灰毡毛忍冬开花的调控基因及其应用
文献发布时间:2024-04-18 19:58:21
技术领域
本发明涉及植物基因工程领域技术,尤其是指一种灰毡毛忍冬开花的调控基因及其应用。
背景技术
灰毡毛忍冬为忍冬科(Caprifoliaceae)、忍冬属(Lonicera)多年生常绿灌木或藤本,是中国传统中药“山银花”的主要基原植物之一,其药用部位为干燥花蕾及初开的花,具有很高的药用价值和经济价值。普通灰毡毛忍冬抗性差、蕾期短、易凋谢、产量和有效成分含量低;而花蕾型灰毡毛忍冬新品种‘龙花’、‘金翠蕾’等,其花期花冠一直不展开,花蕾整齐均呈棒状,花期长达15~25天,解决了同时段大规模集中采摘困难等问题,为灰毡毛忍冬植物提供了新的优良种质资源。为更好的指导实际生产和迎合未来分子育种的发展趋势,研究花蕾型灰毡毛忍冬蕾期长、花蕾整齐不开放等现象背后的分子机制则显得十分必要。
MADS-box基因家族是植物中一类生物功能丰富的转录调控因子,参与调控植物的生长发育的各个时期,尤其与花的形态发生密切相关,MADS-box基因在调控开花时间、花冠展开等花器官发育方面发挥着重要的调控作用。目前,MADS-box家族基因已从大量不同植物种类中克隆得到,但灰毡毛忍冬MADS-box相关基因的报道微乎其微,仅见AP1、AGL19基因和蕾期延长相关基因AGL15克隆和表达分析以及内参基因筛选等零星报道。从现有文献报道来看,调控花蕾型灰毡毛忍冬蕾期长、花冠不展开的关键基因尚需进一步挖掘与验证。
AGL82基因是该基因家族中的一员,目前尚未有对其的研究报道;申请号为201710819491.6的发明公开了一种甘蔗开花调控蛋白ScFT-2及其编码基因,现有技术未从灰毡毛忍冬中克隆得到AGL82基因,并以灰毡毛忍冬AGL82基因为切入点,通过载体构建、遗传转化拟南芥等方法研究灰毡毛忍冬AGL82基因的功能及其在植物花器官发育中的应用;因此,基于此现状,迫切需要开发一种灰毡毛忍冬开花的调控基因及其应用,以满足实际使用的需求。
发明内容
有鉴于此,本发明针对现有技术存在之缺失,其主要目的是提供一种灰毡毛忍冬开花的调控基因及其应用,其提供了灰毡毛忍冬花器官发育相关的AGL82基因及其编码蛋白序列,以及制备AGL82基因的方法和制备过程中所需的引物,并通过AGL82基因过表达载体的成功构建、遗传转化拟南芥等技术手段验证了AGL82基因可促进植物开花的功能。
为实现上述目的,本发明采用如下之技术方案:
一种灰毡毛忍冬开花的调控基因,所述调控基因为AGL82,其基因序列如SEQ IDNO.1所示;所述调控基因的编码蛋白质的氨基酸序列如SEQ ID NO.2所示。
作为一种优选方案:所述调控基因能够通过RNA提取及反转录进行克隆。
作为一种优选方案:所述调控基因进行克隆的方法包括如下步骤:
第一:以灰毡毛忍冬为材料,提取RNA,并将提取的RNA进行反转录生成cDNA第一链,作为PCR扩增的模板;
第二:将获得的灰毡毛忍冬AGL82基因进行序列分析并设计PCR反应引物;
第三:以灰毡毛忍冬cDNA第一链为模板,采用AGL82PCR反应引物进行PCR扩增,PCR扩增产物进行1.0%琼脂糖凝胶电泳,并按琼脂糖凝胶纯化回收试剂盒回收PCR产物;回收PCR产物与pTOPO载体连接并转化至大肠杆菌DH5α感受态细胞,挑取阳性克隆并测序。
作为一种优选方案:所述PCR反应引物为LmAGL82-F,其引物序列为:ATGGGACGAGCTAGAGTCAGAA。
作为一种优选方案:所述PCR反应引物为LmAGL82-R,其引物序列为:CATGGCTAATCAGTAATTAATTGAAC。
作为一种优选方案:所述PCR扩增PCR扩增的反应体系为:2×Phanta Max Buffer12.5μL,dNTP Mix 0.5μL,SVP-F和SVP-R各1μL,Phanta Max Super-Fidelity DNAPolymerase 0.5μL,cDNA模板1.5μL,ddH2O 8.0μL;PCR反应程序为:95℃预变性3min;95℃变性15s,60℃退火15s,72℃延伸1min,35个循环;72℃彻底延伸5min;12℃持续。
所述灰毡毛忍冬开花的调控基因的应用,所述调控基因用于调控灰毡毛忍冬开花期。
作为一种优选方案:所述调控基因应用于植物过表达载体的构建,该调控基因能够促进灰毡毛忍冬提前开花。
作为一种优选方案:所述植物过表达载体的构建具体方法为:采用同源重组的方法构建pCAMBIA1301-LmAGL82植物过表达载体,使用同源重组引物扩增出目的片段,然后重组至线性化载体pCAMBIA1301载体上,将重组反应转化大肠杆菌并挑取单克隆鉴定测序。
作为一种优选方案:所述调控基因用于获取转基因植株。
本发明与现有技术相比具有明显的优点和有益效果,具体而言,由上述技术方案可知,本申请提供了灰毡毛忍冬花器官发育相关的AGL82基因及其编码蛋白序列,以及制备AGL82基因的方法和制备过程中所需的引物,并通过AGL82基因过表达载体的成功构建、遗传转化拟南芥等技术手段验证了AGL82基因可促进植物开花的功能,同时也提供了该基因在植物性状改良中的一种应用路径,表明了该基因具有良好的应用前景,为培育花类药材和花类观赏植物新品种提供了宝贵的基因资源。
为更清楚地阐述本发明的结构特征和功效,下面结合附图与具体实施例来对其进行详细说明。
附图说明
图1为本发明之灰毡毛忍冬LmAGL82基因ORF克隆示意图;
图2为本发明之灰毡毛忍冬LmAGL82蛋白的NJ系统进化树示意图;
图3为本发明之LmAGL82基因在灰毡毛忍冬‘龙花’和‘金翠蕾’品种不同发育期中表达模式示意图;
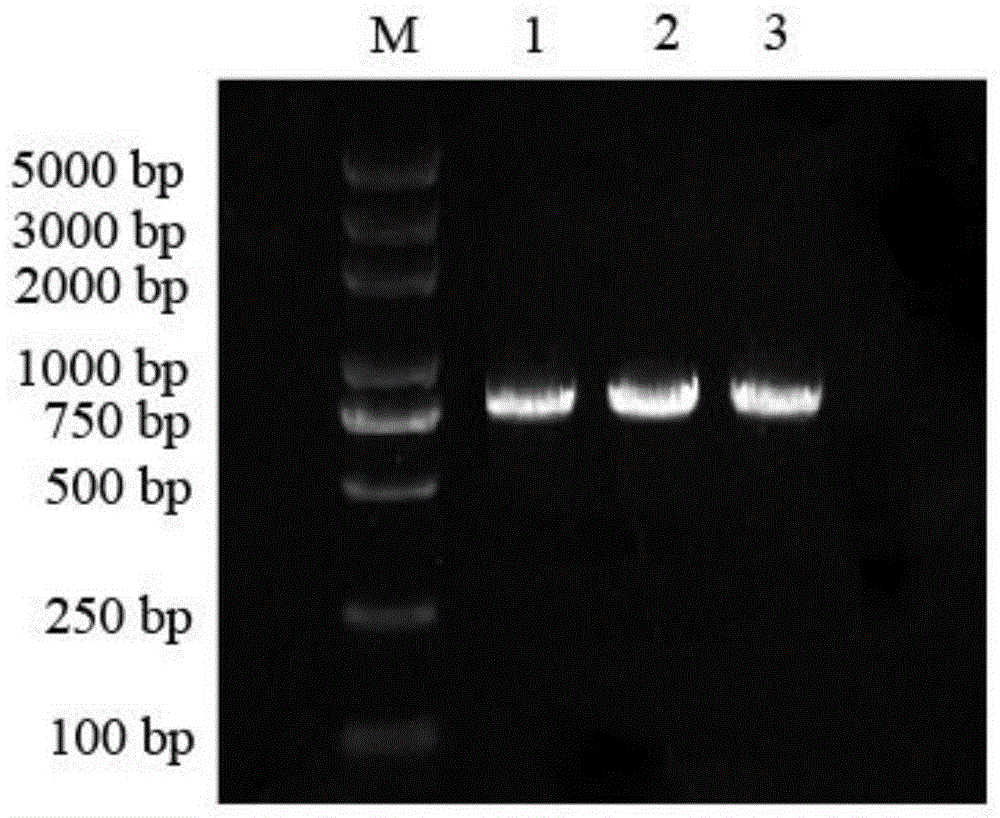
图4为本发明之LmAGL82重组菌液PCR扩增片段示意图;
图5为本发明之拟南芥转基因株系的筛选与鉴定示意图。
具体实施方式
本发明如图1至图5所示,一种灰毡毛忍冬开花的调控基因,该调控基因为AGL82,其基因序列如SEQ ID NO.1所示;该调控基因的编码蛋白质的氨基酸序列如SEQ ID NO.2所示。
该调控基因能够通过RNA提取及反转录进行克隆。
该调控基因进行克隆的方法包括如下步骤:
第一:以灰毡毛忍冬为材料,提取RNA,并将提取的RNA进行反转录生成cDNA第一链,作为PCR扩增的模板;
第二:将获得的灰毡毛忍冬AGL82基因进行序列分析并设计PCR反应引物;
第三:以灰毡毛忍冬cDNA第一链为模板,采用AGL82PCR反应引物进行PCR扩增,PCR扩增产物进行1.0%琼脂糖凝胶电泳,并按琼脂糖凝胶纯化回收试剂盒回收PCR产物;回收PCR产物与pTOPO载体连接并转化至大肠杆菌DH5α感受态细胞,挑取阳性克隆并测序。
该PCR反应引物为LmAGL82-F,其引物序列为:ATGGGACGAGCTAGAGTCAGAA;该PCR反应引物为LmAGL82-R,其引物序列为:CATGGCTAATCAGTAATTAATTGAAC。
该PCR扩增PCR扩增的反应体系为:2×Phanta Max Buffer12.5μL,dNTP Mix0.5μL,SVP-F和SVP-R各1μL,Phanta Max Super-Fidelity DNA Polymerase 0.5μL,cDNA模板1.5μL,ddH2O 8.0μL;PCR反应程序为:95℃预变性3min;95℃变性15s,60℃退火15s,72℃延伸1min,35个循环;72℃彻底延伸5min;12℃持续。
该调控基因用于调控灰毡毛忍冬开花期。
该调控基因应用于植物过表达载体的构建,该调控基因能够促进灰毡毛忍冬提前开花。
该植物过表达载体的构建具体方法为:采用同源重组的方法构建pCAMBIA1301-LmAGL82植物过表达载体,使用同源重组引物扩增出目的片段,然后重组至线性化载体pCAMBIA1301载体上,将重组反应转化大肠杆菌并挑取单克隆鉴定测序。
该调控基因用于获取转基因植株。
实施例1:灰毡毛忍冬LmAGL82基因的克隆及分子特征分析:
(1)以灰毡毛忍冬‘龙花’品种为材料,提取RNA,并将提取的RNA进行反转录生成cDNA第一链,作为PCR扩增的模板。
(2)从转录组获得的灰毡毛忍冬LmAGL82基因进行序列分析设计特异性引物,PCR反应引物序列如表1:
表1:PCR反应引物
(3)以灰毡毛忍冬cDNA第一链为模板,采用LmAGL82特异性引物进行PCR扩增,反应体系(25μL):2×Phanta Max Buffer 12.5μL,dNTP Mix 0.5μL,SVP-F和SVP-R各1μL,Phanta Max Super-Fidelity DNA Polymerase 0.5μL,cDNA模板1.5μL,ddH
(4)对上述克隆获得的LmAGL82基因及其编码蛋白进行生物信息学分析。
结果分析:从灰毡毛忍冬转录组中获得该基因的CDS序列,以灰毡毛忍冬cDNA作为模板,采用RT-PCR的方法进行PCR扩增。以1%琼脂糖凝胶电泳检测PCR扩增产物,结果显示在靠近Marker的850bp附近出现了一条清晰明显的条带,与预期大小相符,见图1。鉴定阳性单克隆并送测序,比对测序结果发现与转录组中序列一致,序列如SEQ ID NO.1。
用EditSeq和ExPASy ProtParam在线分析结果表明:克隆得到的LmAGL82基因的开放阅读框为846,编码281个氨基酸的蛋白质(SEQ ID NO.2),推测其分子式为C
为进一步了解LmAGL82蛋白的系统进化情况,用大豆(Glycine max)、林烟草(Nicotiana sylvestris)、马铃薯(Solanum tuberosum)、君迁子(Diospyros lotus)、毛花猕猴桃(Actinidia eriantha)、大花窄叶油茶(Camellia lanceoleosa)、向日葵(Helianthus annuus)、喜马拉雅凤仙花(Impatiens glandulifera)、烟草(Nicotianatabacum)、拟绒毛烟草(Nicotiana tomentosiformis)、红马银花(Rhododendron vialii)、西班牙鼠尾草(Salvia hispanica)、芝麻(Sesamumindicum)、欧白英(Solanumdulcamara)、茶(Camellia sinensis)等与LmAGL82氨基酸序列具有一定同源性的MADS-box家族的氨基酸序列构建进化树。结果表明,克隆得到的灰毡毛忍冬基因属于AGL82类(图2),被命名为LmAGL82。
实施例2:灰毡毛忍冬LmAGL82基因的表达特性分析:
利用qRT-PCR研究LmAGL82基因在灰毡毛忍冬‘龙花’和‘金翠蕾’品种花蕾和叶中的表达模式。
(1)提取灰毡毛忍冬‘龙花’和‘金翠蕾’品种叶片、早期花蕾(花蕾平均长14.49mm)以及晚期花蕾(花蕾平均长41.85mm)的总RNA并反转录成cDNA;
(2)选用灰毡毛忍冬的18SrRNA基因作为qPCR分析的内参基因,运用PrimerPremier 6.0设计内参基因和目标基因的特异性引物,引物如表2:
表2灰毡毛忍冬实时荧光定量PCR引物
(3)以反转录获得的灰毡毛忍冬cDNA第一链为模板,对LmAGL82基因在灰毡毛忍冬不同品种、不同组织以及花蕾不同发育时期中的表达情况进行qPCR分析。反应体系:2×RealStar Green Fast Mixture PCR 5μL,Forward primer 0.4μL,Reverse primer 0.4μL,cDNA 1μL,ddH
结果分析:
以灰毡毛忍冬18S rRNA为内参基因,利用qRT-PCR技术对灰毡毛忍冬‘龙花’和‘金翠蕾’品种不同组织部位(花、叶)及不同花发育时期LmAGL82基因的表达情况进行分析,见图3。结果显示,无论是在叶片还是花蕾早期、晚期中,‘金翠蕾’品种中LmAGL82基因表达量整体高于‘龙花’品种;LmAGL82基因在‘龙花’和‘金翠蕾’花蕾和叶片中的表达存在极显著差异(**P<0.01),叶片中表达量显著高于花蕾中的表达量;‘龙花’品种中,LmAGL82基因在花蕾早期、晚期中的表达量差异不明显,‘金翠蕾’品种中,LmAGL82基因在花蕾早期中表达量显著高于花蕾晚期中的表达量。
实施例3:灰毡毛忍冬LmAGL82基因过表达载体构建及拟南芥遗传转化
(1)植物过表达载体构建:
根据LmAGL82基因序列自身含有的酶切位点及所用的植物过表达载体pCAMBIA1301所含多克隆位点的特征,选择出最合适的酶Nco I和EcoR I,设计带有载体同源序列的特异性引物,引物名称及序列如下:
p-LmAGL82-F:ACGGGGGACTCTTGA
p-LmAGL82-R:GCAACAGGATTCAAT
使用限制性内切酶QuickCut Nco I和EcoR I双酶切pCAMBIA1301载体,以实施例1中测序正确的菌液为模板,PCR扩增出插入片段,酶切产物和PCR扩增产物经电泳检测、回收、纯化后使用ClonExpress IIOneStep Cloning Kit构建进行同源重组反应,获得重组产物后转化大肠杆菌DH5α感受态细胞,培养后对单克隆进行鉴定及测序,比对测序结果并提取测序正确的菌液质粒。
提取的重组载体质粒转化农杆菌GV3101,使用花器官侵染法将含有pCAMBIA1301-LmAGL82载体质粒农杆菌遗传转化拟南芥。
表3:质粒pCAMBIA1301双酶切反应体系
反应条件:37℃反应1h。
表4:重组反应体系
反应条件为:37℃反应30min。
(2)转基因拟南芥的Hyg抗性筛选与鉴定:
收取侵染后拟南芥植株所结的T
提取T
PCR反应体系:2×Taq mix10μL;genomic DNA2μL;引物0.8μL;ddH
PCR反应条件:94℃3min;94℃30sec,60℃30sec,72℃1min;35cycles;72℃5min。
(3)转基因拟南芥的表型观察:
观察LmAGL82转基因拟南芥及野生型拟南芥生长状况,统计其抽葶时间(以花葶伸出1cm为准)、第一朵花蕾出现时间、花冠展开时间及对应时间的莲座叶数量,数据统计每个株系包含12个植株。
表5转LmAGL82基因拟南芥T2代表型统计
结果分析:
采用同源重组的方法构建pCAMBIA1301-LmAGL82植物过表达载体,使用同源重组引物扩增出目的片段,然后重组至线性化载体pCAMBIA1301载体上,将重组反应转化大肠杆菌并挑取单克隆鉴定测序;菌液PCR后电泳检测结果(图4)表明目的片段连接至载体,比对测序结果与转录组中序列一致,pCAMBIA1301-LmAGL82植物过表达载体构建成功。
采用花器官侵染法将含有pCAMBIA1301-LmAGL82载体质粒农杆菌遗传转化拟南芥,种子成熟后收取晾干,消毒后点播至1/2MS固体培养基上,以野生型拟南芥为对照,将抗性筛选后生长健壮的拟南芥幼苗移栽继续培养,并提取DNA进行PCR检测,检测结果如图5。经过潮霉素筛选和PCR鉴定共获得LmAGL82转基因拟南芥16个株系,待鉴定后的转基因拟南芥植株种子成熟后,收取种子筛选至T3代备用。
以野生型拟南芥WT为对照,对过表达LmAGL82转基因拟南芥的发育状况进行观察;结果发现,LmAGL82转基因植株抽薹时间及花蕾出现时间均比野生型拟南芥显著提前。其中LmAGL82-#2抽薹开花最早,仅需23.85d抽薹达到1cm,第一朵花蕾出现时间为32.67d,花冠展开时间为35.56d;而野生型拟南芥在30.86d后才能抽薹1cm,第一朵花蕾出现时间为40.25d,花冠展开时间为42.38d。除抽薹及花冠展开时间外,拟南芥的花期研究中常以开花时莲座叶数量来反映拟南芥的发育时间,进而衡量花期变化。本研究发现LmAGL82转基因植株第一朵花开放时对应的莲座叶数量也与野生型拟南芥有差异,其中LmAGL82-#2莲座叶数量为9.85,野生型拟南芥为12.66。这些结果均表明LmAGL82在拟南芥中异源表达能促进植株提前开花。
以野生型拟南芥WT为对照,观察过表达LmAGL82转基因拟南芥生长发育状况发现,LmAGL82转基因植株三个株系花冠展开时间分别为35.56d、40.72d、36.25d,较野生型植株花冠展开时间显著提前,说明该基因可以促进拟南芥提前开花。
本发明的设计重点在于,本申请提供了灰毡毛忍冬花器官发育相关的AGL82基因及其编码蛋白序列,以及制备AGL82基因的方法和制备过程中所需的引物,并通过AGL82基因过表达载体的成功构建、遗传转化拟南芥等技术手段验证了AGL82基因可促进植物开花的功能,同时也提供了该基因在植物性状改良中的一种应用路径,表明了该基因具有良好的应用前景,为培育花类药材和花类观赏植物新品种提供了宝贵的基因资源。
以上所述,仅是本发明的较佳实施例而已,并非对本发明的技术范围作任何限制,故凡是依据本发明的技术实质对以上实施例所做的任何细微修改、等同变化与修饰,均仍属于本发明技术方案的范围内。
/>
- 灰毡毛忍冬金银花结蕾不开花品种的选育方法
- 灰毡毛忍冬金银花结蕾不开花品种的选育方法