茶树糖基转移酶CsUGT84J2基因及其增强植物耐铝能力的应用
文献发布时间:2024-01-17 01:26:37
技术领域
本发明涉及基因工程技术领域,具体涉及一种茶树糖基转移酶CsUGT84J2基因及其增强植物耐铝能力的应用。
背景技术
据研究报道,世界上大约百分之五十的可耕作性土地为酸性土壤。在我国华南地区的土壤也以酸性土壤为主。土壤富含铝,主要以氧化铝、硅化铝的形式存在,当土壤溶液酸化后(pH<5),铝就会从氧化物或硅酸盐中释放出来,通常被认为是一种植物毒素,铝分解成可溶性Al
与大多数植物不同的是,茶树是一种典型的耐铝植物。茶树通常富含铝,据报道,铝主要在茶树叶子中积累,特别是老叶,茶树的生长过程中,铝是不可缺少的,茶树需要浓度适宜的铝帮助其到达最适生长状况,同时可以获得高品质的茶叶,提升了茶叶的商用价值。但铝促进茶树生长的机理尚不清晰,茶树对于铝元素的需求较其它植物相对特殊,低浓度铝对茶树生长发育是必需的,尤其是茶树根系的生长,然而相关机理研究相对匮乏。
发明内容
本发明的目的在于克服现有技术的不足,提供了一种茶树糖基转移酶CsUGT84J2基因及其增强植物耐铝能力的应用。
本发明通过以下技术方案来实现上述目的:
本发明提供了一种茶树糖基转移酶CsUGT84J2基因,该基因具有如SEQ ID NO.1所示的核苷酸序列,该基因是从茶树组织中分离获得的,该基因受铝诱导并对黄酮醇和生长素具有糖基化活性。
进一步改进在于,所述茶树组织包括芽、二叶、四叶、成熟叶、茎或根。
本发明还提供了一种上述茶树糖基转移酶CsUGT84J2基因的编码蛋白,所述编码蛋白具有如SEQ ID NO.2所示的氨基酸序列。
本发明还提供了一种含有上述茶树糖基转移酶CsUGT84J2基因的生物材料,所述生物材料为质粒载体或工程菌。
进一步改进在于,所述质粒载体包括但不限于pMal-c2X、pDONR207、pCB2004;所述工程菌包括但不限于大肠杆菌BL21、农杆菌GV3101。
本发明还提供了一种上述CsUGT84J2基因、上述编码蛋白或上述生物材料的以下任一应用:
(1)用于促进植物根系生长;
(2)用于提高酸性土壤中铝敏感植物的耐铝性。
进一步改进在于,所述植物为茶树或拟南芥。
进一步改进在于,通过催化植物的黄酮醇和生长素的糖苷化,并调节植物内源性生长素的稳态平衡以促进植物根系生长。
本发明还提供了一种增强植物耐铝能力的方法,所述方法选自如下①或②:
①使植物表达权利要求3所述CsUGT84J2基因的编码蛋白;
②在植物中过表达权利要求1-2任一所述的CsUGT84J2基因;
所述过表达的方式通过导入具有所述CsUGT84J2基因的过表达载体,将所述CsUGT84J2基因通过过表达载体导入植物中或通过基因工程手段整合到植物的染色体上。
本发明还提供了一种增强拟南芥耐铝性的方法,构建植物过量表达载体pCB2004-CsUGT84J2,通过化学法转化将pCB2004-CsUGT84J2质粒转移至表达宿主菌株GV3101中,通过花侵染法将农杆菌转入野生型拟南芥中,使得CsUGT84J2基因在拟南芥的野生型中进行异源表达。
本发明提具有如下有益效果:本发明提供的茶树糖基转移酶CsUGT84J2基因,在体内体外均能催化黄酮醇和生长素的糖苷化,并进一步调节植物内源性生长素的稳态平衡,从而促进茶树、拟南芥根系生长。本研究为铝促进茶树生长机理的研究奠定基础,对茶树的栽培育种具有一定的指导意义。
附图说明
图1为CsUGT84J2基因的铝诱导达;
图2为CsUGT84J2基因的组成型表达和铝诱导表达;
图3为CsUGT84J2的琼脂糖电泳图;
图4为重组蛋白rCsUGT84J的SDS-PAGE电泳图;
图5为rCsUGT84J2酶反应产物的UPLC分析;
图6为rCsUGT84J2酶产物QqQ-UPLC-MS/MS检测分析;
图7为rCsUGT84J2酶的底物特异性分析;
图8为CsUGT84J2过表达拟南芥中酚酸、黄酮醇和生长素的积累模式;
图9为CsUGT84J2过表达拟南芥中酚酸、黄酮醇和生长素途径相关基因的表达模式;
图10为CsUGT84J2过表达的拟南芥表型分析;
图11为CsUGT84J2过表达的拟南芥铝耐受能力分析;
图12为茶树中CsUGT84J2过表达和CsUGT84J2沉默中酚酸、黄酮醇和生长素相关的基因表达和代谢积累。
具体实施方式
下面结合附图对本申请作进一步详细描述,有必要在此指出的是,以下具体实施方式只用于对本申请进行进一步的说明,不能理解为对本申请保护范围的限制,该领域的技术人员可以根据上述申请内容对本申请作出一些非本质的改进和调整。
1、材料
本实施例所用方法如无特别说明均为本领域的技术人员所知晓的常规方法,所用的试剂等材料,如无特别说明,均为市售购买产品。
选取舒茶早的芽、二叶、四叶、成熟叶、茎、根以及2mM铝处理的茶树根为实验材料,取样后立即用液氮冷冻,然后保存在-80℃的冰箱中备用。用于本发明中基因的克隆和基因的表达分析。
对于茶园的铝处理实验,将安徽农业大学实验茶园(中国安徽省合肥市,
31.52°N117.14°E)-农萃园的五年生“舒茶早”茶树用浓度为2mM的铝溶液灌溉处理半年,从2020年7月至2020年12月,每两周处理一次,每次处理需两升含铝溶液。
拟南芥种植于黑色小方盆中,放置于温度为20±2℃的恒温培养室中自然生长,其光周期为光照16小时,黑暗8小时。
2、方法
2.1基因克隆
2.1.1RNA的提取
(1)取100-200mg茶树组织,加入等量的PVPP,置于液氮中,充分研磨,将lmL枪头液氮预冷后,将粉末移入2mLRNAase-free离心管中。
(2)向离心管中加入1mLFruit-mate,充分混匀,35℃裂解25min,然后4℃,12000r/min,离心5min。
(3)小心吸取上清液500μL,置于新的1.5mL的离心管中,加入等体积的RNAisoplus,盖紧离心管盖子,用力震荡,待溶液充分乳化后,低温静止5min。
(4)分别加入1/5体积的氯仿,约200μL,反复颠倒混匀,待溶液充分乳化后,低温静置5min。4℃,12000r/min,离心5min。
(5)吸取500μL上清液转移至新的1.5mL离心管中,加入等体积的异丙醇上下颠倒充分混匀后,低温静置10min,4℃,12000r/min,离心10min。
(6)小心弃去上清,加入75%乙醇1mL,轻轻上下颠倒混匀,至RNA从离心管壁脱落,4℃,12000r/min,离心5min。重复两次,第三次用无水乙醇。
(7)小心弃去乙醇后,再5000r/min,空离心1min,用枪头吸走多余的乙醇,以便后续真空干燥,干燥时,开离心管盖,旋转1min。
(8)观察离心管底有没有乙醇残留,RNA呈湿润状态即可,加入30μL的RNAas e-free水,溶解沉淀,保存在-80℃备用。
2.1.2基因全长克隆及验证
使用Premier5.0软件对CsUGT84J2基因(该基因具有如SEQ ID NO.1所示的核苷酸序列)设计了开放阅读框扩增所用引物,和针对pMal-c2X载体,pDONR207,pCB2004的多克隆位点分别设计了带酶切位点的特异引物。引物序列为:
CsUGT84J2-ORF-F:ATGGCTGCCATGGAAAAGAAAG;
CsUGT84J2-ORF-R:TCACGTACTAGAATCCCCAAT;
CsUGT84J2-PMAL-F:GGGGACAAGTTTGTACAAAAAAGCAGGCTATGGCTGCCATGGAAAAGAAAG;
CsUGT84J2-PMAL-R:GGGACCACTTTGTACAAGAAAGCTGGGTTCACGTACTA GAATCCCCAAT。
根据TAKARA高保真酶的程序,以茶树根的cDNA为模板克隆了目的基因的全长序列,使用康为胶回收试剂盒回收目的条带以后,送通用公司测序,结果与预测一致。
2.2基因的表达分析
对于目的基因的组成型表达分析,以舒茶早的不同组织器官(芽,二叶,四叶,成熟叶,茎和根)的cDNA为模板;对于目的基因的诱导型表达分析,以2mM铝处理和对照的舒茶早侧根(根尖1cm)的cDNA为模板,进行荧光定量分析。荧光定量的引物序列为CsUGT84J2-RT-F:ACACGGCAAAGGAGGGAGA;CsUGT84J2-RT-R:GGTCCGT CCACTGTGGATAAG。
结果如图1和图2所示:
CsUGT84J2基因在成熟叶和根中的表达量最高,在芽中的表达量最低。铝处理后根中CsUGT84J2的表达水平是对照的2.3倍。该结果表明CsUGT84J2基因是一个受铝诱导的UGT。且铝在茶树中的积累以根和成熟叶中居多,CsUGT84J2基因的组织特异性表达与铝的积累部位是一致的。
2.3基因的原核表达
2.3.1原核表达载体的构建
将含有目的基因ORF编码框的质粒通过体外酶切酶连法,构建到pMal-c2X载体中。再将带有目的基因的pMal-CsUGT84J2转入到表达宿主菌株BL21中。通过挑选和验证的阳性菌株,加入50%的甘油后,保存在-80℃备用。
2.3.2重组融合蛋白的纯化及SDS-PAGE分析
2.3.2.1蛋白纯化
将目的菌株和阳性对照菌株,通过活化,扩大,加入40%的葡萄糖和IPTG诱导后,5500r/min,10℃,多次离心收集菌体,保存在-20℃备用。
将收集的菌体放置于冰上解冻,用上柱缓冲溶液充分重悬,后用超声细胞破碎仪以6℃,25%的功率,破碎菌液30min。后用低温高速离心机以5500r/min,10℃,离心8min30s。用上柱缓冲溶液多次冲洗填充好且带有麦芽糖标签的直链淀粉树脂蛋白纯化柱,去除残留的杂蛋白,使其保持一个相对干净的环境,后将上清蛋白溶液导入蛋白纯化柱,以1mL/min的流速上柱,用上柱缓冲溶液冲洗三次,洗脱杂蛋白,再用洗脱缓冲溶液冲洗并收集。将收集的液体用超滤管离心浓缩,得到蛋白样品。用考马斯亮蓝显色法制备蛋白样品,并用分光反光度计测蛋白浓度(595nm下测OD值)。
2.3.2.2SDS-PAGE蛋白分析
制备蛋白电泳样品时,取20μg的蛋白样品,加入8μL5×loadingbuffer,用纯水补足至20μL。吹打混匀,用电磁炉煮沸10分钟。将蛋白电泳样品用移液枪移取10μL至上样孔中,初以80V的电压在浓缩胶中电泳,当样品条带浓缩成一条线并进入分离胶时,将电压调为150V。当指示剂迁移至距胶底端1cm左右时,关闭电源,停止电泳,从电泳槽中取出凝胶玻璃板,轻轻撬开玻璃板,将胶转移至培养皿中。向培养皿中加入染色液,没过凝胶即可,加热一分钟左右至胶底色呈现蓝色。回收染色液,加入脱色液进行脱色,每隔一段时间更换一次洗脱液并加热脱色,直到底色干净并出现清晰的电泳条带,用凝胶成像系统扫描凝胶并成像。
如图3和图4所示:重组质粒能被BamH1/Sal1和BamH1/Sal2双酶切,表明CsUGT84J2基因的完整片段已成功连接到pMal-BL21质粒载体上。SDS-PAGE电泳图能看到明显且长度正确的蛋白条带,表明重组蛋白rCsUGT84J2(具有如SEQ ID NO.2所示的氨基酸序列)被成功纯化。
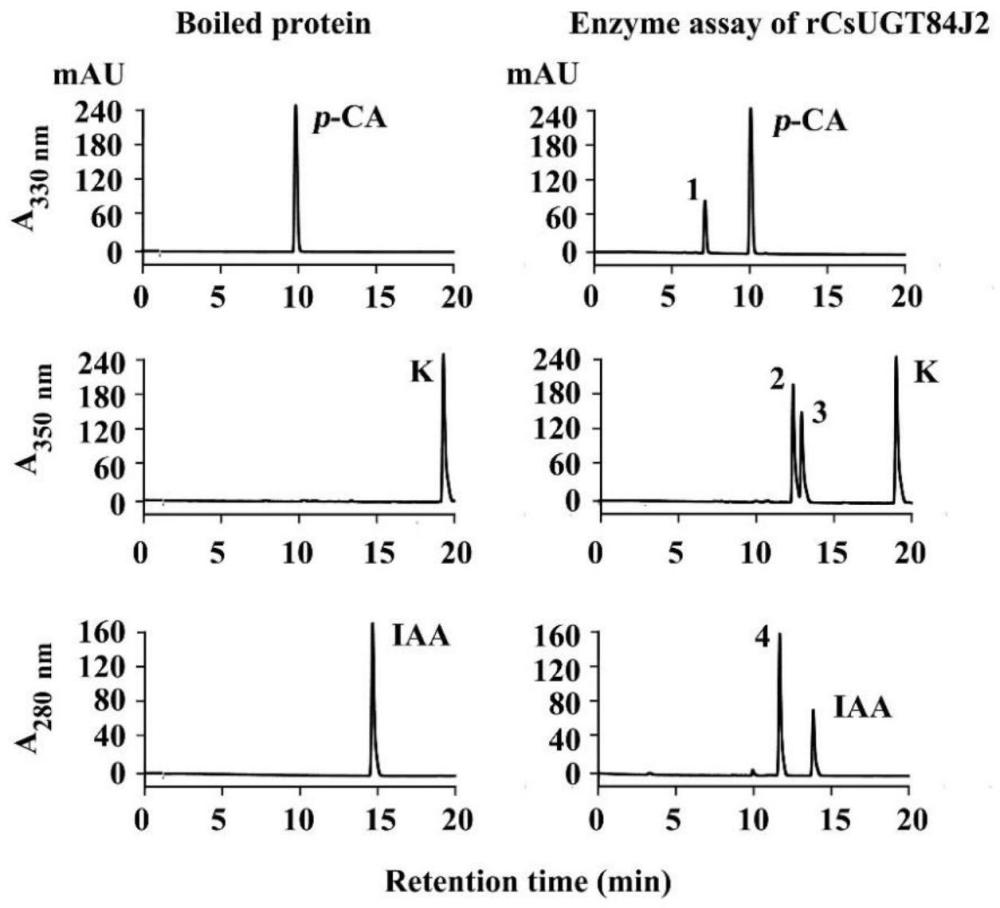
2.3.3酶活检测和产物鉴定
本发明以对酚酸类,黄酮醇类和激素类三类底物作为候选基因的糖受体,以UDP-葡萄糖或UDP-鼠李糖为糖供体,在体外酶活性实验中分析其底物糖基化活性。所有反应混合物的总体系为50μL,其中含有20μg的rCsUGT84J2重组蛋白、2mM的UDP-葡萄糖、0.5mM糖受体,用缓冲液补足至50μL。在30℃的水浴锅中反应60min,加入等体积色谱级别的甲醇,涡旋混匀,使反应迅速终止。并用UPLC检测酶活产物。
对于酚酸类底物的活性检测,反应为50μL酶活测定体系包括2mM的UDP-葡萄糖、0.3mM酚酸类化合物(没食子酸,对羟基苯甲酸,对香豆酸,咖啡酸和芥子酸),约20μg纯化后的重组蛋白,于200mM,pH=7.5的Tris缓冲溶液中进行。
对于黄酮类底物的活性检测,反应为50μL酶活测定体系包括2mM的UDP-葡萄糖、0.3mM黄酮类化合物(山奈酚和槲皮素),约20μg纯化后的重组蛋白,于200mM,pH=7.5的Tris缓冲溶液中进行。
对于激素类底物的酶活检测,反应为50μL酶活测定体系包括2mM的UDP-葡萄糖、0.3mM激素类化合物(吲哚-3-乙酸,吲哚-3-丁酸,水杨酸和脱落酸),约20μg纯化后的重组蛋白,于200mM,pH=7.5的Tris缓冲溶液中进行。
酶反应产物经产物标准品,以及UPLC及QqQ-UPLC-MS/MS相结合的方法进行鉴定。如图5和图6所示,重组蛋白rCsUGT84J2对于酚酸,黄酮醇和生长素三类底物都具有活性。
2.3.4酶动力学分析
酶动力学参数是通过米氏曲线获得。以最适反应条件为前提,选择UDP-葡萄糖作为糖供体,分别选择三类底物中的对香豆酸,山奈酚和吲哚-3-乙酸为糖受体,进行酶动力学分析。
反应为50μL酶活测定体系包括2mM的UDP-葡萄糖、10-400uM的对香豆酸,山奈酚和吲哚-3-乙酸,约20μg纯化后的重组蛋白,用最适pH的缓冲溶液补足至50μL。在最适反应温度下,反应90min,用等体积的色谱甲醇终止反应,立即吹打混匀,离心。取50μL上清液进行HPLC分析。
酶活性的计算基于反应后产物峰面积的增加。如图7所示,rCUGT84J2对山奈酚(KM=0.127μM)的亲和力最高,然后对对香豆酸(KM=0.243μM)和IAA(KM=0.522μM)的亲和力较低。因此,动力学参数表明rCsUGT84J2对山奈酚的催化活性高于对香豆酸和吲哚-3-乙酸。
2.4CsUGT84J2基因的异源表达
2.4.1CsUGT84J2在拟南芥中的质粒构建和异源表达
通过PCR反应将带有attb接头的引物序列连接到CsUGT84J2完整的ORF编码框上。将PCR产物使用GatewayBP克隆酶混合物进行纯化,并克隆到入门载体pDONR207中,随后使用GatewayLR克隆酶系统克隆到表达载体pCB2004中,然后将带有目的基因的表达载体(pCB2004-CsUGT84J2)和不带有目的基因的空白表达载体(pCB2004)转移到表达宿主菌株GV3101中。
引物序列为:CsUGT84J2-F(attb):GGGGACAAGTTTGTACAAAAAAGCAGGCTATGGCTGCCATGGAAAAGAAAG;CsUGT84J2-R(attb):GGGGACCACTTTGTACAAGAAAGCTGGGTTCACGTACTAGAATCCCCAAT。
通过化学法转化将重组pCB2004-CsUGT84J2质粒转移至表达宿主菌株GV3101中。野生型拟南芥(Col-0)用于农杆菌介导的转化,通过花侵染法将农杆菌转入野生型拟南芥中。
2.4.2酚类化合物和生长素的提取和定量
使用UPLC分离方法系统地分析了从活性测定和动力学参数分析测定中获得的产物。取对照和转基因三个品系(line8,line10,line11)的生长21天的拟南芥幼苗(整颗植株)各100mg,三个重复。加入500μL的80%的甲醇,通过低温超声的方法,提取拟南芥幼苗中酚酸糖苷和黄酮醇糖苷,重复三次,最后定容到2ml。分析拟南芥幼苗叶片中的酚酸苷和黄酮醇苷。相对定量基于主要MS/MS信号(M-H)+的面积。
对于酰胺缀合物、IAA和IAGlc定量,使用了大约200mg全植物组织的样品(对照和转基因三个品系(line8,line10,line11)的生长21天的拟南芥幼苗)。在球磨机中用1mL乙酸乙酯在25℃下萃取10分钟后,将样品离心。然后真空浓缩除去上清液中的有机溶剂,沉淀用50μL甲醇溶解,加入100μL的屈臣氏水,吹打混匀,离心并通过UPL C-QqQ-MS/MS分析。相对定量基于主要MS/MS信号(M-H)+的面积。
如图8所示,结果表明,与对照相比,从过表达CsUGT84J2的拟南芥中提取的苯丙酸含量没有显著增加,例如松柏素、丁香苷和芥子葡萄糖,反而在个别过表达品系中显著下调。但可以证实,黄酮醇苷的含量显著高于对照,尤其是line10,其中主要的二黄酮苷增加了3倍以上,例如,K-3-R-7-R、K-3-G-7-R和Q-3-R-7-R。除了苯丙酸糖苷和黄酮醇糖苷外,我们还观察到生长素及其衍生物水平显著增加,例如IAA、IAA-Glc、IAA-Glu。
如图9所示,苯丙烷途径基因的表达情况并没有显著的变化,与此相反的是,CsUGT84J2基因的过表达导致与黄酮醇途径有关的大多数内源基因的表达上调,尤其是AtF3'H和AtFLS1。调节黄酮醇合成的三种MYB转录因子的表达量也显著上调,尤其是MYB11,表达量上调多达3倍左右。
生长素的积累受生长素的合成、调节、流入和流出的影响。在CsUGT84J2过表达的拟南芥中,TAA1的表达变化不大,但四个关键的YUCCA基因表达量均上调。GH3基因催化生长素向生长素-氨基酸偶联物结合态的转化,在过表达CsUGT84J2的拟南芥中GH3家族基因的表达也上调,例如GH3.1和GH3.3。同时,IAA稳态受生长素输入和生长素输出的调控,输入载体(AUX1)的表达略有增加。另一方面,PIN家族的表达水平也整体增加。此外,生长素响应因子(ARF)的表达水平也普遍上调。生长素的合成、调节、流入和流出共同调节植物中的游离IAA的水平,形成一个生长素稳态平衡。总而言之,这些与生长素途径密切相关的基因表达量的上调共同促进了体内生长素的积累。
2.5拟南芥的铝处理
对于拟南芥的播种实验,用25%浓度的84消毒液对种子进行表面消毒3分钟,然后用无菌去离子水清洗3-4次,洗去种子表面的84消毒液,避免种子死亡,不发芽。将过表达CsUGT84J2的3个品系和对照的种子播种在直径为90mm,高度为15mm的一次性培养皿中,培养皿中装有25mL的MS固体培养基,在20±2℃的恒温室(光照16小时,黑暗8小时)中培养6天。待种子萌发,幼苗生长,用于后续的实验。
对于拟南芥铝处理实验,将生长5天的幼苗从MS蔗糖琼脂板上小心取出,浸入添加了Al
2.6拟南芥生理指标测定
为测定过表达CsUGT84J2拟南芥幼苗的生理指标,将幼苗在MS蔗糖琼脂培养基上垂直生长5天或12天,同时测量生长5天幼苗和12天幼苗的初生根长度、下胚轴长度和侧根的数量。生物学实验重复3次,每次处理60株幼苗。
对于拟南芥的铝处理实验,我们在含有不同铝浓度的固体培养基中将5日龄且长势一致的拟南芥幼苗处理5天,然后测量CsUGT84J2过表达和对照幼苗的初级根长度和次级根数。生物实验重复3次,每次处理60株幼苗。
结果表明,含有CsUGT84J2过表达构建体的转基因拟南芥植株表现出促进生长的表型,尤其是下胚轴和侧根。半定量结果分析显示了CsUGT84J2的mRNA显著水平。从图10中可看出,生长五天的过表达拟南芥幼苗的下胚轴长度显著增加,长度约为对照的两倍。于此同时,初生根的长度也显著增加。此时,侧根还未萌发。随着幼苗的生长,生长九天的CsUGT84J2过表达拟南芥幼苗初生根的差异减小,已无显著效果,而次生根的数量和长度显著增加。与对照相比,生长9天的过表达拟南芥幼苗的次生根数量(1cm)增加。
因此,在CsUGT84J2过表达的拟南芥中,黄酮醇途径和生长素途径基因的上调表达促进了相关代谢物的积累,导致拟南芥下胚轴伸长和次生根发育。
从图11中可看出,在浓度为60μM或100μM的铝处理下,对照和过表达CsUGT84J2的拟南芥中初级根的生长均受到抑制。然而,铝对CsUGT84J2过表达系的抑制作用明显弱于对照。如上所述,CsUGT84J2促进了无铝情况下次生根的生长发育。此外,尽管100μM铝抑制了拟南芥的生长,但在60或100μM铝处理下,CsUGT84J2过表达系的次生根数量显著高于对照植株。
2.7CsUGT84J2在茶树中的过表达和沉默
如图12所示,qRT-PCR分析表明,在CsUGT84J2过表达和CsUGT84J2沉默的茶树叶片中,CsUGT84J2表达分别显著上调或下调。同时,CsUGT84J2过表达茶叶中Cs4CLa、黄酮醇途径中的CsCHSa和CsFLSa以及生长素途径中的CsPIN3表达明显上调,特别是CsCHSa。相反,在CsUGT84J2沉默的茶叶中,这些基因的表达显著下调。
随后,检查酚类化合物的代谢变化,验证CsUGT84J2在茶树中的功能。在CsUGT84J2过表达茶叶中,K-7-O-Glc和p-CA衍生物(如3-p-CQA和5-p-CQA)的浓度显着增强。同时,其他黄酮醇苷的水平没有显着变化。相比之下,在CsUGT84J2沉默的茶叶中,与CsUGT84J2过表达茶叶相比,K-7-O-Glc,3-p-CQA和5-p-CQA的含量显著降低。
以上所述实施例仅表达了本发明的几种实施方式,其描述较为具体和详细,但并不能因此而理解为对本发明专利范围的限制。应当指出的是,对于本领域的普通技术人员来说,在不脱离本发明构思的前提下,还可以做出若干变形和改进,这些都属于本发明的保护范围。
- 茶树己糖激酶CsHXK3基因在调控植物生长发育和增强抗寒能力中的应用
- 茶树己糖激酶CsHXK3基因在调控植物生长发育和增强抗寒能力中的应用