一种高产γ-氨基丁酸的谷氨酸脱羧酶GADZ11及其基因和应用
文献发布时间:2023-06-19 09:54:18
技术领域
本发明涉及基因工程领域,具体地,本发明涉及一种高产γ-氨基丁酸的谷氨酸脱羧酶GADZ11及其基因和应用。
背景技术
γ-氨基丁酸(γ-aminobutyric acid,GABA)是自然界普遍存在的一种四碳非蛋白质氨基酸,主要以兼性离子的方式存于水溶液中(Dhakal et al., 2012)。在微生物中,GABA的生成过程与菌体的耐酸机制直接相关(Wang et al., 2013)。研究发现,大多真核微生物细胞内GABA的存在可促进孢子生长(Hao et al., 1991);另外,GABA在酵母抵御活性氧及有机酸等逆境胁迫中也发挥着重要作用(Cao et al., 2013)。在植物中,GABA影响着植物碳源和氮源的存储,能诱导生物体中乙烯的合成或调节胞内pH的平衡,同时防止植物体受氧化胁迫或渗透压等外界环境的影响,并在抵抗真菌和害虫等多种生理生化反应中均有参与(Bouché et al., 2004)。此外,在动物体内GABA同样发挥着重要的生理功效,主要表现为:(1) 血压与心率的调节(Diana et al., 2014);(2) 促进生长激素分泌(Di Cagnoet al., 2010);(3) 抗衰老(McCann et al., 1984);(4) 保肝利肾(Leventhal et al.,2003);(5) 治疗哮喘(Foster et al., 2006);(6) 其余功能:除上述功能外,GABA还具备其他多种生理功效,如改善机体的脂质代谢、减缓血管动脉硬化、防止皮肤的老化、预防肥胖、抑制癌细胞生长和促进生殖等(Khakhalin et al., 2011),因此GABA在医药领域得到越来越多的关注。GABA减少活动量、促进生长的作用,使其还可以作为一种新型饲料添加剂应用于畜牧业。
目前工业制备GABA包括化学合成和生物转化两种常用方法,生物转化法相比较化学合成法具有反应过程简单、条件温和的优势,而围绕生物转化法的核心便是谷氨酸脱羧酶。谷氨酸脱羧酶(Glutamate decarboxylase,GAD)是一种依赖于磷酸吡哆醛(pyridoxal-5-phosphate,PLP)的II类氨基酸脱羧酶,可催化l-谷氨酸或谷氨酸盐脱羧生成GABA,并释放出CO
发明内容
本发明的目的是提供一种能高效应用的谷氨酸脱羧酶。
本发明的再一目的是提供编码上述谷氨酸脱羧酶的基因。
本发明的另一目的是提供包含上述基因的重组载体。
本发明的另一目的是提供包含上述基因的重组菌株。
本发明的另一目的是提供一种制备上述谷氨酸脱羧酶的基因工程方法。
本发明的另一目的提供上述谷氨酸脱羧酶的应用。
本发明提供了一种从芽孢杆菌中分离得到一种谷氨酸脱羧酶GADZ11,其氨基酸序列如SEQ ID NO:1所示。该酶包括489个氨基酸,不含信号肽序列。
本发明提供了编码上述谷氨酸脱羧酶的基因
本发明还提供了包含上述谷氨酸脱羧酶GADZ11的重组载体,优选为pET28a
本发明还提供了包含上述谷氨酸脱羧酶GADZ11的重组菌株,优选所述菌株为大肠杆菌、酵母菌、芽孢杆菌或乳酸杆菌。
其中,优选所述宿主细胞为大肠杆菌细胞,优选将重组大肠杆菌表达质粒转化至大肠杆菌细胞BL21(DE3),得到重组菌株BL21/
本发明还提供了一种制备谷氨酸脱羧酶GADZ11的方法,包括以下步骤:
1)用上述的重组载体转化宿主细胞,得重组菌株;
2)培养重组菌株,诱导重组谷氨酸脱羧酶表达;
3)回收并纯化所表达的谷氨酸脱羧酶GADZ11。
本发明还提供了上述谷氨酸脱羧酶GADZ11的应用。
本发明首先所要解决的技术问题是克服现有技术的不足,提供一种性质优良的、适合于在饲料、食品工业中应用的新型谷氨酸脱羧酶。本发明获得一种谷氨酸脱羧酶GADZ11,最适温度40℃,最适pH5.0。这些性质符合畜禽动物消化生理特点,能提高饲料消化能和代谢能,降低配方成本,减少环境污染。
此谷氨酸脱羧酶的理论分子量为55.4 kDa。该谷氨酸脱羧酶GADZ11的最适pH为5.0,在pH4.0-5.5的范围内,酶活性均维持在最大酶活性的60%以上。利用大肠杆菌异源表达并纯化后,比活为91.7 U/mg。
本发明还提供了编码上述谷氨酸脱羧酶GADZ11的基因
本发明通过PCR的方法分离克隆了这一谷氨酸脱羧酶基因
本发明还提供了包含上述谷氨酸脱羧酶基因的重组载体,优选为pET28a
本发明还提供了上述谷氨酸脱羧酶的应用。
本发明首先所要解决的技术问题是克服现有技术的不足,提供一种性质优良的、适合于在饲料、食品工业中应用的新型谷氨酸脱羧酶。谷氨酸脱羧酶在室温有较高的活性,因其不需要外加能量供酶促反应,所以在工业和农业上的应用可以节省大量的能源。本发明的谷氨酸脱羧酶最适pH为5.0,在pH4~6都有较高的酶活性。因此本发明的谷氨酸脱羧酶可作为饲料添加剂使用,减少畜禽活动量,减少应激反应,从而促进生长。与序列一致性较高的谷氨酸脱羧酶GADZ20相比,酶活显著增加。
附图说明
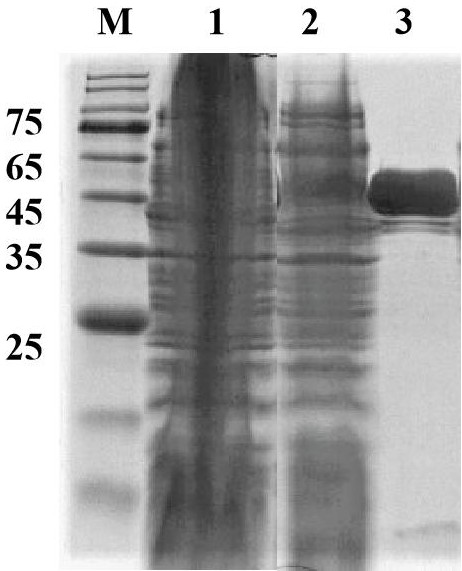
图1显示在大肠杆菌中表达的重组谷氨酸脱羧酶的SDS-PAGE分析,其中,M:低分子量蛋白质Marker;1:诱导的含有空载体的大肠杆菌培养上清液;2:诱导的含有谷氨酸脱羧酶基因的大肠杆菌培养上清液浓缩液;3:通过镍柱纯化的达到电泳纯的Z20蛋白。
图2显示重组谷氨酸脱羧酶的最适pH。
图3显示重组谷氨酸脱羧酶的最适温度。
图4显示l-Glu转化成γ-氨基丁酸的转化率试验。
图5显示GADZ11与GADZ20氨基酸序列比对图。
具体实施方式
试验材料和试剂
1、菌株及载体:大肠杆菌表达载体pET28a(+)及菌株
2、酶类及其它生化试剂:内切酶购自TaKaRa公司,连接酶购自Invitrogen公司。感受态细胞购自南京诺唯赞生物科技有限公司。γ-氨基丁酸、磷酸吡哆醛(PLP)购自Sigma公司,其它都为国产分析纯试剂(均可从普通生化试剂公司购买得到)。
3、培养基:
(1)LB培养基(g/l):酵母粉 5.0,蛋白胨10.0,NaCl 10.0,pH7.0。
(2)平板筛选培养基(g/l):酵母粉 5.0,蛋白胨10.0,NaCl 10.0,琼脂15.0,pH7.0。
说明:以下实施例中未作具体说明的分子生物学实验方法,均参照《分子克隆实验指南》(第三版)J.萨姆布鲁克一书中所列的具体方法进行,或者按照试剂盒和产品说明书进行。
实施例1芽孢杆菌中Z11基因组DNA的提取
枯草芽孢杆菌Z11在LB培养基中30 ℃过夜培养后,在12,000
实施例2 枯草芽孢杆菌Z11来源的谷氨酸脱羧酶编码基因
根据序列设计合成了扩增引物
以上述的基因组总DNA为模板进行PCR扩增。降落PCR反应参数为:95℃变性5 min;95℃变性30 sec,55℃退火30 sec,72℃延伸5 min,25个循环,4℃保温10 min。得到一约1500 bp片段,将该片段回收后与pET28a(+)载体相连送天一辉远生物技术有限公司测序。
通过序列拼接获得该片段的完整序列,全序列共长1470 bp,编码489个氨基酸和一个终止密码子。
实施例3 重组谷氨酸脱羧酶的制备。
将表达载体pET28a(+)进行双酶切 (
取含有重组质粒的BL21菌株,接种于40 mL LB(50 µg/mL的卡那酶素)培养液中,37 ℃培养过夜。按1%的接种量接菌于400 mL LB(含有50 µg/mL的卡那酶素)培养基,于37℃振荡培养约2~3 h后(OD
实施例4 重组谷氨酸脱羧酶GADZ11的活性分析、及性质测定
HPLC法:具体方法如下:在pH5.0,40℃条件下,1 mL的反应体系包括50 μL酶液,500 μL底物,400 μL Buffer,50 μL PLP。反应30 min,加入1 mL 80%的乙醇终止反应。样品离心后取500 µL,加100 µL的NaHCO
1、重组谷氨酸脱羧酶Z11的最适pH的测定方法如下:
将实施例3纯化的重组谷氨酸脱羧酶在不同的pH下进行酶促反应以测定其最适pH。底物L-Glu用不同pH的0.1 M柠檬酸-磷酸氢二钠缓冲液中40℃下进行谷氨酸脱羧酶活力测定。结果(图2)表明,Z11的最适pH为5.0,在pH4.5~5.5的范围内,酶活性均维持在最大酶活性的60%以上。
2、谷氨酸脱羧酶的最适温度测定方法如下:
谷氨酸脱羧酶的最适温度的测定为在柠檬酸-磷酸氢二钠缓冲液(pH6.0)缓冲液体系及不同温度下进行酶促反应。耐温性测定为谷氨酸脱羧酶在不同温度下处理不同时间,再在40℃下进行酶活性测定。酶反应最适温度测定结果(图3)表明, 其最适温度为40℃。重组酶在35-45℃时稳定性非常好。
3、谷氨酸脱羧酶的
用不同浓度的l-Glu为底物,在pH5.0的l-Glu和柠檬酸-磷酸氢二钠缓冲液的混合体系中,在40℃下测定酶活性,计算出
表1. 谷氨酸脱羧酶GAD-Z11的动力学常数
4、转化率试验
人如图4所示,将携带
实施例5 重组谷氨酸脱羧酶GADZ20的性质测定
谷氨酸脱羧酶GADZ20,其氨基酸序列如SEQ ID NO. 5所示。如图5所示,与本发明的谷氨酸脱羧酶GADZ11的氨基酸序列一致性为96.32%,近20个氨基酸的差显著影响了两种谷氨酸脱羧酶的性质。
该谷氨酸脱羧酶GADZ20的最适pH为5.0,在pH4.0-5.5的范围内,酶活性均维持在最大酶活性的60%以上。然而,利用大肠杆菌异源表达并纯化后,比活为13.5 U/mg,显著低于本发明的谷氨酸脱羧酶GADZ11的比活91.7 U/mg。
谷氨酸脱羧酶GADZ20的的
用不同浓度的l-Glu为底物,在pH5.0的l-Glu和柠檬酸-磷酸氢二钠缓冲液的混合体系中,在40℃下测定酶活性,计算出
表2. 谷氨酸脱羧酶GADZ20的动力学常数
由此可见,谷氨酸脱羧酶GADZ11与谷氨酸脱羧酶GADZ20与底物的结合特性差异显著。尽管谷氨酸脱羧酶GADZ11的
序列表
<110> 中国农业科学院北京畜牧兽医研究所
<120> 一种高产γ-氨基丁酸的谷氨酸脱羧酶GADZ11及其基因和应用
<160> 5
<170> SIPOSequenceListing 1.0
<210> 1
<211> 489
<212> PRT
<213> 枯草芽孢杆菌(Bacillus)
<400> 1
Met Ser Lys Asp Arg Lys Ala Asp Thr Gln Ser Asp Thr Tyr Gly Gly
1 5 10 15
Lys Glu Ile Lys Pro Asp Gln Gln Gln Arg Leu Pro His His Met Glu
20 25 30
Met Glu Leu Pro His Glu Leu Ser Ile Asn Pro Leu Phe Ala Arg Glu
35 40 45
Gly Glu Ser Thr Ile Pro Arg Phe His Leu Ala Asp Gly Gly Met Leu
50 55 60
Pro Glu Thr Ala Tyr Gln Ile Val His Asp Glu Ile Thr Leu Asp Gly
65 70 75 80
Asn Ala Arg Leu Asn Leu Ala Thr Phe Val Ser Thr Trp Met Glu Pro
85 90 95
Ala Ala Glu Arg Leu Tyr Ala Gln Ser Phe Asp Lys Asn Met Ile Asp
100 105 110
Lys Asp Glu Tyr Pro Gln Thr Ala Gln Ile Glu Glu Arg Cys Val Arg
115 120 125
Ile Leu Ala Asp Leu Trp His Ser Pro Glu Pro Leu Lys Thr Ile Gly
130 135 140
Val Ser Thr Thr Gly Ser Ser Glu Ala Cys Met Leu Gly Gly Leu Ala
145 150 155 160
Leu Lys Arg Arg Trp Gln Asn Ala Arg Lys Lys Lys Gly Lys Pro Val
165 170 175
Asp Arg Pro Asn Ile Val Phe Ser Ser Ala Val Gln Val Val Trp Glu
180 185 190
Lys Phe Ala Asn Tyr Trp Glu Val Glu Pro Arg Tyr Val Lys Val Thr
195 200 205
Pro Glu His Pro Gln Leu Asn Pro Glu Gly Val Leu Ala Ala Val Asp
210 215 220
Glu Asn Thr Ile Gly Val Val Ala Ile Leu Gly Glu Thr Tyr Thr Gly
225 230 235 240
Leu Tyr Glu Pro Ile Ala Ala Ile Ala Lys Ala Leu Asp Glu Leu Gln
245 250 255
Glu Lys Ser Gly Leu Asn Ile Pro Met His Val Asp Ala Ala Ser Gly
260 265 270
Gly Phe Ile Ala Pro Phe Leu Gln Pro Asp Leu Val Trp Asp Phe Gln
275 280 285
Leu Pro Arg Val Lys Ser Ile Asn Val Ser Gly His Lys Tyr Gly Leu
290 295 300
Val Tyr Pro Gly Leu Gly Trp Ile Ile Trp Arg Glu Ala Lys Asp Leu
305 310 315 320
Pro Glu Glu Leu Ile Phe Arg Val Ser Tyr Leu Gly Gly Asn Met Pro
325 330 335
Thr Phe Ala Leu Asn Phe Ser Arg Pro Gly Ala Gln Val Leu Leu Gln
340 345 350
Tyr Tyr Asn Tyr Leu Arg Leu Gly Lys Glu Gly Tyr Tyr Glu Val Gln
355 360 365
Lys Ala Ser Gln Asn Val Ala Leu Phe Leu Ser Lys Glu Ile Gln Asn
370 375 380
Met Gly Pro Phe Glu Leu Leu Ser Asn Gly Ser Asp Ile Pro Val Phe
385 390 395 400
Ala Trp Arg Leu Lys Asp Asp Ala Thr Ser His Trp Thr Leu Phe Asp
405 410 415
Leu Ser Arg Gln Met Arg Val Phe Gly Trp Gln Val Pro Ala Tyr Pro
420 425 430
Leu Pro Pro Ala Met Glu Thr Val Thr Ile Met Arg Val Val Val Arg
435 440 445
Asn Gly Phe Ser Met Asp Leu Ala His Leu Phe Leu Val Asn Leu Lys
450 455 460
Gln Ala Val Ala Phe Leu Asp Thr Leu Asp Ala Pro Met Ala His Asp
465 470 475 480
Thr Lys Tyr Asp Asn Gly Phe His His
485
<210> 2
<211> 1470
<212> DNA
<213> 枯草芽孢杆菌(Bacillus)
<400> 2
atgtccaagg atcgaaaagc agacactcaa agcgatacat acggaggaaa agagatcaag 60
cccgatcagc aacaacgatt accgcatcac atggaaatgg agcttccaca cgaactctcc 120
ataaaccctc tgtttgcccg cgaaggtgaa tccaccattc cccgctttca tctggcggat 180
ggggggatgc taccagaaac agcctatcag atcgtccatg atgaaatcac tcttgatggc 240
aatgcacgct tgaacctcgc tacattcgtc agcacgtgga tggagcctgc cgcagagcgt 300
ttatatgccc aatcattcga taaaaacatg atagacaagg atgaatatcc gcagacagcc 360
caaatcgagg agagatgcgt ccgcatttta gccgatctct ggcattcacc cgagcctttg 420
aagacgatag gggtttccac aacggggtcg tctgaagcct gtatgcttgg aggactggcg 480
ttgaaaaggc gctggcaaaa cgcacgcaaa aagaaaggga aaccggtaga tcgccccaat 540
atcgtgttta gttctgctgt tcaggtcgtt tgggaaaaat tcgcgaacta ttgggaagtg 600
gaacctcgct atgtaaaggt caccccggag catccccaat tgaatcctga gggagtcctc 660
gctgcagtgg atgaaaatac gatcggggta gtcgccattc ttggcgaaac gtataccggt 720
ctttatgaac ccatagctgc cattgccaaa gcactggacg agctgcagga gaaatccgga 780
ctcaacattc cgatgcatgt ggatgcagcg tcagggggat ttatcgcacc gttccttcaa 840
cctgacctgg tttgggattt ccaactgccg agggtgaagt ccatcaatgt atccggacat 900
aagtatggat tggtctaccc gggattgggt tggatcatct ggagggaagc caaggacctc 960
cctgaggaac tcatcttccg cgtctcctat ctcggcggga acatgccgac tttcgccctg 1020
aatttctccc gccccggcgc gcaggtactg ctgcagtatt ataattacct gcgtctaggg 1080
aaagaggggt actatgaggt ccaaaaggct tcacagaacg ttgccctctt cctgagcaag 1140
gagattcaga acatgggacc gttcgaactt ttatccaatg gttccgatat acccgtcttc 1200
gcttggcgac tgaaagatga tgccacatcc cattggacgc tatttgattt atccagacaa 1260
atgcgtgtgt tcggctggca ggtccccgcc tatccattgc ctcctgcgat ggaaacggtg 1320
acgatcatgc gggtcgtggt ccggaatggt ttctccatgg accttgccca tttatttttg 1380
gtgaacctca aacaggccgt tgcttttctc gatacactgg atgcacctat ggcacatgat 1440
acgaagtatg ataatgggtt tcatcattag 1470
<210> 3
<211> 30
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 3
ctgaattcat gtccaaggat cgaaaagcag 30
<210> 4
<211> 38
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 4
ttcgccggcg aagcggccgc ctaatgatga aacccatt 38
<210> 5
<211> 489
<212> PRT
<213> 枯草芽孢杆菌(Bacillus)
<400> 5
Met Ser Lys Asp Arg Lys Ala Asp Arg Gln Asn Asp Thr Tyr Gly Gly
1 5 10 15
Lys Glu Ile Lys Pro Asp Gln Gln Gln Arg Leu Pro His Leu Met Glu
20 25 30
Met Glu Leu Pro His Glu Leu Ser Ile Asn Pro Leu Phe Ala Arg Glu
35 40 45
Gly Glu Ser Thr Val Pro Arg Phe His Leu Ala Asp Gly Gly Met Leu
50 55 60
Pro Glu Thr Ala Tyr Gln Ile Val His Asp Glu Ile Thr Leu Asp Gly
65 70 75 80
Asn Ala Arg Leu Asn Leu Ala Thr Phe Val Ser Thr Trp Met Glu Pro
85 90 95
Ser Ala Glu Arg Leu Tyr Ala Gln Ser Phe Asp Lys Asn Met Ile Asp
100 105 110
Lys Asp Glu Tyr Pro Gln Thr Ala Gln Ile Glu Glu Arg Cys Val Arg
115 120 125
Ile Leu Ala Asp Leu Trp His Ser Pro Glu Pro Leu Lys Thr Met Gly
130 135 140
Val Ser Thr Thr Gly Ser Ser Glu Ala Cys Met Leu Gly Gly Leu Ala
145 150 155 160
Leu Lys Arg Arg Trp Gln Asn Ala Arg Lys Lys Glu Gly Lys Pro Thr
165 170 175
Asp Arg Pro Asn Ile Val Phe Ser Ser Ala Val Gln Val Val Trp Glu
180 185 190
Lys Phe Ala Asn Tyr Trp Glu Val Glu Pro Arg Tyr Val Lys Val Thr
195 200 205
His Glu His Pro Gln Leu Asn Pro Glu Gly Val Leu Ala Ala Val Asp
210 215 220
Glu Asn Thr Ile Gly Val Val Ala Ile Leu Gly Glu Thr Tyr Thr Gly
225 230 235 240
Leu Tyr Glu Pro Ile Val Ser Ile Ala Lys Ala Leu Asp Asp Leu Gln
245 250 255
Glu Lys Ser Gly Leu Asn Ile Pro Met His Val Asp Ala Ala Ser Gly
260 265 270
Gly Phe Ile Ala Pro Phe Leu Gln Pro Asp Leu Val Trp Asp Phe Gln
275 280 285
Leu Pro Arg Val Lys Ser Ile Asn Val Ser Gly His Lys Tyr Gly Leu
290 295 300
Val Tyr Pro Gly Leu Gly Trp Ile Ile Trp Arg Glu Ala Lys Asp Leu
305 310 315 320
Pro Glu Glu Leu Ile Phe Arg Val Ser Tyr Leu Gly Gly Asn Met Pro
325 330 335
Thr Phe Ala Leu Asn Phe Ser Arg Pro Gly Ala Gln Val Leu Leu Gln
340 345 350
Tyr Tyr Asn Tyr Leu Arg Leu Gly Lys Glu Gly Tyr Phe Glu Val Gln
355 360 365
Lys Ala Ser Gln Asn Val Ala Leu Phe Leu Ser Lys Glu Ile Gln Asn
370 375 380
Met Gly Pro Phe Glu Leu Leu Ser Asp Gly Ser Asp Ile Pro Val Phe
385 390 395 400
Ala Trp Arg Leu Lys Glu Asp Asp Thr Ser His Trp Thr Leu Phe Asp
405 410 415
Leu Ser Arg Gln Met Arg Val Phe Gly Trp Gln Val Pro Ala Tyr Pro
420 425 430
Leu Pro Pro Asp Met Glu Thr Val Thr Ile Met Arg Val Val Val Arg
435 440 445
Asn Gly Phe Ser Met Asp Leu Ala His Leu Phe Leu Val Asn Leu Lys
450 455 460
Gln Ala Val Ala Phe Leu Asp Thr Leu Asp Ala Pro Met Pro His Asp
465 470 475 480
Thr Lys Tyr Asp Asn Gly Phe His His
485
- 一种高产γ-氨基丁酸的谷氨酸脱羧酶GADZ11及其基因和应用
- 一种高产γ-氨基丁酸的谷氨酸脱羧酶GADZ1及其基因和应用