种黄精。
李子树修剪处理:在每年在11月-次年1月,对李子树修剪,长势较差的枝条剪去,提高坐果率。修剪后的枝条、枝叶就地处理,剪断回土增肥。
以上为本发明李树和黄精套栽方法的整体概要描述,套种方法的重点在于选择黄精和李树搭配栽种,利用李树的林下阴凉环境,为黄精生长提供良好的基础环境状态。同时,黄精地下根系和李树地下根系相同互作用,相互促进,使得李树林间套栽的黄精L生长态势良好,如图7所示。
我们推测,黄精的生长状态还获益于李树根系和黄精地下根系的相互作用,以及李树修剪后的枝条、枝叶,在剪断后就地处理回土增肥,对于黄精生长具有独特的促进效果。
实施例3
为了进一步详细说明本发明李树和多花黄精套栽的方法形成的独特协同促进优势,将李树和多花黄精套种过程中获得的园艺数据进行对比分析研究。具体研究包括多花黄精套栽在李树林间的生长状态和同品种同批次多花黄精单种的生长情况的对比分析。
如实施例2记载,在准备黄精种植根状茎的时候,选择根状茎直径约5厘米的多花黄精,切割成长度3-5厘米的截断,确保每段上有2-3芽节;黄精截断后,在截断切口处涂抹草木灰,将黄精伤口全部被覆盖草木灰,置于阴凉处晾干,得到切段黄精根状茎,备用。
获得的切段黄精根状茎被分成了两组,其中一组用于实施例2记载的李树下林间套栽记为黄精L(即实施例2栽种的林下黄精L),另一组用于单独种植记作黄精CK。
单独种植的黄精,以下记为“单作黄精CK”。选择的土壤环境为:气候较为阴湿的半高山,种植区域土层湿润肥沃、且深厚疏松,整体环境肥力良好。单作黄精CK,栽种于泡沫箱中,栽种深度和实施例2中林下黄精栽种深度一致,并将泡沫箱置于图2中标记22和21区域作为黄精单种区域A和B。这些泡沫箱所处位置同样是李树下,和其实施例2栽种的黄精L布置状态相同,取得相同的遮荫效果,具体如图6所示。
由于泡沫箱储水和漏水性能相比于大地更差,每日观察泡沫箱中土壤含水率,及时补充水分,确保和泡沫箱中土壤含水率和林下黄精栽种的土壤水分一致(误差不超过20%,以林下土壤含水率为基准)。
单作黄精CK和林间黄精L种植时间相同,即实施例2李树林间种植黄精的同一时期。具体为,春季3-4月份,按行株距30cm×20cm(受限于泡沫箱大小,略有调整),开穴栽种,每穴栽1段根茎。栽后,待黄精地上部分出苗达到45厘米时,施用实施例2相同的农家肥,即实施例2记录每亩栽种黄精数量,计算得到的每棵黄精使用农家肥重量。向每棵黄精施用的农家肥和实施例2中每棵黄精施用的农家肥重量相同(根据每亩栽种黄精数量、施肥量,确定单株黄精对应农家肥重量,然后用于单作黄精CK施肥量控制)。理论上单作黄精处于泡沫箱孤立种植土壤中,不会有李树争肥的情况,即单作黄精施用农家肥量优于林下套栽黄精L。
单作黄精CK的田间管理同实施例2中李树林间种植的黄精L,出苗后经常松土除草,松土要浅,避免伤根。每年施肥2次,每年施农家肥总量与林下黄精L相同(按照每棵黄精换算)。除黄精出苗后首次追肥,其余年份在春、夏各追肥1次,肥料均为实施例2相同的农家肥。
在黄精生长的第3年,进行黄精农艺性状测量,测量过程中选择苗圃中部的黄精植株进行测量,避开苗圃边缘或接近畦边缘的黄精植株。分别测试五个样本,汇总整体农艺性状测试结果如下表所示。下表中,L为实施例2种植的李树下黄精L,对应图2中标记12和11区域;CK为单作黄精CK,对应图2中标记22和21区域。序号代表测试样本的编号。
表1李树下黄精和单作黄精的农艺性状比较结果
从农艺性状测试结果可以看出,李树下黄精L和单作黄精CK相比,李树下黄精的叶宽大幅度增加,表明多花黄精的生长态势更好,具有显著性。黄精的节间距降低19%,说明黄精根状茎的分枝倾向增加,黄精结节膨大后,药性增强,和传统黄精种植过程中所说的“鸡头黄精”说法一致,即黄精的药用价值提高。
另外,黄精茎粗略微增加,不显著,需要进一步观察测量更多样品进行分析,黄精茎粗意味着黄精的生长状态,对于黄精根茎增大可能和黄精关键基因表达有一定的关联。
最后,黄精的叶绿素和叶片数均有一定幅度的提升,具有显著性,黄精叶绿素和叶片数的增加,对于黄精生长具有正面意义。叶绿素含量受到光照、温度、植株基因等多种因素共同影响,叶绿素含量通常处于动态平衡中,可见光光照都能促使绿素形成,但如果光照过强反而会发生光氧化破坏叶绿素。保持较高的叶绿素含量,对于叶片增强光合作用是有利的,有利于提高黄精叶片的捕光能力,吸收更多光,利用光能完成光合作用。李树林间套在的多花黄精叶绿素和叶片数都有明显的增加,和李树下黄精生长速度增加的结果一致。
以上农艺性状测试结果表明,李树套栽多花黄精L相比于单作黄精CK,林下黄精L的生长态势有明显的促进,且多花黄精的产量和品质都有明显提升。
如果单纯就林下黄精L和单作黄精CK的栽种过程中的差异而言,仅仅在于单作黄精CK使用泡沫箱单独控制,在同等农家肥施用的情况下,由于单作黄精不存在李树争肥的问题,实际获得养分更多。单作黄精CK取得了同样的李树遮荫环境作用,但实际生长状态来看,单作黄精CK却不及林下黄精L,还需要进一步的研究分析。
实施例4
为了进一步详细说明本发明李树和多花黄精套栽,对于多花黄精品质和李树产量、品质的影响。对于实施例2套栽黄精以后的土壤进行采集测试,对应图2中标记12和11区域分别采集五个样品。标记21、22的区域采集样品记载H(单作黄精CK),标记11、12的区域采集样品记载L(林下黄精L),分别编号,下表中序号代表测试样本的编号。
表2土壤理化性质分析
备注:由于测试过程中出现了采样错误,引入的人为误差,故表中部分数据删除。这些误差数据和平均值相比相差200%以上,经过分析确认为取样到未均匀分散的农家肥,或未充分翻耕的林下土壤。同时,后续黄精取样的时候,避开农家肥施用不均匀区域的黄精样品,前面实施例3取样黄精样品不涉及这些需排除的黄精植株。
经过测试各个区域的林下黄精L和单作黄精CK分别取样五个土壤样品,测试结果差异并不显著,都属于正常波动范围内。即两个区域的全磷含量测试结果以及有效磷含量测试结果存在一定差异,主要是因为部分样品存在较大差异波动有关,这可能和苗圃人工管理培育过程施用肥料的分布状态有关。
另外,测得水解氮和速效钾含量存在较大波动,同样的观察到部分样品测试数据存在较大跳变,表明样品采集可能受到污染,或者向土壤施用肥料存在较大的不均匀性问题。
土壤理化性质分析结果未见明显的差异,我们认为土壤的环境因素对于林下黄精L和单作黄精CK的生长影响较小,并不是造成两者形状差异的主要原因。
实施例5
为了进一步详细研究本发明李树和多花黄精套栽,对于多花黄精品质和李树产量、品质的影响。选择作畦开沟过程中套栽在李树下的多花黄精,记作林下黄精L(同实施例2取样情况),栽种靠近畦宽边缘处的黄精,记作M,分别测试取五个样品。
黄精样品M也是实施例2栽种的黄精,只是挑选前述实验中未取样到的畦边缘黄精进行农艺性状分析。主要是比较黄精种植过程中,作畦开沟的区域位置的影响,作畦开沟的区域差异,对于水肥略有影响,但不显著。采用实施例3相同的方式进行农艺性状测试分析,比较结果如下表所示。
表3李树下黄精和靠近畦宽边缘处黄精的农艺性状比较
分析李树下套栽黄精根据栽种的位置不同,黄精农艺形状基本相同,仅仅在叶宽和节间距形状上略有差异,主要表现为林下黄精叶宽更宽一些,同时黄精节间距更短,基本上可以确定黄精的生长状态稍好。
基于上述分析,我们认为在作畦开沟的时候,黄精种植位置对于黄精的影响较小,林下黄精的种植位置对于农艺性状影响较小,不是导致黄精生长差异的主要因素。为了进一步研究黄精的套栽园艺技术,进一步开展黄精关键酶分析研究。
对套栽黄精和单作黄精进行地下部分多糖合成关键酶基因表达量测试,采用RT-qpcr方法,分析黄精根茎基因相对表达量。参考以下文献方法(陶鹏,基于转录组的黄精多糖代谢关键酶基因的筛选与验证,2023,doi:10.13422/j.cnki.syfjx.20230414)。划分为林下黄精L和单作黄精CK。林下黄精L为实施例2的李树下套栽黄精,单作黄精CK为实施例3中“单作黄精”。
黄精根茎基因表达情况分析,对比前述林下黄精L(同实施例2)和单作黄精CK(同实施例3)的根茎中关键酶基因sacA(β-果糖苷酶)、PMM(磷酸甘露糖异构酶)、SPS(蔗糖磷酸合成酶)的相对表达量变化。以及测试林下黄精L和单作黄精CK叶子中的黄精多糖含量,测试方法为蒽酮-硫酸法,结果显示。
表4黄精根茎中多糖合成关键酶基因相对表达量和多糖含量测试结果
测试结果显示,林下黄精L关键酶基因表达显著高于单作黄精CK,虽然林下黄精L和单作黄精CK都采用李树遮荫的环境进行栽种,并控制水肥环境因素一致。我们分析可能和林下黄精L对于李树是处于同一土壤环境,以及李树修剪枝条以后,回土增肥处理有关。单作黄精CK处于孤立的土壤环境(泡沫箱中),单作黄精CK的根系和李树根系相互独立;而林下黄精L的根系和李树根系存在直接作用,推测李树根系向土壤中释放化学信号使得林下黄精L的关键酶基因表达显著升高。具体作用机理还需要进一步的研究,但可以肯定黄精和李树套栽具有独特的优势,是由于李树和黄精处于相同的土壤环境,而不仅仅是李树提供树荫形成的阴凉气候环境,是其他数目和黄精套种所不具有的特性。
同时,林下黄精L和单作黄精CK相比,黄精多糖含量基本没有差别,黄精多糖含量对于培育多花黄精的产量具有正相关性。在两者黄精多糖含量差异不大的情况下,我们认为林下黄精L和单作黄精CK的产量差异不大。但是林下黄精和单作黄精相比,关键酶的表达具有显著差异,这可能会影响培育得到的黄精品质。
RT-qpcr分析确定β-果糖苷酶基因sacA表达显著增加,这对于黄精表达产生果糖具有很好的正面促进作用。中国专利CN113004433A一种黄精果聚糖及其制备方法和应用公开,使用黄精多糖经过分离提纯,可以获得以β(2→6)果糖糖苷键为主链,侧链果糖以β(2→1)糖苷键连接到主链上形成的黄精果聚糖产品。该黄精果聚糖具有促进益生菌增殖的活性,具有巨大的药物/保健品开发潜力。本发明套栽于李树林间的多花黄精L的β-果糖苷酶基因sacA表达显著上调,对于获得黄精果聚糖产物具有很好的促进意义,有利于开展黄精多糖下游产业深入研究,可以提供更多符合相关研究方向的优质黄精原料。
测试结果还显示蔗糖磷酸合成酶(Sucrose phosphate synthase,SPS)基因表达显著提高,蔗糖磷酸合酶SPS是蔗糖合成的关键限速酶,蔗糖是由葡萄糖和果糖分子组成的二糖,蔗糖磷酸合酶SPS表达升高后,黄精半乳聚糖、黄精果聚糖的合成都会增强。而磷酸甘露糖异构酶PMM基因表达显著提高,则会使得磷酸甘露糖变位酶含量增加,可催化D-甘露糖-6-磷酸和D-甘露糖-1-磷酸之间的转化,D-甘露糖-1-磷酸是GDP-甘露糖合成的底物。甘露糖-6-磷酸可以抑制参与葡萄糖代谢的三种酶,进而促进半乳糖多糖生成。
如中国专利CN113024681A黄精半乳聚糖及其制备方法和应用,多花黄精经分离提纯得到黄精半乳聚糖具有促进益生菌增殖的活性,对于改善肠道健康具有积极作用,可以用于制备促进消化、肠道蠕动的食品、药物或保健品。
因此,本发明林下黄精L和单作黄精CK虽然都采用相同的培育环境,都应用了李树提供的阴凉环境,以及农家肥作为有机肥补充养分,促进黄精生长,但是李树下套栽的黄精由于地下根系的相互作用,使得黄精的关键酶基因sacA(β-果糖苷酶)、PMM(磷酸甘露糖异构酶)、SPS(蔗糖磷酸合成酶)的相对表达量显著升高,对于黄精种植过程中多糖的合成选择具有积极促进作用,可以使得黄精多糖中黄精果聚糖、黄精半乳聚糖含量增加,进而可以产生促进益生菌增殖的活性,具有非常高的食品、药品或保健品开发价值。
所以,通过黄精与李子树套栽工艺方法,收获得到的多花黄精品质提升,具有更好的药品开发、保健品开发或养生应用的价值,具有积极意义。
最后,在9月份采摘李子果以后,比较套栽多花黄精的李子树产量相比于前有一年未套栽李子树时李子果产量变化。称重结果显示,在李子树林间套栽多花黄精以后,李子果产量略微增加,并不显著,这可能与套栽多花黄精以后,林间除草浇水次数略有增加有关。在李子果产量并无减少的情况下,本发明实现了多花黄精和李子树套栽,额外增加了土壤栽种多花黄精的收获,具有较为明显的经济效益。
- 一种鉴别长梗黄精和多花黄精的特征序列、引物及方法
- 鉴别长梗黄精和多花黄精的特征序列、引物及方法
- 一种多花黄精多糖的提取方法及多花黄精多糖的应用
- 一种区分多花黄精和制多花黄精的质量评价方法及应用