用于蛋白质测序的方法及组合物
文献发布时间:2023-06-19 12:16:29
相关申请的交叉引用
本申请依据35 U.S.C.§119(e)主张2019年9月27日申请的美国临时专利申请第62/907,507号,及2018年11月15日申请的美国临时专利申请第62/768,076号的优先权,这些申请中的每一个以全文引用的方式并入本文中。
背景技术
在生物系统研究中,蛋白质组学已成为基因组学及转录组学的重要且必要补充。对单个生物体的蛋白质组学分析可提供深入细胞过程及反应模式的洞察,其使诊断及治疗策略得到改良。围绕蛋白质结构、组成及修饰的复杂性为测定生物样品的大规模蛋白质测序信息带来挑战。
发明内容
在一些方面中,本申请提供了用于从多肽测定氨基酸序列信息(例如,用于对一种或多种多肽进行测序)的方法及组合物。在一些实施方案中,可针对单一多肽分子测定氨基酸序列信息。在一些实施方案中,测定多肽中两个或更多个氨基酸的相对位置,例如针对单一多肽分子进行测定。在一些实施方案中,标记多肽的一个或多个氨基酸(例如,直接地或间接地),并测定多肽中带标记的氨基酸的相对位置。
在一些方面中,本申请提供了方法,其包括在多肽降解过程中获得数据。在一些实施方案中,方法进一步包括分析数据,以确定对应于以下的数据部分:在降解过程中在多肽的末端依次暴露的氨基酸。在一些实施方案中,方法进一步包括输出代表多肽的氨基酸序列。在一些实施方案中,数据指示在降解过程中多肽的末端的氨基酸身分。在一些实施方案中,数据指示在降解过程中由一种或多种氨基酸识别分子与末端的不同类型的末端氨基酸结合所产生的信号。在一些实施方案中,数据指示在降解过程中产生的发光信号。在一些实施方案中,数据指示在降解过程中产生的电信号。
在一些实施方案中,分析数据进一步包括检测裂解事件系列,及确定连续裂解事件之间的数据部分。在一些实施方案中,分析数据进一步包括针对单独部分中的每一个,确定一种类型的氨基酸。在一些实施方案中,单独部分中的每一个包括脉冲图案(例如,特征图案),且分析数据进一步包括针对这些部分中的一个或多个,基于其各自的脉冲图案确定一种类型的氨基酸。在一些实施方案中,确定氨基酸的类型进一步包括在数据超过阈值时,鉴定一部分内的时间量,及将该时间量与该部分的持续时间进行比较。在一些实施方案中,确定氨基酸的类型进一步包括针对一个或多个部分中的每一个,鉴定至少一个脉冲持续时间。在一些实施方案中,确定氨基酸的类型进一步包括针对一个或多个部分中的每一个,鉴定至少一个脉冲间(interpulse)持续时间。在一些实施方案中,氨基酸序列包括对应于这些部分的氨基酸系列。
在一些方面中,本申请提供了系统,其包括至少一种硬件处理器,及至少一种储存处理器可执行指令的非暂时性电脑可读储存介质,当由该至少一种硬件处理器执行时,这些处理器可执行指令使该至少一种硬件处理器进行根据本申请的方法。在一些方面中,本申请提供了至少一种储存处理器可执行指令的非暂时性电脑可读储存介质,当由至少一种硬件处理器执行时,这些处理器可执行指令使该至少一种硬件处理器进行根据本申请的方法。
在一些方面中,本申请提供了多肽测序的方法。在一些实施方案中,方法包括使单一多肽分子与一种或多种末端氨基酸识别分子接触。在一些实施方案中,方法进一步包括检测信号脉冲系列,该系列指示一种或多种末端氨基酸识别分子与单一多肽分子降解时在单一多肽分子的末端暴露的连续氨基酸的缔合,从而获得关于单一多肽分子的序列信息。在一些实施方案中,测定单一多肽分子的大部分或全部的氨基酸序列。在一些实施方案中,信号脉冲系列为实时信号脉冲系列。
在一些实施方案中,一种或多种末端氨基酸识别分子与在末端暴露的各类型的氨基酸的缔合在信号脉冲系列中产生特征图案,所述特征图案不同于在末端暴露的其他类型的氨基酸。在一些实施方案中,该特征图案的信号脉冲对应于末端氨基酸识别分子与在末端暴露的氨基酸之间的单独缔合事件。在一些实施方案中,特征图案对应于与在单一多肽分子的末端暴露的氨基酸的可逆末端氨基酸识别分子结合相互作用系列。在一些实施方案中,特征图案指示在单一多肽分子的末端暴露的氨基酸,及相邻位置处的氨基酸(例如,相同类型或不同类型的氨基酸)。
在一些实施方案中,单一多肽分子由裂解试剂降解,该裂解试剂将一个或多个氨基酸从单一多肽分子末端移除。在一些实施方案中,方法进一步包括检测指示裂解试剂与末端缔合的信号。在一些实施方案中,裂解试剂包括可检测标记(例如,发光标记、导电性标记)。在一些实施方案中,单一多肽分子被固定至表面。在一些实施方案中,单一多肽分子通过远离一种或多种末端氨基酸识别分子所缔合的末端的末端被固定至表面。在一些实施方案中,单一多肽分子通过连接子(例如,包括生物分子的增溶连接子)被固定至表面。
在一些方面中,本申请提供了对多肽进行测序的方法,这些方法包括使单一多肽分子在反应混合物中与包括一种或多种末端氨基酸识别分子及裂解试剂的组合物接触。在一些实施方案中,方法进一步包括在裂解试剂存在下,检测指示一种或多种末端氨基酸识别分子与单一多肽分子的末端缔合的信号脉冲系列。在一些实施方案中,信号脉冲系列指示由于末端氨基酸被裂解试剂裂解而导致的随时间推移在末端暴露的氨基酸系列。
在一些方面中,本申请提供了对多肽进行测序的方法,其包括(a)鉴定单一多肽分子的末端的第一氨基酸,(b)移除第一氨基酸以暴露单一多肽分子的末端的第二氨基酸,及(c)鉴定单一多肽分子的末端的第二氨基酸。在一些实施方案中,(a)至(c)在单一反应混合物中进行。在一些实施方案中,(a)至(c)依次发生。在一些实施方案中,(c)在(a)及(b)之前发生。在一些实施方案中,单一反应混合物包括一种或多种末端氨基酸识别分子。在一些实施方案中,单一反应混合物包括裂解试剂。在一些实施方案中,第一氨基酸由裂解试剂移除。在一些实施方案中,方法进一步包括重复以下步骤:移除及鉴定单一多肽分子的末端的一个或多个氨基酸,从而确定单一多肽分子的序列(例如,部分序列或完整序列)。
在一些方面中,本申请提供了鉴定多肽的氨基酸的方法,其包括使单一多肽分子与结合单一多肽分子的一种或多种氨基酸识别分子接触。在一些实施方案中,方法进一步包括在多肽降解条件下,检测指示一种或多种氨基酸识别分子与单一多肽分子缔合的信号脉冲系列。在一些实施方案中,方法进一步包括基于信号脉冲系列中的第一特征图案,鉴定单一多肽分子中第一类型的氨基酸。
在一些方面中,本申请提供了鉴定多肽的末端氨基酸(例如,N末端或C末端氨基酸)的方法。在一些实施方案中,方法包括使多肽与一种或多种带标记的亲和试剂(例如,一种或多种氨基酸识别分子)接触,所述一种或多种带标记的亲和试剂在多肽的末端选择性结合一种或多种类型的末端氨基酸。在一些实施方案中,方法进一步包括通过检测多肽与一种或多种带标记的亲和试剂的相互作用,鉴定多肽末端的末端氨基酸。
在其他方面中,本申请提供了通过艾德曼(Edman)型降解反应进行的多肽测序的方法。在一些实施方案中,出于检测或裂解的目的,艾德曼型降解反应可通过使多肽与不同反应混合物接触进行(例如,相比于可涉及使用单一反应混合物进行检测及裂解的动态测序反应)。
因此,在一些方面中,本申请提供了确定多肽的氨基酸序列的方法,其包括(i)使多肽与一种或多种带标记的亲和试剂接触,所述一种或多种试剂在多肽的末端选择性地结合一种或多种类型的末端氨基酸。在一些实施方案中,方法进一步包括(ii)通过检测多肽与一种或多种带标记的亲和试剂的相互作用,鉴定多肽末端的末端氨基酸(例如,N末端或C末端氨基酸)。在一些实施方案中,方法进一步包括(iii)移除末端氨基酸。在一些实施方案中,方法进一步包括(iv)在多肽的末端重复(i)至(iii)一次或多次,以确定多肽的氨基酸序列。
在一些实施方案中,方法进一步包括在(i)之后且(ii)之前,移除不选择性结合末端氨基酸的一种或多种带标记的亲和试剂中的任一种。在一些实施方案中,方法进一步包括在(ii)之后且(iii)之前,移除选择性结合末端氨基酸的一种或多种带标记的亲和试剂中的任一种。
在一些实施方案中,移除末端氨基酸(例如,(iii))包括通过使末端氨基酸与异硫氰酸酯(例如,异硫氰酸苯酯)接触来修饰末端氨基酸,以及使被修饰的末端氨基酸与特异性结合并移除被修饰的末端氨基酸的蛋白酶接触。在一些实施方案中,裂解末端氨基酸(例如,(iii))包括通过使末端氨基酸与异硫氰酸酯接触来修饰末端氨基酸,以及使被修饰的末端氨基酸经历足以移除被修饰的末端氨基酸的酸性或碱性条件。
在一些实施方案中,鉴定末端氨基酸包括将末端氨基酸鉴定为一种或多种带标记的亲和试剂所结合的一种或多种类型的末端氨基酸中的一种类型。在一些实施方案中,鉴定末端氨基酸包括将末端氨基酸鉴定为除一种或多种带标记的亲和试剂所结合的一种或多种类型的末端氨基酸外的类型。
在一些方面中,本申请提供了氨基酸识别分子,其包括屏蔽元件,例如用于增强多肽测序反应中的光稳定性。在一些方面中,本申请提供了式(I)的氨基酸识别分子:
A-(Y)
(I),
其中:A是包括至少一种氨基酸识别分子的氨基酸结合组分;Y在各情况下是形成共价或非共价连接基团的聚合物;n是1至10的整数(包括端点);且D是包括至少一种可检测标记的标记组分。在一些实施方案中,D的直径小于
在一些方面中,本申请提供了式(II)的氨基酸识别分子:
A-Y
(II),
其中:A是包括至少一种氨基酸识别分子的氨基酸结合组分;Y
在一些方面中,本申请提供了氨基酸识别分子,其包括:核酸;连接至核酸上的第一连接位点的至少一种氨基酸识别分子;及连接至核酸上的第二连接位点的至少一种可检测标记,其中核酸在至少一种氨基酸识别分子与至少一种可检测标记之间形成共价或非共价连接基团。在一些实施方案中,核酸包括第一寡核苷酸链。在一些实施方案中,核酸进一步包括与第一寡核苷酸链杂交的第二寡核苷酸链。
在一些方面中,本申请提供了氨基酸识别分子,其包括:多价蛋白,其包括至少两个配体结合位点;至少一种氨基酸识别分子,其通过与蛋白质上的第一配体结合位点结合的第一配体部分连接至蛋白质;及至少一种可检测标记,其通过与蛋白质上的第二配体结合位点结合的第二配体部分连接至蛋白质。在一些实施方案中,多价蛋白为抗生物素蛋白(avidin protein)。
在一些实施方案中,被屏蔽的氨基酸识别分子可用于根据本申请的多肽测序方法,或本领域中已知的任何方法。因此,在一些方面中,本申请提供了多肽测序的方法(例如,在艾德曼型降解反应中、在动态测序反应中或本领域中已知的其他方法),其包括使多肽分子与本申请的一种或多种被屏蔽的氨基酸识别分子接触。举例而言,在一些实施方案中,方法包括使多肽分子与包括根据本申请的屏蔽物或屏蔽元件的至少一种氨基酸识别分子接触,及检测至少一种氨基酸识别分子与多肽分子的缔合。
在一些方面中,本申请提供了在混合样品中鉴定所关注蛋白质的方法。在一些实施方案中,方法包括裂解混合蛋白质样品以产生多个多肽片段。在一些实施方案中,方法进一步包括在根据本申请的方法的方法中,确定多个多肽片段中的至少一个多肽片段的氨基酸序列。在一些实施方案中,方法进一步包括若所关注蛋白质可唯一地鉴定氨基酸序列,则在混合样品中鉴定所关注蛋白质。
在一些实施方案中,鉴定混合样品中的所关注蛋白质的方法包括裂解混合蛋白质样品以产生多个多肽片段。在一些实施方案中,方法进一步包括用一种或多种不同发光标记标记多个多肽片段中的一种或多种类型的氨基酸。在一些实施方案中,方法进一步包括针对多个带标记的多肽中的至少一个带标记的多肽,随时间推移测量发光。在一些实施方案中,方法进一步包括基于所检测到的发光确定至少一个带标记的多肽的氨基酸序列。在一些实施方案中,方法进一步包括若所关注蛋白质可唯一地鉴定氨基酸序列,则在混合样品中鉴定所关注蛋白质。
因此,在一些实施方案中,待根据本申请分析的多肽分子或所关注蛋白质可为混合或纯化样品的形式。在一些实施方案中,多肽分子或所关注蛋白质从生物样品(例如,血液、组织、唾液、尿液或其他生物来源)获得。在一些实施方案中,多肽分子或所关注蛋白质从患者样品(例如,人类样品)获得。
本发明的某些实施方案的细节阐述于如下文所描述的某些实施方案的实施方式中。本发明的其他特征、目标及优点将根据定义、实施例、图式及申请专利范围而显而易见。
附图说明
本领域技术人员本领域技术人员将理解,本文所描述的图仅出于说明的目的。应理解,在一些情况下,为辅助理解本发明,本发明的各种方面可能被放大或扩大展示。在图式中,在通篇各种图中,相同参考标号一般是指相同特征、功能上类似和/或结构上类似的元件。图式未必为按比例的,而实际上重点在于说明教示内容的原理。图式不意欲以任何方式限制本教示内容的范畴。
本发明的特征及优点将从下文结合图式所阐述的实施方式而变得更显而易见。
当参考图式描述实施例时,可使用方向参考(“之上”、“之下”、“顶部”、“底部”、“左侧”、“右侧”、“水平”、“垂直”等)。此类参考仅意欲作为读者在正常定向上观看图式的辅助。这些方向参考不意欲描述实施装置的较佳或唯一定向。装置可以其他定向实施。
如根据实施方式显而易见,在本申请通篇,图中描绘及出于说明的目的进一步描述的实施例描述非限制性实例,且在一些情况下可出于更清楚说明的目的简化某些方法或省略特征或步骤。
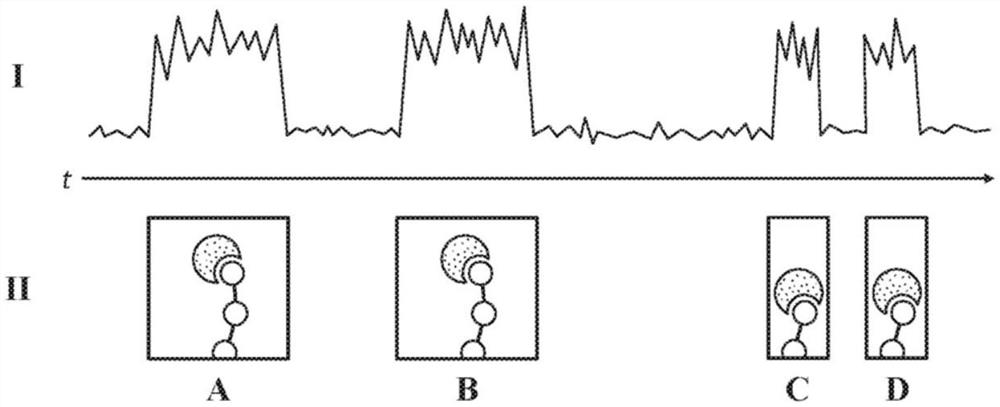
图1A至图1B展示了通过单分子结合相互作用的检测(图1A)及分析(图1B)进行多肽测序的实施例。
图1C至图1E展示了根据本申请的带标记的亲和试剂及使用方法的各种实施例。图1C描绘带标记的亲和试剂的示例性构造,包括选择性结合一种或多种类型的末端氨基酸的带标记的酶及带标记的适体。图1D大体上描绘使用带标记的亲和试剂进行的基于降解的多肽测序过程。图1E展示了通过末端氨基酸检测、修饰及裂解的重复循环,使用带标记的适体进行多肽测序的实施例。
图2展示了使用各自选择性结合且裂解不同类型的末端氨基酸的带标记的外肽酶进行实时多肽测序的实施例。
图3A至图3B展示了通过评估末端和/或内部氨基酸与带标记的亲和试剂及带标记的裂解试剂(例如,带标记的非特异性外肽酶)的结合相互作用,进行实时多肽测序的实施例。图3A展示通过检测信号输出中的脉冲系列,进行实时测序的实施例。图3B示意性地描绘温度依赖性测序过程。
图4展示了通过评估末端及内部氨基酸与带标记的亲和试剂及带标记的非特异性外肽酶的结合相互作用,进行实时多肽测序的实施例。
图5A至图5E展示了被屏蔽元件标记的亲和试剂的非限制性实例。图5A示出了用亲和试剂进行的单分子肽测序,该亲和试剂通过常规共价连接标记。图5B示出了用亲和试剂进行的单分子肽测序,该亲和试剂包括屏蔽元件。图5C至图5E示出了根据本申请的屏蔽元件的各种实施例。
图6展示了基于带标记的多肽中所检测到的氨基酸的独特组合来鉴定多肽的实施例。
图7展示了通过检测经历末端氨基酸修饰及裂解的重复循环的带标记的多肽的发光,进行多肽测序的实施例。
图8A至图8C展示了通过带标记的多肽的进行性(processive)酶裂解,进行多肽测序的实施例。图8A展示了通过固定化末端肽酶进行性酶裂解带标记的多肽来进行测序的实施例。图8B展示了通过末端肽酶进行性酶裂解固定的带标记的多肽来进行测序的实施例。图8C示意性地示出了根据图8B进行的实时测序过程的实施例。
图9示意性地示出了使用固定化ATP依赖型蛋白酶、供体标记的ATP及多肽底物的受体标记的氨基酸,通过基于辅因子的FRET进行测序的实施例。
图10A至图10C展示了针对根据本申请的多肽及蛋白质的分析,制备样品及样品孔表面的各种实施例。图10A大体上描绘了由蛋白质样品制备末端修饰的多肽的示例性过程。图10B大体上描绘了将增溶连接子缀合至多肽的示例性过程。图10C展示了具有可用于促进单分子固定至底表面的被修饰的表面的样品孔的示例性示意图。
图11为根据本文所描述的技术的一些实施方案,用于分析在多肽降解过程中获得的数据的说明性序列数据处理流水线的图解。
图12为根据本文所描述的技术的一些实施方案,用于确定多肽分子的氨基酸序列的说明性过程的流程图。
图13为根据本文所描述的技术的一些实施方案,用于确定代表多肽的氨基酸序列的说明性过程的流程图。
图14为可用于实施本文所描述的技术的一些实施方案的说明性电脑系统的方块图。
图15A至图15C展示了针对由不同增溶连接子提供的提高溶解度制备及评估的所选肽-连接子缀合物的实验数据。图15A展示了被合成及评估的肽-连接子缀合物的示例性结构。图15B展示了来自LCMS的结果,其展现N末端的肽裂解。图15C展示了来自负载实验的结果。
图16展示了基于实验结果对所选外肽酶的氨基酸裂解活性的概述。
图17A至图17C展示了用于检测及裂解末端氨基酸的染料/肽共轭物分析的实验数据。图17A展示了用于进行染料/肽共轭物分析的示例性方案及结构。图17B展示在芯片上分析中负载至样品孔中的肽-连接子缀合物的成像结果。图17C展示检测肽-缀合物负载及末端氨基酸裂解的示例性信号迹线。
图18A至图18F展示了用于检测及裂解末端氨基酸的FRET染料/肽缀合物分析的实验数据。图18A展示了用于进行FRET染料/肽缀合物分析的示例性方案及结构。图18B展示了不同时间点的FRET成像结果。图18C展示了不同时间点处的剪切效率。图18D展示了不同时间点中的每一个处所显示的剪切。图18E展示用来自鼠疫耶氏杆菌(Yersinia pestis)的脯氨酸亚胺基肽酶(yPIP)进行的不同时间点的额外FRET成像结果。图18F展示了用来自解蛋白弧菌(Vibrio proteolyticus)的氨肽酶(VPr)进行的不同时间点的FRET成像结果。
图19A至图19H展示了由带标记的亲和试剂进行末端氨基酸区分的实验数据。图19A展示针对这些实验标记的ClpS2蛋白的晶体结构。图19B展示了单分子强度迹线,其说明由带标记的ClpS2蛋白进行的N末端氨基酸区分。图19C为展示不同末端氨基酸的平均脉冲持续时间的曲线图。图19D为展示不同末端氨基酸的平均脉冲间持续时间的曲线图。图19E展示了进一步示出不同末端氨基酸当中判别脉冲持续时间的曲线图。图19F、图19G及图19H展示了来自表明亮氨酸由来自细长嗜热聚球藻(Thermosynochoccus elongatus)的ClpS蛋白(teClpS)识别的停留时间分析的示例性结果。图19I展示来自表明苯丙氨酸、亮氨酸、色氨酸及酪氨酸由根癌农杆菌(A.tumefaciens)ClpS1可区分识别的停留时间分析的示例性结果。图19J展示了来自停留时间分析的示例性结果,其展现由细长聚球藻(S.elongatus)ClpS2进行的亮氨酸识别。图19K至图19L展示了来自停留时间分析的示例性结果,其展现由GID4进行的脯氨酸识别。
图20A至图20D展示了来自使用同一反应混合物中的带标记的ClpS2识别蛋白及氨肽酶裂解试剂,实时进行多肽测序反应的示例性结果。图20A展示了第一测序反应的信号迹线数据。图20B展示了图20A中所展示的信号迹线数据的脉冲持续时间统计数据。图20C展示了第二测序反应的信号迹线数据。图20D展示了图20C中所展示的信号迹线数据的脉冲持续时间统计数据。
图21A至图21F展示了由带标记的外肽酶进行的末端氨基酸鉴定及裂解的实验数据。图21A展示了针对这些实验位点特异性标记的脯氨酸亚胺基肽酶(yPIP)的晶体结构。图21B展示了纯化蛋白质产品的标记程度。图21C为证实yPIP的位点特异性标记的SDS page的影像。图21D为证实位点特异性标记的SDS page凝胶的过度曝光影像。图21E为证实带标记的蛋白质产品的纯度的考马斯(Coomassie)染色凝胶的影像。图21F为表明带标记的外肽酶的裂解活性的HPLC迹线。序列YPYPYPK对应于SEQ ID NO:82。序列PYPYPK对应于SEQ ID NO:83。
图22A至图22F展示了来自评估含有特定翻译后修饰的氨基酸识别的实验的数据。图22A展示了表明磷酸化酪氨酸由含SH2结构域的蛋白质识别的代表性迹线;图22B展示了对应于图22A的迹线的脉冲持续时间数据;且图22C展示了针对迹线测定的统计数据。图22D至图22F展示了来自阴性对照实验的代表性迹线。
图23为展示了来自评估次末端(penultimate)氨基酸对脉冲持续时间的影响的实验的中值脉冲持续时间的曲线图。
图24A至图24C展示了来自评估由差异性标记的识别分子进行的同时氨基酸识别的实验的数据。图24A展示了代表性迹线。图24B为比较在这些实验期间针对各识别分子获得的脉冲持续时间数据的曲线图。图24C展示了这些实验的脉冲持续时间统计数据。
图25A至图25C展示了来自评估在单分子识别期间肽的光稳定性的实验的数据。图25A展示了来自使用距氨基酸结合位点约2nm用染料标记的atClpS2-V1进行识别的代表性迹线。图25B展示了这些实验中所使用的ClpS2蛋白结构的视觉化。图25C展示了来自使用距氨基酸结合位点>10nm经DNA/蛋白质连接子用染料标记的ClpS2进行识别的代表性迹线。
图26A至图26D展示了来自在氨肽酶裂解试剂存在下,使用被DNA/抗生物素蛋白链菌素(streptavidin)连接子标记的ClpS2识别蛋白,在互补金氧半导体(CMOS)芯片上实时进行多肽测序反应的代表性迹线。
图27展示了来自在掘越氏火球菌(Pyrococcus horikoshii)TET氨肽酶裂解试剂存在下,使用被DNA/抗生物素蛋白链菌素连接子标记的atClpS2-V1识别蛋白实时进行多肽测序反应的代表性迹线。
图28A至图28J展示了来自使用具有差异性裂解特异性的多个类型的外肽酶实时进行多肽测序反应的代表性迹线数据。图28A展示了来自用hTET外肽酶进行的反应的代表性迹线,扩展脉冲图案区域展示于图28B中。图28A中的序列YAAWAAFADDDWK对应于SEQ IDNO:78。图28C展示了来自用hTET及yPIP外肽酶两者进行的反应的代表性迹线,扩展脉冲图案区域展示于图28D中,且额外代表性迹线展示于图28E中。图28C中的序列FYPLPWPDDDYK对应于SEQ ID NO:80。图28F展示了来自用hTET及yPIP外肽酶两者进行的另一反应的代表性迹线,扩展脉冲图案区域展示于图28G中,且额外代表性迹线展示于图28H中。图28I展示了来自用PfuTET及yPIP外肽酶两者进行的反应的代表性迹线,扩展脉冲图案区域展示于图28J中。图28F及图28I中的序列YPLPWPDDDYK对应于SEQ ID NO:81。
具体实施方式
本申请的方面涉及蛋白质测序及鉴定的方法、多肽测序及鉴定的方法、氨基酸鉴定的方法及用于进行此类方法的组合物。
在一些方面中,本申请涉及多肽测序技术的发现,这些技术可使用现有分析仪器,几乎不进行或不进行装置修改即可实施。举例而言,先前的多肽测序策略涉及不同试剂混合物通过含有所分析的多肽的反应容器迭代循环。此类策略可能需要对现有分析仪器,诸如核酸测序仪器进行修改,现有分析仪器可能未配备有能够进行试剂循环的流槽或类似设备。诸位发明人已认识到且了解,本申请的某些多肽测序技术不需要迭代试剂循环,从而允许使用现有仪器,而不需进行可能增加仪器大小的大量修改。因此,在一些方面中,本申请提供了允许使用较小测序仪器的多肽测序方法。在一些方面中,本申请涉及多肽测序技术的发现,这些技术允许使用同一测序仪器进行基因组分析及蛋白质组分析。
诸位发明人已进一步认识到且了解,在多肽测序中,差异性结合相互作用可以为常规标记策略提供另外的或替代的方法。常规多肽测序可涉及用可唯一鉴定的标记标记各类型的氨基酸。此过程费力且容易出错,因为存在至少二十种不同类型的天然存在的氨基酸,此外还有其大量翻译后变化形式。在一些方面中,本申请涉及一些技术的发现,这些技术涉及使用与不同类型的氨基酸差异性缔合的氨基酸识别分子,以产生指示多肽的氨基酸序列的可检测的特征标志。因此,本申请的方面提供了一些技术,这些技术不需要某些常规多肽测序方法中所使用的多肽标记和/或苛刻化学试剂,从而提高了获自样品的序列信息的通量和/或准确度。
在一些方面中,本申请涉及以下发现,即,可仅使用单一反应混合物(例如,而不需要通过反应容器进行迭代试剂循环)实时监测多肽测序反应。如上文所详述,常规多肽测序反应可涉及将多肽暴露于不同试剂混合物,以在氨基酸检测与氨基酸裂解步骤之间循环。因此,在一些方面中,本申请涉及二代测序中的改进,允许在整个正在进行中的降解反应中通过氨基酸检测实时进行多肽分析。通过动态测序进行此类多肽分析的方法在下文描述。
如本文所描述,在一些方面中,本申请提供了通过以下对多肽进行测序的方法:在多肽降解过程中获得数据,及分析数据,以确定对应于以下的数据部分:在降解过程中在多肽的末端依次暴露的氨基酸。在一些实施方案中,这部分数据包括一系列信号脉冲,这一系列信号脉冲指示一种或多种氨基酸识别分子与多肽末端暴露的连续氨基酸的缔合(例如,在降解期间)。在一些实施方案中,这一系列信号脉冲对应于降解过程中多肽末端的一系列可逆的单分子结合相互作用。
通过检测多肽降解过程中的单分子结合相互作用进行多肽测序的非限制性实例在图1A中示意性地示出。示例性信号迹线(I)与图(II)系列一起展示,图(II)系列描绘了对应于信号变化时的不同缔合事件。如所展示,氨基酸识别分子(带点(stippled)形状)与多肽(展示为线珠结构(beads-on-a-string))的末端的氨基酸之间的缔合事件产生了信号幅度的变化,该变化持续一定持续时间。
图(A)及图(B)描绘了氨基酸识别分子与多肽末端暴露的第一氨基酸(例如,第一末端氨基酸)之间的不同缔合事件。各缔合事件产生了信号迹线(I)的变化,该变化的特征在于缔合事件的持续时间内持续存在的信号幅度的变化。因此,图(A)及图(B)的缔合事件之间的持续时间可对应于一个持续时间,在该持续时间内,多肽未与氨基酸识别分子可检测地缔合。
图(C)及图(D)描绘了氨基酸识别分子与多肽末端暴露的第二氨基酸(例如,第二末端氨基酸)之间的不同缔合事件。如本文所描述,在多肽的末端“暴露”的氨基酸是这样一种氨基酸,它仍连接至多肽,且在降解期间在移除先前的末端氨基酸(例如,单独或与一个或多个额外氨基酸一起)后变为末端氨基酸。因此,图(II)系列的第一及第二氨基酸提供了在多肽的末端暴露的连续氨基酸的说明性示例,其中第二氨基酸在第一氨基酸移除后变为末端氨基酸。
如大体上所描绘,图(C)及图(D)的缔合事件产生了信号迹线(I)的变化,这些变化的特征在于与图(A)及图(B)相比,幅度变化持续的持续时间相对较短,且与图(A)及图(B)相比,图(C)与图(D)的缔合事件之间的持续时间相对较短。如本文所描述,在一些实施方案中,这些独特信号变化中的一种或两种可用于确定信号迹线(I)中的特征图案,这些特征图案可区分不同类型的氨基酸。在一些实施方案中,从一种特征图案转变至另一特征图案指示氨基酸裂解。如本文所用,在一些实施方案中,氨基酸裂解是指从多肽的末端移除至少一个氨基酸(例如,从多肽移除至少一个末端氨基酸)。在一些实施方案中,氨基酸裂解通过基于特征图案之间的持续时间的推断来确定。在一些实施方案中,氨基酸裂解通过检测由带标记的裂解试剂与多肽末端的氨基酸的缔合产生的信号的变化来确定。随着在降解期间氨基酸从多肽的末端依次裂解,检测到幅度变化系列,或信号脉冲系列。在一些实施方案中,信号脉冲数据可如图1B中所示出来分析。
在一些实施方案中,可通过将阈值水平应用于信号数据的一个或多个参数,来分析信号数据,以提取信号脉冲信息。举例而言,图(III)描绘了阈值幅度水平(“M
提取出的信号脉冲信息展示于图(III)中,出于说明的目的,叠加了示例性信号迹线(I)。在一些实施方案中,信号脉冲的幅度的峰值通过对持续高于M
图(IV)展示了从示例性信号迹线(I)提取的信号脉冲信息。在一些实施方案中,可分析信号脉冲信息,以基于信号脉冲系列中的不同特征图案来鉴定序列中不同类型的氨基酸。举例而言,如图(IV)中所展示,信号脉冲信息基于第一特征图案(“CP
亦如图(IV)中所展示,各信号脉冲包括脉冲持续时间(“pd”),其对应于氨基酸识别分子与具有特征图案的氨基酸之间的缔合事件。在一些实施方案中,脉冲持续时间为结合的解离速率的特征。亦如所展示,特征图案的各信号脉冲由脉冲间持续时间(“ipd”)与特征图案的另一信号脉冲间隔开。在一些实施方案中,脉冲间持续时间为结合的缔合速率的特征。在一些实施方案中,可基于基线与信号脉冲的峰值之间的差值来确定信号脉冲的幅度变化(“ΔM”)。在一些实施方案中,基于脉冲持续时间来确定特征图案。在一些实施方案中,基于脉冲持续时间及脉冲间持续时间来确定特征图案。在一些实施方案中,基于脉冲持续时间、脉冲间持续时间及幅度变化中的任何一种或多种来确定特征图案。
因此,如由图1A至图1B所示出,在一些实施方案中,通过在进行中的降解反应中,检测指示一种或多种氨基酸识别分子与多肽末端暴露的连续氨基酸的缔合的信号脉冲系列,进行多肽测序。可分析信号脉冲系列,以确定信号脉冲系列中的特征图案,且特征图案的时程可用于确定多肽的氨基酸序列。
在一些实施方案中,信号脉冲系列包括光信号幅度随时间推移的变化系列。在一些实施方案中,光信号的变化系列包括在缔合事件期间所产生的发光的变化系列。在一些实施方案中,发光由与测序反应的一种或多种试剂缔合的可检测标记产生。举例而言,在一些实施方案中,一种或多种氨基酸识别分子中的每一个包括发光标记。在一些实施方案中,裂解试剂包括发光标记。根据本申请的发光标记及其用途的实例提供于本文别处。
在一些实施方案中,信号脉冲系列包括电信号幅度随时间推移的变化系列。在一些实施方案中,电信号的变化系列包括在缔合事件期间产生的电导的变化系列。在一些实施方案中,导电性由与测序反应的一种或多种试剂缔合的可检测标记产生。举例而言,在一些实施方案中,一种或多种氨基酸识别分子中的每一个包括导电性标记。根据本申请的导电性标记及其用途的实例提供于本文别处。已描述了使用导电性标记来鉴定单一分子的方法(参见例如美国专利公开案第2017/0037462号)。
在一些实施方案中,电导的变化系列包括通过纳米孔的电导的变化系列。举例而言,已描述了使用纳米孔来评估受体-配体相互作用的方法(参见例如Thakur,A.K.及Movileanu,L.(2019)Nature Biotechnology 37(1))。诸位发明人已认识到且了解,此类纳米孔可用于监测根据本申请的多肽测序反应。因此,在一些实施方案中,本申请提供了多肽测序的方法,其包括使单一多肽分子与一种或多种氨基酸识别分子接触,其中单一多肽分子固定至纳米孔。在一些实施方案中,方法进一步包括检测通过纳米孔的电导的变化系列,从而对单一多肽分子进行测序,其中所述变化系列指示一种或多种末端氨基酸识别分子与单一多肽降解时在单一多肽的末端暴露的连续氨基酸的缔合。
在一些方面中,本申请提供了通过鉴定来自混合物的多肽的一种或多种类型的氨基酸,对蛋白质的复杂混合物中的单独蛋白质进行测序和/或鉴定的方法。在一些实施方案中,标记多肽的一种或多种氨基酸(例如,末端氨基酸和/或内部氨基酸)(例如,直接地或间接地,例如使用诸如氨基酸识别分子的结合剂),并测定多肽中带标记的氨基酸的相对位置。在一些实施方案中,使用一系列氨基酸标记及裂解步骤来确定多肽中氨基酸的相对位置。然而,在一些实施方案中,可无需从多肽中移除氨基酸,但需要通过使带标记的多肽通过孔隙(例如,蛋白质通道)移位,并在通过孔隙移位期间检测来自(多个)带标记的氨基酸的信号(例如,FRET信号),以便确定多肽分子中带标记的氨基酸的相对位置,来确定多肽分子中带标记的氨基酸的相对位置。
在一些实施方案中,评定末端氨基酸(例如,N末端或C末端氨基酸)的身分,其后移除末端氨基酸,并评定末端的下一氨基酸的身分,并重复此过程直至多肽中的多个连续氨基酸得到评定。在一些实施方案中,评定氨基酸的身分包括确定存在的氨基酸的类型。在一些实施方案中,确定氨基酸的类型包括确定实际氨基酸身分,例如通过确定末端氨基酸为天然存在的20种氨基酸中的哪一种(例如,使用对单独末端氨基酸具有特异性的结合剂)。在一些实施方案中,氨基酸的类型选自丙氨酸、精氨酸、天冬酰胺、天冬氨酸、半胱氨酸、谷氨酰胺、谷氨酸、甘氨酸、组氨酸、异亮氨酸、亮氨酸、赖氨酸、甲硫氨酸、苯丙氨酸、脯氨酸、硒半胱氨酸、丝氨酸、苏氨酸、色氨酸、酪氨酸及缬氨酸。
然而,在一些实施方案中,评定末端氨基酸类型的身分可包括确定可在多肽的末端存在的潜在氨基酸的子集。在一些实施方案中,这可通过确定氨基酸并非一种或多种特定氨基酸(并因此可为其他氨基酸中的任一种)来实现。在一些实施方案中,这可通过确定氨基酸的哪一指定子集(例如,基于大小、电荷、疏水性、翻译后修饰、结合特性)可处于多肽的末端(例如,使用与两种或更多种末端氨基酸的指定子集结合的结合剂)来实现。
在一些实施方案中,评定末端氨基酸类型的身分包括确定氨基酸包含翻译后修饰。翻译后修饰的非限制性实例包括乙酰化、ADP-核糖基化、凋亡蛋白酶(caspase)裂解、瓜氨酸化(citrullination)、甲酰化、N-连接糖基化、O-连接糖基化、羟基化、甲基化、豆蔻酰化、类泛素化(neddylation)、硝化、氧化、棕榈酰化、磷酸化、异戊烯化、S-亚硝基化、硫酸化、类小泛素化(sumoylation)及泛素化。
在一些实施方案中,评定末端氨基酸类型的身分包括确定氨基酸包含特征在于一种或多种生物化学特性的侧链。举例而言,氨基酸可包含非极性脂族侧链、带正电侧链、带负电侧链、非极性芳族侧链或极性不带电侧链。包含非极性脂族侧链的氨基酸的非限制性实例包括丙氨酸、甘氨酸、缬氨酸、亮氨酸、甲硫氨酸及异亮氨酸。包含带正电侧链的氨基酸的非限制性实例包括赖氨酸、精氨酸及组氨酸。包含带负电侧链的氨基酸的非限制性实例包括天冬氨酸及谷氨酸。包含非极性芳族侧链的氨基酸的非限制性实例包括苯丙氨酸、酪氨酸及色氨酸。包含极性不带电侧链的氨基酸的非限制性实例包括丝氨酸、苏氨酸、半胱氨酸、脯氨酸、天冬酰胺及谷氨酰胺。
在一些实施方案中,可以将蛋白质或多肽消化成多个较小多肽,并可从这些较小多肽中的一个或多个获得序列信息(例如,使用涉及依次评定多肽的末端氨基酸并移除该氨基酸以暴露末端下一氨基酸的方法)。
在一些实施方案中,多肽从其氨基(N)末端测序。在一些实施方案中,多肽从其羧基(C)末端测序。在一些实施方案中,将多肽的第一末端(例如,N或C末端)固定,且另一末端(例如,C或N末端)如本文所描述测序。
如本文所用,对多肽进行测序是指确定多肽的序列信息。在一些实施方案中,这可涉及确定多肽的部分(或全部)的各顺序氨基酸的身分。然而,在一些实施方案中,这可涉及评定多肽内的氨基酸的子集的身分(例如,并在不确定多肽中各氨基酸的身分的情况下,确定一种或多种氨基酸类型的相对位置)。然而,在一些实施方案中,可在不直接确定多肽中不同类型的氨基酸的相对位置的情况下,从多肽获得氨基酸内容信息。单独的氨基酸内容可用于推断存在的多肽的身分(例如,通过将氨基酸内容与多肽信息的数据库进行比较,并确定哪一(些)多肽具有相同的氨基酸内容)。
在一些实施方案中,可分析获自较长多肽或蛋白质的多种多肽产物(例如,经由酶和/或化学裂解)的序列信息,以重构或推断较长多肽或蛋白质的序列。
因此,在一些实施方案中,通过检测选择性结合该一种或多种类型的氨基酸的一种或多种带标记的亲和试剂的发光来鉴定一种或多种类型的氨基酸。在一些实施方案中,通过检测带标记的多肽的发光来鉴定一种或多种类型的氨基酸。
诸位发明人已进一步认识到且了解,尤其相比于常规多肽测序技术,本文所描述的多肽测序技术可涉及产生新的多肽测序数据。因此,当应用于使用本文所描述的多肽测序技术产生的数据时,用于分析多肽测序数据的常规技术可能不足够。
举例而言,涉及迭代试剂循环的常规多肽测序技术可产生与所测序的多肽的单独氨基酸相关的数据。在此类情况下,分析所产生的数据可能仅仅涉及确定在特定时间时检测到哪一氨基酸,因为所检测到的数据仅对应于一种氨基酸。相比之下,本文所描述的多肽测序技术可在多肽降解过程中产生数据,同时检测多肽分子的多个氨基酸,得到的数据可能难以在对应于多肽的不同氨基酸的数据段之间进行辨别。因此,诸位发明人已开发了用于分析通过本文所描述的多肽测序技术产生的此类数据的新计算技术,这些技术涉及确定对应于单独氨基酸的数据段,诸如通过将数据分段成对应于各自的氨基酸缔合事件的部分。随后可进一步分析这些段,以鉴定在那些单独段期间所检测到的氨基酸。
作为另一实施例,涉及对各类型的氨基酸使用可唯一鉴定的标记的常规测序技术可能涉及仅仅分析在特定时间时检测到哪一标记,而不考虑单独氨基酸如何与其他分子相互作用的任何动力学。相比之下,本文所描述的多肽测序技术产生了指示氨基酸如何与识别分子相互作用的数据。如上文所论述,数据可包括对应于氨基酸与其各自的识别分子之间的缔合事件的特征图案系列。因此,诸位发明人已开发了用于分析特征图案以确定对应于数据的该部分的一种类型的氨基酸的新计算技术,从而允许通过分析不同特征图案系列来测定多肽的氨基酸序列。
带标记的亲和试剂及使用方法
在一些实施方案中,本文所提供的方法包括使多肽与选择性结合一种类型的末端氨基酸的带标记的亲和试剂(在本文中也被称作氨基酸识别分子,其可包括或可不包括标记)接触。如本文所用,在一些实施方案中,末端氨基酸可以指多肽的氨基末端氨基酸或多肽的羧基末端氨基酸。在一些实施方案中,带标记的亲和试剂选择性结合一种类型的末端氨基酸,而不是其他类型的末端氨基酸。在一些实施方案中,带标记的亲和试剂选择性结合一种类型的末端氨基酸,而不是相同类型的内部氨基酸。在其他实施例中,带标记的亲和试剂选择性结合处于多肽的任何位置处的一种类型的氨基酸,例如作为末端氨基酸及内部氨基酸的相同类型的氨基酸。
如本文所用,在一些实施方案中,一种类型的氨基酸是指二十种天然存在的氨基酸中的一种或其类型的子集。在一些实施方案中,一种类型的氨基酸是指二十种天然存在的氨基酸中的一种的被修饰的变体,或其未被修饰的和/或被修饰的变体的子集。被修饰的氨基酸变体的实例包括(但不限于)翻译后修饰变体(例如,乙酰化、ADP-核糖基化、凋亡蛋白酶裂解、瓜氨酸化、甲酰化、N-连接糖基化、O-连接糖基化、羟基化、甲基化、豆蔻酰化、类泛素化、硝化、氧化、棕榈酰化、磷酸化、异戊烯化、S-亚硝基化、硫酸化、类小泛素化及泛素化)、化学修饰变体、非天然氨基酸及蛋白型(proteinogenic)氨基酸(诸如硒半胱氨酸及吡咯赖氨酸)。在一些实施方案中,氨基酸类型的子集包括具有一种或多种类似生物化学特性的多于一种且少于二十种氨基酸。举例而言,在一些实施方案中,一种类型的氨基酸是指选自以下的一种类型:具有带电侧链(例如,带正电和/或带负电侧链)的氨基酸、具有极性侧链(例如,极性不带电侧链)的氨基酸、具有非极性侧链(例如,非极性脂族和/或芳族侧链)的氨基酸及具有疏水性侧链的氨基酸。
在一些实施方案中,本文所提供的方法包括使多肽与一种或多种带标记的亲和试剂接触,所述一种或多种带标记的亲和试剂选择性地结合一种或多种类型的末端氨基酸。作为说明性且非限制性实例,其中在本申请的方法中使用了四种带标记的亲和试剂,任一种试剂选择性结合一种类型的末端氨基酸,其不同于另外三种中的任一种所选择性结合的另一类型的氨基酸(例如,第一试剂结合第一类型,第二试剂结合第二类型,第三试剂结合第三类型且第四试剂结合第四类型的末端氨基酸)。出于此论述的目的,在本文所描述的方法的情形下,一种或多种带标记的亲和试剂可被替代地称作一组带标记的亲和试剂。
在一些实施方案中,一组带标记的亲和试剂包括至少一种且至多六种带标记的亲和试剂。举例而言,在一些实施方案中,一组带标记的亲和试剂包括一种、两种、三种、四种、五种或六种带标记的亲和试剂。在一些实施方案中,一组带标记的亲和试剂包括十种或更少带标记的亲和试剂。在一些实施方案中,一组带标记的亲和试剂包括八种或更少带标记的亲和试剂。在一些实施方案中,一组带标记的亲和试剂包括六种或更少带标记的亲和试剂。在一些实施方案中,一组带标记的亲和试剂包括四种或更少带标记的亲和试剂。在一些实施方案中,一组带标记的亲和试剂包括三种或更少带标记的亲和试剂。在一些实施方案中,一组带标记的亲和试剂包括两种或更少带标记的亲和试剂。在一些实施方案中,一组带标记的亲和试剂包括四种带标记的亲和试剂。在一些实施方案中,一组带标记的亲和试剂包括至少两种且至多二十种(例如,至少两种且至多十种、至少两种且至多八种、至少四种且至多二十种、至少四种且至多十种)带标记的亲和试剂。在一些实施方案中,一组带标记的亲和试剂包括多于二十种(例如,20种至25种、20种至30种)亲和试剂。然而,应了解,根据本申请的方法可使用任何数目的亲和试剂以适应所需用途。
根据本申请,在一些实施方案中,通过检测带标记的亲和试剂(例如,包含发光标记的氨基酸识别分子)的发光来鉴定一种或多种类型的氨基酸。在一些实施方案中,带标记的亲和试剂包括选择性结合一种类型的氨基酸的亲和试剂,及具有发光的与亲和试剂缔合的发光标记。以此方式,可使发光(例如,发光寿命、发光强度及本文别处所描述的其他发光特性)与亲和试剂的选择性结合相关联,以鉴定多肽的氨基酸。在一些实施方案中,可在根据本申请的方法中使用多种类型的带标记的亲和试剂,其中各类型包括具有可从多个当中唯一地鉴定的发光的发光标记。适合的发光标记可包括发光分子,诸如荧光团染料,且在本文别处描述。
在一些实施方案中,通过检测带标记的亲和试剂的一种或多种电特征来鉴定一种或多种类型的氨基酸。在一些实施方案中,带标记的亲和试剂包括选择性结合一种类型的氨基酸的亲和试剂,及与亲和试剂缔合的导电性标记。以此方式,可使一种或多种电特征(例如,电荷、电流振荡色彩及其他电特征)与亲和试剂的选择性结合相关联,以鉴定多肽的氨基酸。在一些实施方案中,可在根据本申请的方法中使用多种类型的带标记的亲和试剂,其中各类型包括产生可从多个当中唯一地鉴定的电信号变化(例如,电导变化,诸如导电率幅度变化及特征图案的导电性转变)的导电性标记。在一些实施方案中,多种类型的带标记的亲和试剂各自包括导电性标记,其具有不同数目的带电基团(例如,不同数目的带负电和/或带正电基团)。因此,在一些实施方案中,导电性标记为电荷标记。电荷标记的实例包括树状体、纳米粒子、核酸及具有多个带电基团的其他聚合物。在一些实施方案中,导电性标记可由其净电荷(例如,净正电荷或净负电荷)、由其电荷密度和/或由其带电基团的数目唯一地鉴定。
在一些实施方案中,亲和试剂(例如,氨基酸识别分子)可由本领域技术人员使用常规技术进行工程改造。在一些实施方案中,期望的特性可包括只有当一种类型的氨基酸位于多肽的末端(例如,N末端或C末端)处时,才选择性地且以高亲和力结合至该类型的氨基酸的能力。在其他实施例中,期望的特性可包括当一种类型的氨基酸位于多肽的末端(例如,N末端或C末端)处时,以及当其位于多肽的内部位置处时,选择性地且以高亲和力结合至该类型的氨基酸的能力。在一些实施方案中,期望的特性包括选择性地且以低亲和力(例如,以约50nM或更高,例如,在约50nM与约50μM之间、在约100nM与约10μM之间、在约500nM与约50μM之间的K
如本文所用,在一些实施方案中,术语“选择性(selective)”及“特异性(specific)”(及其变化形式,例如选择性(selectively/selectivity)、特异性(specifically/specificity))是指优先结合相互作用。举例而言,在一些实施方案中,选择性结合一种类型的氨基酸的带标记的亲和试剂优先结合这一种类型的氨基酸,而不是另一类型的氨基酸。选择性结合相互作用将区分一种类型的氨基酸(例如,一种类型的末端氨基酸)与其他类型的氨基酸(例如,其他类型的末端氨基酸),通常高于约10至100倍或更高(例如,高于约1,000或10,000倍)。因此,应了解,选择性结合相互作用可以指相比于其他类型的氨基酸,一种类型的氨基酸可唯一地鉴定的任何结合相互作用。举例而言,在一些方面中,本申请提供了通过获得指示一种或多种氨基酸识别分子与多肽分子缔合的数据来进行多肽测序的方法。在一些实施方案中,数据包括对应于以下的信号脉冲系列:与多肽分子的氨基酸的可逆氨基酸识别分子结合相互作用系列,且数据可用于确定氨基酸的身分。如此,在一些实施方案中,“选择性”或“特异性”结合相互作用是指区分一种类型的氨基酸与其他类型的氨基酸的所检测到的结合相互作用。
在一些实施方案中,带标记的亲和试剂(例如,氨基酸识别分子)以低于约10
在一些实施方案中,带标记的亲和试剂(例如,氨基酸识别分子)以低于约10
根据本文所提供的方法及组合物,图1C展示了带标记的亲和试剂的各种示例性构造及用途。在一些实施方案中,带标记的亲和试剂100包括发光标记110(例如,标记),及选择性结合多肽120的一种或多种类型的末端氨基酸的亲和试剂(展示为带点形状)。在一些实施方案中,亲和试剂具有针对处于末端位置或处于末端位置及内部位置两者处的一种类型的氨基酸或一子集(例如,少于二十种常见类型的氨基酸)类型的氨基酸的选择性。
如本文所描述,亲和试剂(也被称作“识别分子”)可为能够选择性或特异性结合一种分子,而不是另一种分子(例如,一种类型的氨基酸,而不是另一种类型的氨基酸,如在本文所提及的“氨基酸识别分子”的情况下)的任何生物分子。在一些实施方案中,亲和试剂不为肽酶或不具有肽酶活性。举例而言,在一些实施方案中,本申请的多肽测序的方法涉及使多肽分子与一种或多种亲和试剂及裂解试剂接触。在此类实施例中,一种或多种亲和试剂不具有肽酶活性,且由裂解试剂进行一个或多个氨基酸从多肽分子的移除(例如,从多肽分子的末端进行氨基酸移除)。
亲和试剂(例如,识别分子)包括例如蛋白质及核酸,其可为合成或重组的。在一些实施方案中,亲和试剂或识别分子可为抗体或抗体的抗原结合部分、含SH2结构域蛋白或其片段或酶生物分子,诸如肽酶、氨基转移酶、核酶、人工适体酶(aptazyme)或tRNA合成酶(包括氨基酰基-tRNA合成酶),及描述于2016年9月2日申请的标题为“用于迭代多肽分析及加工的分子及方法(MOLECULES AND METHODS FOR ITERATIVE POLYPEPTIDE ANALYSIS ANDPROCESSING)”的美国专利申请第15/255,433号中的相关分子。
在一些实施方案中,本申请的亲和试剂或识别分子为降解途径蛋白。适合用作识别分子的降解途径蛋白的实例包括(但不限于)N端规则途径蛋白,诸如Arg/N端规则途径蛋白、Ac/N端规则途径蛋白及Pro/N端规则途径蛋白。在一些实施方案中,识别分子为选自以下的N端规则途径蛋白:Gid蛋白(例如,Gid4或Gid10蛋白)、UBR盒(UBR box)蛋白(例如,UBR1、UBR2)或其含UBR盒结构域的蛋白片段、p62蛋白或其含ZZ结构域的片段及ClpS蛋白(例如,ClpS1、ClpS2)。
在一些实施方案中,本申请的亲和试剂或识别分子为ClpS蛋白,诸如根癌农杆菌ClpS1、根癌农杆菌ClpS2、细长聚球藻ClpS1、细长聚球藻ClpS2、细长嗜热聚球藻ClpS、大肠杆菌(Escherichia coli)ClpS或恶性疟原虫(Plasmodium falciparum)ClpS。在一些实施方案中,识别分子为L/F转移酶,诸如大肠杆菌亮氨酰基/苯丙氨酰基-tRNA-蛋白转移酶。在一些实施方案中,识别分子为D/E亮氨酸转移酶,诸如创伤弧菌(Vibrio vulnificus)天冬氨酸/谷氨酸亮氨酰转移酶Bpt。在一些实施方案中,识别分子为UBR蛋白或UBR盒结构域,诸如人类UBR1及UBR2或酿酒酵母(Saccharomyces cerevisiae)UBR1的UBR蛋白或UBR盒结构域。在一些实施方案中,识别分子为p62蛋白,诸如智人(H.sapiens)p62蛋白或褐家鼠(Rattus norvegicus)p62蛋白,或其最少包括ZZ结构域的截短变体。在一些实施方案中,识别分子为Gid4蛋白,诸如智人GID4或酿酒酵母GID4。在一些实施方案中,识别分子为Gid10蛋白,诸如酿酒酵母GID10。在一些实施方案中,识别分子为N-豆蔻酰基转移酶(N-meristoyltransferase),诸如硕大利什曼原虫(Leishmania major)N-豆蔻酰基转移酶或智人N-豆蔻酰基转移酶NMT1。在一些实施方案中,识别分子为BIR2蛋白,诸如黑腹果蝇(Drosophila melanogaster)BIR2。在一些实施方案中,识别分子为酪氨酸激酶或酪氨酸激酶的SH2结构域,诸如智人Fyn SH2结构域、智人Src酪氨酸激酶SH2结构域,或其变体,诸如智人Fyn SH2结构域三重突变体超结合子(superbinder)。在一些实施方案中,识别分子为抗体或抗体片段,诸如针对磷酸化酪氨酸或本文所描述的另一翻译后修饰氨基酸变体的单链抗体可变片段(scFv)。
表1提供了氨基酸识别分子的示例性序列的列表。表1中还展示了就多肽的末端位置处的氨基酸身分而言,各分子的氨基酸结合偏好,除非另外规定。应了解,这些序列及本文所描述的其他实例意欲为非限制性的,且根据本申请的识别分子可包括其任何同系物、变体,或其最少含有负责肽识别的结构域或子结构域的片段。
表1.氨基酸识别蛋白的非限制性实例。
*结合偏好根据公开科学文献推断和/或由诸位发明人在如本文所描述的单分子实验中进一步证明。
**与磷酸化酪氨酸的结合可出现在肽末端或内部位置处。
因此,在一些实施方案中,本申请提供了氨基酸识别分子,其具有选自表1的氨基酸序列(或具有与选自表1的氨基酸序列具有至少50%、至少60%、至少70%、至少80%、80%至90%、90%至95%、95%至99%或更高氨基酸序列一致性的氨基酸序列)。在一些实施方案中,氨基酸识别分子与表1中所列的氨基酸识别分子具有25%至50%、50%至60%、60%至70%、70%至80%、80%至90%、90%至95%或95%至99%或更高氨基酸序列一致性。在一些实施方案中,氨基酸识别分子为被修饰的氨基酸识别分子,且相对于表1中所列举的序列,包括一种或多种氨基酸突变。
在一些实施方案中,氨基酸识别分子包括标签序列,该标签序列提供除氨基酸结合外的一个或多个功能。举例而言,在一些实施方案中,标签序列包括生物素连接酶识别序列,其准许对识别分子进行生物素标记(例如,并入一个或多个生物素分子,包括生物素及双生物素部分)。标签序列中的官能性序列的额外实例包括纯化标签、裂解位点及适用于识别分子的纯化和/或修饰的其他部分。表2提供了末端标签序列的非限制性序列的列表,这些序列中的任何一个或多个可与本申请的氨基酸识别分子中的任一种组合(例如,与表1中所列举的序列组合)使用。应了解,表2中所展示的标签序列意欲为非限制性的,且根据本申请的识别分子可包括在识别分子多肽的N末端或C末端、在N末端与C末端之间分割或如本领域中所实践以其他方式重排的标签序列(例如,His标签和/或生物素标记标签)中的任何一种或多种。
表2.末端标签序列的非限制性实例。
在一些实施方案中,本申请的识别分子或亲和试剂为肽酶。肽酶,也称作蛋白酶(protease/proteinase),是催化肽键的水解的酶。肽酶将多肽消化成较短片段,且一般可分类为内肽酶及外肽酶,其分别在内部及末端裂解多肽链。在一些实施方案中,带标记的亲和试剂100包括已被修饰以使外肽酶或内肽酶活性失活的肽酶。以此方式,带标记的亲和试剂100选择性结合来自多肽的氨基酸,而不同时将氨基酸从多肽裂解。在其他实施例中,可使用尚未被修饰以使外肽酶或内肽酶活性失活的肽酶。举例而言,在一些实施方案中,带标记的亲和试剂包括带标记的外肽酶102。
根据本申请的某些实施方案,蛋白质测序方法可包括在多肽的末端迭代检测及裂解。在一些实施方案中,带标记的外肽酶102可用作进行氨基酸检测及裂解步骤两者的单一试剂。如大体上所描绘,在一些实施方案中,带标记的外肽酶102具有氨肽酶或羧肽酶活性,使得其分别选择性结合并裂解来自多肽的N末端或C末端氨基酸。应了解,在某些实施方案中,如本文所描述,带标记的外肽酶102可由本领域技术人员催化失活,使得带标记的外肽酶102保留选择性结合特性,用作非裂解性带标记的亲和试剂100。
外肽酶一般需要多肽底物,以包括其氨基末端的游离氨基或其羧基末端的游离羧基中的至少一种。在一些实施方案中,根据本申请的外肽酶水解处于或接近于多肽的末端的键。在一些实施方案中,外肽酶水解距多肽末端不超过三个残基的键。举例而言,在一些实施方案中,由外肽酶催化的单一水解反应从多肽末端裂解单一氨基酸、二肽或三肽。
在一些实施方案中,根据本申请的外肽酶为氨肽酶或羧肽酶,其分别从氨基末端或羧基末端裂解单一氨基酸。在一些实施方案中,根据本申请的外肽酶为二肽基-肽酶或肽基-二肽酶,其分别从氨基末端或羧基末端裂解二肽。在其他实施例中,根据本申请的外肽酶为三肽基-肽酶,其从氨基末端裂解三肽。肽酶分类及其各类别或子类别的活性为熟知的,且描述于文献中(参见例如Gurupriya,V.S.及Roy,S.C.Proteases and ProteaseInhibitors in Male Reproduction.Proteases in Physiology and Pathology 195-216(2017);及Brix,K.及
可基于测序反应的方向性来选择或工程改造根据本申请的外肽酶。举例而言,在从多肽的氨基末端向羧基末端测序的实施例中,外肽酶包括氨肽酶活性。相反,在从多肽的羧基末端向氨基末端测序的实施例中,外肽酶包括羧肽酶活性。识别特定羧基末端氨基酸的羧肽酶(其可用作带标记的外肽酶,或被失活以用作本文所描述的非裂解性带标记的亲和试剂)已描述于文献中(参见例如Garcia-Guerrero,M.C.等人(2018)PNAS 115(17))。
适合用作裂解试剂和/或亲和试剂(例如,识别分子)的肽酶包括选择性结合一种或多种类型的氨基酸的氨肽酶。在一些实施方案中,氨肽酶识别分子被修饰以使氨肽酶活性失活。在一些实施方案中,氨肽酶裂解试剂为非特异性的,使得其从多肽末端裂解大部分或所有类型的氨基酸。在一些实施方案中,相比于多肽末端的其他类型的氨基酸,氨肽酶裂解试剂在裂解多肽末端的一种或多种类型的氨基酸时更高效。举例而言,根据本申请的氨肽酶特异性裂解丙氨酸、精氨酸、天冬酰胺、天冬氨酸、半胱氨酸、谷氨酰胺、谷氨酸、甘氨酸、组氨酸、异亮氨酸、亮氨酸、赖氨酸、甲硫氨酸、苯丙氨酸、脯氨酸、硒半胱氨酸、丝氨酸、苏氨酸、色氨酸、酪氨酸和/或缬氨酸。在一些实施方案中,氨肽酶为脯氨酸氨肽酶。在一些实施方案中,氨肽酶为脯氨酸亚胺基肽酶。在一些实施方案中,氨肽酶为谷氨酸/天冬氨酸特异性氨肽酶。在一些实施方案中,氨肽酶为甲硫氨酸特异性氨肽酶。在一些实施方案中,氨肽酶为表3中所列举的氨肽酶。在一些实施方案中,氨肽酶裂解试剂裂解如表3中所列举的肽底物。
在一些实施方案中,氨肽酶为非特异性氨肽酶。在一些实施方案中,非特异性氨肽酶为锌金属蛋白酶。在一些实施方案中,非特异性氨肽酶为表4中所列举的氨肽酶。在一些实施方案中,非特异性氨肽酶裂解如表4中所列举的肽底物。
因此,在一些实施方案中,本申请提供了氨肽酶(例如,氨肽酶识别分子、氨肽酶裂解试剂),其具有选自表3或表4的氨基酸序列(或具有与选自表3或表4的氨基酸序列具有至少50%、至少60%、至少70%、至少80%、80%至90%、90%至95%、95%至99%或更高氨基酸序列一致性的氨基酸序列)。在一些实施方案中,氨肽酶与表3或表4中所列的氨肽酶具有25%至50%、50%至60%、60%至70%、70%至80%、80%至90%、90%至95%或95%至99%或更高氨基酸序列一致性。在一些实施方案中,氨肽酶为被修饰的氨肽酶,且相对于表3或表4中所列举的序列,包括一种或多种氨基酸突变。
表3.氨肽酶的非限制性实例。
表4.非特异性氨肽酶的非限制性实例。
*裂解效率(从最高至最低):精氨酸>赖氨酸>疏水性残基(包括丙氨酸、亮氨酸、甲硫氨酸及苯丙氨酸)>脯氨酸(参见例如Matthews Biochemistry 47,2008,5303-5311)。
**裂解效率(从最高至最低):亮氨酸>丙氨酸>精氨酸>苯丙氨酸>脯氨酸;在谷氨酸及天冬氨酸后不裂解。
出于比较两种或更多种氨基酸序列的目的,可通过以下计算第一氨基酸序列与第二氨基酸序列之间的“序列一致性”(在本文也称为“氨基酸一致性”)百分比:[第一氨基酸序列中与第二氨基酸序列中对应位置处的氨基酸残基一致的氨基酸残基的数目]除以[第一氨基酸序列中的氨基酸残基的总数目]并乘以[100],其中与第一氨基酸序列相比,第二氨基酸序列中各氨基酸残基缺失、插入、取代或添加视为单一氨基酸残基(位置)处的差异。可替代地,两种氨基酸序列之间的序列一致性程度可使用已知电脑算法(例如,通过Smith及Waterman(1970)Adv.Appl.Math.2:482c的局部同源性算法,通过Needleman及Wunsch,J.Mol.Biol.(1970)48:443的同源性比对算法,通过Pearson及Lipman.Proc.Natl.Acad.Sci.USA(1998)85:2444的搜索相似度方法,或通过可以Blast、Clustal Omega获得的算法或其他序列比对算法的电脑化实施),并例如使用标准设置来计算。通常,为了根据上文概述的计算方法测定两种氨基酸序列之间的“序列一致性”百分比,具有最大氨基酸残基数目的氨基酸序列将被视为“第一”氨基酸序列,且另一氨基酸序列将被视为“第二”氨基酸序列。
另外或替代地,可评定两种或更多种序列的序列之间的一致性。在两种或更多种核酸或氨基酸序列的情形下,术语“一致”或百分比“一致性”是指相同的两种或更多种序列或子序列。当比较且比对比较窗或指明的区域上如使用以上序列比较算法中的一种或通过手动比对及目视检验测量的最大一致性时,若两种序列在指定区域上或完整序列上具有指定百分比的相同(例如,至少80%、85%、90%、95%、96%、97%、98%、99%、99.5%、99.6%、99.7%、99.8%或99.9%一致)氨基酸残基或核苷酸,则两种序列“实质上一致”。任选地,,一致性存在于长度为至少约25、50、75或100个氨基酸的区域上,或长度为100至150、150至200、100至200或200或更多个氨基酸的区域上。
另外或替代地,可评定两种或更多种序列的序列之间的对准(alignment)。在两种或更多种核酸或氨基酸序列的情形下,术语“对准”或百分比“对准”是指相同的两种或更多种序列或子序列。当比较且比对比较窗或指明的区域上如使用以上序列比较算法中的一种或通过手动比对及目视检验测量的最大一致性时,若两种序列在指定区域上或完整序列上具有指定百分比的相同(例如,至少80%、85%、90%、95%、96%、97%、98%、99%、99.5%、99.6%、99.7%、99.8%或99.9%一致)氨基酸残基或核苷酸,则两种序列“实质上对准”。任选地,,对准存在于长度为至少约25、50、75或100个氨基酸的区域上,或长度为100至150、150至200、100至200或200或更多个氨基酸的区域上。
除蛋白质分子以外,核酸分子具有用作根据本申请的亲和试剂(例如,氨基酸识别分子)的多种有利特性。
核酸适体为已经工程改造以便以高亲和力及选择性结合所需目标的核酸分子。因此,核酸适体可使用本领域中已知的选择和/或富集技术进行工程改造,以选择性结合所需类型的氨基酸。因此,在一些实施方案中,亲和试剂包括核酸适体(例如,DNA适体、RNA适体)。如图1C中所展示,在一些实施方案中,带标记的亲和试剂为选择性结合一种类型的末端氨基酸的带标记的适体104。举例而言,在一些实施方案中,如本文所描述,带标记的适体104选择性结合多肽的末端的一种类型的氨基酸(例如,单一类型的氨基酸或一子集类型的氨基酸)。尽管未图示,但应了解,根据本申请的方法,带标记的适体104可被工程改造,以选择性结合多肽的任何位置处(例如,多肽的末端位置处或末端及内部位置处)的一种类型的氨基酸。
在一些实施方案中,带标记的亲和试剂包括具有结合诱导型发光的标记。举例而言,在一些实施方案中,带标记的适体106包括供体标记112及受体标记114,且如图1C的图(I)及图(II)中所示出的那样起作用。如图(I)中所描绘,呈游离分子形式的带标记的适体106采用一种构形,其中供体标记112与受体标记114间隔开一段距离,该距离限制了标记之间的可检测FRET(例如,约10nm或更大)。如图(II)中所描绘,呈选择性结合的分子形式的带标记的适体106采用一种构形,其中供体标记112与受体标记114在一段距离之内,该距离促进了标记之间的可检测FRET(例如,约10nm或更小)。在其他实施例中,带标记的适体106包括淬灭部分,且类似于分子信标起作用,其中带标记的适体106的发光以游离分子形式在内部淬灭,且以选择性结合的分子形式复原(参见例如Hamaguchi等人(2001)AnalyticalBiochemistry 294,126-131)。在不希望受理论束缚的情况下,认为用于结合诱导型发光的这些及其他类型的机制可有利地减少或消除背景发光,以提高本文所描述的方法的整体灵敏度及准确度。
除鉴定多肽的末端氨基酸的方法以外,本申请提供了使用带标记的亲和试剂对多肽进行测序的方法。在一些实施方案中,测序的方法可涉及使多肽末端经历末端氨基酸检测及末端氨基酸裂解的重复循环。举例而言,在一些实施方案中,本申请提供了确定多肽的氨基酸序列的方法,其包括使多肽与本文所描述的一种或多种带标记的亲和试剂接触,以及使多肽经历艾德曼降解。
常规艾德曼降解涉及修饰及裂解多肽的末端氨基酸的重复循环,其中鉴定各连续裂解的氨基酸,以确定多肽的氨基酸序列。作为常规艾德曼降解的说明性实例,使用异硫氰酸苯酯(PITC)修饰多肽的N末端氨基酸,以形成PITC衍生化N末端氨基酸。随后使用酸性条件、碱性条件和/或高温裂解PITC衍生化N末端氨基酸。还已经展示,裂解PITC衍生化N末端氨基酸的步骤可以酶方式实现,使用来自原虫克氏锥虫(Trypanosoma cruzi)的被修饰的半胱氨酸蛋白酶,其涉及在中性或接近于中性pH下的相对较温和裂解条件。适用酶的非限制性实例描述于2016年9月2日申请的标题为“用于迭代多肽分析及加工的分子及方法(MOLECULES AND METHODS FOR ITERATIVE POLYPEPTIDE ANALYSIS AND PROCESSING)”的美国专利申请第15/255,433号中。
使用根据本申请的带标记的亲和试剂通过艾德曼降解进行测序的实例描绘于图1D中。在一些实施方案中,通过艾德曼降解进行测序包括提供多肽122,其通过连接子124固定至固体载体的表面130(例如,固定至样品孔的底部或侧壁表面)。在一些实施方案中,如本文所描述,多肽122在一个末端(例如,氨基末端氨基酸或羧基末端氨基酸)固定,使得另一末端对于进行末端氨基酸的检测及裂解而言是游离的。因此,在一些实施方案中,本文所描述的艾德曼降解方法中所使用的试剂优先与多肽122的非固定(例如,游离)末端的末端氨基酸相互作用。以此方式,多肽122在检测及裂解的重复循环内保持固定。为此目的,在一些实施方案中,连接子124可根据用于检测及裂解的所需条件组设计,例如以限制多肽122在化学裂解条件下从表面130脱离。适用于将多肽固定至表面的连接子组合物及技术详细描述于本文别处。
根据本申请,在一些实施方案中,通过艾德曼降解进行测序的方法包括步骤(1):使多肽122与一种或多种带标记的亲和试剂接触,所述一种或多种带标记的亲和试剂选择性地结合一种或多种类型的末端氨基酸。如所展示,在一些实施方案中,带标记的亲和试剂108通过选择性地结合末端氨基酸而与多肽122相互作用。在一些实施方案中,步骤(1)进一步包括移除不选择性结合多肽122的末端氨基酸(例如,游离末端氨基酸)的一种或多种带标记的亲和试剂中的任一种。
在一些实施方案中,该方法进一步包括通过检测带标记的亲和试剂108来鉴定多肽122的末端氨基酸。在一些实施方案中,检测包括检测来自带标记的亲和试剂108的发光。如本文所描述,在一些实施方案中,发光与带标记的亲和试剂108唯一关联,从而发光与亲和试剂108所选择性结合的氨基酸的类型相关联。如此,在一些实施方案中,通过确定带标记的亲和试剂108的一种或多种发光特性来鉴定氨基酸的类型。
在一些实施方案中,通过艾德曼降解进行测序的方法包括步骤(2):移除多肽122的末端氨基酸。在一些实施方案中,步骤(2)包括从多肽122移除带标记的亲和试剂108(例如,选择性结合末端氨基酸的一种或多种带标记的亲和试剂中的任一种)。在一些实施方案中,步骤(2)包括通过使末端氨基酸与异硫氰酸酯(例如,PITC)接触以形成经异硫氰酸酯修饰的末端氨基酸,来修饰多肽122的末端氨基酸(例如,游离末端氨基酸)。在一些实施方案中,与未被修饰的末端氨基酸相比,经异硫氰酸酯修饰的末端氨基酸对由裂解试剂(例如,化学或酶裂解试剂)进行的移除较敏感。
在一些实施方案中,步骤(2)包括通过使多肽122与特异性结合并裂解经异硫氰酸酯修饰的末端氨基酸的蛋白酶140接触,来移除末端氨基酸。在一些实施方案中,蛋白酶140包括被修饰的半胱氨酸蛋白酶。在一些实施方案中,蛋白酶140包括被修饰的半胱氨酸蛋白酶,诸如来自克氏锥虫的半胱氨酸蛋白酶(参见例如Borgo等人(2015)Protein Science24:571-579)。在其他实施例中,步骤(2)包括通过使多肽122经历足以裂解经异硫氰酸酯修饰的末端氨基酸的化学(例如,酸性、碱性)条件,来移除末端氨基酸。
在一些实施方案中,通过艾德曼降解进行测序的方法包括步骤(3):在末端氨基酸裂解之后洗涤多肽122。在一些实施方案中,洗涤包括移除蛋白酶140。在一些实施方案中,洗涤包括使多肽122复原至中性pH条件(例如,在由酸性或碱性条件进行化学裂解之后)。在一些实施方案中,通过艾德曼降解进行测序的方法包括重复步骤(1)至步骤(3),持续多个循环。
通过艾德曼降解进行测序的示例性方法展示于图1E中。在一些实施方案中,可使用常用酶将含有多肽的复杂混合物(例如,蛋白质的混合物)的样品降解成大约6至40个氨基酸的短多肽片段。在一些实施方案中,根据本申请的方法对此多肽库进行测序将展现初始复杂混合物中存在的多肽中的每一个的身分及丰度。如本文及文献中所描述,6至40个氨基酸的大小范围内的大部分多肽可通过确定多肽链内恰好四个氨基酸的数目及位置唯一地鉴定。
因此,在一些实施方案中,通过艾德曼降解进行测序的方法可使用一组带标记的适体150进行,带标记的适体150包括四种DNA适体类型,各类型识别不同的N末端氨基酸。各适体类型可经不同的发光标记标记,使得可基于一种或多种发光特性来区别不同的适体类型。出于说明的目的,示例性带标记的适体150组包括:被第一发光标记(“染料1”)标记的半胱氨酸特异性适体;被第二发光标记(“染料2”)标记的赖氨酸特异性适体;被第三发光标记(“染料3”)标记的色氨酸特异性适体;及被第四发光标记(“染料4”)标记的谷氨酸特异性适体。
在一些实施方案中,根据本申请的通过艾德曼降解进行测序的方法根据图1E中所展示的过程152进行。在一些实施方案中,在步骤(1)之前,将来自多肽库的单一多肽分子固定至固体载体的表面,例如在样品孔阵列的样品孔的底部或侧壁表面处。在一些实施方案中,如本文别处所描述,能够实现表面固定的部分(例如,生物素)或提高溶解度的部分(例如,寡核苷酸)可以化学方式或酶方式连接至多肽的C末端。为了确定各多肽的序列,在一些实施方案中,使固定化多肽经历N末端氨基酸检测及N末端氨基酸裂解的重复循环,如过程152所示出。在一些实施方案中,过程152包括试剂添加及洗涤步骤,其通过使用从动化流体系统注射至检测表面上方的流槽来进行。在一些实施方案中,步骤(1)至(4)示出了使用带标记的适体150进行检测及裂解的一个循环。
在一些实施方案中,根据过程152通过艾德曼降解进行测序的方法包括步骤(1):流入四种正交标记的DNA适体的混合物,并孵育,以允许适体结合至在N末端含有四种适当氨基酸中的一种的任何固定化多肽(例如,在阵列的样品孔内固定的多肽)。在一些实施方案中,方法进一步包括洗涤固定化多肽,以移除未结合的适体。在一些实施方案中,方法进一步包括对固定化多肽进行成像(“成像步骤1”)。在一些实施方案中,所获取的影像含有足以确定适体结合的多肽的位置(例如,样品孔阵列内的位置)及四种适体中的哪一种在各位置处结合的信息。在一些实施方案中,方法进一步包括使用适当缓冲液洗涤固定化多肽,以将适体从固定化多肽上移除。
在一些实施方案中,根据过程152测序的方法包括步骤(2):流入含有特异性修饰N末端氨基的反应性分子(例如,PITC,如所展示)的溶液。诸如PITC的异硫氰酸酯分子,在一些实施方案中,将N末端氨基酸修饰成用于由被修饰的蛋白酶(诸如来自克氏锥虫的半胱氨酸蛋白酶克氏锥虫半胱氨酸蛋白酶(cruzain))进行裂解的底物。
在一些实施方案中,根据过程152的测序方法包括步骤(3):洗涤固定化多肽,随后流入合适的被修饰的蛋白酶,该被修饰的蛋白酶识别被修饰的N末端氨基酸并将其从固定化多肽上裂解。在一些实施方案中,方法包括步骤(4):在酶裂解之后洗涤固定化多肽。在一些实施方案中,步骤(1)至步骤(4)描绘了艾德曼降解的一个循环。因此,如所展示的步骤(1')为下一反应循环的开始,该循环以如上文针对步骤(1)至步骤(4)所描述进行的步骤(1')至步骤(4')继续进行。在一些实施方案中,步骤(1)至步骤(4)重复大约20至40个循环。
在一些实施方案中,带标记的异硫氰酸酯(例如,染料标记的PITC)可用于监测样品负载。举例而言,在一些实施方案中,在使多肽样品经历如过程152中所展示的测序方法之前,使用染料标记的PITC对末端进行修饰,使多肽样品与发光标记在末端预先缀合。以此方式,可通过在启动过程152之前检测来自标记的发光,来监测多肽样品向样品孔阵列中的负载。在一些实施方案中,使用发光来确定阵列中样品孔的单一占有率(例如,含有单一多肽分子的样品孔的分率),其可有利地增加针对给定样品可靠地获得的信息的量。一旦所需样品负载状态由发光确定,则可如所描述的那样,通过化学或酶裂解来启动过程152,随后以步骤(1)进行。
在一些实施方案中,带标记的异硫氰酸酯(例如,染料标记的PITC)可用于监测阵列中多肽样品的反应进展。举例而言,在一些实施方案中,步骤(2)包括流入含有染料标记的PITC的溶液,该染料标记的PITC特异性修饰并标记样品中多肽的N末端氨基。在一些实施方案中,可在步骤(2)期间或之后检测来自标记的发光,以评估样品中多肽的N末端PITC修饰。因此,在一些实施方案中,使用发光来判定是否或何时自步骤(2)继续进行至步骤(3)。在一些实施方案中,可在步骤(3)期间或之后检测来自标记的发光,以评估样品中多肽的N末端氨基酸裂解,例如以判定是否或何时自步骤(3)继续进行至步骤(4)。
根据过程152的测序方法利用不同试剂来检测及裂解多肽的末端氨基酸。尽管如此,在一些方面中,本申请提供了测序的方法,其中使用单一试剂(包括肽酶)来检测及裂解多肽的末端氨基酸。图2展示了使用一组带标记的外肽酶200进行多肽测序的实施例,其中各带标记的外肽酶选择性结合并裂解不同类型的末端氨基酸。
如图2的实施例中大体上示出,带标记的外肽酶200包括:包括第一发光标记的赖氨酸特异性外肽酶、包括第二发光标记的甘氨酸特异性外肽酶、包括第三发光标记的天冬氨酸特异性外肽酶及包括第四发光标记的亮氨酸特异性外肽酶。根据本文所描述的某些实施方案,仅当氨基酸处于多肽的氨基或羧基末端时,带标记的外肽酶200中的每一种才会选择性结合并裂解其各自的氨基酸。因此,由于通过此方法进行的测序从肽的一个末端向另一末端进行,因此带标记的外肽酶200被工程改造或选择,以使得组中所有试剂都将具有氨肽酶或羧肽酶活性。
如图2中进一步所展示,过程202示意性地示出了使用带标记的外肽酶200进行的实时测序反应。图(I)至图(IX)示出了事件进展,涉及与下方展示的信号输出相关,且对应于各图中所描绘的事件的多肽的末端迭代检测及裂解。出于说明的目的,展示了具有“KLDG…”的任意选择的氨基酸序列的多肽(从一个末端向另一末端进行)。
图(I)描绘了测序反应的开始,其中多肽被固定至固体载体的表面,诸如样品孔的底部或侧壁表面。在一些实施方案中,根据本申请的测序方法包括实时单分子测序。在一些实施方案中,多个单分子测序反应在样品孔阵列中同时进行。在此类实施方案中,多肽固定通过将多肽锚定在用于单分子分析的样品孔内,防止多肽扩散离开样品孔。
图(II)描绘了检测事件,其中来自带标记的亲和试剂200的组的赖氨酸特异性外肽酶选择性结合多肽的末端赖氨酸残基。如图(I)及图(II)下方的信号迹线中所展示,信号输出通过显示信号强度增加来报告此结合作用,信号强度增加可用于鉴定赖氨酸特异性外肽酶的发光标记,从而鉴定末端氨基酸。图(III)示出了在选择性结合末端氨基酸之后,带标记的肽酶裂解末端氨基酸。因此,这些组分可自由扩散远离用于发光检测的观测区,其在信号输出中由信号强度下降报告,如图(III)下方的迹线中所展示。图(IV)至图(IX)类似于如针对图(I)至图(III)所描述的过程进行。亦即,带标记的外肽酶结合并裂解对应末端氨基酸,以分别在信号输出中产生对应增加及减少。
在一些方面中,本申请提供了通过评估末端氨基酸与带标记的氨基酸识别分子(例如,带标记的亲和试剂)及带标记的裂解试剂(例如,带标记的非特异性外肽酶)的结合相互作用,进行实时多肽测序的方法。图3A展示了测序方法的实施例,其中离散结合事件引起信号输出300的信号脉冲。图3A的插图示出了通过此方法实时测序的通用方案。如所展示,带标记的亲和试剂310与末端氨基酸(此处展示为赖氨酸)选择性缔合(例如,结合)并从末端氨基酸解离,引起信号输出300中的脉冲系列,脉冲系列可用于鉴定末端氨基酸。在一些实施方案中,脉冲系列提供脉冲图案(例如,特征图案),其可诊断对应末端氨基酸的身分。
在不希望受理论束缚的情况下,带标记的亲和试剂310根据结合亲和力(K
如插图中所展示,测序反应混合物进一步包括带标记的非特异性外肽酶320,其包括不同于带标记的亲和试剂310的发光标记的发光标记。在一些实施方案中,带标记的非特异性外肽酶320以低于带标记的亲和试剂310的浓度的浓度存在于混合物中。在一些实施方案中,带标记的非特异性外肽酶320显示宽特异性,使得其裂解大部分或所有类型的末端氨基酸。因此,动态测序方法可涉及在由外肽酶裂解活性催化的降解反应的过程中,在多肽的末端监测亲和试剂结合。
如由信号输出300的进展所示出,在一些实施方案中,由带标记的非特异性外肽酶320进行的末端氨基酸裂解引起信号脉冲,并且与带标记的亲和试剂310的结合脉冲相比,这些事件以较低频率出现。以此方式,可在实时测序过程中对多肽的氨基酸进行计数和/或鉴定。如信号输出300中进一步示出,在一些实施方案中,可使用多种带标记的亲和试剂,其各自具有诊断性脉冲图案(例如,特征图案),诊断性脉冲图案可用于鉴定对应末端氨基酸。举例而言,在一些实施方案中,不同特征图案(如在信号输出300中由赖氨酸、苯丙氨酸及谷氨酰胺中的每一种所示出的)对应于多于一种带标记的亲和试剂与不同类型的末端氨基酸的缔合。如本文所描述,应了解,根据本申请可使用与多于一种类型的氨基酸缔合的单一亲和试剂。因此,在一些实施方案中,不同特征图案对应于一种带标记的亲和试剂与不同类型的末端氨基酸的缔合。
如本文所描述,基于信号脉冲系列中的特征图案,信号脉冲信息可用于鉴定氨基酸。在一些实施方案中,特征图案包括多个信号脉冲,各信号脉冲包括脉冲持续时间。在一些实施方案中,多个信号脉冲的特征可为特征图案中脉冲持续时间的分布的概括统计量(summary statistic)(例如,平均值、中值、时间衰变常数)。在一些实施方案中,特征图案的平均脉冲持续时间在约1毫秒与约10秒之间(例如,在约1ms与约1s之间、在约1ms与约100ms之间、在约1ms与约10ms之间、在约10ms与约10s之间、在约100ms与约10s之间、在约1s与约10s之间、在约10ms与约100ms之间或在约100ms与约500ms之间)。在一些实施方案中,对应于单一多肽中不同类型的氨基酸的不同特征图案可基于概括统计量中的统计显著差异区别于彼此。举例而言,在一些实施方案中,可基于至少10毫秒(例如,在约10ms与约10s之间、在约10ms与约1s之间、在约10ms与约100ms之间、在约100ms与约10s之间、在约1s与约10s之间或在约100ms与约1s之间)的平均脉冲持续时间差异,区别一种特征图案与另一特征图案。应了解,在一些实施方案中,不同特征图案之间平均脉冲持续时间中的较小差异可能需要各特征图案内较大数目的脉冲持续时间,以便以统计置信度区别一种特征图案与另一特征图案。
如上文所详述,如图3A所示出的实时测序过程一般可涉及末端氨基酸识别及末端氨基酸裂解的循环,其中识别及裂解的相对发生率可受带标记的亲和试剂310与带标记的非特异性外肽酶320之间的浓度差(concentration differential)控制。在一些实施方案中,浓度差可被最佳化,以使得在单独氨基酸识别期间所检测到的信号脉冲的数目为鉴定提供所需的置信区间。举例而言,若初始测序反应提供的信号数据具有过少的裂解事件之间的信号脉冲,以致于不允许以所需置信区间确定特征图案,则可使用相对于亲和试剂浓度降低的非特异性外肽酶重复测序反应。
在一些实施方案中,根据本申请的多肽测序可通过使多肽与测序反应混合物接触进行,该测序反应混合物包括一种或多种氨基酸识别分子(例如,亲和试剂)和/或一种或多种裂解试剂(例如,外肽酶)。在一些实施方案中,测序反应混合物以在约10nM与约10μM之间的浓度包括氨基酸识别分子。在一些实施方案中,测序反应混合物以在约500nM与约500μM之间的浓度包括裂解试剂。
在一些实施方案中,测序反应混合物以以下浓度包括氨基酸识别分子:在约100nM与约10μM之间、在约250nM与约10μM之间、在约100nM与约1μM之间、在约250nM与约1μM之间、在约250nM与约750nM之间或在约500nM与约1μM之间。在一些实施方案中,测序反应混合物以约100nM、约250nM、约500nM、约750nM或约1μM的浓度包括氨基酸识别分子。
在一些实施方案中,测序反应混合物以以下浓度包括裂解试剂:在约500nM与约250μM之间、在约500nM与约100μM之间、在约1μM与约100μM之间、在约500nM与约50μM之间、在约1μM与约100μM之间、在约10μM与约200μM之间或在约10μM与约100μM之间。在一些实施方案中,测序反应混合物以约1μM、约5μM、约10μM、约30μM、约50μM、约70μM或约100μM的浓度包括裂解试剂。
在一些实施方案中,测序反应混合物以在约10nM与约10μM之间的浓度包括氨基酸识别分子,且以在约500nM与约500μM之间的浓度包括裂解试剂。在一些实施方案中,测序反应混合物以在约100nM与约1μM之间的浓度包括氨基酸识别分子,且以在约1μM与约100μM之间的浓度包括裂解试剂。在一些实施方案中,测序反应混合物以在约250nM与约1μM之间的浓度包括氨基酸识别分子,且以在约10μM与约100μM之间的浓度包括裂解试剂。在一些实施方案中,测序反应混合物以约500nM的浓度包括氨基酸识别分子,且以在约25μM与约75μM之间的浓度包括裂解试剂。
在一些实施方案中,测序反应混合物以约500:1、约400:1、约300:1、约200:1、约100:1、约75:1、约50:1、约25:1、约10:1、约5:1、约2:1或约1:1的比率包括氨基酸识别分子及裂解试剂。在一些实施方案中,测序反应混合物以在约10:1与约200:1之间的比率包括氨基酸识别分子及裂解试剂。在一些实施方案中,测序反应混合物以在约50:1与约150:1之间的比率包括氨基酸识别分子及裂解试剂。
尽管由图3A示出的实施例涉及使用带标记的裂解试剂的测序过程,但测序过程并不意欲在此方面中受限。如本文别处所描述,诸位发明人已证明了使用不带标记的裂解试剂的单分子测序。在一些实施方案中,裂解试剂移除连续末端氨基酸的大约频率是已知的,例如基于所使用的酶的已知活性和/或浓度。在一些实施方案中,由试剂进行的末端氨基酸裂解例如基于针对氨基酸识别检测到的信号或缺少检测到的信号推断。诸位发明人已认识到用于控制实时测序反应的另外技术,其可与如所描述的浓度差方法组合使用,或可替代浓度差方法使用。
温度依赖性实时测序过程的实施例展示于图3B中。图(I)至图(III)示出了测序反应,其涉及温度依赖性末端氨基酸识别及末端氨基酸裂解的循环。测序反应的各循环在两个温度范围内进行:第一温度范围(“T
在一些实施方案中,第一温度范围T
在一些实施方案中,第一温度范围T
图(I)描绘了在第一温度范围T
图(II)描绘了在第二温度范围T
图(III)描绘了测序反应中下一循环的开始,其中反应混合物温度已降低回至第一温度范围T
因此,动态测序方法可涉及反应循环,其在反应混合物内的一种或多种蛋白质的蛋白质活性或功能的水平上受控。应了解,图3B中所描绘且上文所描述的温度依赖性多肽测序过程可说明通过条件依赖性识别及裂解的受控循环进行多肽测序的一般方法。举例而言,在一些实施方案中,本申请提供了使用发光活化试剂的发光依赖性测序过程。在一些实施方案中,发光依赖性测序过程涉及发光依赖性氨基酸识别及裂解的循环。测序反应的各循环可通过使测序反应混合物暴露于两种不同发光条件进行:第一发光条件,相比于外肽酶活性,其对于亲和试剂活性而言最佳(例如,以促进氨基酸识别);及第二发光条件,相比于亲和试剂活性,其对于外肽酶活性而言最佳(例如,以促进氨基酸裂解)。测序反应通过在使反应混合物暴露于第一发光条件(以启动氨基酸识别)与使反应混合物暴露于第二发光条件(以启动氨基酸裂解)之间交替而进展。借助于实施例而非限制,在一些实施方案中,两种不同发光条件包括第一波长及第二波长。
在一些方面中,本申请提供了通过评估一种或多种带标记的亲和试剂与末端及内部氨基酸的结合相互作用,及带标记的非特异性外肽酶与末端氨基酸的结合相互作用,进行实时多肽测序的方法。图4展示了测序方法的实施例,其中针对图3A至图3B中的方法描述且说明的方法通过使用带标记的亲和试剂410修改,带标记的亲和试剂选择性结合至以下且从以下解离:末端及内部位置两者处的一种类型的氨基酸(此处展示为赖氨酸)(图4,插图)。如先前方法中所描述,选择性结合引起信号输出400中的脉冲系列。然而,在此方法中,脉冲系列以一定比率出现,该比率由整个多肽中该类型的氨基酸的数目决定。因此,在一些实施方案中,对应于结合事件的脉冲速率将诊断多肽中当前存在的同源氨基酸的数目。
如在先前方法中,与带标记的亲和试剂410相比,带标记的非特异性肽酶420将以相对较低浓度存在,例如以提供在裂解事件之间的最佳时间窗(图4,插图)。另外,在某些实施方案中,带标记的非特异性肽酶420的可唯一鉴定的发光标记将指示裂解事件何时发生。随着多肽经历迭代裂解,每当末端氨基酸被带标记的非特异性肽酶420裂解时,对应于被带标记的亲和试剂410结合的脉冲速率都将以逐步方式下降。此概念由曲线图402说明,该曲线图大体上描绘了随时间函数变化的脉冲速率,其中实时裂解事件由箭头表示。因此,在一些实施方案中,可基于脉冲图案和/或在裂解事件之间所检测到的图案内存在的脉冲速率,以此方法鉴定氨基酸,并从而对多肽进行测序。
在一些实施方案中,末端多肽序列信息(例如,如本文所描述确定)可与获自一种或多种其他来源的多肽序列信息组合。举例而言,末端多肽序列信息可与内部多肽序列信息组合。在一些实施方案中,如本文所描述,可使用与内部氨基酸缔合的一种或多种氨基酸识别分子获得内部多肽序列信息。内部或其他多肽序列信息可在多肽降解过程之前或过程中获得。在一些实施方案中,获自这些方法的序列信息可与使用其他技术的多肽序列信息,例如使用一种或多种内部氨基酸识别分子获得的序列信息组合。
被屏蔽的识别分子
根据本文所描述的实施例,可通过用激发光照射表面固定多肽,并检测由连接至氨基酸识别分子(例如,带标记的亲和试剂)的标记产生的发光,来进行单分子多肽测序方法。在一些情况下,由标记产生的辐射和/或非辐射衰变可导致对多肽的光损伤。举例而言,图5A示出了示例性测序反应,其中展示了识别分子与固定至表面的多肽缔合。
在激发照射存在下,标记可经由辐射衰变产生荧光,引起可检测的缔合事件。然而,在一些情况下,标记产生非辐射衰变,可导致活性氧物质500的形成。活性氧物质500可最终损伤固定肽,使得反应在获得多肽的完整序列信息之前结束。此光损伤可例如在暴露的多肽末端(顶部空心箭头)、在内部位置(中间空心箭头)或在将多肽连接至表面的表面连接子(底部空心箭头)处发生。
诸位发明人已发现,可通过将屏蔽元件并入至氨基酸识别分子中,来减轻光损伤且延长识别时间。图5B示出了使用被屏蔽的识别分子的示例性测序反应,该被屏蔽的识别分子包括屏蔽物502。屏蔽物502形成共价或非共价连接基团,其在标记与多肽之间提供增加的距离,使得来自活性氧物质500的损伤效应可归因于在标记-多肽间隔距离上的自由基衰变而减小。屏蔽物502也可提供空间屏障,其通过吸收来自活性氧物质500及辐射和/或非辐射衰变的损伤来屏蔽多肽免受标记影响。
在不希望受理论束缚的情况下,认为定位于识别组分与标记组分之间的屏蔽物可吸收、偏转或以其他方式阻断由标记组分发射的辐射和/或非辐射衰变。在一些实施方案中,屏蔽物阻止一种或多种标记(例如,发光标记)与一种或多种氨基酸识别分子相互作用,或限制相互作用的程度。在一些实施方案中,屏蔽物阻止一种或多种标记与氨基酸识别分子缔合的一种或多种分子(例如,与识别分子缔和的多肽)相互作用,或限制相互作用的程度。因此,在一些实施方案中,术语屏蔽一般可以指由识别组分与标记组分之间形成的连接基团的某一部分提供的保护或屏蔽效应。
在一些实施方案中,屏蔽物连接至一种或多种氨基酸识别分子(例如,识别组分)且连接至一种或多种标记(例如,标记组分)。在一些实施方案中,识别及标记组分在屏蔽物上不相邻位点处连接。举例而言,一种或多种氨基酸识别分子可连接至屏蔽物的第一侧,且一种或多种标记可连接至屏蔽物的第二侧,其中屏蔽物的第一侧与第二侧彼此远离。在一些实施方案中,连接位点在屏蔽物的大约对侧上。
屏蔽物连接至识别分子的位点与屏蔽物连接至标记的位点之间的距离可为穿过空间的线性测量值,或跨越屏蔽物的表面的非线性测量值。屏蔽物上识别分子连接位点与标记连接位点之间的距离可通过对屏蔽物的三维结构建模测量。在一些实施方案中,此距离可为至少2nm、至少4nm、至少6nm、至少8nm、至少10nm、至少12nm、至少15nm、至少20nm、至少30nm、至少40nm或更大。可替代地,可通过将屏蔽物的结构视为二次曲面(例如,椭球面、椭圆柱面),来描述屏蔽物上识别分子与标记的相对位置。在一些实施方案中,识别分子连接位点与标记连接位点间隔开的距离为环绕表示屏蔽物的椭球形形状的距离的至少八分之一。在一些实施方案中,识别分子与标记间隔开的距离为环绕表示屏蔽物的椭球形形状的距离的至少四分之一。在一些实施方案中,识别分子与标记间隔开的距离为环绕表示屏蔽物的椭球形形状的距离的至少三分之一。在一些实施方案中,识别分子与标记间隔开的距离为环绕表示屏蔽物的椭球形形状的距离的一半。
屏蔽物的大小应使得当氨基酸识别分子与多肽缔合时,标记不能或不大可能直接接触多肽。屏蔽物的大小也应使得当氨基酸识别分子与多肽缔合时,所连接的标记可检测。举例而言,大小应使得所连接的发光标记在待激发的照明体积内。
应了解,从业者可根据多种参数评估屏蔽效应。一般而言,可通过在具有屏蔽元件的组合物与不具有屏蔽元件的组合物之间进行比较性评定,来评估屏蔽元件的作用。举例而言,屏蔽元件可延长氨基酸识别分子的识别时间。在一些实施方案中,识别时间是指在如本文所描述的多肽测序反应中,可观测到识别分子与多肽之间的缔合事件的时间长度。在一些实施方案中,与除了氨基酸识别分子不具有屏蔽元件但在其他方面相似或相同,在相同条件下进行的多肽测序反应相比,识别时间延长约10%至25%、25%至50%、50%至75%、75%至100%或超过100%,例如延长约2倍、3倍、4倍、5倍或更多。在一些实施方案中,屏蔽元件可增加测序准确度和/或序列读段长度(例如,相对于在如上文所描述的比较条件下进行的测序反应,增加至少5%、至少10%、至少15%、至少25%或更多)。
因此,在一些方面中,本申请提供了被屏蔽的识别分子,其包括至少一种氨基酸识别分子,至少一种可检测标记,及在识别分子与标记之间形成共价或非共价连接基团的屏蔽元件(例如,“屏蔽物”)。在一些实施方案中,屏蔽元件的长度为至少2nm、至少5nm、至少10nm、至少12nm、至少15nm、至少20nm或更大(例如,在水溶液中)。在一些实施方案中,屏蔽元件的长度在约2nm与约100nm之间(例如,在约2nm与约50nm之间、在约10nm与约50nm之间、在约20nm与约100nm之间)。
在一些实施方案中,屏蔽物(例如,屏蔽元件)在一种或多种氨基酸识别分子(例如,识别组分)与一种或多种标记(例如,标记组分)之间形成共价或非共价连接基团。如本文所用,在一些实施方案中,共价及非共价连接或连接基团是指识别组分及标记组分与屏蔽物的连接的性质。
在一些实施方案中,共价连接或共价连接基团是指经共价键或一系列相邻共价键连接至识别组分及标记组分中的每一个的屏蔽物。共价连接一或两个组分可通过本领域中已知的共价缀合方法来达成。举例而言,在一些实施方案中,点击化学(click chemistry)技术(例如,铜催化、应力促进(strain-promoted)、无铜点击化学等)可用于将一或两个组分连接至屏蔽物。此类方法一般涉及将一个反应性部分缀合至另一反应性部分,以在反应性部分之间形成一个或多个共价键。因此,在一些实施方案中,可使屏蔽物的第一反应性部分与识别或标记组分的第二反应性部分接触,以形成共价连接。反应性部分的实施例包括(但不限于)反应性氨、叠氮化合物、炔烃、硝酮、烯烃(例如,环烯烃)、四嗪、四唑及适于点击反应及类似偶联技术的其他反应性部分。
在一些实施方案中,非共价连接或非共价连接基团是指经由一种或多种非共价偶联手段,包括(但不限于)受体-配体相互作用及寡核苷酸链杂交,连接至识别组分及标记组分中的一或两个的屏蔽物。本文提供了受体-配体相互作用的实施例,并且其包括(但不限于)蛋白质-蛋白质复合物、蛋白质-配体复合物、蛋白质-适体复合物及适体-核酸复合物。用于寡核苷酸链杂交的各种构造及策略描述于本文中并且是本领域中已知的(参见例如美国专利公开案第2019/0024168号)。
在一些实施方案中,屏蔽物502包括聚合物,诸如生物分子或树状聚合物。图5C描绘了本申请的聚合物屏蔽物及被屏蔽的识别分子的构造的实施例。第一被屏蔽的构建体504展示了蛋白质屏蔽物530的实施例。在一些实施方案中,蛋白质屏蔽物530在识别分子与标记之间形成共价连接基团。举例而言,在一些实施方案中,蛋白质屏蔽物530经一个或多个共价键,例如通过经蛋白质屏蔽物530的天然或非天然氨基酸的侧链共价连接,连接至识别分子及标记中的每一个。在一些实施方案中,蛋白质屏蔽物530在识别分子与标记之间形成非共价连接基团。举例而言,在一些实施方案中,蛋白质屏蔽物530为包括一个或多个配体结合位点的单体或多聚蛋白。在一些实施方案中,非共价连接基团经结合至一个或多个配体结合位点的一个或多个配体部分形成。由蛋白质屏蔽物形成的非共价连接的额外实施例描述于本文别处。
第二被屏蔽的构建体506展示了双链核酸屏蔽物的实施例,双链核酸屏蔽物包括第一寡核苷酸链532,其与第二寡核苷酸链534杂交。如所展示,在一些实施方案中,双链核酸屏蔽物可包括连接至第一寡核苷酸链532的识别分子,及连接至第二寡核苷酸链534的标记。以此方式,双链核酸屏蔽物经由寡核苷酸链杂交在识别分子与标记之间形成非共价连接基团。在一些实施方案中,识别分子及标记可连接至相同寡核苷酸链,这可提供单链核酸屏蔽物,或经由与另一寡核苷酸链杂交,提供双链核酸屏蔽物。在一些实施方案中,链杂交可在连接基团内提供提高的刚性,以进一步增强识别分子与标记之间之间隔。
在屏蔽元件502包括核酸时,标记与识别分子之间的间隔距离可由核酸上连接位点(例如,直接连接或间接连接,诸如经一种或多种额外屏蔽聚合物连接)之间的距离测量。在一些实施方案中,核酸上连接位点之间的距离可由标记与识别分子之间出现的核酸内的核苷酸的数目测量。应理解,核苷酸的数目可以指单链核酸中的核苷酸碱基的数目,或双链核酸中的核苷酸碱基对的数目。
因此,在一些实施方案中,识别分子的连接位点与标记的连接位点可由在5个与200个之间的核苷酸(例如,在5个与150个之间的核苷酸、在5个与100个之间的核苷酸、在5个与50个之间的核苷酸、在10个与100个之间的核苷酸)间隔开。应了解,核酸中的任何位置可充当识别分子、标记或一种或多种额外聚合屏蔽物的连接位点。在一些实施方案中,连接位点可处于或大约处于5'或3'端,或处于沿核酸的链的内部位置。
第二被屏蔽的构建体506的非限制性构造示出了屏蔽物的实施例,其经由链杂交形成非共价连接。非共价连接的另一实施例由第三被屏蔽的构建体508示出,其包括寡核苷酸屏蔽物536。在一些实施方案中,寡核苷酸屏蔽物536为核酸适体,其结合识别分子以形成非共价连接。在一些实施方案中,识别分子为核酸适体,且寡核苷酸屏蔽物536包括寡核苷酸链,其与适体杂交以形成非共价连接。
第四被屏蔽的构建体510展示了树状聚合物屏蔽物538的实施例。如本文所用,在一些实施方案中,树状聚合物一般是指多元醇或树状体。多元醇及树状体已描述于本领域中,且可包括针对特定构造最佳化的分支链树状结构。在一些实施方案中,树状聚合物屏蔽物538包括聚乙二醇、四乙二醇、聚(酰氨基氨)、聚(丙烯亚氨)、聚(丙烯氨)、碳硅烷、聚(L-赖氨酸)或其一种或多种的组合。
树状体或树状突(dendron)为重复分支链分子,其通常围绕核对称,且可采用球形三维形态。参见例如Astruc等人(2010)Chem.Rev.110:1857。将此类结构并入至本申请的屏蔽物中可以通过标记与与其缔合的一种或多种生物分子(例如,识别分子和/或与识别分子缔合的多肽)之间的接触的空间抑制,来提供保护效应。经由分子的一级结构的变化(包括树状体表面的潜在官能化)改进树状体的化学及物理特性,允许按需要调节屏蔽效应。如本领域中已知,树状体可使用广泛范围的材料及分支链反应通过多种技术合成。此类合成变化允许视需要定制树状体的特性。可根据本申请的屏蔽物使用的多元醇及树状体化合物的实施例包括(但不限于)美国专利公开案第20180346507号中所描述的化合物。
图5D描绘了本申请的被屏蔽的识别分子的另外示例性构造。蛋白质-核酸构建体512展示了屏蔽物的实施例,其包括呈蛋白质及双链核酸形式的多于一种聚合物。在一些实施方案中,屏蔽物的蛋白质部分经由共价连接连接至屏蔽物的核酸部分。在一些实施方案中,连接是经由非共价连接。举例而言,在一些实施方案中,屏蔽物的蛋白质部分为单价或多价蛋白,其经由配体部分形成至少一个非共价连接,该配体部分连接至该单价或多价蛋白的配体结合位点。在一些实施方案中,屏蔽物的蛋白质部分包括抗生物素蛋白。
在一些实施方案中,本申请的被屏蔽的识别分子为抗生物素-核酸构建体514。在一些实施方案中,抗生物素-核酸构建体514包括屏蔽物,其包括抗生物素蛋白540及双链核酸。如本文中所描述,抗生物素蛋白540可用于在一种或多种氨基酸识别分子与一种或多种标记之间,直接地或间接地(诸如经本文所描述的一种或多种额外屏蔽聚合物)形成非共价连接。
抗生物素蛋白为生物素结合蛋白,一般在抗生物素蛋白的四个亚单元中的每一个处具有生物素结合位点。抗生物素蛋白包括例如抗生物素、抗生物素蛋白链菌素、突变抗生物素蛋白链菌素(traptavidin)、金顶侧耳抗生物素(tamavidin)、慢生型根瘤菌抗生物素(bradavidin)、爪蟾抗生物素(xenavidin)及其同系物及变体。在一些情况下,可使用抗生物素蛋白的单体、二聚或四聚形式。在一些实施方案中,抗生物素蛋白复合物的抗生物素蛋白为呈四聚形式(例如,均四聚体)的抗生物素蛋白链菌素。在一些实施方案中,抗生物素蛋白的生物素结合位点为本文所描述的一种或多种氨基酸识别分子、一种或多种标记和/或一种或多种额外屏蔽聚合物提供连接位点。
抗生物素蛋白复合物的说明图解展示于图5D的插图中。如插图中所展示,抗生物素蛋白540可在蛋白质的四个亚单元中的每一个处包括结合位点542,其可结合至生物素部分(展示为白色圆形)。抗生物素蛋白540的多价可允许各种连接构造,其出于说明的目的大体上展示。举例而言,在一些实施方案中,生物素连接部分544可用于提供与抗生物素蛋白540的单一连接点。在一些实施方案中,双生物素连接部分546可用于提供两个与抗生物素蛋白540的连接点。如由抗生物素-核酸构建体514所示出,抗生物素蛋白复合物可由两个双生物素连接部分形成,两个双生物素连接部分形成反式构造,以在识别分子与标记之间提供增加的间隔距离。
展示了抗生物素蛋白屏蔽物组态的各种另外的实施例。第一抗生物素构建体516展示了抗生物素屏蔽物的实施例,其经双生物素连接部分连接至识别分子,且经独立的生物素连接部分连接至两个标记。第二抗生物素构建体518展示抗生物素屏蔽物的实施例,其经独立的生物素连接部分连接至两个识别分子,且经双生物素连接部分连接至标记。第三抗生物素构建体520展示抗生物素屏蔽物的实施例,其经独立的生物素连接部分连接至两个识别分子,且经核酸的各链的生物素连接部分连接至带标记的核酸。第四抗生物素构建体522展示抗生物素屏蔽物的实施例,其经独立的双生物素连接部分连接至识别分子且连接至带标记的核酸。如所展示,标记由标记与核酸之间的树状聚合物进一步与识别分子屏蔽。
应了解,图5A至图5D中所展示的被屏蔽的识别分子的示例性构造出于说明的目的提供。诸位发明人已构想使用一种或多种不同聚合物的各种其他屏蔽物构造,这些聚合物在被屏蔽的识别分子的识别组分与标记组分之间形成共价或非共价连接。借助于实施例,图5E示出了根据本申请的屏蔽物构造的模块化。
如图5E顶部处所展示,被屏蔽的识别分子一般包括识别组分550、屏蔽元件552及标记组分554。为了易于说明,识别组分550描绘为一种氨基酸识别分子,且标记组分554描绘为一种标记。应了解,本申请的被屏蔽的识别分子可包括屏蔽元件552,其连接至一种或多种氨基酸识别分子及一种或多种标记。在识别组分550包括多于一种识别分子时,各识别分子可在屏蔽元件552上的一个或多个连接位点处连接至屏蔽元件552。在标记组分554包括多于一种标记时,各标记可在屏蔽元件552上的一个或多个连接位点处连接至屏蔽元件552。
在一些实施方案中,屏蔽元件552包括蛋白质560。在一些实施方案中,蛋白质560为单价或多价蛋白。在一些实施方案中,蛋白质560为单体或多聚蛋白,诸如蛋白质同二聚体、蛋白质异二聚体、蛋白质寡聚物或其他蛋白质(proteinaceous)分子。在一些实施方案中,屏蔽元件552包括蛋白质复合物,其由非共价结合至至少一种其他分子的蛋白质形成。举例而言,在一些实施方案中,屏蔽元件552包括蛋白质-蛋白质复合物562。在一些实施方案中,蛋白质-蛋白质复合物562包括一种特异性结合至另一蛋白质分子的蛋白质分子。在一些实施方案中,蛋白质-蛋白质复合物562包括结合至抗原的抗体或抗体片段(例如,scFv)。在一些实施方案中,蛋白质-蛋白质复合物562包括结合至蛋白质配体的受体。蛋白质-蛋白质复合物的额外实施例包括(但不限于)胰蛋白酶-抑肽酶(trypsin-aprotinin)、雄性不育基因-恢复基因(barnase-barstar)及大肠杆菌素(colicin)E9-Im9免疫蛋白。
在一些实施方案中,屏蔽元件552包括蛋白质-配体复合物564。在一些实施方案中,蛋白质-配体复合物564包括单价蛋白质及非蛋白质配体部分。举例而言,在一些实施方案中,蛋白质-配体复合物564包括结合至小分子抑制剂部分的酶。在一些实施方案中,蛋白质-配体复合物564包括结合至非蛋白质配体部分的受体。
在一些实施方案中,屏蔽元件552包括多价蛋白质复合物,其由非共价结合至一种或多种配体部分的多价蛋白质形成。在一些实施方案中,屏蔽元件552包括抗生物素蛋白复合物,其由非共价结合至一种或多种生物素连接部分的抗生物素蛋白形成。构建体566、568、570及572提供抗生物素蛋白复合物的说明性实施例,其中的任何一种或多种可并入至屏蔽元件552中。
在一些实施方案中,屏蔽元件552包括双向抗生物素复合物566,其包括结合至两个双生物素连接部分的抗生物素蛋白。在一些实施方案中,屏蔽元件552包括三向抗生物素复合物568,其包括结合至两个生物素连接部分及一个双生物素连接部分的抗生物素蛋白。在一些实施方案中,屏蔽元件552包括四向抗生物素复合物570,其包括结合至四个生物素连接部分的抗生物素蛋白。
在一些实施方案中,屏蔽元件552包括抗生物素蛋白,其包括工程改造到抗生物素蛋白中的一或两个非官能性结合位点。举例而言,在一些实施方案中,屏蔽元件552包括二价抗生物素复合物572,其包括在两个亚单元中的每一个处结合至生物素连接部分的抗生物素蛋白,其中抗生物素蛋白在两个其他亚单元中的每一个处包括非官能性配体结合位点548。如所展示,在一些实施方案中,二价抗生物素复合物572包括反-二价抗生物素蛋白,不过可视所需实施方案而定使用顺-二价抗生物素蛋白。在一些实施方案中,抗生物素蛋白为三价抗生物素蛋白。在一些实施方案中,三价抗生物素蛋白在一个亚单元处包括非官能性配体结合位点548,且在其他亚单元处结合至三个生物素连接部分,或一个生物素连接部分及一个双生物素连接部分。
在一些实施方案中,屏蔽元件552包括树状聚合物574。在一些实施方案中,树状聚合物574为多元醇或树状体,如文本别处所描述。在一些实施方案中,树状聚合物574为分支链多元醇或分支链树状体。在一些实施方案中,树状聚合物574包括单糖-TEG、二糖、N-乙酰基单糖、TEMPO-TEG、水溶性维生素E(trolox)-TEG或甘油树状体。根据本申请的被屏蔽的识别分子适用的多元醇的实施例包括聚醚多元醇及聚酯多元醇,例如聚乙二醇、聚丙二醇及本领域中熟知的类似的此类聚合物。在一些实施方案中,树状聚合物574包括以下式的化合物:-(CH
在一些实施方案中,屏蔽元件552包括核酸。在一些实施方案中,核酸为单链的。在一些实施方案中,标记组分554直接地或间接地连接至单链核酸的一端(例如,5'端或3'端),且识别组分550直接地或间接地连接至单链核酸的另一端(例如,3'端或5'端)。举例而言,单链核酸可包括连接至核酸的5'端的标记,及连接至核酸的3'端的氨基酸识别分子。
在一些实施方案中,屏蔽元件552包括双链核酸576。如所展示,在一些实施方案中,双链核酸576可经由链杂交在识别组分550与标记组分554之间形成非共价连接。然而,在一些实施方案中,双链核酸576可经由与相同寡核苷酸链的连接在识别组分550与标记组分554之间形成共价连接。在一些实施方案中,标记组分554直接地或间接地连接至双链核酸的一端,且识别组分550直接地或间接地连接至双链核酸的另一端。举例而言,双链核酸可包括连接至一链的5'端的标记,及连接至另一链的5'端的氨基酸识别分子。
在一些实施方案中,屏蔽元件552包括核酸,其形成可适用于增加屏蔽物的空间体积(steric bulk)的一种或多种结构基元。核酸结构基元的实施例包括(但不限于)茎-环、三向连结体(例如,由两个或更多个茎-环基元形成)、四向连结体(例如,霍利迪连结体(Holliday junction))及凸环(bulge loop)。
在一些实施方案中,屏蔽元件552包括形成茎-环578的核酸。茎-环或发夹环为当寡核苷酸链折叠且与相同链的另一段形成碱基对时形成的寡核苷酸链上的核苷酸的未配对环。在一些实施方案中,茎-环578的未配对环包括三至十个核苷酸。因此,茎-环578可由具有杂交形成茎的反向互补序列的寡核苷酸链的两个区域形成,其中两个区域由形成未配对环的三至十个核苷酸间隔开。在一些实施方案中,茎-环578的茎可被设计以具有一个或多个G/C核苷酸,相比于A/T核苷酸,其可通过形成的附加氢键结相互作用而提供增加的稳定性。在一些实施方案中,茎-环578的茎包括紧邻未配对环序列的G/C核苷酸。在一些实施方案中,茎-环578的茎在与未配对环序列相邻之前2、3、4或5个核苷酸内包括G/C核苷酸。在一些实施方案中,茎-环578的未配对环包括一个或多个连接位点。在一些实施方案中,连接位点存在于未配对环中的无碱基位点处。在一些实施方案中,连接位点存在于未配对环的碱基处。
在一些实施方案中,茎-环578由双链核酸形成。如本文所描述,在一些实施方案中,双链核酸可经由第一寡核苷酸链与第二寡核苷酸链的链杂交形成非共价连接基团。然而,在一些实施方案中,屏蔽元件552包括单链核酸,其形成茎-环基元,例如以提供共价连接基团。在一些实施方案中,屏蔽元件552包括形成两个或更多个茎-环基元的核酸。举例而言,在一些实施方案中,核酸包括两个茎-环基元。在一些实施方案中,一个茎-环基元的茎与另一茎-环基元的茎相邻,使得基元共同形成三向连结体。在一些实施方案中,屏蔽元件552包括形成四向连结体578的核酸。在一些实施方案中,四向连结体578经由两个或更多个寡核苷酸链(例如,2、3或4个寡核苷酸链)的杂交形成。
在一些实施方案中,屏蔽元件552包括一种或多种选自图5E的560、562、564、566、568、570、572、574、576、578及580的聚合物。应了解,560、562、564、566、568、570、572、574、576、578及580中的每一个上所展示的连接部分及连接位点出于说明的目的被展示,且并不意欲描绘较佳连接或连接位点构造。
在一些方面中,本申请提供了式(I)的氨基酸识别分子:
A-(Y)
(I),
其中:A是包括至少一种氨基酸识别分子的氨基酸结合组分;Y在各情况下是形成共价或非共价连接基团的聚合物;n是1至10的整数(包括端点);且D是包括至少一种可检测标记的标记组分。在一些实施方案中,本申请提供了包括式(I)的可溶性氨基酸识别分子的组合物。
在一些实施方案中,A包括多种氨基酸识别分子。在一些实施方案中,多种氨基酸识别分子中的各氨基酸识别分子连接至Y上的不同连接位点。在一些实施方案中,多种氨基酸识别分子中的至少两种氨基酸识别分子连接至Y上的单一连接位点。在一些实施方案中,氨基酸识别分子为识别蛋白或核酸适体,例如如本文别处所描述。
在一些实施方案中,可检测标记为发光标记或导电性标记。在一些实施方案中,发光标记包括至少一种荧光团染料分子。在一些实施方案中,D包括20种或更少荧光团染料分子。在一些实施方案中,荧光团染料分子的数目与氨基酸识别分子的数目的比率在1:1与20:1之间。在一些实施方案中,发光标记包括至少一个FRET对,其包括供体标记及受体标记。在一些实施方案中,供体标记与受体标记的比率为1:1、2:1、3:1、4:1或5:1。在一些实施方案中,受体标记与供体标记的比率为1:1、2:1、3:1、4:1或5:1。
在一些实施方案中,D的直径小于
在一些实施方案中,氨基酸识别分子是以下式中的一个:
A-Y
其中:Y
在一些实施方案中,核酸包括第一寡核苷酸链。在一些实施方案中,核酸包括与第一寡核苷酸链杂交的第二寡核苷酸链。在一些实施方案中,核酸经第一寡核苷酸链形成共价连接。在一些实施方案中,核酸经杂交的第一寡核苷酸链及第二寡核苷酸链形成非共价连接。
在一些实施方案中,多肽为单价或多价蛋白。在一些实施方案中,单价或多价蛋白通过配体部分形成至少一个非共价连接,所述配体部分连接至单价或多价蛋白的配体结合位点。在一些实施方案中,A、Y或D包括配体部分。
在一些实施方案中,氨基酸识别分子是以下式中的一个:
A-(Y)
其中:Y
在一些方面中,本申请提供了式(II)的氨基酸识别分子:
A-Y
(II),
其中:A是包括至少一种氨基酸识别分子的氨基酸结合组分;Y
在一些实施方案中,Y
在一些实施方案中,Y
在一些实施方案中,氨基酸识别分子是以下式中的一个:
A-Y
其中:Y在各情况下是形成共价或非共价连接基团的聚合物;且n是1至10的整数(包括端点)。在一些实施方案中,Y在各情况下独立地是生物分子、多元醇或树状体。
在其他方面中,本申请提供了氨基酸识别分子,其包括:核酸;连接至核酸上的第一连接位点的至少一种氨基酸识别分子;及连接至核酸上的第二连接位点的至少一种可检测标记。在一些实施方案中,核酸在至少一种氨基酸识别分子与至少一种可检测标记之间形成共价或非共价连接基团。
在一些实施方案中,核酸为双链核酸,其包括与第二寡核苷酸链杂交的第一寡核苷酸链。在一些实施方案中,第一连接位点在第一寡核苷酸链上,且其中第二连接位点在第二寡核苷酸链上。在一些实施方案中,至少一种氨基酸识别分子通过蛋白质连接至第一连接位点,该蛋白质在至少一种氨基酸识别分子与核酸之间形成共价或非共价连接基团。在一些实施方案中,至少一种可检测标记通过蛋白质连接至第二连接位点,该蛋白质在至少一种可检测标记与核酸之间形成共价或非共价连接基团。在一些实施方案中,第一连接位点与第二连接位点在核酸上相隔5个至100个核苷酸碱基或核苷酸碱基对。
在其他方面中,本申请提供了氨基酸识别分子,其包括:多价蛋白,其包括至少两个配体结合位点;至少一种氨基酸识别分子,其通过与蛋白质上的第一配体结合位点结合的第一配体部分连接至蛋白质;及至少一种可检测标记,其通过与蛋白质上的第二配体结合位点结合的第二配体部分连接至蛋白质。
在一些实施方案中,多价蛋白为包括四个配体结合位点的抗生物素蛋白。在一些实施方案中,配体结合位点为生物素结合位点,且其中配体部分为生物素部分。在一些实施方案中,生物素部分中的至少一个为双生物素部分,且其中双生物素部分结合至抗生物素蛋白上的两个生物素结合位点。在一些实施方案中,至少一种氨基酸识别分子通过包含第一配体部分的核酸连接至蛋白质。在一些实施方案中,至少一种可检测标记通过包含第二配体部分的核酸连接至蛋白质。
如本文别处所描述,本申请的被屏蔽的识别分子可用于根据本申请的多肽测序方法,或本领域中已知的任何方法。举例而言,在一些实施方案中,本文所提供的被屏蔽的识别分子可用于本文所提供或本领域中常规的艾德曼型降解反应,其可涉及多肽测序反应中多个反应混合物的迭代循环。在一些实施方案中,本文所提供的被屏蔽的识别分子可用于本申请的动态测序反应,其涉及单一反应混合物中的氨基酸识别及降解。
通过带标记的多肽的降解进行测序
在一些方面中,本申请提供了通过鉴定对应于已知多肽序列的氨基酸的独特组合,对多肽进行测序的方法。举例而言,图6展示了通过检测带标记的多肽600的被选择性标记的氨基酸进行测序的方法。在一些实施方案中,带标记的多肽600包括被选择性修饰的氨基酸,使得不同氨基酸类型包括不同发光标记。如本文所用,除非另外指示,否则带标记的多肽是指包括一种或多种被选择性标记的氨基酸侧链的多肽。选择性标记的方法及涉及带标记的多肽的制备及分析的细节是本领域中已知的(参见例如Swaminathan等人PLoSComput Biol.2015,11(2):e1004080)。
如所展示,在一些实施方案中,带标记的多肽600被固定且曝露于激发源。检测来自带标记的多肽600的聚集发光,且在一些实施方案中,随时间的推移,曝露于发光会导致所检测到的信号由于发光标记降解(例如,由于光漂白(photobleaching)的降解)而损失。在一些实施方案中,带标记的多肽600包括被选择性标记的氨基酸的独特组合,其引起初始检测到的信号。如大体上所示出,随时间推移的发光标记降解导致针对被光漂白的带标记的多肽602检测到的信号的对应降低。在一些实施方案中,可通过分析一种或多种发光特性来解卷积信号(例如,通过发光寿命分析来进行信号解卷积)。在一些实施方案中,带标记的多肽600的被选择性标记的氨基酸的独特组合已在计算上预计算,且在经验上被验证——例如基于蛋白质组的已知多肽序列。在一些实施方案中,将所检测到的氨基酸标记的组合与生物体的蛋白质组的已知序列的数据库进行比较,以鉴定对应于带标记的多肽600的数据库中的特定多肽。
在一些实施方案中,图6中所示出的方法可通过确定用于进行使大规模并行分析中的取样达到最大的测序反应的最佳样品浓度来修改。在一些实施方案中,选择浓度以使得阵列的样品孔的所需分率(例如,30%)在任何给定时间被占据。在不希望受理论束缚的情况下,认为尽管多肽在一段时间被漂白,但相同孔继续可用于进一步分析。经由扩散,每3分钟可将阵列样品孔的大约30%用于分析。作为说明性实施例,在一百万样品孔芯片中,每小时可取样6,000,000个多肽,或在4小时时段内取样24,000,000个。
在一些方面中,本申请提供了通过检测经历了末端氨基酸修饰及裂解的重复循环的带标记的多肽的发光,而对多肽进行测序的方法。举例而言,图7展示了根据本申请的通过艾德曼降解对带标记的多肽进行测序的方法。在一些实施方案中,该方法一般如本文针对通过艾德曼降解进行测序的其他方法所描述进行。举例而言,在一些实施方案中,图7中所展示的步骤(1)及步骤(2)可分别如本文针对末端氨基酸修饰及末端氨基酸裂解所描述,在艾德曼降解反应中进行。
如图7中所描绘的实施例所展示,在一些实施方案中,方法包括步骤(1):修饰带标记的多肽的末端氨基酸。如本文别处所描述,在一些实施方案中,修饰包括使末端氨基酸与异硫氰酸酯(例如,PITC)接触,以形成经异硫氰酸酯修饰的末端氨基酸。在一些实施方案中,异硫氰酸酯修饰710将末端氨基酸转化为对由裂解试剂(例如,化学或酶裂解试剂,如本文所描述)移除更敏感的形式。因此,在一些实施方案中,方法包括步骤(2):使用本文别处针对艾德曼降解详述的化学或酶手段移除被修饰的末端氨基酸。
在一些实施方案中,方法包括重复步骤(1)至步骤(2),持续多个循环,在其期间检测带标记的多肽的发光,并且对应于带标记的氨基酸从末端移除的裂解事件可以检测为所检测到的信号的降低。在一些实施方案中,如图7中所展示的步骤(2)之后信号无变化,可以鉴定是未知类型的氨基酸。因此,在一些实施方案中,部分序列信息可通过在每个相继轮次期间评估步骤(2)之后所检测到的信号来确定,即,通过基于所检测到的信号的变化而确定的身分来指派氨基酸类型,或基于所检测到的信号无变化将氨基酸类型鉴定为未知。
在一些方面中,根据本申请对多肽进行测序的方法包括通过带标记的多肽的进行性酶裂解进行测序,如图8A至图8C中大体上所示出。如所展示,在一些实施方案中,带标记的多肽经历使用被修饰的进行性外肽酶进行的降解,该外肽酶从一个末端向另一末端连续裂解末端氨基酸。本文别处详细描述外肽酶。图8A描绘了实施例,其中带标记的多肽800经历由固定化的进行性外肽酶810进行的降解。图8B描绘了实施例,其中固定化的带标记的多肽820经历由进行性外肽酶830进行的降解。
图8C示意性地示出了根据图8B中所描绘的方法进行的实时测序过程的实施例。如所展示,图(I)至图(IV)展示了带标记的多肽降解的进展,在各图下方展示了随时间推移的对应信号迹线。如所展示,对应于带标记的氨基酸的各裂解事件引起信号中的伴随下降。在一些实施方案中,进行性外肽酶830的进行(processivity)速率已知,使得所检测到的信号降低之间的时序可用于计算各检测事件之间不带标记的氨基酸的数目。举例而言,若40个氨基酸的多肽以使得每秒移除一个氨基酸的方式裂解,则具有3个信号的带标记的多肽将最初展示全部3个((图(I)),随后2个(图(II)),随后1个(图(III))且最后无信号。以此方式,可确定带标记的氨基酸的顺序。因此,这些方法可用于确定部分序列信息,例如用于基于多肽片段测序的蛋白质组分析。
在一些实施方案中,可使用例如如图9中所示出的基于ATP的福斯特共振能量转移(
举例而言,在一些实施方案中,所提取的蛋白质被变性,且半胱氨酸及赖氨酸被荧光染料标记。在一些实施方案中,可使用蛋白质移位酶(例如,细菌ClpX)的工程改造形式来结合单独底物蛋白质,将蛋白质外展,且使蛋白质通过其纳米通道移位。在一些实施方案中,移位酶被供体染料标记,且当底物通过纳米通道时,在移位酶上的供体与底物上的两种或更多种相异受体染料之间发生FRET。随后可根据FRET信号确定带标记的氨基酸的顺序。在一些实施方案中,可使用表5中所展示的以下非限制性带标记的ATP类似物中的一种或多种。
表5.带标记的ATP类似物的非限制性实例。
用于测序的样品的制备
在测序之前,多肽样品可被修饰。在一些实施方案中,多肽的N末端氨基酸或C末端氨基酸被修饰。图10A示出了用于由蛋白质样品制备末端修饰的多肽的末端修饰的非限制性实例。在步骤(1)中,将蛋白质样品1000片段化以产生多肽片段1002。可通过裂解(例如,以化学方式)和/或消化(例如,以酶方式,例如使用肽酶,例如胰蛋白酶)所关注多肽,将多肽片段化。片段化可在标记之前或之后进行。在一些实施方案中,在标记整个蛋白质之后进行片段化。可在裂解之前或之后标记一个或多个氨基酸以产生带标记的多肽。在一些实施方案中,在化学或酶片段化之后,选择多肽的大小。在一些实施方案中,移除较小多肽(例如,<2kDa)且保留较大多肽以用于序列分析。大小选择可使用诸如凝胶过滤、SEC、透析、PAGE凝胶提取、微流张力流或任何其他适合的技术实现。在步骤(2)中,多肽片段1002的N末端或C末端被修饰以产生末端修饰的多肽1004。在一些实施方案中,修饰包括添加固定部分。在一些实施方案中,修饰包括添加偶联部分。
因此,本发明提供了用使得能够固定至表面(例如,用于蛋白质分析的芯片上的样品孔的表面)的部分来修饰蛋白质及多肽的末端的方法。在一些实施方案中,此类方法包括修饰带标记的多肽的末端,所述带标记的多肽将要根据本申请被分析。在其他实施方案中,此类方法包括修饰蛋白质或酶的末端,所述蛋白质或酶降解根据本申请的蛋白质或多肽底物,或使根据本申请的蛋白质或多肽底物移位。
在一些实施方案中,蛋白质或多肽的羧基末端在包括以下的方法中被修饰:(i)阻断蛋白质或多肽的游离羧酸基;(ii)使蛋白质或多肽变性(例如,通过加热和/或化学手段);(iii)阻断蛋白质或多肽的游离硫醇基;(iv)消化蛋白质或多肽以产生至少一个多肽片段,其包括游离C末端羧酸基;及(v)将官能性部分缀合(例如,以化学方式)至游离C末端羧酸基。在一些实施方案中,该方法进一步包括在(i)之后且(ii)之前,透析包括蛋白质或多肽的样品。
在一些实施方案中,蛋白质或多肽的羧基末端在包括以下的方法中被修饰:(i)使蛋白质或多肽变性(例如,通过加热和/或化学手段);(ii)阻断蛋白质或多肽的游离硫醇基;(iii)消化蛋白质或多肽以产生至少一个多肽片段,其包括游离C末端羧酸基;(iv)阻断游离C末端羧酸基以产生至少一个多肽片段,其包括被阻断的C末端羧酸基;及(v)将官能性部分缀合(例如,以酶方式)至被阻断的C末端羧酸基。在一些实施方案中,该方法进一步包括在(iv)之后且(v)之前,透析包括蛋白质或多肽的样品。
在一些实施方案中,阻断游离羧酸基是指对这些基团进行化学修饰,相对于未被修饰的羧酸,其改变化学反应性。适合的羧酸阻断方法是本领域中已知的,且应修饰侧链羧酸基,以在化学上不同于待官能化的多肽的羧基末端羧酸基。在一些实施方案中,阻断游离羧酸基包括将多肽的游离羧酸基酯化或酰氨化。在一些实施方案中,阻断游离羧酸基包括将多肽的游离羧酸基甲酯化,例如通过使多肽与甲醇HCl反应进行。适用于阻断游离羧酸基的试剂及技术的额外实施例包括(但不限于)4-磺基-2,3,5,6-四氟苯酚(STP)和/或碳化二亚胺(诸如N-(3-二甲氨基丙基)-N'-乙基碳化二亚胺盐酸盐(EDAC))、脲鎓试剂、重氮甲烷、用于费歇尔酯化(Fischer esterification)的醇及酸、使用N-羟基丁二酰亚胺(NHS)形成NHS酯(潜在地作为后续酯或氨形成的中间体)、或与羰基二咪唑(CDI)反应或形成混合酸酐,或潜在地经由形成酯或酰氨进行的修饰或阻断羧酸的任何其他方法。
在一些实施方案中,阻断游离硫醇基是指对这些基团进行化学修饰,相对于未被修饰的硫醇,其改变化学反应性。在一些实施方案中,阻断游离硫醇基包括将蛋白质或多肽的游离硫醇基还原及烷化。在一些实施方案中,还原及烷化通过使多肽与二硫苏糖醇(DTT)及碘乙酰氨与碘乙酸中的一个或两个接触而进行。可使用的额外及替代半胱氨酸还原性试剂的实施例为人所熟知,且包括(但不限于)2-巯基乙醇、三(2-羧乙基)膦盐酸盐(TCEP)、三丁基膦、二硫丁氨(DTBA)或能够还原硫醇基的任何试剂。可使用的额外及替代半胱氨酸阻断性(例如,半胱氨酸烷化)试剂的实施例为人所熟知,且包括(但不限于)丙烯酰氨、4-乙烯基吡啶、N-乙基顺丁烯二酰亚胺(NEM)、N-ε-顺丁烯二酰亚胺基己酸(EMCA)或修饰半胱氨酸从而阻止二硫键形成的任何试剂。
在一些实施方案中,消化包括酶消化。在一些实施方案中,消化通过在消化条件下使蛋白质或多肽与内肽酶(例如,胰蛋白酶)接触进行。在一些实施方案中,消化包括化学消化。适用于化学及酶消化的试剂的实施例是本领域中已知的,且包括(但不限于)胰蛋白酶、胰凝乳蛋白酶、Lys-C、Arg-C、Asp-N、Lys-N、BNPS-粪臭素(Skatole)、CNBr、凋亡蛋白酶、甲酸、谷氨酰基内肽酶、羟氨、亚碘酰基苯甲酸(iodosobenzoic acid)、嗜中性白血球弹性蛋白酶、胃蛋白酶、脯氨酸内肽酶、蛋白酶K、葡萄球菌肽酶I、嗜热菌蛋白酶及凝血酶。
在一些实施方案中,官能性部分包括生物素分子。在一些实施方案中,官能性部分包括反应性化学部分,诸如炔基。在一些实施方案中,缀合官能性部分包括由羧肽酶Y对羧基末端羧基甲酯基进行生物素标记,如本领域中已知。
在一些实施方案中,将增溶部分添加至多肽中。图10B示出了例如使用将增溶连接子缀合至多肽的过程,将增溶部分添加至多肽的末端氨基酸中的非限制性实例。
在一些实施方案中,将末端修饰的多肽1010(其包括连接子缀合部分1012)缀合至增溶连接子1020(其包括多肽缀合部分1022)。在一些实施方案中,增溶连接子包括增溶聚合物,诸如生物分子(例如,展示为带点形状)。在一些实施方案中,包括形成于1012与1022之间的连接1032的所得连接子缀合多肽1030进一步包括表面缀合部分1034。因此,在一些实施方案中,本文所提供的方法及组合物适用于用提高其溶解度的部分修饰多肽的末端。在一些实施方案中,增溶部分适用于由片段化(例如,酶片段化,例如使用胰蛋白酶进行)产生且溶解度相对低的小多肽。举例而言,在一些实施方案中,可通过将聚合物(例如,短寡核苷酸、糖或其他带电聚合物)缀合至多肽,使多肽库中的短多肽溶解。
在一些实施方案中,样品孔的一个或多个表面(例如,样品孔的侧壁)可被修饰。对样品孔侧壁进行钝化和/或防积垢的非限制性实施例展示于图10C中,其中示出了样品孔的示例性示意图,其具有可用于促进单分子固定至底表面的被修饰的表面。在一些实施方案中,1040为SiO
发光标记
如本文所用,发光标记为吸收一个或多个光子且可随后在一个或多个持续时间之后发射一个或多个光子的分子。在一些实施方案中,视情形而定,该术语可与“标记”或“发光分子”互换使用。根据本文所描述的某些实施方案的发光标记可以指带标记的亲和试剂的发光标记、带标记的肽酶(例如,带标记的外肽酶、带标记的非特异性外肽酶)的发光标记、带标记的肽的发光标记、带标记的辅因子的发光标记或本文所描述的另一带标记的组合物。在一些实施方案中,根据本申请的发光标记是指包括一种或多种带标记的氨基酸的带标记的多肽中的带标记的氨基酸。
在一些实施方案中,发光标记可包括第一及第二发色团。在一些实施方案中,第一发色团的激发态能够经由能量传递至第二发色团弛豫。在一些实施方案中,能量传递为福斯特共振能量转移(FRET)。此类FRET对可能适用于为发光标记提供特性,这些特性使得该标记更易于在混合物中在多种发光标记当中进行区分,例如如本文针对图1C的带标记的适体106所示出且描述的那样。在其他实施方案中,FRET对包括第一发光标记的第一发色团及第二发光标记的第二发色团,例如如本文针对使用带标记的辅因子对带标记的肽进行测序所示出且描述的那样(参见例如图9)。在某些实施方案中,FRET对可吸收第一光谱范围内的激发能量且发射第二光谱范围内的发光。
在一些实施方案中,发光标记是指荧光团或染料。通常,发光标记包括芳族或杂芳族化合物且可为芘、蒽、萘、萘氨、吖啶、茋、吲哚、苯并吲哚、噁唑、咔唑、噻唑、苯并噻唑、苯并噁唑、菲啶、吩噁嗪、卟啉、喹啉、乙锭、苯甲酰氨、花青、羰花青、水杨酸盐、邻氨基苯甲酸酯、香豆素、荧光素(fluoroscein)、若丹明(rhodamine)、二苯并哌喃或其他类似化合物。
在一些实施方案中,发光标记包括选自以下中的一种或多种的染料:5/6-羧基若丹明(Carboxyrhodamine)6G、5-羧基若丹明6G、6-羧基若丹明6G、6-TAMRA、
发光
在一些方面中,本申请涉及基于发光标记的一种或多种发光特性的多肽测序和/或鉴定。在一些实施方案中,基于发光寿命、发光强度、亮度、吸收光谱、发射光谱、发光量子产率或其两者或多于两者的组合来鉴定发光标记。在一些实施方案中,可基于不同发光寿命、发光强度、亮度、吸收光谱、发射光谱、发光量子产率或其两者或多于两者的组合来彼此区别多种类型的发光标记。鉴定可能意味着指派与发光标记缔合的一种类型的氨基酸(例如,单一类型或一子集类型)的准确身分和/或数量,且也可能意味着相对于其他类型的氨基酸,指派多肽中的氨基酸位置。
在一些实施方案中,通过使发光标记曝露于独立光脉冲系列并评估从标记发射的各光子的时序或其他特性来检测发光。在一些实施方案中,将从标记依次发射的多个光子的信息汇总并进行评估,以鉴定标记并从而鉴定相关联类型的氨基酸。在一些实施方案中,根据从标记依次发射的多个光子来确定标记的发光寿命,并使用发光寿命来鉴定标记。在一些实施方案中,根据从标记依次发射的多个光子来确定标记的发光强度,并使用发光强度来鉴定标记。在一些实施方案中,根据从标记依次发射的多个光子来确定标记的发光寿命及发光强度,并使用发光寿命及发光强度来鉴定标记。
在本申请的一些方面中,使单一多肽分子曝露于多个独立光脉冲并检测并分析发射光子系列。在一些实施方案中,发射光子系列提供关于单一多肽分子的信息,该单一多肽分子在实验时间内在反应样品中存在且未改变。然而,在一些实施方案中,发射光子系列提供关于一系列不同分子的信息,这些分子在不同时间存在于反应样品中(例如,随着反应或过程进展)。借助于实施例而非限制,此类信息可用于对经历根据本申请的化学或酶降解的多肽进行测序和/或鉴定。
在某些实施方案中,发光标记吸收一个光子且在持续时间之后发射一个光子。在一些实施方案中,可通过测量持续时间确定或估计标记的发光寿命。在一些实施方案中,可通过测量多个脉冲事件及发射事件的多个持续时间来确定或估计标记的发光寿命。在一些实施方案中,可通过测量持续时间,在多种类型的标记的发光寿命中区分标记的发光寿命。在一些实施方案中,可通过测量多个脉冲事件及发射事件的多个持续时间,在多种类型的标记的发光寿命中区分标记的发光寿命。在某些实施方案中,通过确定或估计标记的发光寿命,在多种类型的标记中鉴定或区分标记。在某些实施方案中,通过在多种类型的标记的多个发光寿命中区分标记的发光寿命,在多种类型的标记中鉴定或区分标记。
确定发光标记的发光寿命可使用任何适合的方法(例如,通过使用适合的技术测量寿命,或通过确定发射的时间相依特征)进行。在一些实施方案中,确定一种标记的发光寿命包括相对于另一标记确定寿命。在一些实施方案中,确定标记的发光寿命包括相对于参考确定寿命。在一些实施方案中,确定标记的发光寿命包括测量寿命(例如,荧光寿命)。在一些实施方案中,确定标记的发光寿命包括确定指示寿命的一种或多种时间特征。在一些实施方案中,可基于相对于激发脉冲,贯穿一个或多个时间闸控窗发生的多个发射事件(例如,1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、30、40、50、60、70、80、90、100个或更多个发射事件)的分布,确定标记的发光寿命。举例而言,可基于相对于激发脉冲测量的光子出现次数的分布,区别标记的发光寿命与具有不同发光寿命的多种标记。
应了解,发光标记的发光寿命指示在标记达到激发态之后发射的光子的时序,且标记可由指示光子时序的信息区别。一些实施方案可包括通过测量与由标记发射的光子相关的时间,基于标记的发光寿命,来区别标记与多种标记。时间分布可提供发光寿命的指示,发光寿命可由分布来确定。在一些实施方案中,可基于时间分布,诸如通过将时间分布与对应于已知标记的参考分布进行比较,来区分标记与多种标记。在一些实施方案中,发光寿命的值由时间分布来确定。
如本文所用,在一些实施方案中,发光强度是指每单位时间发射光子的数目,这些光子由受脉冲激发能量的递送激发的发光标记发射。在一些实施方案中,发光强度是指所检测到的每单位时间发射光子的数目,这些光子由受脉冲激发能量的递送激发的发光标记发射,且由特定传感器或传感器组检测。
如本文所用,在一些实施方案中,亮度是指报告每发光标记平均发射强度的参数。因此,在一些实施方案中,“发射强度”可用于大体上指代包括一种或多种标记的组合物的亮度。在一些实施方案中,标记的亮度等于其量子产率与消光是数的乘积。
如本文所用,在一些实施方案中,发光量子产率是指引起发射事件,在给定波长下或给定光谱范围内的激发事件的分率,且通常小于1。在一些实施方案中,本文所描述的发光标记的发光量子产率在0与约0.001之间、在约0.001与约0.01之间、在约0.01与约0.1之间、在约0.1与约0.5之间、在约0.5与0.9之间或在约0.9与1之间。在一些实施方案中,通过确定或估计发光量子产率鉴定标记。
如本文所用,在一些实施方案中,激发能量为来自光源的光脉冲。在一些实施方案中,激发能量在可见光谱中。在一些实施方案中,激发能量在紫外线光谱中。在一些实施方案中,激发能量在红外线光谱中。在一些实施方案中,激发能量处于或接近于发光标记的最大吸收,将要从该发光标记检测多个发射光子。在某些实施方案中,激发能量在约500nm与约700nm之间(例如,在约500nm与约600nm之间、在约600nm与约700nm之间、在约500nm与约550nm之间、在约550nm与约600nm之间、在约600nm与约650nm之间或在约650nm与约700nm之间)。在某些实施方案中,激发能量可为单色的或受限于光谱范围。在一些实施方案中,光谱范围具有在约0.1nm与约1nm之间、在约1nm与约2nm之间或在约2nm与约5nm之间的范围。在一些实施方案中,光谱范围具有在约5nm与约10nm之间、在约10nm与约50nm之间或在约50nm与约100nm之间的范围。
测序
本申请的方面涉及对生物聚合物,诸如多肽及蛋白质进行测序。如本文所用,参考多肽或蛋白质,“测序”、“序列确定”、“确测序列”及类似术语包括确定多肽或蛋白质的部分序列信息以及全序列信息。亦即,术语包括序列比较、指纹识别(fingerprinting)、机率性指纹识别及关于目标分子的类似水平的信息,以及感兴趣区域内目标分子的各氨基酸的明确鉴定及排序。在一些实施方案中,术语包括鉴定多肽的单一氨基酸。在其他实施方案中,鉴定多肽的多于一个氨基酸。如本文所用,在一些实施方案中,参考氨基酸,“鉴定”、“确定身分”及类似术语包括确定氨基酸的明确身分,以及确定氨基酸的明确身分的机率。举例而言,在一些实施方案中,通过确定氨基酸具有特定类型的机率(例如,0%至100%),或通过确定多种特定类型中的每一种的机率,来鉴定氨基酸。因此,在一些实施方案中,如本文所用,术语“氨基酸序列”、“多肽序列”及“蛋白质序列”可以指多肽或蛋白质材料自身,且不限于在生物化学上表征特定多肽或蛋白质的特定序列信息(例如,表示氨基酸从一个末端向另一个末端的顺序的一连串字母)。
在一些实施方案中,对多肽分子进行测序包括鉴定多肽分子中的至少两个(例如,至少3个、至少4个、至少5个、至少6个、至少7个、至少8个、至少9个、至少10个、至少11个、至少12个、至少13个、至少14个、至少15个、至少16个、至少17个、至少18个、至少19个、至少20个、至少25个、至少30个、至少35个、至少40个、至少45个、至少50个、至少60个、至少70个、至少80个、至少90个、至少100个或更多个)氨基酸。在一些实施方案中,该至少两个氨基酸为相邻氨基酸。在一些实施方案中,该至少两个氨基酸为非相邻氨基酸。
在一些实施方案中,对多肽分子进行测序包括鉴定多肽分子中所有氨基酸的少于100%(例如,少于99%、少于95%、少于90%、少于85%、少于80%、少于75%、少于70%、少于65%、少于60%、少于55%、少于50%、少于45%、少于40%、少于35%、少于30%、少于25%、少于20%、少于15%、少于10%、少于5%、少于1%或更少)。举例而言,在一些实施方案中,对多肽分子进行测序包括鉴定多肽分子中一种类型的氨基酸的少于100%(例如,鉴定多肽分子中一种类型的所有氨基酸的一部分)。在一些实施方案中,对多肽分子进行测序包括鉴定多肽分子中各类型的氨基酸的少于100%。
在一些实施方案中,对多肽分子进行测序包括鉴定多肽中至少1种、至少5种、至少10种、至少15种、至少20种、至少25种、至少30种、至少35种、至少40种、至少45种、至少50种、至少55种、至少60种、至少65种、至少70种、至少75种、至少80种、至少85种、至少90种、至少95种、至少100种或更多种类型的氨基酸。
在一些实施方案中,本申请提供了通过随时间推移鉴定在多肽的末端存在的氨基酸系列(例如,通过迭代检测及裂解末端的氨基酸),从而对多肽进行测序的组合物及方法。在其他实施方案中,本申请提供了通过鉴定多肽的带标记的氨基内容并与参考序列数据库进行比较,从而对多肽进行测序的组合物及方法。
在一些实施方案中,本申请提供了通过对多肽的多个片段进行测序,从而对多肽进行测序的组合物及方法。在一些实施方案中,对多肽进行测序包括将多个多肽片段的序列信息组合,以鉴定和/或确定多肽的序列。在一些实施方案中,组合序列信息可由电脑硬件及软件进行。本文所描述的方法可允许一组相关多肽,诸如生物体的全部蛋白质组被测序。在一些实施方案中,根据本申请的方面,并行(例如,在单一芯片上)进行多个单分子测序反应。举例而言,在一些实施方案中,多个单分子测序反应在单一芯片上的单独样品孔中各自进行。
在一些实施方案中,本文所提供的方法可用于对包括蛋白质的复杂混合物的样品中的单独蛋白质进行测序及鉴定。在一些实施方案中,本申请提供了唯一地鉴定蛋白质的复杂混合物中的单独蛋白质的方法。在一些实施方案中,通过确定蛋白质的部分氨基酸序列,在混合样品中检测单独蛋白质。在一些实施方案中,蛋白质的部分氨基酸序列在大约5至50个氨基酸的连续延伸段内。
不希望受任何特定理论束缚,相信大部分人类蛋白质可使用不完全序列信息,参考蛋白质组数据库而鉴定。举例而言,人类蛋白质组的简单建模已表明,大约98%的蛋白质可通过在6至40个氨基酸的延伸段内检测仅四种类型的氨基酸而唯一地鉴定(参见例如Swaminathan等人PLoS Comput Biol.2015,11(2):e1004080;及Yao等人Phys.Biol.2015,12(5):055003)。因此,可将蛋白质的复杂混合物降解(例如,以化学方式降解、以酶方式降解)成大约6至40个氨基酸的短多肽片段,且对此多肽库进行测序将展现初始复杂混合物中存在的蛋白质中的每一个的身分及丰度。用于选择性氨基酸标记及通过确定部分序列信息鉴定多肽的组合物及方法详细描述于2015年9月15日申请的标题为“单分子肽测序(SINGLEMOLECULE PEPTIDE SEQUENCING)”的美国专利申请第15/510,962号中,该申请以全文引用的方式并入。
实施方案能够以高准确度对单一多肽分子进行测序,该准确度诸如至少约50%、60%、70%、75%、80%、85%、90%、95%、96%、97%、98%、99%、99.9%、99.99%、99.999%或99.9999%的准确度。在一些实施方案中,单分子测序中所使用的目标分子为固定至固体载体的表面的多肽,该表面诸如样品孔的底表面或侧壁表面。样品孔也可含有根据本申请的测序反应所需的任何其他试剂,诸如一种或多种适合的缓冲液、辅因子、带标记的亲和试剂及酶(例如,催化活性或无催化活性外肽酶,其可被发光标记的或不标记)。
如上文所描述,在一些实施方案中,根据本申请的测序包括通过确定氨基酸具有特定类型的机率来鉴定氨基酸。常规蛋白质鉴定系统需要鉴定多肽中的各氨基酸以鉴定多肽。然而,难以准确鉴定多肽中的各氨基酸。举例而言,从第一识别分子与第一氨基酸缔合的相互作用收集的数据与从第二识别分子与第二氨基酸缔合的相互作用收集的数据的差异可能不足以区分两种氨基酸。在一些实施方案中,根据本申请的测序通过使用蛋白质鉴定系统而避免此问题,该蛋白质鉴定系统不同于常规蛋白质鉴定系统,不需要(但不排除)鉴定蛋白质中的各氨基酸。
因此,在一些实施方案中,根据本申请的测序可使用蛋白质鉴定系统进行,该系统使用机器学习技术来鉴定蛋白质。在一些实施方案中,系统通过以下操作:(1)使用实时蛋白质测序装置收集关于蛋白质的多肽的数据;(2)使用机器学习模型及所收集的数据来鉴定各自的位置处某些氨基酸为多肽的一部分的机率;及(3)使用所鉴定的机率作为“机率性指纹”来鉴定蛋白质。在一些实施方案中,可使用选择性结合氨基酸的试剂获得关于蛋白质的多肽的数据。作为一实施例,试剂和/或氨基酸可用发光标记来标记,该发光标记回应于激发能量的施加而发射光。在此实施例中,蛋白质测序装置可在试剂与样品中的氨基酸的结合相互作用期间,将激发能量施加至蛋白质(例如,多肽)的样品。在一些实施方案中,测序装置中的一个或多个传感器(例如,光检测器、电传感器和/或任何其他适合类型的传感器)可检测结合相互作用。继而,可将从所检测到的光发射收集和/或导出的数据提供至机器学习模型。机器学习模型及相关系统及方法详细描述于2019年6月12日申请的标题为“启用机器学习的蛋白质鉴定(MACHINE LEARNING ENABLED PROTEIN IDENTIFICATION)”的美国临时专利申请第62/860,750号,该申请以全文引用的方式并入。
根据本申请的测序在一些方面中可涉及将多肽固定于基板(例如,固体载体,例如芯片,例如如本文所描述的集成装置)的表面上。在一些实施方案中,可将多肽固定于基板上的样品孔的表面上(例如,样品孔的底表面上)。在一些实施方案中,多肽的N末端氨基酸被固定(例如,连接至表面)。在一些实施方案中,多肽的C末端氨基酸被固定(例如,连接至表面)。在一些实施方案中,一种或多种非末端氨基酸被固定(例如,连接至表面)。(多种)固定氨基酸可使用例如如本申请中所描述的任何适合的共价或非共价连接连接。在一些实施方案中,将多种多肽连接至例如基板上的样品孔阵列中的多个样品孔(例如,其中一种多肽连接至各样品孔的表面,例如底表面)。
根据本申请的测序在一些方面中可使用准许单分子分析的系统进行。系统可包括集成装置及被配置为与集成装置介接的仪器。集成装置可包括像素阵列,其中单独像素包括样品孔及至少一个光检测器。集成装置的样品孔可在集成装置的表面上或通过集成装置的表面形成,并被配置为接纳放置于集成装置的表面上的样品。集合地,样品孔可视为样品孔阵列。多个样品孔可具有适合的大小及形状,使得样品孔的至少一部分接纳单一样品(例如,单分子,诸如多肽)。在一些实施方案中,样品孔内样品的数目可分布于集成装置的样品孔当中,使得一些样品孔含有一种样品,而其他样品孔含有零、两种或更多种样品。
从集成装置外部的一种或多种光源向集成装置提供激发光。集成装置的光学组件可接收来自光源的激发光,并将光朝集成装置的样品孔阵列导向,并照射样品孔内的照射区域。在一些实施方案中,样品孔可具有一定构造,该构造允许样品被保留在样品孔表面附近,可使向样品递送激发光以及检测样品发射光更容易。定位于照射区域内的样品可回应于由激发光照射而发射发射光。举例而言,样品可被荧光标记物标记,该荧光标记物回应于经由激发光照射达成的激发态而发射光。随后可由对应于具有所分析样品的样品孔的像素内的一个或多个光检测器检测由样品发射的发射光。当遍及样品孔阵列(根据一些实施方案,其范围可在大约10,000个像素至1,000,000个像素之间的数目内)进行时,多个样品可并行分析。
集成装置可包括用于接收激发光并将激发光在样品孔阵列当中导向的光学系统。光学系统可包括一个或多个光栅耦合器,其被配置为将激发光耦合至集成装置,并将激发光导向其他光学组件。光学系统可包括将激发光自光栅耦合器朝样品孔阵列导向的光学组件。此类光学组件可包括光分路器(optical splitter)、光合路器(optical combiner)及波导。在一些实施方案中,一个或多个光分路器可耦合来自光栅耦合器的光,并将激发光递送至波导中的至少一个。根据一些实施方案,光分路器可具有一种构造,该构造允许在所有波导中基本上均一地递送激发光,使得波导中的每一个接收基本上相似量的激发光。此类实施方案可通过提高由集成装置的样品孔接收的激发光的均一性,来提高集成装置的效能。集成装置中包括的适用于例如将激发光耦合至样品孔和/或将发射光导向至光检测器的组件的实施例描述于2015年8月7日申请的标题为“用于探测、检测及分析分子的集成装置(INTEGRATED DEVICE FOR PROBING,DETECTING AND ANALYZING MOLECULES)”的美国专利申请第14/821,688号,及2014年11月17日申请的标题为“用于探测、检测及分析分子的具有外部光源的集成装置(INTEGRATED DEVICE WITH EXTERNAL LIGHT SOURCE FORPROBING,DETECTING,AND ANALYZING MOLECULES)”的美国专利申请第号14/543,865中,这些申请均以全文引用的方式并入。可实施于集成装置中的适合光栅耦合器及波导的实施例描述于2017年12月15日申请的标题为“光学耦合器及波导系统(OPTICAL COUPLER ANDWAVEGUIDE SYSTEM)”的美国专利申请第15/844,403号中,该申请以全文引用的方式并入。
额外光子结构可定位于样品孔与光检测器之间,并被配置为减少或阻止激发光到达光检测器,否则其可在检测发射光时促成信号噪声。在一些实施方案中,可充当集成装置的电路系统的金属层也可充当空间滤光器。适合的光子结构的实施例可包括光谱滤光器、偏振滤光器及空间滤光器,且描述于2018年7月23日申请的标题为“光学排斥光子结构(OPTICAL REJECTION PHOTONIC STRUCTURES)”的美国专利申请第16/042,968号中,该申请以全文引用的方式并入。
位于集成装置外的组件可用于将激发源向集成装置定位并对准。此类组件可包括光学组件,其包括透镜、反射镜、棱镜、窗、孔、衰减器和/或光纤。可在仪器中包括额外机械组件以允许控制一种或多种对准组件。此类机械组件可包括致动器、步进马达和/或把手。适合的激发源及对准机构的实施例描述于2016年5月20日申请的标题为“脉冲雷射及系统(PULSED LASER AND SYSTEM)”的美国专利申请第号15/161,088中,该申请以全文引用的方式并入。光束引导模块的另一实施例描述于2017年12月14日申请的标题为“紧凑光束整形及引导组合件(COMPACT BEAM SHAPING AND STEERING ASSEMBLY)”的美国专利申请第15/842,720号中,该申请以引用的方式并入本文中。适合的激发源的额外实施例描述于2015年8月7日申请的标题为“用于探测、检测及分析分子的集成装置(INTEGRATED DEVICE FORPROBING,DETECTING AND ANALYZING MOLECULES)”的美国专利申请第14/821,688号中,该申请以全文引用的方式并入。
伴随集成装置的单独像素定位的(多个)光检测器可被配置并定位,以检测来自像素的对应样品孔的发射光。适合的光检测器的实施例描述于2015年8月7日申请的标题为“用于所接收的光子的时间分组的集成装置(INTEGRATED DEVICE FOR TEMPORAL BINNINGOF RECEIVED PHOTONS)”的美国专利申请第14/821,656号中,该申请以全文引用的方式并入。在一些实施方案中,样品孔及其各自的(多个)光检测器可沿公共轴对准。以此方式,(多个)光检测器可与像素内的样品孔重叠。
所检测到的发射光的特征可为鉴定与发射光相关的标记物提供指示。此类特征可包括任何适合类型的特征,包括由光检测器检测的光子到达时间、光检测器随时间推移所累积的光子的量和/或遍及两个或更多个光检测器的光子分布。在一些实施方案中,光检测器可具有一种构造,该构造允许检测与样品的发射光相关的一种或多种时序特征(例如,发光寿命)。光检测器可检测激发光脉冲通过集成装置传播之后光子到达时间的分布,且到达时间的分布可提供样品的发射光的时序特征的指示(例如,发光寿命的代理)。在一些实施方案中,一个或多个光检测器提供由标记物发射的发射光的机率(例如,发光强度)的指示。在一些实施方案中,多个光检测器可被设定大小并布置,以捕获发射光的空间分布。随后可使用来自一个或多个光检测器的输出信号,从多种标记物当中区别一种标记物,其中该多种标记物可用于鉴定样品内的样品。在一些实施方案中,样品可由多个激发能量激发,且由样品回应于多个激发能量而发射的发射光的发射光和/或时序特征可区别标记物与多种标记物。
在操作时,通过使用激发光激发孔内样品中的一些或全部,且用光检测器检测来自样品发射的信号进行样品孔内的样品的并行分析。来自样品的发射光可由对应光检测器检测且转换为至少一个电信号。电信号可沿集成装置的电路系统中的导线传输,导线可连接至与集成装置介接的仪器。电信号随后可经处理和/或分析。处理或分析电信号可在位于仪器上或仪器外的适合计算装置上进行。
仪器可包括用于控制仪器和/或集成装置的操作的使用者接口。使用者接口可被配置以允许使用者将信息输入至仪器中,诸如用于控制仪器的功能的命令和/或设置。在一些实施方案中,使用者接口可包括按钮、开关、拨号盘及用于话音命令的麦克风。使用者接口可允许使用者接收对仪器和/或集成装置的效能的反馈,诸如正确对准和/或由来自集成装置上的光检测器的读出信号获得的信息。在一些实施方案中,使用者接口可使用扬声器以提供可听反馈来提供反馈。在一些实施方案中,使用者接口可包括用于向使用者提供视觉反馈的指示灯和/或显示荧幕。
在一些实施方案中,仪器可包括被配置以与计算装置连接的电脑接口。电脑接口可为USB接口、FireWire接口或任何其他适合的电脑接口。计算装置可为任何通用电脑,诸如膝上型电脑或桌上型电脑。在一些实施方案中,计算装置可为服务器(例如,基于云端的服务器),其可经由适合的电脑接口经一无线网络接入。电脑接口可使仪器与计算装置之间的信息通信便利。可将用于控制和/或构造仪器的输入信息提供至计算装置,且经电脑接口传输至仪器。计算装置可经由电脑接口接收由仪器产生的输出信息。输出信息可包括关于仪器效能、集成装置效能的反馈和/或由光检测器的读出信号产生的数据。
在一些实施方案中,仪器可包括处理装置,其被配置以分析从集成装置的一个或多个光检测器接收到的数据和/或将控制信号传输至(多个)激发源。在一些实施方案中,处理装置可包括通用处理器、专门适配的处理器(例如,中央处理单元(CPU)(诸如一种或多种微处理器或微控制器核心)、现场可编程门阵列(FPGA)、特殊应用集成电路(ASIC)、定制集成电路、数位信号处理器(DSP)或其组合)。在一些实施方案中,可由仪器的处理装置及外部计算装置两者进行对来自一个或多个光检测器的数据的处理。在其他实施例中,外部计算装置可省略,且可由集成装置的处理装置单独进行对来自一个或多个光检测器的数据的处理。
根据一些实施方案,被配置以基于发光发射特征来分析样品的仪器可检测不同发光分子之间的发光寿命和/或强度的差异,和/或不同环境中相同发光分子的寿命和/或强度之间的差异。诸位发明人已认识到且了解,发光发射寿命的差异可用于在存在或不存在不同发光分子之间进行辨别和/或在发光分子经历的不同环境或条件之间进行辨别。在一些情况下,基于寿命(而非例如发射波长)辨别发光分子可简化系统的方面。作为一实施例,在基于寿命辨别发光分子时,可减少区分波长的光学器件(诸如波长滤光器、各波长的专用检测器、不同波长下的专用脉冲光学源和/或绕射光学器件)的数目或除去这些区分波长的光学装置。在一些情况下,在单一特征波长下操作的单一脉冲光学源可用于激发不同发光分子,这些发光分子在光谱的相同波长范围内发射但具有可测量的不同寿命。使用单一脉冲光学源,而非在不同波长下操作的多个源,以激发且辨别在相同波长区域内发射的不同发光分子的分析系统可较易于操作及维护、较紧凑且可以较低成本制造。
尽管基于发光寿命分析的分析系统可具有某些益处,但由分析系统获得的信息的量和/或检测准确度可通过允许额外检测技术而增加。举例而言,系统的一些实施方案可另外被配置,以基于发光波长和/或发光强度来辨别样品的一个或多个特性。在一些实施方案中,可另外或替代地使用发光强度区别不同发光标记。举例而言,尽管一些发光标记的衰变速率可能类似,但其可以显著不同强度发射或其激发机率具有显著差异(例如,至少约35%的差异)。通过将分组信号与所测量的激发光参比,或许有可能基于强度水平来区别不同发光标记。
根据一些实施方案,可用光检测器区别不同发光寿命,该光检测器被配置,以对发光标记激发之后的发光发射事件进行时间分组。时间分组可发生在光检测器的单一电荷积聚循环期间。电荷积聚循环为读出事件之间的间隔,在此期间,光生载流子积聚于时间分组光检测器的仓中。时间分组光检测器的实施例描述于2015年8月7日申请的标题为“用于对所接收的光子进行时间分组的集成装置(INTEGRATED DEVICE FOR TEMPORAL BINNING OFRECEIVED PHOTONS)”的美国专利申请第14/821,656号中,该申请以引用的方式并入本文中。在一些实施方案中,时间分组光检测器可在光子吸收/载流子产生区域中产生电荷载流子,且将电荷载流子直接转移至电荷载流子储存区域中的储仓中。在此类实施例中,时间分组光检测器可不包括载流子行进/捕获区域。此类时间分组光检测器可称作“直接分组像素”。时间分组光检测器(包括直接分组像素)的实施例描述于2017年12月22日申请的标题为“具有直接分组像素的集成光检测器(INTEGRATED PHOTODETECTOR WITH DIRECTBINNING PIXEL)”的美国专利申请第15/852,571号中,该申请以引用的方式并入本文中。
在一些实施方案中,不同数目个相同类型的荧光团可连接至样品中的不同试剂,使得可基于发光强度来鉴定各试剂。举例而言,两个荧光团可连接至第一带标记的亲和试剂,且四个或多于四个荧光团可连接至第二带标记的亲和试剂。因为荧光团的数目不同,所以可存在与不同亲和试剂相关的不同激发及荧光团发射机率。举例而言,在信号积聚间隔期间可存在第二带标记的亲和试剂的更多发射事件,使得分组的表观强度显著高于第一带标记的亲和试剂的表观强度。
诸位发明人已认识到且了解,基于荧光团衰变速率和/或荧光团强度区别核苷酸或任何其他生物或化学样品可使得光学激发及检测系统能够简化。举例而言,可用单一波长源(例如,产生一种特征波长的源而非多个源或在多个不同特征波长下操作的源)进行光学激发。另外,检测系统中可不需要区分波长的光学器件及滤光器。此外,各样品孔可使用单一光检测器以检测来自不同荧光团的发射。词组“特征波长”或“波长”用以指有限辐射频宽内的中心或主导波长(例如,由脉冲光学源输出的20nm频宽内的中心或峰值波长)。在一些情况下,“特征波长”或“波长”可用以指通过源输出的总辐射频宽内的峰值波长。
计算技术
本申请的一些方面涉及用于分析由本文所描述的多肽测序技术产生的数据的计算技术。如上文例如结合图1A及图1B所论述,使用这些测序技术产生的数据可包括信号脉冲系列,其指示氨基酸识别分子与所测序的多肽的末端暴露的氨基酸缔合的情况。随着降解过程继续移除连续氨基酸,随时间推移,视当前处于末端的氨基酸的类型而定,信号脉冲系列可具有变化的一种或多种特征(例如,脉冲持续时间、脉冲间持续时间、幅度的变化)。所得信号迹线可包括特征图案,其由与各自的氨基酸相关的变化的一种或多种特征引起。本文所描述的计算技术可作为使用这些测序技术分析所获得的此类数据以鉴定氨基酸序列的一部分来实施。
一些实施方案可涉及在多肽降解过程中获得数据;分析数据,以确定对应于以下的数据部分:在降解过程中在多肽的末端依次暴露的氨基酸;及输出代表多肽的氨基酸序列。图11为用于通过分析使用本文所描述的多肽测序技术获得的数据,来鉴定氨基酸序列的说明性处理流水线1100的图解。如图11中所展示,分析测序数据1102可涉及使用缔合事件鉴定技术1104及氨基酸鉴定技术1106来输出(多种)氨基酸序列1108。
如本文所论述,可在多肽降解过程中获得测序数据1102。在一些实施方案中,测序数据1102指示在降解过程中多肽的末端的氨基酸身分。在一些实施方案中,测序数据1102指示在降解过程中由一种或多种氨基酸识别分子与末端的不同类型的末端氨基酸结合所产生的信号。例示性测序数据展示于图1A及图1B中,其如上文所论述。
视信号在降解过程中如何产生而定,测序数据1102可指示一种或多种不同类型的信号。在一些实施方案中,测序数据1102指示在降解过程中产生的发光信号。举例而言,发光标记可用于标记氨基酸识别分子,且由发光标记发射的发光可随着氨基酸识别分子与特定氨基酸缔合,产生发光信号而被检测到。在一些实施方案中,测序数据1102指示在降解过程中产生的电信号。举例而言,所测序的多肽分子可固定至纳米孔,且电信号(例如,电导的变化)可随着氨基酸识别分子与特定氨基酸缔合而被检测到。
一些实施方案涉及分析测序数据1102,以确定对应于在降解过程中在多肽的末端依次暴露的氨基酸的测序数据1102的部分。如图11中所展示,缔合事件鉴定技术1104可存取测序数据1102,并分析测序数据以鉴定对应于缔合事件的测序数据1102的部分。缔合事件可对应于数据中的特征图案,诸如图1B中所展示的CP
一些实施方案涉及针对测序数据1102所确定的部分中的一种或多种来鉴定一种类型的氨基酸。如图11中所展示,氨基酸鉴定技术1106可用于针对通过缔合事件鉴定技术1104鉴定的缔合事件中的一种或多种,来确定一种类型的氨基酸。在一些实施方案中,通过缔合事件鉴定技术1104鉴定的数据的单独部分可包括脉冲图案,且氨基酸鉴定技术1106可针对这些部分中的一种或多种,基于其各自的脉冲图案来确定一种类型的氨基酸。参见图1B,氨基酸鉴定技术1106可鉴定CP
通过鉴定测序数据1102的连续部分的一种类型的氨基酸,氨基酸鉴定技术1106可输出代表多肽的(多种)氨基酸序列1108。在一些实施方案中,氨基酸序列包括对应于使用缔合事件鉴定技术1104鉴定的数据的部分的氨基酸系列。
图12为根据本文所描述的技术的一些实施方案,用于确定多肽分子的氨基酸序列的说明性过程1200的流程图。过程1200可在任何适合的(多种)计算装置(例如,单一计算装置、共置于单一实体位置中或位于远离彼此的多个实体位置中的多个计算装置、云端计算系统的一个或多个计算装置部分等)上进行,因为本文所描述的技术的方面在此方面中不受限制。在一些实施方案中,缔合事件鉴定技术1104及氨基酸鉴定技术1106可进行过程1200中的一些或全部以确定(多种)氨基酸序列。
过程1200以操作1202开始,其涉及使单一多肽分子与一种或多种末端氨基酸识别分子接触。接下来,过程1200继续进行至操作1104,其涉及检测信号脉冲系列,该系列指示一种或多种末端氨基酸识别分子与单一多肽降解时在单一多肽的末端暴露的连续氨基酸的缔合。脉冲系列可允许对单一多肽分子进行测序,诸如通过使用缔合事件鉴定技术1104及氨基酸鉴定技术1106。
在一些实施方案中,过程1200可包括操作1206,其涉及基于信号脉冲系列中的第一特征图案鉴定单一多肽分子中的第一类型的氨基酸,诸如通过使用氨基酸鉴定技术1106。
图13为根据本文所描述的技术的一些实施方案,用于确定代表多肽的氨基酸序列的说明性过程1300的流程图。过程1300可在任何适合的(多种)计算装置(例如,单一计算装置、共置于单一实体位置中或位于远离彼此的多个实体位置中的多个计算装置、云端计算系统的一个或多个计算装置部分等)上进行,因为本文所描述的技术的方面在此方面中不受限制。在一些实施方案中,缔合事件鉴定技术1104及氨基酸鉴定技术1106可进行过程1300中的一些或全部以确定(多种)氨基酸序列。
过程1300以操作1302开始,其中获得多肽降解过程中的数据。在一些实施方案中,数据指示在降解过程中多肽的末端的氨基酸身分。在一些实施方案中,数据指示在降解过程中由一种或多种氨基酸识别分子与末端的不同类型的末端氨基酸结合所产生的信号。在一些实施方案中,数据指示在降解过程中产生的发光信号。在一些实施方案中,数据指示在降解过程中产生的电信号。
接下来,过程1300继续进行至操作1304,其中分析数据,以确定对应于以下的数据部分:在降解过程中在多肽的末端依次暴露的氨基酸,诸如通过使用缔合事件鉴定技术1104及氨基酸鉴定技术1106。在一些实施方案中,分析数据进一步包括检测裂解事件系列,及确定连续裂解事件之间的数据的部分,诸如通过使用缔合事件鉴定技术1104。
在一些实施方案中,分析数据进一步包括针对单独部分中的每一个确定一种类型的氨基酸,诸如通过使用氨基酸鉴定技术1106。在一些实施方案中,单独部分中的每一个包括脉冲图案,且分析数据进一步包括针对这些部分中的一种或多种,基于其各自的脉冲图案确定一种类型的氨基酸。在一些实施方案中,确定氨基酸的类型进一步包括在数据超过阈值时,鉴定一部分内的时间量,及将该时间量与该部分的持续时间进行比较。在一些实施方案中,确定氨基酸的类型进一步包括针对一个或多个部分中的每一个,鉴定至少一个脉冲持续时间。在一些实施方案中,确定氨基酸的类型进一步包括针对一个或多个部分中的每一个,鉴定至少一个脉冲间持续时间。
接下来,过程1300继续进行至操作1306,其中输出代表多肽的氨基酸序列,诸如经由使用者接口。在一些实施方案中,氨基酸序列包括对应于这些部分的氨基酸系列。
图14中展示了可与本文所描述的技术的实施例中的任一种结合使用的电脑系统1400的说明性实施方案。电脑系统1400包括一种或多种处理器1410及一种或多种制品,其包括非暂时性电脑可读储存介质(例如,内存1420及一种或多种非易失性储存介质1430)。处理器1410可以任何适合方式控制向内存1420及非易失性储存装置1430写入数据及从其中读取数据,因为本文所描述的技术的方面在此方面中不受限制。为了执行本文所描述的功能中的任一种,处理器1410可执行储存于一个或多个非暂时性电脑可读储存介质(例如,内存1420)中的一种或多种处理器可执行指令,其可充当储存用于由处理器1410执行的处理器可执行指令的非暂时性电脑可读储存介质。
计算装置1400也可包括网络输入/输出(I/O)接口1440,计算装置可经由该接口与其他计算装置通信(例如,在网络上);且也可包括一个或多个使用者I/O接口1450,计算装置可经由该接口提供向使用者的输出及接收来自使用者的输入。使用者I/O接口可包括诸如键盘、鼠标、麦克风、显示装置(例如,监视器或触控式荧幕)、扬声器、相机的装置和/或各种其他类型的I/O装置。
上述实施方案可以众多方法中的任一种来实施。举例而言,这些实施方案可使用硬件、软件或其组合来实施。当实施于软件中时,软件代码可在任何适合处理器(例如,微处理器)或处理器集合(不管在单一计算装置中提供或分布于多个计算装置当中)上执行。应了解,执行上文所描述的功能的任何组件或组件的集合可大体上视为控制上述功能的一个或多个控制器。可以众多方式实施一种或多种控制器,诸如通过专用硬件,或通过使用微码或软件以执行上文所概述的功能的经程序化的通用硬件(例如,一种或多种处理器)。
在此方面中,应了解,本文所描述的实施例的一种实施方案包括编码有电脑程序(亦即,多个可执行指令)的至少一种电脑可读储存介质(例如,RAM、ROM、EEPROM、快闪存储器或其他内存技术、CD-ROM、数位多功能光盘(DVD)或其他光盘储存器、盒式磁带、磁带、磁盘储存器或其他磁储存装置,或其他有形非暂时性电脑可读储存介质),当在一种或多种处理器上执行时,该电脑程序执行一个或多个实施例的上述功能。电脑可读介质可为可输送的,使得储存于其上的程序可加载于任何计算装置上以实施本文中所论述的技术的方面。另外,应了解,对在执行时执行上述功能中的任一种的电脑程序的参考不限于在主电脑上执行的应用程序。实际上,术语电脑程序及软件在本文中以通用含义使用,以指代可用于程序化一种或多种处理器以实施本文所论述的技术的方面的任何类型的电脑代码(例如,应用软件、韧体、微码或任何其他形式的电脑指令)。
实施例
实施例1.通过化学裂解进行的艾德曼降解
提供大约3个至大约30个氨基酸(n=3-30)的经表面连接的寡肽,其中氨基酸残基R
实施例2.用于肽表面固定的增溶连接子
试图提高寡肽在水性缓冲液中的溶解度,确定了可将肽片段与寡核苷酸连接子缀合,以同时提高水溶解度,且提供官能性部分,用于在单分子水平上进行肽的表面固定。合成了不同肽-连接子缀合物,其具有图15A中针对肽-DNA缀合物及肽-PEG缀合物描绘的示例性结构。对于所评估的不同肽-连接子缀合物中的每一个,观测到连接子缀合极大地提高了肽在水溶液中的溶解度。
通过N末端氨肽酶评估肽-连接子缀合物在肽N-末端的氨基酸裂解(表6,以下)。
表6.肽-连接子缀合物的末端氨基酸裂解。
将表6中所展示的肽-连接子缀合物与脯氨酸亚胺基肽酶(“PIP”)或大鼠氨肽酶N(“大鼠APN”)一起孵育,并通过LCMS监测肽裂解。图15B中展示了LCMS的实施例,证明了来自表6的条目5的裂解。所有其他裂解反应以类似方式测量。如表6中所展示,尽管带正电的肽-DNA缀合物(“寡核苷酸”及“寡核苷酸-PEG连接子))未被所测试的氨肽酶裂解,但具有DNA寡核苷酸连接子的所有其他缀合物类别(带负电、芳族、疏水性)均被裂解。相比之下,表明带正电的肽-PEG缀合物被氨基肽酶中的至少一种裂解。
使用带标记的肽-连接子缀合物,表明具有不同组成的肽可固定至单独样品孔表面,以便进行单分子分析。对于这些实验,用染料(例如,如图15A中针对肽-DNA缀合物所描绘的)标记DNA连接子,且通过染料荧光测量不同肽-DNA缀合物向单独样品孔中的负载。图15C中展示了示例性负载实验。通过测量带标记的肽-DNA缀合物(50pM)的荧光发射,确定芯片上至少18%的样品孔以每样品孔单一占有率负载有经表面固定的缀合物。这些实验证明,与非缀合肽对应物相比,肽-连接子缀合物显示提高的水溶解度;缀合连接子不阻止不同氨肽酶对肽的末端氨基酸裂解;及具有不同组成的肽-连接子缀合物可在单分子水平上固定至芯片表面。
实施例3.多肽底物的外肽酶裂解
测试了各种氨肽酶的裂解能力。表7中展示了一组裂解分析实验的条件及结果,包括肽底物浓度、酶浓度、缓冲液条件、温度及孵育时间。使用高效液相色谱(HPLC)分析指定酶对肽底物的裂解。表7中的“HPLC分析转化率”值指示转化为裂解产物的肽底物的百分比。为了确定“HPLC分析转化率”值,制备了含有相同起始肽浓度的两种溶液。一种溶液经历酶消化,而另一溶液不含任何酶,但用当量的用于储存酶的缓冲液稀释。反应在指定时间淬灭。通过以下方法确定转化为产物的反应物的量:将在酶消化之后剩余的起始材料的HPLC获得的峰的面积除以未消化的肽的对照溶液的峰面积,且随后将此比率乘以100。在表7中,“NH2”指示氨基,“yPIP”是指鼠疫耶氏杆菌脯氨酸亚胺基肽酶,“NPEPPS”是指嘌呤霉素敏感型氨肽酶,“VPr”是指解蛋白弧菌氨肽酶且“EDAPN”是指嗜肺性退伍军人杆菌M1氨肽酶。
表7.氨肽酶对肽底物的裂解。
图16中展示了表7的所选外肽酶的氨基酸裂解活性的概述。展示了以下酶的特异性裂解活性:“cVPr”(解蛋白弧菌氨肽酶)、“yPIP”(鼠疫耶氏杆菌脯氨酸亚胺基肽酶)、“D/EAPN”(嗜肺性退伍军人杆菌M1氨肽酶)、hTET(掘越氏火球菌TET氨肽酶)及Pfu API(表7中的“PfuTET”)。相对于末端氨基酸的特异活性如所展示分类,对氨基酸使用单字母缩写(“XP-”表示具有相邻或次末端脯氨酸(P)残基的任何末端氨基酸(X))。
实施例4.在单分子水平上固定肽的末端氨基酸裂解
使用带标记的肽缀合物开发了对固定肽的芯片上氨基酸裂解的分析。分析经设计以提供一种用于确定外肽酶对固定肽的酶识别及裂解活性的方法,其可允许测量动力学结合参数及总体结合亲和力。
为了评估肽的N末端氨基酸裂解,设计且合成了经染料标记的肽,其含有借助于PEG间隔基连接至该染料的N末端天冬氨酸。此肽也含有与被修饰的天冬氨酸相邻的脯氨酸残基,其由酶脯氨酸亚胺基肽酶(来自鼠疫耶氏杆菌,别处已知且在本文中被称作“yPIP”)特异性识别。酶yPIP应仅裂解脯氨酸残基上游的N末端氨基酸。
在表明该标记的肽及其他带标记的肽由yPIP批量有效剪切(例如,如实施例2中所描述)之后,开发了芯片上染料/肽缀合物分析,以在单分子水平上观测N末端氨基酸裂解。图17A展示了染料/肽缀合物分析的通用方案(插图)。如所展示,肽带有标记,该标记经间隔基连接至N末端氨基酸,该肽借助于连接子固定至表面。在暴露于肽酶之后,N末端氨基酸裂解引起带标记的残基自可检测观测体积移除,并通过来自标记的信号的伴随损失来测量。插图右侧的酶-肽复合物大体上描绘了N末端裂解位点。
图17A展示了带标记的肽构建体(在底部),其经设计并合成以用于染料/肽缀合物分析。在这些实验中,将若丹明染料(ATTO Rho6G)连接至肽的N末端天冬氨酸残基,该肽在N末端具有次末端脯氨酸残基。如所展示,肽进一步缀合至增溶DNA连接子,该增溶DNA连接子具有生物素部分,以便进行表面固定。
将带标记的肽缀合物负载至具有样品孔阵列的玻璃芯片上。在负载之前及之后获取芯片的影像,以通过若丹明荧光确定单一占有时样品孔的负载百分比。随后将酶yPIP引入至负载芯片上,且使其在37℃下孵育两小时。在引入yPIP之后取得芯片的影像,且计算损失的绿色染料的百分比,以评估N末端氨基酸裂解。图17B展示了来自实验的成像结果,这些结果显示负载阶段中有6%至7%负载,且在与yPIP一起孵育之后,先前负载的孔中有91%信号损失,这指示N末端氨基酸裂解。图17C展示了来自这些实验的代表性信号迹线,其证明在带标记的肽的负载后染料信号的检测到的增加,及在暴露于yPIP之后染料信号的检测到的损失。
开发了芯片上FRET分析以评估外肽酶识别及裂解活性,作为对单分子水平上N末端氨基酸裂解的进一步确认。图18A大体上描绘了FRET肽缀合物分析(A图)及FRET酶缀合物分析(B图)。在FRET肽缀合物分析(A图)中,固定肽构建体包括连接至连接子的FRET供体标记及在N末端连接的FRET受体标记。当暴露于肽酶时,由来自FRET受体标记的信号的损失检测N末端氨基酸裂解。另外,此设计准许通过追踪来自FRET供体标记的发射,在整个实验中监测肽缀合物的负载。
在FRET酶缀合物分析(B图)中,固定肽构建体包括连接至连接子的FRET对的第一标记,且肽酶经FRET对的第二标记被标记。由可归因于FRET相互作用的荧光增强检测N末端氨基酸裂解,FRET相互作用将在肽酶与肽足够接近且N末端的滞留时间足够的情况下发生。另外,此分析准许通过检测在进行性裂解的情况下随时间推移增加的FRET信号,评估由进行性外肽酶进行的进行性氨基酸裂解。
图18A还在A图下方展示了FRET肽构建体,其经设计及合成以用于A图的FRET肽缀合物分析。如所展示,FRET肽构建体包括若丹明染料(ATTO 647N),其连接至肽的N末端天冬氨酸残基,肽在N末端具有次末端脯氨酸残基。肽进一步缀合至增溶DNA连接子,其连接至用于FRET的花青染料(Cy3B)及用于表面固定的生物素部分。
在此实验中,将FRET肽构建体负载至具有样品孔阵列的玻璃芯片上,且首先由绿色滤光器对所收集的光进行滤光,随后由红色滤光器对所收集的光进行滤光。通过测量通过绿色滤光器及红色滤光器两者的信号来检测FRET肽构建体的负载。当信号仅在绿色滤光器中可测量时检测到末端氨基酸裂解,这表明来自FRET肽构建体的红色染料缀合的N末端氨基酸被yPIP裂解。此检测图案在C图中示出。如所展示,若在添加yPIP之前两种染料均可检测,且在与yPIP一起孵育之后仅绿色染料可见,则可合理地推断检测图案中的此变化是由肽的裂解而非光漂白或肽整体上损失所致。另外,将预期来自单独绿色染料的荧光增加,因为其发射不再由红色染料吸收。
在将FRET肽构建体负载至芯片(已使用膦酸及硅烷通过表面钝化对其进行修饰)上之后,引入yPIP且在数个时间点处获得影像。为了评定整体裂解趋势,对各实验计算(绿色)/(绿色+红色)的比率。此比率随发生的裂解的程度提高。图18B为在与yPIP一起孵育的不同时间点处遍及所有孔的FRET发射比率的曲线图。如所展示,在与yPIP一起孵育期间,绿色染料对荧光发射的比率的贡献随时间推移增加,表明较多N末端天冬氨酸残基已被裂解,留下仅具有绿色染料的截短肽。
随后通过确定在哪些时间点观测到染料荧光,在不同时间点处评估剪切效率。这利用简单的定限进行,例如若在激发期间平均染料发射信号>2.5,则确定染料存在(当施加各对应滤光器时)。展现剪切的孔将随后在实验的负载阶段期间显示绿色染料及红色染料两者,但在暴露于yPIP的时间点处仅显示绿色染料。如图18C中所展示,随着芯片暴露于与yPIP一起孵育的时间变长,逐渐观测到较多剪切。图18D展示了在三个经yPIP处理的时间点中的每一个处显示的展示剪切的示例性信号迹线。
使用已使用葡聚糖通过表面钝化而修饰的芯片,用yPIP及其他肽酶进行额外实验,产生了类似结果,结果表明在将肽酶引入至芯片上之后,随时间推移末端氨基酸裂解增加。图18E为在与yPIP一起孵育的不同时间点处遍及经负载孔的FRET发射比率的曲线图。图18F为在与氨肽酶一起孵育的不同时间点处遍及经负载孔的FRET发射比率的曲线图。总体而言,此处实验证明,使用不同外肽酶及不同标记策略,可在单分子水平上实时检测N末端氨基酸裂解。
实施例5.通过带标记的亲和试剂进行的末端氨基酸区分
蛋白分解途径中所涉及的一种接附蛋白(adaptor protein)被鉴定为了潜在候选物,其用作用于检测N末端芳族残基的带标记的亲和试剂。表达该接附蛋白,即,来自α-变形菌(α-proteobacterium)(根癌农杆菌)的ClpS2,并在暴露的半胱氨酸残基处标记。图19A展示了ClpS2蛋白的晶体结构,其中暴露的半胱氨酸残基展示为棍。暴露的半胱氨酸残基经若丹明染料(ATTO 532)标记。
制备具有不同N末端芳族残基的肽,以测试带标记的ClpS2是否能够在单分子水平上进行N末端氨基酸区分。图19B中展示了来自这些实验的示例性单分子强度迹线。如所展示,信号迹线证明了残基特异性缔合-解离结合图案,其对应于可逆结合具有以下的肽的N末端的带标记的亲和试剂:N末端苯丙氨酸残基(F,顶部信号迹线)、N末端酪氨酸残基(Y,中间信号迹线)或N末端色氨酸残基(W,底部信号迹线)。
进行对单分子轨迹的进一步分析,结果展示于图19C至图19E中。图19C为展示当被带标记的ClpS2可逆结合时,三种N末端残基当中的判别脉冲持续时间(信号峰的持续时间)的曲线图。图19D为展示三种N末端残基当中的判别脉冲间持续时间(信号脉冲之间的持续时间)的曲线图。图19E展示了曲线图,其进一步示出了肽N末端的苯丙氨酸、酪氨酸及色氨酸当中的判别脉冲持续时间。不同N末端残基的平均脉冲持续时间由直方图(A)至(B)及分层直方图(C)视觉化。
针对用作用于亮氨酸识别的带标记的亲和试剂,评估了另一接附蛋白,即来自细长嗜热聚球藻的ClpS(teClpS)。如图19F至图19H中所展示,获自停留时间分析的数据证明,带标记的teClpS蛋白产生了与多肽的末端亮氨酸残基的可检测结合相互作用,其中平均脉冲持续时间为0.71秒。这些实验中所使用的teClpS蛋白的氨基酸序列展示于表1中。
进行了类似实验,以评估作为用于亮氨酸识别的潜在试剂的根癌农杆菌ClpS1及细长聚球藻ClpS2,及作为用于脯氨酸识别的潜在试剂的GID4。图19I展示了来自停留时间分析的示例性结果,其展示了由根癌农杆菌ClpS1对苯丙氨酸、亮氨酸、色氨酸及酪氨酸的可区分识别。图19J展示了来自停留时间分析的示例性结果,其展现了由细长聚球藻ClpS2进行的亮氨酸识别。图19K至图19L展示了来自停留时间分析的示例性结果,其展现了由GID4进行的脯氨酸识别。
实施例6.在降解期间通过识别进行的多肽测序
进行实验以评估在进行中的降解反应期间通过N末端氨基酸识别进行的肽测序。图20A至图20D中展示了来自这些实验的示例性结果,其展示了在相同反应混合物中,使用带标记的ClpS2蛋白及氨肽酶实时进行两个独立多肽测序反应中所获得的单分子强度迹线。在各反应中,通过将肽组合物(10pM)负载至芯片上20分钟,将序列YAAWAAFADDDWK(SEQID NO:78)的多肽经C末端赖氨酸残基固定至芯片表面,并在带标记的亲和试剂(经ATTO542标记的根癌农杆菌ClpS2-V1,500nM)及氨肽酶裂解试剂(VPr,8μM)存在下监测固定肽。
图20A及图20C展示了两种不同测序运行的信号迹线数据,其中顶部图(图20A中的图1,图20C中的图2)展示了全迹线,且底部图(Y、W、F)展示了对应于全迹线中突出显示区域中的每一个的放大区域。图20B及图20D分别以直方图展示了如图20A及图20C中所标记的对应图的迹线数据的脉冲持续时间统计数据。如各测序运行的全信号迹线(图1、图2)所展示,在反应过程中观测到信号脉冲的三个分开的时间间隔。如由放大区域(Y图、W图、F图)突出显示,可基于信号脉冲图案中可观测的差异将三个间隔在视觉上彼此区别。
为了进一步分析信号脉冲数据,对各时间间隔确定脉冲持续时间统计数据(图20B及图20D)。确定脉冲持续时间分布中的差异,对应于针对这些氨基酸分别在利用ClpS2的稳态芯片上结合分析中所观测到的那些,且信号脉冲信息在来自测序运行及单独氨基酸结合分析的间隔之间在表型上一致。
如对信号脉冲信息的分析所证实,在各测序运行的进展中观测到的信号脉冲的三个时间间隔分别对应于Y、W及F的识别图案(图1、图2)。信号脉冲图案之间的中间时段由ClpS2-V1的选择性所致,ClpS2-V1不结合N末端丙氨酸残基。如由全信号迹线所示出,第一间隔对应于Y识别,其之后为在VPr肽酶剪切Y及两个丙氨酸残基时的暂停;之后为对应于W识别的第二间隔,其之后为在VPr肽酶剪切W及两个丙氨酸残基时的另一暂停;且最后第三间隔对应于F识别,随后VPr肽酶剪切掉F且在剩余的ADDDWK肽处停止。这些结果展示,在进行中的降解反应期间通过末端氨基酸识别获得的脉冲持续时间信息,可用于确定区分不同类型的末端氨基酸的特征图案。
实施例7.由带标记的外肽酶进行的末端氨基酸鉴定及裂解
进行了一些研究,以研究单一试剂的潜能,即,能够鉴定肽的末端氨基酸,且同时从肽上裂解末端氨基酸。作为单一试剂,外肽酶必须能够结合至肽,同时保留对末端残基的裂解活性。因此,通过靶向不同外肽酶的原生表面暴露氨基酸,采用传统标记策略进行初始方法。在这些实验中,用荧光染料标记表面暴露的半胱氨酸(-SH)或赖氨酸(-NH
为了更精确地控制标记在外肽酶上存在的位置,且确保各外肽酶分子被单一荧光染料标记(以及消除该染料的脱靶反应性),研究了新标记策略。在这些实验中,使用位点特异性标记策略制备了带标记的外肽酶,其中将含有反应性官能基的非天然氨基酸引入至外肽酶中(参见例如Chin,J.W.等人J Am Chem Soc.2002年8月7日;124(31):9026-9027)。
通过将位置287处的赖氨酸残基突变为具有对叠氮苯丙氨酸(pAzF)侧链的残基,修饰来自鼠疫耶氏杆菌的脯氨酸亚胺基肽酶(yPIP)。图21A展示了yPIP的晶体结构,其中由pAzF的化学结构指示的突变用展示为棍的K287侧链展示。此突变位点是基于由此位置处的α螺旋提供的稳定性而选择的,且其经选择以确保新叠氮官能基是溶剂暴露的。
获得了pEVOL质粒,其含有将pAzF并入至氨基酸链中所必需的突变氨基tRNA合成酶及突变tRNA。随后使用QuickChange II突变诱发试剂盒,将对特异性并入pAzF而言必需的琥珀终止密码子(TAG)引入至cDNA中。随后对cDNA进行测序,并确认TAG密码子位置。此后,共转染含有yPIP琥珀突变体的pET21b+质粒,及含有用于将pAzF装入琥珀密码子的tRNA的细胞机构的pEVOL质粒两者。随后使共转染细胞生长至0.8ODU,在于2L LB中的2mM pAzF存在下,用0.02%阿拉伯糖及1mM IPTG诱导,并使用化学分解收集。使用5mL亲和色谱管柱进行纯化,并在100mM咪唑中洗脱蛋白质。随后将所得蛋白质透析并浓缩至50mM HEPES pH8.0及0.5M KCl中,等分,并急骤冷冻,随后储存在-20℃下。
为了确认叠氮基存在于纯化蛋白质中,使DBCO-Cy3(2mM)与pAzF-yPIP变体(220μM)反应(反应条件:50mM HEPES pH 8.0,0.5mM KCl,20%DMSO;37℃下10小时,室温下48小时)。通过尺寸排阻色谱纯化蛋白质反应产物,并确定所得蛋白质100%被叠氮化合物反应性DBCO-Cy3试剂标记(图21B),指示非天然氨基酸的稳健并入。
通过对不带标记的及带标记的pAzF变体的SDS-PAGE分析确认最终产物的蛋白质标记及纯度。图21C展示了SDS-PAGE凝胶的图片,其证实pAzF-yPIP的Cy3标记(图21D中展示了凝胶的过度曝光影像以展示梯状条带(ladder))。图21E展示了考马斯染色凝胶的图片,其证实染料及蛋白质两者共同迁移且是纯的。
在活性分析中使用了被染料标记的pAzF-yPIP,以确认在标记及纯化之后酶仍然具有活性。如图21F中所展示,如通过HPLC测量,使用1000倍过量底物,Cy3-pAzF-yPIP能够在1小时内水解100%的肽底物。这些实验证明了一种方法,该方法允许在对天然蛋白质结构/功能扰动极小的情况下,对外肽酶进行位点特异性修饰及标记。
实施例8.在多肽测序中对被修饰的氨基酸的识别
进行了实验,以评估对含有特定翻译后修饰的氨基酸的识别。测试了来自酪氨酸激酶Fyn的Src同源2(SH2)结构域的三重突变变体(T8V、S10A、K15L)作为肽测序中磷酸化酪氨酸残基的潜在识别分子。将变体蛋白固定至样品孔底部,且在添加含有N末端磷酸化酪氨酸的荧光标记的肽后,收集单分子信号迹线。如图22A中的代表性迹线所展示,在这些实验期间,检测了由固定蛋白质进行的肽结合。图22B中展示了在这些实验期间所收集的脉冲持续时间数据(顶部、中间及底部曲线图分别对应于图22A的顶部、中间及底部迹线)。图22C中展示了脉冲持续时间及脉冲间持续时间统计数据(分别顶部图及底部图)。
进行了对照实验,以确认Fyn蛋白对磷酸化酪氨酸具有特异性。针对三种不同肽中的每一种重复实验:第一肽,其含有N末端未被修饰的酪氨酸(Y;图22D);第二肽,其含有N末端及次末端未被修饰的酪氨酸(YY;图22E);及第三肽,其含有N末端磷酸化丝氨酸(图22F)。如所展示,在阴性对照实验中所使用的肽中的任一种的情况下,未检测到结合。
实施例9.在多肽测序中对次末端氨基酸的识别
进行了实验,以确定次末端氨基酸对根癌农杆菌ClpS2-V1的脉冲持续时间的影响。制备了四十九种不同的荧光标记的肽,在N末端含有独特的二肽序列,其中N末端氨基酸为F、W或Y,且次末端位置为20种天然氨基酸中的一种。对于各实验,将ClpS2-V1固定在样品孔底部,且在添加荧光标记的肽中的一种后,收集单分子信号迹线10至20分钟。对于各肽,针对最少50个样品孔收集脉冲持续时间数据。
图23展示了50种肽中的每一个的中值脉冲持续时间,其中数据点由次末端氨基酸(x轴)分组,且N末端氨基酸用不同符号表示。
实施例10.利用多种识别分子进行的同时氨基酸识别
进行了单分子肽识别实验,以证明多于一种带标记的识别分子对固定肽的末端氨基酸识别。将含有N末端苯丙氨酸的单一肽分子(FYPLPWPDDDY(SEQ ID NO:79))固定于芯片的样品孔中。添加含有500nM atClpS1(根癌农杆菌ClpS1;序列提供于表1中)及atClpS2-V1(根癌农杆菌ClpS2变体1;序列提供于表1中)中的每一个的缓冲液,其中atClpS1及atClpS2-V1分别被Cy3及Cy3B标记。由于Cy3B的强度高于Cy3,因此atClpS2-V1结合事件可容易地与atClpS1结合事件区别。
图24A至图24C展示了实验的结果,其展示了利用差异性标记的识别分子的单分子肽识别。图24A中显示了代表性迹线。对于各结合子,脉冲持续时间分布相异(图24B),且对应于如单一结合子实验中所观测到的其动力学概况。对于atClpS1,平均脉冲持续时间为1.3秒;且对于atClpS2-V1,平均脉冲持续时间为为1.0秒(图24C)。脉冲速率也相异:对于atClpS1,8.1个脉冲/分钟;且对于atClp2-V1,14.1个脉冲/分钟(图24C)。因此,当包括多于一种识别分子以便动态识别固定肽时,各识别分子的结合特征(包括脉冲持续时间、脉冲间持续时间及脉冲速率)可同时提供关于肽序列的信息。
实施例11.用识别分子连接子增强光稳定性
进行了实验,以评估在单分子测序期间固定肽的光稳定性。在532nm激发光的存在下,将实施例5中所描述的经染料标记的atClpS2-V1添加至含有固定肽底物的样品孔中,以通过来自ATTO 532的发射来监测识别。图25A中展示了代表性迹线。如顶部图中所展示,观测到识别在进入实验大约600秒时停止。底部图为展示进入反应大约180秒至430秒时的信号脉冲的放大视图。
图25B展示了这些实验中所使用的ClpS2蛋白的晶体结构的视觉化。如所展示,充当染料缀合位点的半胱氨酸残基距末端氨基酸结合位点大约2nm。假设对肽的光损伤是由在结合期间染料与肽的N末端接近而引起。为了减轻染料接近的潜在光损伤效应,将ClpS2蛋白经连接子进行染料标记,该连接子使染料与肽的N末端之间的距离增加超过10nm。连接子包括抗生物素蛋白链菌素及双链核酸;该双链核酸用两个Cy3B染料分子标记,且经双生物素部分连接至抗生物素蛋白链菌素,且ClpS2蛋白经生物素部分连接至抗生物素蛋白链菌素上其余两个结合位点中的每一个。图25C中展示了使用该染料屏蔽的ClpS2分子的代表性迹线。如顶部图中所展示,识别时间延长至进入实验大约6,000秒。底部图为展示进入反应大约750秒至930秒时的信号脉冲的放大视图。
用连接子产生了DNA-抗生物素蛋白链菌素识别分子,该连接子含有被两个Cy3B染料分子标记且经双生物素部分连接至抗生物素蛋白链菌素的双链核酸,及经双生物素部分连接至抗生物素蛋白链菌素上其余两个结合位点的单一ClpS2蛋白。在单分子肽测序反应中使用了该构建体,且来自这些实验的代表性迹线展示于图26A至图26D中。
重复实施例6中所描述的测序实验,其中反应条件如下改变:将DNA-抗生物素蛋白链菌素ClpS2识别分子与hTET氨基酸裂解试剂组合使用。代表性信号迹线展示于图27中。
实施例12.在由多种外肽酶降解期间通过识别进行的测序
进行了实验,以评估具有差异性裂解特异性的多种类型的外肽酶在单分子肽测序反应混合物中的用途。将单一肽分子(YAAWAAFADDDWK(SEQ ID NO:78))经C末端赖氨酸残基固定于芯片的样品孔中。添加含有用于氨基酸识别的atClpS2-V1及用于氨基酸裂解的hTET的缓冲液。代表性迹线显示于图28A中,脉冲图案区域的展开图展示于图28B中。
进行了实验,以评估在具有差异性特异性的两种类型的外肽酶存在下的测序反应。将单一肽分子(FYPLPWPDDDYK(SEQ ID NO:80))经C末端赖氨酸残基固定于芯片的样品孔中。添加含有用于氨基酸识别的atClpS2-V1及用于氨基酸裂解的hTET及yPIP两者的缓冲液。代表性迹线显示于图28C中,脉冲图案区域的展开图展示于图28D中。来自这些反应条件的额外代表性迹线展示于图28E中。
进行了进一步实验,以评估在具有差异性特异性的两种类型的外肽酶存在下的测序反应。将单一肽分子(YPLPWPDDDYK(SEQ ID NO:81))经C末端赖氨酸残基固定于芯片的样品孔中。在一个实验中,添加缓冲液,缓冲液含有用于氨基酸识别的atClpS2-V1及用于氨基酸裂解的hTET及yPIP两者。代表性迹线显示于图28F中,脉冲图案区域的展开图展示于图28G中。来自这些反应条件的额外代表性迹线展示于图28H中。在进一步实验中,添加缓冲液(50mM MOPS,60mM KOAc,200μM Co(OAc)
等效物及范畴
在申请专利范围中,除非相反地指示或另外从上下文显而易见,否则诸如“一(a/an)”及“该”的冠词可意味一或超过一。除非相反地指示或以其他方式从上下文显而易见,否则若一个、多于一个或所有群成员存在于给定产物或方法中、用于给定产物或方法中或以其他方式与给定产物或方法相关,则在该群的一个或多个成员之间包括“或”的申请专利范围或描述视为满足。本发明包括群中恰好一个成员存在于给定产物或方法中、用于给定产物或方法中或以其他方式与给定产物或方法相关的实施例。本发明包括多于一个或所有群成员存在于给定产物或方法中、用于给定产物或方法中或以其他方式与给定产物或方法相关的实施例。
此外,本发明涵盖其中来自一或多条所列权利要求的一个或多个限制、要素、条款及描述性用语经引入另一条权利要求中的所有变化、组合及排列。举例而言,依附于另一权利要求的任何申权利要求可经修改以包括在依附于同一基本权利要求的任何其他权利要求中可见的一个或多个限制。在要素以列表形式,例如呈马库什(Markush)组形式呈现的情况下,亦揭示要素的各子组,且可从该组移除任何要素。应理解,一般而言,在本发明或本发明的方面称为包括具体要素和/或特征时,本发明或本发明方面的某些实施方案由此类要素和/或特征组成或主要由此类要素和/或特征组成。出于简单的目的,这些实施例尚未具体地以词语阐述在本文中。
如本文在说明书及申请专利范围中使用,词组“和/或”应理解为意味如此结合的要素的“任一种或两者”,亦即,在一些情况下结合地存在且在其他情况下未结合地存在的要素。使用“和/或”列出的多个要素应以相同方式解释,亦即,如此结合的“一个或多个”要素。可任选地,存在除了通过“和/或”条项所确切地鉴定的要素以外的其他要素,无论与确切地鉴定的这些要素相关抑或不相关。因此,作为非限制性实例,参考“A和/或B”在结合诸如“包括”的开放式措辞使用时,在一个实施例中,可仅指A(任选地,包括除了B以外的要素);在另一实施例中,可仅指B(任选地,包括除了A以外的要素);在另一实施例中,可指A及B两者(任选地,包括其他要素);等。
如在本说明书及申请专利范围中所用,“或”应理解为具有与如上文所定义的“和/或”相同的含义。举例而言,当分离列表中的项目时,“或”或“和/或”应解释为包括性的,亦即,包括一些或一列要素中的至少一个而且包括多于一个,及(任选地,)其他未列出的项目。只有指示截然相反的术语,诸如“中的仅一者”或“中的恰好一者”或当用于申请专利范围中时“由……组成”将指包括一些或一列要素中的恰好一个要素。一般而言,当置于排他性术语,诸如“任一种”、“中的一者”、“中的仅一者”或“中的恰好一者”之前时,如本文中所使用的术语“或”应仅解释为表明排他性替代方式(亦即“一者或另一者但非二者皆”)。当用于申请专利范围中时,“主要由……组成”应具有如其在专利法律领域中所使用的普通含义。
如本说明书及申请专利范围中所用,参考一个或多个要素的列表的词组“至少一个”应理解为意味选自要素列表中的任何一个或多个要素的至少一个要素,但未必包括要素列表内具体列出的每一个要素中的至少一者,且未必排除要素列表中的要素的任何组合。此定义也允许可任选地,存在除词组“至少一个”所指的要素列表内具体鉴定的要素以外的要素,而无论与具体鉴定的这些要素相关抑或不相关。因此,作为非限制性实例,“A及B中的至少一者”(或等效地“A或B中的至少一者”或等效地“A和/或B中的至少一者”)在一个实施例中可指至少一个(任选地,包括多于一个)A而不存在B(且任选地,包括除了B以外的要素);在另一实施例中,指至少一个(任选地,包括多于一个)B而不存在A (且任选地,包括除了A以外的要素);在又一实施例中,指至少一个(任选地,包括多于一个)A及至少一个(任选地,包括多于一个)B(且任选地,包括其他要素);等。
还应理解,除非截然相反地指示,否则在本文中所主张的包括超过一个步骤或操作的任何方法中,该方法的步骤或操作的顺序并非必需限制于列举该方法的步骤或操作的顺序。
在申请专利范围中以及在上述说明书中,所有过渡性词组,诸如“包括”、“包括”、“携有”、“具有”、“含有”、“涉及”、“拥有”、“由……构成”及其类似词组,应理解为开放的,亦即,意味包括但不限于。仅过渡词组“由……组成”及“主要由……组成”应分别为封闭或半封闭过渡词组,如美国专利局专利审查程序手册(United States Patent Office Manualof Patent Examining Procedures)第2111.03节中所阐述。应了解,在替代实施例中,亦涵盖此文件中使用开放式过渡词组(例如“包括”)描述的实施例,如“由……组成”及“主要由……组成”由开放式过渡词组描述的特征。举例而言,若本申请描述“包括A及B的组合物”,则本申请亦涵盖替代实施例“由A及B组成的组合物”及“主要由A及B组成的组合物”。
当给出范围时,包括端点。此外,除非另外指示或另外从上下文及一般技术者的理解显而易见,否则表示为范围的值可在本发明的不同实施例中采用所述范围内的任何特定值或子范围,除非上下文另外明确规定,否则达到该范围下限的单位的十分之一。
本申请提及各种颁予的专利、公开的专利申请、期刊文章及其他出版物,以上所有者均以引用的方式并入本文中。若任何并入的参考文献与本说明书之间存在冲突,则应以本说明书为准。另外,本发明的属于先前技术的任何特定实施例可明确地从申请专利范围中的任一或多项排除。因为认为此类实施例为一般技术者所已知,所以其可经排除,即使未在本文中明确地阐述该排除。本发明的任何特定实施例可出于任何原因从任何权利要求排除,无论是否与先前技术的存在相关。
本领域技术人员最多使用常规实验将认识到或能够确定本文所描述的特定实施例的许多等效物。本文所描述的本发明实施例的范畴并不意欲限于以上描述,而是实际上如所附申请专利范围中所阐述。一般技术者将了解,可在不脱离如以下申请专利范围所定义的本发明的精神或范畴的情况下对本说明书进行各种改变及修改。
本文中的变量的任何定义下的化学基团列表的列举包括呈任何单一基团或所列基团组合的该变量的定义。本文中的变量的实施例的列举包括呈任何单一实施例或与任何其他实施例或其部分组合的形式的实施例。本文中的实施例的列举包括呈任何单一实施例或与任何其他实施例或其部分组合的形式的实施例。
序列表
<110> 宽腾矽公司
<120> 用于蛋白质测序的方法及组合物
<130> R0708.70042WO00
<140> 尚未指定
<141> 与此同日
<150> US 62/907,507
<151> 2019-09-27
<150> US 62/768,076
<151> 2018-11-15
<160> 158
<170> PatentIn version 3.5
<210> 1
<211> 103
<212> PRT
<213> 根癌农杆菌
<400> 1
Met Ser Asp Ser Pro Val Asp Leu Lys Pro Lys Pro Lys Val Lys Pro
1 5 10 15
Lys Leu Glu Arg Pro Lys Leu Tyr Lys Val Met Leu Leu Asn Asp Asp
20 25 30
Tyr Thr Pro Met Ser Phe Val Thr Val Val Leu Lys Ala Val Phe Arg
35 40 45
Met Ser Glu Asp Thr Gly Arg Arg Val Met Met Thr Ala His Arg Phe
50 55 60
Gly Ser Ala Val Val Val Val Cys Glu Arg Asp Ile Ala Glu Thr Lys
65 70 75 80
Ala Lys Glu Ala Thr Asp Leu Gly Lys Glu Ala Gly Phe Pro Leu Met
85 90 95
Phe Thr Thr Glu Pro Glu Glu
100
<210> 2
<211> 103
<212> PRT
<213> 根癌农杆菌
<400> 2
Met Ser Asp Ser Pro Val Asp Leu Lys Pro Lys Pro Lys Val Lys Pro
1 5 10 15
Lys Leu Glu Arg Pro Lys Leu Tyr Lys Val Met Leu Leu Asn Asp Asp
20 25 30
Tyr Thr Pro Arg Glu Phe Val Thr Val Val Leu Lys Ala Val Phe Arg
35 40 45
Met Ser Glu Asp Thr Gly Arg Arg Val Met Met Thr Ala His Arg Phe
50 55 60
Gly Ser Ala Val Val Val Val Cys Glu Arg Asp Ile Ala Glu Thr Lys
65 70 75 80
Ala Lys Glu Ala Thr Asp Leu Gly Lys Glu Ala Gly Phe Pro Leu Met
85 90 95
Phe Thr Thr Glu Pro Glu Glu
100
<210> 3
<211> 103
<212> PRT
<213> 根癌农杆菌
<400> 3
Met Ser Asp Ser Pro Val Asp Leu Lys Pro Lys Pro Lys Val Lys Pro
1 5 10 15
Lys Leu Glu Arg Pro Lys Leu Tyr Lys Val Met Leu Leu Asn Asp Asp
20 25 30
Tyr Thr Pro Arg Glu Phe Val Thr Val Val Leu Lys Ala Val Phe Arg
35 40 45
Met Ser Glu Asp Thr Gly Arg Arg Val Met Met Thr Ala His Arg Phe
50 55 60
Gly Ser Ala Val Val Val Val Ser Glu Arg Asp Ile Ala Glu Thr Lys
65 70 75 80
Ala Lys Glu Ala Thr Asp Leu Gly Lys Glu Ala Gly Phe Pro Leu Met
85 90 95
Phe Thr Thr Glu Pro Glu Glu
100
<210> 4
<211> 117
<212> PRT
<213> 根癌农杆菌
<400> 4
Met Ile Ala Glu Pro Ile Cys Met Gln Gly Glu Gly Asp Gly Glu Asp
1 5 10 15
Gly Gly Thr Asn Arg Gly Thr Ser Val Ile Thr Arg Val Lys Pro Lys
20 25 30
Thr Lys Arg Pro Asn Leu Tyr Arg Val Leu Leu Leu Asn Asp Asp Tyr
35 40 45
Thr Pro Met Glu Phe Val Ile His Ile Leu Glu Arg Phe Phe Gln Lys
50 55 60
Asp Arg Glu Ala Ala Thr Arg Ile Met Leu His Val His Gln His Gly
65 70 75 80
Val Gly Glu Cys Gly Val Phe Thr Tyr Glu Val Ala Glu Thr Lys Val
85 90 95
Ser Gln Val Met Asp Phe Ala Arg Gln His Gln His Pro Leu Gln Cys
100 105 110
Val Met Glu Lys Lys
115
<210> 5
<211> 103
<212> PRT
<213> 根癌农杆菌
<400> 5
Met Ser Asp Ser Pro Val Asp Leu Lys Pro Lys Pro Lys Val Lys Pro
1 5 10 15
Lys Leu Glu Arg Pro Lys Leu Tyr Lys Val Met Leu Leu Asn Asp Asp
20 25 30
Tyr Thr Pro Met Ser Phe Val Thr Val Val Leu Lys Ala Val Phe Arg
35 40 45
Met Ser Glu Asp Thr Gly Arg Arg Val Met Met Thr Ala His Arg Phe
50 55 60
Gly Ser Ala Val Val Val Val Ser Glu Arg Asp Ile Ala Glu Thr Lys
65 70 75 80
Ala Lys Glu Ala Thr Asp Leu Gly Lys Glu Ala Gly Phe Pro Leu Met
85 90 95
Phe Thr Thr Glu Pro Glu Glu
100
<210> 6
<211> 117
<212> PRT
<213> 根癌农杆菌
<400> 6
Met Ile Ala Glu Pro Ile Ser Met Gln Gly Glu Gly Asp Gly Glu Asp
1 5 10 15
Gly Gly Thr Asn Arg Gly Thr Ser Val Ile Thr Arg Val Lys Pro Lys
20 25 30
Thr Lys Arg Pro Asn Leu Tyr Arg Val Leu Leu Leu Asn Asp Asp Tyr
35 40 45
Thr Pro Met Glu Phe Val Ile His Ile Leu Glu Arg Phe Phe Gln Lys
50 55 60
Asp Arg Glu Ala Ala Thr Arg Ile Met Leu His Val His Gln His Gly
65 70 75 80
Val Gly Glu Cys Gly Val Phe Thr Tyr Glu Val Ala Glu Thr Lys Val
85 90 95
Ser Gln Val Met Asp Phe Ala Arg Gln His Gln His Pro Leu Gln Cys
100 105 110
Val Met Glu Lys Lys
115
<210> 7
<211> 117
<212> PRT
<213> 根癌农杆菌
<400> 7
Met Ile Ala Glu Pro Ile Ser Met Gln Gly Glu Gly Asp Gly Glu Asp
1 5 10 15
Gly Gly Thr Asn Arg Gly Thr Ser Val Ile Thr Arg Val Lys Pro Lys
20 25 30
Thr Lys Arg Pro Asn Leu Tyr Arg Val Leu Leu Leu Asn Asp Asp Tyr
35 40 45
Thr Pro Met Glu Phe Val Ile His Ile Leu Glu Arg Phe Phe Gln Lys
50 55 60
Asp Arg Glu Ala Ala Thr Arg Ile Met Leu His Val His Gln His Gly
65 70 75 80
Val Gly Glu Ser Gly Val Phe Thr Tyr Glu Val Ala Glu Thr Lys Val
85 90 95
Ser Gln Val Met Asp Phe Ala Arg Gln His Gln His Pro Leu Gln Ser
100 105 110
Val Met Glu Lys Lys
115
<210> 8
<211> 103
<212> PRT
<213> 根癌农杆菌
<400> 8
Met Ser Asp Ser Pro Val Asp Leu Lys Pro Lys Pro Lys Val Lys Pro
1 5 10 15
Lys Leu Glu Arg Pro Lys Leu Tyr Lys Val Ile Leu Leu Asn Asp Asp
20 25 30
Tyr Thr Pro Met Glu Phe Val Val Glu Val Leu Lys Arg Val Phe Asn
35 40 45
Met Ser Glu Glu Gln Ala Arg Arg Val Met Met Thr Ala His Lys Lys
50 55 60
Gly Lys Ala Val Val Gly Val Cys Pro Arg Asp Ile Ala Glu Thr Lys
65 70 75 80
Ala Lys Gln Ala Thr Asp Leu Ala Arg Glu Ala Gly Phe Pro Leu Met
85 90 95
Phe Thr Thr Glu Pro Glu Glu
100
<210> 9
<211> 103
<212> PRT
<213> 根癌农杆菌
<400> 9
Met Ser Asp Ser Pro Val Asp Leu Lys Pro Lys Pro Lys Val Lys Pro
1 5 10 15
Lys Leu Glu Arg Pro Lys Leu Tyr Lys Val Ile Leu Leu Asn Asp Asp
20 25 30
Tyr Thr Pro Met Glu Phe Val Val Glu Val Leu Lys Arg Val Phe Asn
35 40 45
Met Ser Glu Glu Gln Ala Arg Arg Val Met Met Thr Ala His Lys Lys
50 55 60
Gly Lys Ala Val Val Gly Val Ser Pro Arg Asp Ile Ala Glu Thr Lys
65 70 75 80
Ala Lys Gln Ala Thr Asp Leu Ala Arg Glu Ala Gly Phe Pro Leu Met
85 90 95
Phe Thr Thr Glu Pro Glu Glu
100
<210> 10
<211> 95
<212> PRT
<213> 细长聚球藻
<400> 10
Met Ala Val Glu Thr Ile Gln Lys Pro Glu Thr Thr Thr Lys Arg Lys
1 5 10 15
Ile Ala Pro Arg Tyr Arg Val Leu Leu His Asn Asp Asp Phe Asn Pro
20 25 30
Met Glu Tyr Val Val Met Val Leu Met Gln Thr Val Pro Ser Leu Thr
35 40 45
Gln Pro Gln Ala Val Asp Ile Met Met Glu Ala His Thr Asn Gly Thr
50 55 60
Gly Leu Val Ile Thr Cys Asp Ile Glu Pro Ala Glu Phe Tyr Cys Glu
65 70 75 80
Gln Leu Lys Ser His Gly Leu Ser Ser Ser Ile Glu Pro Asp Asp
85 90 95
<210> 11
<211> 136
<212> PRT
<213> 细长聚球藻
<400> 11
Met Ser Pro Gln Pro Asp Glu Ser Val Leu Ser Ile Leu Gly Val Pro
1 5 10 15
Arg Pro Cys Val Lys Lys Arg Ser Arg Asn Asp Ala Phe Val Leu Thr
20 25 30
Val Leu Thr Cys Ser Leu Gln Ala Ile Ala Ala Pro Ala Thr Ala Pro
35 40 45
Gly Thr Thr Thr Thr Arg Val Arg Gln Pro Tyr Pro His Phe Arg Val
50 55 60
Ile Val Leu Asp Asp Asp Val Asn Thr Phe Gln His Val Ala Glu Cys
65 70 75 80
Leu Leu Lys Tyr Ile Pro Gly Met Thr Gly Asp Arg Ala Trp Asp Leu
85 90 95
Thr Asn Gln Val His Tyr Glu Gly Ala Ala Thr Val Trp Ser Gly Pro
100 105 110
Gln Glu Gln Ala Glu Leu Tyr His Glu Gln Leu Arg Arg Glu Gly Leu
115 120 125
Thr Met Ala Pro Leu Glu Ala Ala
130 135
<210> 12
<211> 90
<212> PRT
<213> 细长嗜热聚球藻
<400> 12
Met Pro Gln Glu Arg Gln Gln Val Thr Arg Lys His Tyr Pro Asn Tyr
1 5 10 15
Lys Val Ile Val Leu Asn Asp Asp Phe Asn Thr Phe Gln His Val Ala
20 25 30
Ala Cys Leu Met Lys Tyr Ile Pro Asn Met Thr Ser Asp Arg Ala Trp
35 40 45
Glu Leu Thr Asn Gln Val His Tyr Glu Gly Gln Ala Ile Val Trp Val
50 55 60
Gly Pro Gln Glu Gln Ala Glu Leu Tyr His Glu Gln Leu Leu Arg Ala
65 70 75 80
Gly Leu Thr Met Ala Pro Leu Glu Pro Glu
85 90
<210> 13
<211> 106
<212> PRT
<213> 大肠杆菌
<400> 13
Met Gly Lys Thr Asn Asp Trp Leu Asp Phe Asp Gln Leu Ala Glu Glu
1 5 10 15
Lys Val Arg Asp Ala Leu Lys Pro Pro Ser Met Tyr Lys Val Ile Leu
20 25 30
Val Asn Asp Asp Tyr Thr Pro Met Glu Phe Val Ile Asp Val Leu Gln
35 40 45
Lys Phe Phe Ser Tyr Asp Val Glu Arg Ala Thr Gln Leu Met Leu Ala
50 55 60
Val His Tyr Gln Gly Lys Ala Ile Cys Gly Val Phe Thr Ala Glu Val
65 70 75 80
Ala Glu Thr Lys Val Ala Met Val Asn Lys Tyr Ala Arg Glu Asn Glu
85 90 95
His Pro Leu Leu Cys Thr Leu Glu Lys Ala
100 105
<210> 14
<211> 106
<212> PRT
<213> 大肠杆菌
<400> 14
Met Gly Lys Thr Asn Asp Trp Leu Asp Phe Asp Gln Leu Ala Glu Glu
1 5 10 15
Lys Val Arg Asp Ala Leu Lys Pro Pro Ser Met Tyr Lys Val Ile Leu
20 25 30
Val Asn Asp Asp Tyr Thr Pro Ala Glu Phe Val Ile Asp Val Leu Gln
35 40 45
Lys Phe Phe Ser Tyr Asp Val Glu Arg Ala Thr Gln Leu Met Leu Ala
50 55 60
Val His Tyr Gln Gly Lys Ala Ile Cys Gly Val Phe Thr Ala Glu Val
65 70 75 80
Ala Glu Thr Lys Val Ala Met Val Asn Lys Tyr Ala Arg Glu Asn Glu
85 90 95
His Pro Leu Leu Cys Thr Leu Glu Lys Ala
100 105
<210> 15
<211> 192
<212> PRT
<213> 恶性疟原虫
<400> 15
Met Phe Lys Asp Leu Lys Pro Phe Phe Leu Cys Ile Ile Leu Leu Leu
1 5 10 15
Leu Leu Ile Tyr Lys Cys Thr His Ser Tyr Asn Ile Lys Asn Lys Asn
20 25 30
Cys Pro Leu Asn Phe Met Asn Ser Cys Val Arg Ile Asn Asn Val Asn
35 40 45
Lys Asn Thr Asn Ile Ser Phe Pro Lys Glu Leu Gln Lys Arg Pro Ser
50 55 60
Leu Val Tyr Ser Gln Lys Asn Phe Asn Leu Glu Lys Ile Lys Lys Leu
65 70 75 80
Arg Asn Val Ile Lys Glu Ile Lys Lys Asp Asn Ile Lys Glu Ala Asp
85 90 95
Glu His Glu Lys Lys Glu Arg Glu Lys Glu Thr Ser Ala Trp Lys Val
100 105 110
Ile Leu Tyr Asn Asp Asp Ile His Asn Phe Thr Tyr Val Thr Asp Val
115 120 125
Ile Val Lys Val Val Gly Gln Ile Ser Lys Ala Lys Ala His Thr Ile
130 135 140
Thr Val Glu Ala His Ser Thr Gly Gln Ala Leu Ile Leu Ser Thr Trp
145 150 155 160
Lys Ser Lys Ala Glu Lys Tyr Cys Gln Glu Leu Gln Gln Asn Gly Leu
165 170 175
Thr Val Ser Ile Ile His Glu Ser Gln Leu Lys Asp Lys Gln Lys Lys
180 185 190
<210> 16
<211> 236
<212> PRT
<213> 大肠杆菌
<400> 16
Met Arg Leu Val Gln Leu Ser Arg His Ser Ile Ala Phe Pro Ser Pro
1 5 10 15
Glu Gly Ala Leu Arg Glu Pro Asn Gly Leu Leu Ala Leu Gly Gly Asp
20 25 30
Leu Ser Pro Ala Arg Leu Leu Met Ala Tyr Gln Arg Gly Ile Phe Pro
35 40 45
Trp Phe Ser Pro Gly Asp Pro Ile Leu Trp Trp Ser Pro Asp Pro Arg
50 55 60
Ala Val Leu Trp Pro Glu Ser Leu His Ile Ser Arg Ser Met Lys Arg
65 70 75 80
Phe His Lys Arg Ser Pro Tyr Arg Val Thr Met Asn Tyr Ala Phe Gly
85 90 95
Gln Val Ile Glu Gly Cys Ala Ser Asp Arg Glu Glu Gly Thr Trp Ile
100 105 110
Thr Arg Gly Val Val Glu Ala Tyr His Arg Leu His Glu Leu Gly His
115 120 125
Ala His Ser Ile Glu Val Trp Arg Glu Asp Glu Leu Val Gly Gly Met
130 135 140
Tyr Gly Val Ala Gln Gly Thr Leu Phe Cys Gly Glu Ser Met Phe Ser
145 150 155 160
Arg Met Glu Asn Ala Ser Lys Thr Ala Leu Leu Val Phe Cys Glu Glu
165 170 175
Phe Ile Gly His Gly Gly Lys Leu Ile Asp Cys Gln Val Leu Asn Asp
180 185 190
His Thr Ala Ser Leu Gly Ala Cys Glu Ile Pro Arg Arg Asp Tyr Leu
195 200 205
Asn Tyr Leu Asn Gln Met Arg Leu Gly Arg Leu Pro Asn Asn Phe Trp
210 215 220
Val Pro Arg Cys Leu Phe Ser Pro Gln Glu Leu Glu
225 230 235
<210> 17
<211> 231
<212> PRT
<213> 创伤弧菌
<400> 17
Met Ser Ser Asp Ile His Gln Ile Lys Ile Gly Leu Thr Asp Asn His
1 5 10 15
Pro Cys Ser Tyr Leu Pro Glu Arg Lys Glu Arg Val Ala Val Ala Leu
20 25 30
Glu Ala Asp Met His Thr Ala Asp Asn Tyr Glu Val Leu Leu Ala Asn
35 40 45
Gly Phe Arg Arg Ser Gly Asn Thr Ile Tyr Lys Pro His Cys Asp Ser
50 55 60
Cys His Ser Cys Gln Pro Ile Arg Ile Ser Val Pro Asp Ile Glu Leu
65 70 75 80
Ser Arg Ser Gln Lys Arg Leu Leu Ala Lys Ala Arg Ser Leu Ser Trp
85 90 95
Ser Met Lys Arg Asn Met Asp Glu Asn Trp Phe Asp Leu Tyr Ser Arg
100 105 110
Tyr Ile Val Ala Arg His Arg Asn Gly Thr Met Tyr Pro Pro Lys Lys
115 120 125
Asp Asp Phe Ala His Phe Ser Arg Asn Gln Trp Leu Thr Thr Gln Phe
130 135 140
Leu His Ile Tyr Glu Gly Gln Arg Leu Ile Ala Val Ala Val Thr Asp
145 150 155 160
Ile Met Asp His Cys Ala Ser Ala Phe Tyr Thr Phe Phe Glu Pro Glu
165 170 175
His Glu Leu Ser Leu Gly Thr Leu Ala Val Leu Phe Gln Leu Glu Phe
180 185 190
Cys Gln Glu Glu Lys Lys Gln Trp Leu Tyr Leu Gly Tyr Gln Ile Asp
195 200 205
Glu Cys Pro Ala Met Asn Tyr Lys Val Arg Phe His Arg His Gln Lys
210 215 220
Leu Val Asn Gln Arg Trp Gln
225 230
<210> 18
<211> 83
<212> PRT
<213> 酿酒酵母
<400> 18
Met Gly Ser Val His Lys His Thr Gly Arg Asn Cys Gly Arg Lys Phe
1 5 10 15
Lys Ile Gly Glu Pro Leu Tyr Arg Cys His Glu Cys Gly Cys Asp Asp
20 25 30
Thr Cys Val Leu Cys Ile His Cys Phe Asn Pro Lys Asp His Val Asn
35 40 45
His His Val Cys Thr Asp Ile Cys Thr Glu Phe Thr Ser Gly Ile Cys
50 55 60
Asp Cys Gly Asp Glu Glu Ala Trp Asn Ser Pro Leu His Cys Lys Ala
65 70 75 80
Glu Glu Gln
<210> 19
<211> 167
<212> PRT
<213> 智人
<400> 19
Met Ser Gly Ser Lys Phe Arg Gly His Gln Lys Ser Lys Gly Asn Ser
1 5 10 15
Tyr Asp Val Glu Val Val Leu Gln His Val Asp Thr Gly Asn Ser Tyr
20 25 30
Leu Cys Gly Tyr Leu Lys Ile Lys Gly Leu Thr Glu Glu Tyr Pro Thr
35 40 45
Leu Thr Thr Phe Phe Glu Gly Glu Ile Ile Ser Lys Lys His Pro Phe
50 55 60
Leu Thr Arg Lys Trp Asp Ala Asp Glu Asp Val Asp Arg Lys His Trp
65 70 75 80
Gly Lys Phe Leu Ala Phe Tyr Gln Tyr Ala Lys Ser Phe Asn Ser Asp
85 90 95
Asp Phe Asp Tyr Glu Glu Leu Lys Asn Gly Asp Tyr Val Phe Met Arg
100 105 110
Trp Lys Glu Gln Phe Leu Val Pro Asp His Thr Ile Lys Asp Ile Ser
115 120 125
Gly Ala Ser Phe Ala Gly Phe Tyr Tyr Ile Cys Phe Gln Lys Ser Ala
130 135 140
Ala Ser Ile Glu Gly Tyr Tyr Tyr His Arg Ser Ser Glu Trp Tyr Gln
145 150 155 160
Ser Leu Asn Leu Thr His Val
165
<210> 20
<211> 362
<212> PRT
<213> 酿酒酵母
<400> 20
Met Ile Asn Asn Pro Lys Val Asp Ser Val Ala Glu Lys Pro Lys Ala
1 5 10 15
Val Thr Ser Lys Gln Ser Glu Gln Ala Ala Ser Pro Glu Pro Thr Pro
20 25 30
Ala Pro Pro Val Ser Arg Asn Gln Tyr Pro Ile Thr Phe Asn Leu Thr
35 40 45
Ser Thr Ala Pro Phe His Leu His Asp Arg His Arg Tyr Leu Gln Glu
50 55 60
Gln Asp Leu Tyr Lys Cys Ala Ser Arg Asp Ser Leu Ser Ser Leu Gln
65 70 75 80
Gln Leu Ala His Thr Pro Asn Gly Ser Thr Arg Lys Lys Tyr Ile Val
85 90 95
Glu Asp Gln Ser Pro Tyr Ser Ser Glu Asn Pro Val Ile Val Thr Ser
100 105 110
Ser Tyr Asn His Thr Val Cys Thr Asn Tyr Leu Arg Pro Arg Met Gln
115 120 125
Phe Thr Gly Tyr Gln Ile Ser Gly Tyr Lys Arg Tyr Gln Val Thr Val
130 135 140
Asn Leu Lys Thr Val Asp Leu Pro Lys Lys Asp Cys Thr Ser Leu Ser
145 150 155 160
Pro His Leu Ser Gly Phe Leu Ser Ile Arg Gly Leu Thr Asn Gln His
165 170 175
Pro Glu Ile Ser Thr Tyr Phe Glu Ala Tyr Ala Val Asn His Lys Glu
180 185 190
Leu Gly Phe Leu Ser Ser Ser Trp Lys Asp Glu Pro Val Leu Asn Glu
195 200 205
Phe Lys Ala Thr Asp Gln Thr Asp Leu Glu His Trp Ile Asn Phe Pro
210 215 220
Ser Phe Arg Gln Leu Phe Leu Met Ser Gln Lys Asn Gly Leu Asn Ser
225 230 235 240
Thr Asp Asp Asn Gly Thr Thr Asn Ala Ala Lys Lys Leu Pro Pro Gln
245 250 255
Gln Leu Pro Thr Thr Pro Ser Ala Asp Ala Gly Asn Ile Ser Arg Ile
260 265 270
Phe Ser Gln Glu Lys Gln Phe Asp Asn Tyr Leu Asn Glu Arg Phe Ile
275 280 285
Phe Met Lys Trp Lys Glu Lys Phe Leu Val Pro Asp Ala Leu Leu Met
290 295 300
Glu Gly Val Asp Gly Ala Ser Tyr Asp Gly Phe Tyr Tyr Ile Val His
305 310 315 320
Asp Gln Val Thr Gly Asn Ile Gln Gly Phe Tyr Tyr His Gln Asp Ala
325 330 335
Glu Lys Phe Gln Gln Leu Glu Leu Val Pro Ser Leu Lys Asn Lys Val
340 345 350
Glu Ser Ser Asp Cys Ser Phe Glu Phe Ala
355 360
<210> 21
<211> 240
<212> PRT
<213> 未知
<220>
<223> 针对磷酸化酪氨酸的单链抗体可变片段(scFv)
<400> 21
Met Met Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro
1 5 10 15
Gly Ala Ser Val Met Ile Ser Cys Arg Thr Ser Ala Tyr Thr Phe Thr
20 25 30
Glu Asn Thr Val His Trp Val Lys Gln Ser His Gly Glu Ser Leu Glu
35 40 45
Trp Ile Gly Gly Ile Asn Pro Tyr Tyr Gly Gly Ser Ile Phe Ser Pro
50 55 60
Lys Phe Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr
65 70 75 80
Ala Tyr Met Glu Leu Arg Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Arg Ala Gly Ala Tyr Tyr Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Ser Gly Gly Gly Ser
115 120 125
Gly Gly Gly Ser Glu Asn Val Leu Thr Gln Ser Pro Ala Ile Met Ser
130 135 140
Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser
145 150 155 160
Val Ser Ser Ser Tyr Leu His Trp Tyr Arg Gln Lys Ser Gly Ala Ser
165 170 175
Pro Lys Leu Trp Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro
180 185 190
Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile
195 200 205
Ser Ser Val Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Tyr
210 215 220
Ser Gly Tyr Arg Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg
225 230 235 240
<210> 22
<211> 111
<212> PRT
<213> 智人
<400> 22
Met Gly Ala Met Asp Ser Ile Gln Ala Glu Glu Trp Tyr Phe Gly Lys
1 5 10 15
Leu Gly Arg Lys Asp Ala Glu Arg Gln Leu Leu Ser Phe Gly Asn Pro
20 25 30
Arg Gly Thr Phe Leu Ile Arg Glu Ser Glu Thr Thr Lys Gly Ala Tyr
35 40 45
Ser Leu Ser Ile Arg Asp Trp Asp Asp Met Lys Gly Asp His Val Lys
50 55 60
His Tyr Lys Ile Arg Lys Leu Asp Asn Gly Gly Tyr Tyr Ile Thr Thr
65 70 75 80
Arg Ala Gln Phe Glu Thr Leu Gln Gln Leu Val Gln His Tyr Ser Glu
85 90 95
Arg Ala Ala Gly Leu Ser Ser Arg Leu Val Val Pro Ser His Lys
100 105 110
<210> 23
<211> 111
<212> PRT
<213> 智人
<400> 23
Met Gly Ala Met Asp Ser Ile Gln Ala Glu Glu Trp Tyr Phe Gly Lys
1 5 10 15
Leu Gly Arg Lys Asp Ala Glu Arg Gln Leu Leu Ser Phe Gly Asn Pro
20 25 30
Arg Gly Thr Phe Leu Ile Arg Glu Ser Glu Thr Val Lys Gly Ala Tyr
35 40 45
Ala Leu Ser Ile Arg Asp Trp Asp Asp Met Lys Gly Asp His Val Lys
50 55 60
His Tyr Leu Ile Arg Lys Leu Asp Asn Gly Gly Tyr Tyr Ile Thr Thr
65 70 75 80
Arg Ala Gln Phe Glu Thr Leu Gln Gln Leu Val Gln His Tyr Ser Glu
85 90 95
Arg Ala Ala Gly Leu Ser Ser Arg Leu Val Val Pro Ser His Lys
100 105 110
<210> 24
<211> 113
<212> PRT
<213> 智人
<400> 24
Met Gly Ala Met Asp Ser Ile Gln Ala Glu Glu Trp Tyr Phe Gly Lys
1 5 10 15
Ile Thr Arg Arg Glu Ser Glu Arg Leu Leu Leu Asn Ala Glu Asn Pro
20 25 30
Arg Gly Thr Phe Leu Val Arg Glu Ser Glu Thr Thr Lys Gly Ala Tyr
35 40 45
Ser Leu Ser Val Ser Asp Phe Asp Asn Ala Lys Gly Leu Asn Val Lys
50 55 60
His Tyr Lys Ile Arg Lys Leu Asp Ser Gly Gly Phe Tyr Ile Thr Ser
65 70 75 80
Arg Thr Gln Phe Asn Ser Leu Gln Gln Leu Val Ala Tyr Tyr Ser Lys
85 90 95
His Ala Asp Gly Leu Cys His Arg Leu Thr Thr Val Cys Pro Thr Ser
100 105 110
Lys
<210> 25
<211> 113
<212> PRT
<213> 智人
<400> 25
Met Gly Ala Met Asp Ser Ile Gln Ala Glu Glu Trp Tyr Phe Gly Lys
1 5 10 15
Ile Thr Arg Arg Glu Ser Glu Arg Leu Leu Leu Asn Ala Glu Asn Pro
20 25 30
Arg Gly Thr Phe Leu Val Arg Glu Ser Glu Val Thr Lys Gly Ala Tyr
35 40 45
Ala Leu Ser Val Ser Asp Phe Asp Asn Ala Lys Gly Leu Asn Val Lys
50 55 60
His Tyr Leu Ile Arg Lys Leu Asp Ser Gly Gly Phe Tyr Ile Thr Ser
65 70 75 80
Arg Thr Gln Phe Asn Ser Leu Gln Gln Leu Val Ala Tyr Tyr Ser Lys
85 90 95
His Ala Asp Gly Leu Cys His Arg Leu Thr Thr Val Cys Pro Thr Ser
100 105 110
Lys
<210> 26
<211> 310
<212> PRT
<213> 智人
<400> 26
Met Ala Ser Leu Thr Val Lys Ala Tyr Leu Leu Gly Lys Glu Asp Ala
1 5 10 15
Ala Arg Glu Ile Arg Arg Phe Ser Phe Cys Cys Ser Pro Glu Pro Glu
20 25 30
Ala Glu Ala Glu Ala Ala Ala Gly Pro Gly Pro Cys Glu Arg Leu Leu
35 40 45
Ser Arg Val Ala Ala Leu Phe Pro Ala Leu Arg Pro Gly Gly Phe Gln
50 55 60
Ala His Tyr Arg Asp Glu Asp Gly Asp Leu Val Ala Phe Ser Ser Asp
65 70 75 80
Glu Glu Leu Thr Met Ala Met Ser Tyr Val Lys Asp Asp Ile Phe Arg
85 90 95
Ile Tyr Ile Lys Glu Lys Lys Glu Cys Arg Arg Asp His Arg Pro Pro
100 105 110
Cys Ala Gln Glu Ala Pro Arg Asn Met Val His Pro Asn Val Ile Cys
115 120 125
Asp Gly Cys Asn Gly Pro Val Val Gly Thr Arg Tyr Lys Cys Ser Val
130 135 140
Cys Pro Asp Tyr Asp Leu Cys Ser Val Cys Glu Gly Lys Gly Leu His
145 150 155 160
Arg Gly His Thr Lys Leu Ala Phe Pro Ser Pro Phe Gly His Leu Ser
165 170 175
Glu Gly Phe Ser His Ser Arg Trp Leu Arg Lys Val Lys His Gly His
180 185 190
Phe Gly Trp Pro Gly Trp Glu Met Gly Pro Pro Gly Asn Trp Ser Pro
195 200 205
Arg Pro Pro Arg Ala Gly Glu Ala Arg Pro Gly Pro Thr Ala Glu Ser
210 215 220
Ala Ser Gly Pro Ser Glu Asp Pro Ser Val Asn Phe Leu Lys Asn Val
225 230 235 240
Gly Glu Ser Val Ala Ala Ala Leu Ser Pro Leu Gly Ile Glu Val Asp
245 250 255
Ile Asp Val Glu His Gly Gly Lys Arg Ser Arg Leu Thr Pro Val Ser
260 265 270
Pro Glu Ser Ser Ser Thr Glu Glu Lys Ser Ser Ser Gln Pro Ser Ser
275 280 285
Cys Cys Ser Asp Pro Ser Lys Pro Gly Gly Asn Val Glu Gly Ala Thr
290 295 300
Gln Ser Leu Ala Glu Gln
305 310
<210> 27
<211> 310
<212> PRT
<213> 智人
<400> 27
Met Ala Ser Leu Thr Val Lys Ala Tyr Leu Leu Gly Lys Glu Asp Ala
1 5 10 15
Ala Arg Glu Ile Arg Arg Phe Ser Phe Cys Cys Ser Pro Glu Pro Glu
20 25 30
Ala Glu Ala Glu Ala Ala Ala Gly Pro Gly Pro Cys Glu Arg Leu Leu
35 40 45
Ser Arg Val Ala Ala Leu Phe Pro Ala Leu Arg Pro Gly Gly Phe Gln
50 55 60
Ala His Tyr Arg Asp Glu Asp Gly Asp Leu Val Ala Phe Ser Ser Asp
65 70 75 80
Glu Glu Leu Thr Met Ala Met Ser Tyr Val Lys Asp Asp Ile Phe Arg
85 90 95
Ile Tyr Ile Lys Glu Lys Lys Glu Cys Arg Arg Asp His Arg Pro Pro
100 105 110
Cys Ala Gln Glu Ala Pro Arg Asn Met Val His Pro Asn Val Ile Cys
115 120 125
Asp Gly Cys Asn Gly Pro Val Val Gly Thr Arg Tyr Lys Cys Ser Val
130 135 140
Cys Pro Asp Tyr Asp Leu Cys Ser Val Cys Glu Gly Lys Gly Leu His
145 150 155 160
Arg Gly His Thr Lys Leu Ala Phe Pro Ser Pro Phe Gly His Leu Ser
165 170 175
Glu Gly Phe Ser His Ser Arg Trp Leu Arg Lys Val Lys His Gly His
180 185 190
Phe Gly Trp Pro Gly Trp Glu Met Gly Pro Pro Gly Asn Trp Ser Pro
195 200 205
Arg Pro Pro Arg Ala Gly Glu Ala Arg Pro Gly Pro Thr Ala Glu Ser
210 215 220
Ala Ser Gly Pro Ser Glu Asp Pro Ser Val Asn Phe Leu Lys Asn Val
225 230 235 240
Gly Glu Ser Val Ala Ala Ala Leu Ser Pro Leu Gly Ile Glu Val Asp
245 250 255
Ile Asp Val Glu His Gly Gly Lys Arg Ser Arg Leu Thr Pro Val Ser
260 265 270
Pro Glu Ser Ser Ser Thr Glu Glu Lys Ser Ser Ser Gln Pro Ser Ser
275 280 285
Cys Cys Ser Asp Pro Ser Lys Pro Gly Gly Asn Val Glu Gly Ala Thr
290 295 300
Gln Ser Leu Ala Glu Gln
305 310
<210> 28
<211> 310
<212> PRT
<213> 智人
<400> 28
Met Ala Ser Leu Thr Val Lys Ala Tyr Leu Leu Gly Lys Glu Asp Ala
1 5 10 15
Ala Arg Glu Ile Arg Arg Phe Ser Phe Cys Cys Ser Pro Glu Pro Glu
20 25 30
Ala Glu Ala Glu Ala Ala Ala Gly Pro Gly Pro Cys Glu Arg Leu Leu
35 40 45
Ser Arg Val Ala Ala Leu Phe Pro Ala Leu Arg Pro Gly Gly Phe Gln
50 55 60
Ala His Tyr Arg Asp Glu Asp Gly Asp Leu Val Ala Phe Ser Ser Asp
65 70 75 80
Glu Glu Leu Thr Met Ala Met Ser Tyr Val Lys Asp Asp Ile Phe Arg
85 90 95
Ile Tyr Ile Lys Glu Lys Lys Glu Cys Arg Arg Asp His Arg Pro Pro
100 105 110
Cys Ala Gln Glu Ala Pro Arg Asn Met Val His Pro Asn Val Ile Cys
115 120 125
Asp Gly Cys Asn Gly Pro Val Val Gly Thr Arg Tyr Lys Cys Ser Val
130 135 140
Cys Pro Asp Tyr Asp Leu Cys Ser Val Cys Glu Gly Lys Gly Leu His
145 150 155 160
Arg Gly His Thr Lys Leu Ala Phe Pro Ser Pro Phe Gly His Leu Ser
165 170 175
Glu Gly Phe Ser His Ser Arg Trp Leu Arg Lys Val Lys His Gly His
180 185 190
Phe Gly Trp Pro Gly Trp Glu Met Gly Pro Pro Gly Asn Trp Ser Pro
195 200 205
Arg Pro Pro Arg Ala Gly Glu Ala Arg Pro Gly Pro Thr Ala Glu Ser
210 215 220
Ala Ser Gly Pro Ser Glu Asp Pro Ser Val Asn Phe Leu Lys Asn Val
225 230 235 240
Gly Glu Ser Val Ala Ala Ala Leu Ser Pro Leu Gly Ile Glu Val Asp
245 250 255
Ile Asp Val Glu His Gly Gly Lys Arg Ser Arg Leu Thr Pro Val Ser
260 265 270
Pro Glu Ser Ser Ser Thr Glu Glu Lys Ser Ser Ser Gln Pro Ser Ser
275 280 285
Cys Cys Ser Asp Pro Ser Lys Pro Gly Gly Asn Val Glu Gly Ala Thr
290 295 300
Gln Ser Leu Ala Glu Gln
305 310
<210> 29
<211> 440
<212> PRT
<213> 智人
<400> 29
Met Ala Ser Leu Thr Val Lys Ala Tyr Leu Leu Gly Lys Glu Asp Ala
1 5 10 15
Ala Arg Glu Ile Arg Arg Phe Ser Phe Cys Cys Ser Pro Glu Pro Glu
20 25 30
Ala Glu Ala Glu Ala Ala Ala Gly Pro Gly Pro Cys Glu Arg Leu Leu
35 40 45
Ser Arg Val Ala Ala Leu Phe Pro Ala Leu Arg Pro Gly Gly Phe Gln
50 55 60
Ala His Tyr Arg Asp Glu Asp Gly Asp Leu Val Ala Phe Ser Ser Asp
65 70 75 80
Glu Glu Leu Thr Met Ala Met Ser Tyr Val Lys Asp Asp Ile Phe Arg
85 90 95
Ile Tyr Ile Lys Glu Lys Lys Glu Cys Arg Arg Asp His Arg Pro Pro
100 105 110
Cys Ala Gln Glu Ala Pro Arg Asn Met Val His Pro Asn Val Ile Cys
115 120 125
Asp Gly Cys Asn Gly Pro Val Val Gly Thr Arg Tyr Lys Cys Ser Val
130 135 140
Cys Pro Asp Tyr Asp Leu Cys Ser Val Cys Glu Gly Lys Gly Leu His
145 150 155 160
Arg Gly His Thr Lys Leu Ala Phe Pro Ser Pro Phe Gly His Leu Ser
165 170 175
Glu Gly Phe Ser His Ser Arg Trp Leu Arg Lys Val Lys His Gly His
180 185 190
Phe Gly Trp Pro Gly Trp Glu Met Gly Pro Pro Gly Asn Trp Ser Pro
195 200 205
Arg Pro Pro Arg Ala Gly Glu Ala Arg Pro Gly Pro Thr Ala Glu Ser
210 215 220
Ala Ser Gly Pro Ser Glu Asp Pro Ser Val Asn Phe Leu Lys Asn Val
225 230 235 240
Gly Glu Ser Val Ala Ala Ala Leu Ser Pro Leu Gly Ile Glu Val Asp
245 250 255
Ile Asp Val Glu His Gly Gly Lys Arg Ser Arg Leu Thr Pro Val Ser
260 265 270
Pro Glu Ser Ser Ser Thr Glu Glu Lys Ser Ser Ser Gln Pro Ser Ser
275 280 285
Cys Cys Ser Asp Pro Ser Lys Pro Gly Gly Asn Val Glu Gly Ala Thr
290 295 300
Gln Ser Leu Ala Glu Gln Met Arg Lys Ile Ala Leu Glu Ser Glu Gly
305 310 315 320
Arg Pro Glu Glu Gln Met Glu Ser Asp Asn Cys Ser Gly Gly Asp Asp
325 330 335
Asp Trp Thr His Leu Ser Ser Lys Glu Val Asp Pro Ser Thr Gly Glu
340 345 350
Leu Gln Ser Leu Gln Met Pro Glu Ser Glu Gly Pro Ser Ser Leu Asp
355 360 365
Pro Ser Gln Glu Gly Pro Thr Gly Leu Lys Glu Ala Ala Leu Tyr Pro
370 375 380
His Leu Pro Pro Glu Ala Asp Pro Arg Leu Ile Glu Ser Leu Ser Gln
385 390 395 400
Met Leu Ser Met Gly Phe Ser Asp Glu Gly Gly Trp Leu Thr Arg Leu
405 410 415
Leu Gln Thr Lys Asn Tyr Asp Ile Gly Ala Ala Leu Asp Thr Ile Gln
420 425 430
Tyr Ser Lys His Pro Pro Pro Leu
435 440
<210> 30
<211> 439
<212> PRT
<213> 褐家鼠
<400> 30
Met Ala Ser Leu Thr Val Lys Ala Tyr Leu Leu Gly Lys Glu Glu Ala
1 5 10 15
Ala Arg Glu Ile Arg Arg Phe Ser Phe Cys Phe Ser Pro Glu Pro Glu
20 25 30
Ala Glu Ala Ala Ala Gly Pro Gly Pro Cys Glu Arg Leu Leu Ser Arg
35 40 45
Val Ala Val Leu Phe Pro Ala Leu Arg Pro Gly Gly Phe Gln Ala His
50 55 60
Tyr Arg Asp Glu Asp Gly Asp Leu Val Ala Phe Ser Ser Asp Glu Glu
65 70 75 80
Leu Thr Met Ala Met Ser Tyr Val Lys Asp Asp Ile Phe Arg Ile Tyr
85 90 95
Ile Lys Glu Lys Lys Glu Cys Arg Arg Glu His Arg Pro Pro Cys Ala
100 105 110
Gln Glu Ala Arg Ser Met Val His Pro Asn Val Ile Cys Asp Gly Cys
115 120 125
Asn Gly Pro Val Val Gly Thr Arg Tyr Lys Cys Ser Val Cys Pro Asp
130 135 140
Tyr Asp Leu Cys Ser Val Cys Glu Gly Lys Gly Leu His Arg Glu His
145 150 155 160
Ser Lys Leu Ile Phe Pro Asn Pro Phe Gly His Leu Ser Asp Ser Phe
165 170 175
Ser His Ser Arg Trp Leu Arg Lys Leu Lys His Gly His Phe Gly Trp
180 185 190
Pro Gly Trp Glu Met Gly Pro Pro Gly Asn Trp Ser Pro Arg Pro Pro
195 200 205
Arg Ala Gly Asp Gly Arg Pro Cys Pro Thr Ala Glu Ser Ala Ser Ala
210 215 220
Pro Ser Glu Asp Pro Asn Val Asn Phe Leu Lys Asn Val Gly Glu Ser
225 230 235 240
Val Ala Ala Ala Leu Ser Pro Leu Gly Ile Glu Val Asp Ile Asp Val
245 250 255
Glu His Gly Gly Lys Arg Ser Arg Leu Thr Pro Thr Ser Ala Glu Ser
260 265 270
Ser Ser Thr Gly Thr Glu Asp Lys Ser Gly Thr Gln Pro Ser Ser Cys
275 280 285
Ser Ser Glu Val Ser Lys Pro Asp Gly Ala Gly Glu Gly Pro Ala Gln
290 295 300
Ser Leu Thr Glu Gln Met Lys Lys Ile Ala Leu Glu Ser Val Gly Gln
305 310 315 320
Pro Glu Glu Leu Met Glu Ser Asp Asn Cys Ser Gly Gly Asp Asp Asp
325 330 335
Trp Thr His Leu Ser Ser Lys Glu Val Asp Pro Ser Thr Gly Glu Leu
340 345 350
Gln Ser Leu Gln Met Pro Glu Ser Glu Gly Pro Ser Ser Leu Asp Pro
355 360 365
Ser Gln Glu Gly Pro Thr Gly Leu Lys Glu Ala Ala Leu Tyr Pro His
370 375 380
Leu Pro Pro Glu Ala Asp Pro Arg Leu Ile Glu Ser Leu Ser Gln Met
385 390 395 400
Leu Ser Met Gly Phe Ser Asp Glu Gly Gly Trp Leu Thr Arg Leu Leu
405 410 415
Gln Thr Lys Asn Tyr Asp Ile Gly Ala Ala Leu Asp Thr Ile Gln Tyr
420 425 430
Ser Lys His Pro Pro Pro Leu
435
<210> 31
<211> 292
<212> PRT
<213> 酿酒酵母
<400> 31
Met Thr Ser Leu Asn Ile Met Gly Arg Lys Phe Ile Leu Glu Arg Ala
1 5 10 15
Lys Arg Asn Asp Asn Ile Glu Glu Ile Tyr Thr Ser Ala Tyr Val Ser
20 25 30
Leu Pro Ser Ser Thr Asp Thr Arg Leu Pro His Phe Lys Ala Lys Glu
35 40 45
Glu Asp Cys Asp Val Tyr Glu Glu Gly Thr Asn Leu Val Gly Lys Asn
50 55 60
Ala Lys Tyr Thr Tyr Arg Ser Leu Gly Arg His Leu Asp Phe Leu Arg
65 70 75 80
Pro Gly Leu Arg Phe Gly Gly Ser Gln Ser Ser Lys Tyr Thr Tyr Tyr
85 90 95
Thr Val Glu Val Lys Ile Asp Thr Val Asn Leu Pro Leu Tyr Lys Asp
100 105 110
Ser Arg Ser Leu Asp Pro His Val Thr Gly Thr Phe Thr Ile Lys Asn
115 120 125
Leu Thr Pro Val Leu Asp Lys Val Val Thr Leu Phe Glu Gly Tyr Val
130 135 140
Ile Asn Tyr Asn Gln Phe Pro Leu Cys Ser Leu His Trp Pro Ala Glu
145 150 155 160
Glu Thr Leu Asp Pro Tyr Met Ala Gln Arg Glu Ser Asp Cys Ser His
165 170 175
Trp Lys Arg Phe Gly His Phe Gly Ser Asp Asn Trp Ser Leu Thr Glu
180 185 190
Arg Asn Phe Gly Gln Tyr Asn His Glu Ser Ala Glu Phe Met Asn Gln
195 200 205
Arg Tyr Ile Tyr Leu Lys Trp Lys Glu Arg Phe Leu Leu Asp Asp Glu
210 215 220
Glu Gln Glu Asn Gln Met Leu Asp Asp Asn His His Leu Glu Gly Ala
225 230 235 240
Ser Phe Glu Gly Phe Tyr Tyr Val Cys Leu Asp Gln Leu Thr Gly Ser
245 250 255
Val Glu Gly Tyr Tyr Tyr His Pro Ala Cys Glu Leu Phe Gln Lys Leu
260 265 270
Glu Leu Val Pro Thr Asn Cys Asp Ala Leu Asn Thr Tyr Ser Ser Gly
275 280 285
Phe Glu Ile Ala
290
<210> 32
<211> 77
<212> PRT
<213> 智人
<400> 32
Met Gly Pro Leu Gly Ser Leu Cys Gly Arg Val Phe Lys Ser Gly Glu
1 5 10 15
Thr Thr Tyr Ser Cys Arg Asp Cys Ala Ile Asp Pro Thr Cys Val Leu
20 25 30
Cys Met Asp Cys Phe Gln Asp Ser Val His Lys Asn His Arg Tyr Lys
35 40 45
Met His Thr Ser Thr Gly Gly Gly Phe Cys Asp Cys Gly Asp Thr Glu
50 55 60
Ala Trp Lys Thr Gly Pro Phe Cys Val Asn His Glu Pro
65 70 75
<210> 33
<211> 76
<212> PRT
<213> 智人
<400> 33
Met Gly Pro Leu Gly Ser Leu Cys Gly Arg Val Phe Lys Val Gly Glu
1 5 10 15
Pro Thr Tyr Ser Cys Arg Asp Cys Ala Val Asp Pro Thr Cys Val Leu
20 25 30
Cys Met Glu Cys Phe Leu Gly Ser Ile His Arg Asp His Arg Tyr Arg
35 40 45
Met Thr Thr Ser Gly Gly Gly Gly Phe Cys Asp Cys Gly Asp Thr Glu
50 55 60
Ala Trp Lys Glu Gly Pro Tyr Cys Gln Lys His Glu
65 70 75
<210> 34
<211> 421
<212> PRT
<213> 硕大利什曼原虫
<400> 34
Met Ser Arg Asn Pro Ser Asn Ser Asp Ala Ala His Ala Phe Trp Ser
1 5 10 15
Thr Gln Pro Val Pro Gln Thr Glu Asp Glu Thr Glu Lys Ile Val Phe
20 25 30
Ala Gly Pro Met Asp Glu Pro Lys Thr Val Ala Asp Ile Pro Glu Glu
35 40 45
Pro Tyr Pro Ile Ala Ser Thr Phe Glu Trp Trp Thr Pro Asn Met Glu
50 55 60
Ala Ala Asp Asp Ile His Ala Ile Tyr Glu Leu Leu Arg Asp Asn Tyr
65 70 75 80
Val Glu Asp Asp Asp Ser Met Phe Arg Phe Asn Tyr Ser Glu Glu Phe
85 90 95
Leu Gln Trp Ala Leu Cys Pro Pro Asn Tyr Ile Pro Asp Trp His Val
100 105 110
Ala Val Arg Arg Lys Ala Asp Lys Lys Leu Leu Ala Phe Ile Ala Gly
115 120 125
Val Pro Val Thr Leu Arg Met Gly Thr Pro Lys Tyr Met Lys Val Lys
130 135 140
Ala Gln Glu Lys Gly Glu Gly Glu Glu Ala Ala Lys Tyr Asp Glu Pro
145 150 155 160
Arg His Ile Cys Glu Ile Asn Phe Leu Cys Val His Lys Gln Leu Arg
165 170 175
Glu Lys Arg Leu Ala Pro Ile Leu Ile Lys Glu Ala Thr Arg Arg Val
180 185 190
Asn Arg Thr Asn Val Trp Gln Ala Val Tyr Thr Ala Gly Val Leu Leu
195 200 205
Pro Thr Pro Tyr Ala Ser Gly Gln Tyr Phe His Arg Ser Leu Asn Pro
210 215 220
Glu Lys Leu Val Glu Ile Arg Phe Ser Gly Ile Pro Ala Gln Tyr Gln
225 230 235 240
Lys Phe Gln Asn Pro Met Ala Met Leu Lys Arg Asn Tyr Gln Leu Pro
245 250 255
Ser Ala Pro Lys Asn Ser Gly Leu Arg Glu Met Lys Pro Ser Asp Val
260 265 270
Pro Gln Val Arg Arg Ile Leu Met Asn Tyr Leu Asp Ser Phe Asp Val
275 280 285
Gly Pro Val Phe Ser Asp Ala Glu Ile Ser His Tyr Leu Leu Pro Arg
290 295 300
Asp Gly Val Val Phe Thr Tyr Val Val Glu Asn Asp Lys Lys Val Thr
305 310 315 320
Asp Phe Phe Ser Phe Tyr Arg Ile Pro Ser Thr Val Ile Gly Asn Ser
325 330 335
Asn Tyr Asn Leu Leu Asn Ala Ala Tyr Val His Tyr Tyr Ala Ala Thr
340 345 350
Ser Ile Pro Leu His Gln Leu Ile Leu Asp Leu Leu Ile Val Ala His
355 360 365
Ser Arg Gly Phe Asp Val Cys Asn Met Val Glu Ile Leu Asp Asn Arg
370 375 380
Ser Phe Val Glu Gln Leu Lys Phe Gly Ala Gly Asp Gly His Leu Arg
385 390 395 400
Tyr Tyr Phe Tyr Asn Trp Ala Tyr Pro Lys Ile Lys Pro Ser Gln Val
405 410 415
Ala Leu Val Met Leu
420
<210> 35
<211> 496
<212> PRT
<213> 智人
<400> 35
Met Ala Asp Glu Ser Glu Thr Ala Val Lys Pro Pro Ala Pro Pro Leu
1 5 10 15
Pro Gln Met Met Glu Gly Asn Gly Asn Gly His Glu His Cys Ser Asp
20 25 30
Cys Glu Asn Glu Glu Asp Asn Ser Tyr Asn Arg Gly Gly Leu Ser Pro
35 40 45
Ala Asn Asp Thr Gly Ala Lys Lys Lys Lys Lys Lys Gln Lys Lys Lys
50 55 60
Lys Glu Lys Gly Ser Glu Thr Asp Ser Ala Gln Asp Gln Pro Val Lys
65 70 75 80
Met Asn Ser Leu Pro Ala Glu Arg Ile Gln Glu Ile Gln Lys Ala Ile
85 90 95
Glu Leu Phe Ser Val Gly Gln Gly Pro Ala Lys Thr Met Glu Glu Ala
100 105 110
Ser Lys Arg Ser Tyr Gln Phe Trp Asp Thr Gln Pro Val Pro Lys Leu
115 120 125
Gly Glu Val Val Asn Thr His Gly Pro Val Glu Pro Asp Lys Asp Asn
130 135 140
Ile Arg Gln Glu Pro Tyr Thr Leu Pro Gln Gly Phe Thr Trp Asp Ala
145 150 155 160
Leu Asp Leu Gly Asp Arg Gly Val Leu Lys Glu Leu Tyr Thr Leu Leu
165 170 175
Asn Glu Asn Tyr Val Glu Asp Asp Asp Asn Met Phe Arg Phe Asp Tyr
180 185 190
Ser Pro Glu Phe Leu Leu Trp Ala Leu Arg Pro Pro Gly Trp Leu Pro
195 200 205
Gln Trp His Cys Gly Val Arg Val Val Ser Ser Arg Lys Leu Val Gly
210 215 220
Phe Ile Ser Ala Ile Pro Ala Asn Ile His Ile Tyr Asp Thr Glu Lys
225 230 235 240
Lys Met Val Glu Ile Asn Phe Leu Cys Val His Lys Lys Leu Arg Ser
245 250 255
Lys Arg Val Ala Pro Val Leu Ile Arg Glu Ile Thr Arg Arg Val His
260 265 270
Leu Glu Gly Ile Phe Gln Ala Val Tyr Thr Ala Gly Val Val Leu Pro
275 280 285
Lys Pro Val Gly Thr Cys Arg Tyr Trp His Arg Ser Leu Asn Pro Arg
290 295 300
Lys Leu Ile Glu Val Lys Phe Ser His Leu Ser Arg Asn Met Thr Met
305 310 315 320
Gln Arg Thr Met Lys Leu Tyr Arg Leu Pro Glu Thr Pro Lys Thr Ala
325 330 335
Gly Leu Arg Pro Met Glu Thr Lys Asp Ile Pro Val Val His Gln Leu
340 345 350
Leu Thr Arg Tyr Leu Lys Gln Phe His Leu Thr Pro Val Met Ser Gln
355 360 365
Glu Glu Val Glu His Trp Phe Tyr Pro Gln Glu Asn Ile Ile Asp Thr
370 375 380
Phe Val Val Glu Asn Ala Asn Gly Glu Val Thr Asp Phe Leu Ser Phe
385 390 395 400
Tyr Thr Leu Pro Ser Thr Ile Met Asn His Pro Thr His Lys Ser Leu
405 410 415
Lys Ala Ala Tyr Ser Phe Tyr Asn Val His Thr Gln Thr Pro Leu Leu
420 425 430
Asp Leu Met Ser Asp Ala Leu Val Leu Ala Lys Met Lys Gly Phe Asp
435 440 445
Val Phe Asn Ala Leu Asp Leu Met Glu Asn Lys Thr Phe Leu Glu Lys
450 455 460
Leu Lys Phe Gly Ile Gly Asp Gly Asn Leu Gln Tyr Tyr Leu Tyr Asn
465 470 475 480
Trp Lys Cys Pro Ser Met Gly Ala Glu Lys Val Gly Leu Val Leu Gln
485 490 495
<210> 36
<211> 127
<212> PRT
<213> 黑腹果蝇
<400> 36
Met Gly Asp Val Gln Pro Glu Thr Cys Arg Pro Ser Ala Ala Ser Gly
1 5 10 15
Asn Tyr Phe Pro Gln Tyr Pro Glu Tyr Ala Ile Glu Thr Ala Arg Leu
20 25 30
Arg Thr Phe Glu Ala Trp Pro Arg Asn Leu Lys Gln Lys Pro His Gln
35 40 45
Leu Ala Glu Ala Gly Phe Phe Tyr Thr Gly Val Gly Asp Arg Val Arg
50 55 60
Cys Phe Ser Cys Gly Gly Gly Leu Met Asp Trp Asn Asp Asn Asp Glu
65 70 75 80
Pro Trp Glu Gln His Ala Leu Trp Leu Ser Gln Cys Arg Phe Val Lys
85 90 95
Leu Met Lys Gly Gln Leu Tyr Ile Asp Thr Val Ala Ala Lys Pro Val
100 105 110
Leu Ala Glu Glu Lys Glu Glu Ser Thr Ser Ile Gly Gly Asp Thr
115 120 125
<210> 37
<211> 27
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 37
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly Leu Asn Asp
1 5 10 15
Phe Phe Glu Ala Gln Lys Ile Glu Trp His Glu
20 25
<210> 38
<211> 54
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 38
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly Leu Asn Asp
1 5 10 15
Phe Phe Glu Ala Gln Lys Ile Glu Trp His Glu Gly Gly Gly Ser Gly
20 25 30
Gly Gly Ser Gly Gly Gly Ser Gly Leu Asn Asp Phe Phe Glu Ala Gln
35 40 45
Lys Ile Glu Trp His Glu
50
<210> 39
<211> 56
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 39
Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly Leu
1 5 10 15
Asn Asp Phe Phe Glu Ala Gln Lys Ile Glu Trp His Glu Gly Gly Gly
20 25 30
Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly Leu Asn Asp Phe Phe Glu
35 40 45
Ala Gln Lys Ile Glu Trp His Glu
50 55
<210> 40
<211> 38
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 40
Gly His His His His His His His His His His Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Ser Gly Gly Gly Ser Gly Leu Asn Asp Phe Phe Glu Ala Gln
20 25 30
Lys Ile Glu Trp His Glu
35
<210> 41
<211> 65
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 41
Gly His His His His His His His His His His Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Ser Gly Gly Gly Ser Gly Leu Asn Asp Phe Phe Glu Ala Gln
20 25 30
Lys Ile Glu Trp His Glu Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly
35 40 45
Gly Ser Gly Leu Asn Asp Phe Phe Glu Ala Gln Lys Ile Glu Trp His
50 55 60
Glu
65
<210> 42
<211> 67
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 42
Gly Gly Ser His His His His His His His His His His Gly Gly Gly
1 5 10 15
Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly Leu Asn Asp Phe Phe Glu
20 25 30
Ala Gln Lys Ile Glu Trp His Glu Gly Gly Gly Ser Gly Gly Gly Ser
35 40 45
Gly Gly Gly Ser Gly Leu Asn Asp Phe Phe Glu Ala Gln Lys Ile Glu
50 55 60
Trp His Glu
65
<210> 43
<211> 66
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 43
Gly Ser His His His His His His His His His His Gly Gly Gly Ser
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Ser Gly Leu Asn Asp Phe Phe Glu Ala
20 25 30
Gln Lys Ile Glu Trp His Glu Gly Gly Gly Ser Gly Gly Gly Ser Gly
35 40 45
Gly Gly Ser Gly Leu Asn Asp Phe Phe Glu Ala Gln Lys Ile Glu Trp
50 55 60
His Glu
65
<210> 44
<211> 61
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 44
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly Leu Asn Asp
1 5 10 15
Phe Phe Glu Ala Gln Lys Ile Glu Trp His Glu Gly Gly Gly Ser Gly
20 25 30
Gly Gly Ser Gly Gly Gly Ser Gly Leu Asn Asp Phe Phe Glu Ala Gln
35 40 45
Lys Ile Glu Trp His Glu Gly His His His His His His
50 55 60
<210> 45
<211> 901
<212> PRT
<213> 嗜肺性退伍军人杆菌
<400> 45
Met Met Val Lys Gln Gly Val Phe Met Lys Thr Asp Gln Ser Lys Val
1 5 10 15
Lys Lys Leu Ser Asp Tyr Lys Ser Leu Asp Tyr Phe Val Ile His Val
20 25 30
Asp Leu Gln Ile Asp Leu Ser Lys Lys Pro Val Glu Ser Lys Ala Arg
35 40 45
Leu Thr Val Val Pro Asn Leu Asn Val Asp Ser His Ser Asn Asp Leu
50 55 60
Val Leu Asp Gly Glu Asn Met Thr Leu Val Ser Leu Gln Met Asn Asp
65 70 75 80
Asn Leu Leu Lys Glu Asn Glu Tyr Glu Leu Thr Lys Asp Ser Leu Ile
85 90 95
Ile Lys Asn Ile Pro Gln Asn Thr Pro Phe Thr Ile Glu Met Thr Ser
100 105 110
Leu Leu Gly Glu Asn Thr Asp Leu Phe Gly Leu Tyr Glu Thr Glu Gly
115 120 125
Val Ala Leu Val Lys Ala Glu Ser Glu Gly Leu Arg Arg Val Phe Tyr
130 135 140
Leu Pro Asp Arg Pro Asp Asn Leu Ala Thr Tyr Lys Thr Thr Ile Ile
145 150 155 160
Ala Asn Gln Glu Asp Tyr Pro Val Leu Leu Ser Asn Gly Val Leu Ile
165 170 175
Glu Lys Lys Glu Leu Pro Leu Gly Leu His Ser Val Thr Trp Leu Asp
180 185 190
Asp Val Pro Lys Pro Ser Tyr Leu Phe Ala Leu Val Ala Gly Asn Leu
195 200 205
Gln Arg Ser Val Thr Tyr Tyr Gln Thr Lys Ser Gly Arg Glu Leu Pro
210 215 220
Ile Glu Phe Tyr Val Pro Pro Ser Ala Thr Ser Lys Cys Asp Phe Ala
225 230 235 240
Lys Glu Val Leu Lys Glu Ala Met Ala Trp Asp Glu Arg Thr Phe Asn
245 250 255
Leu Glu Cys Ala Leu Arg Gln His Met Val Ala Gly Val Asp Lys Tyr
260 265 270
Ala Ser Gly Ala Ser Glu Pro Thr Gly Leu Asn Leu Phe Asn Thr Glu
275 280 285
Asn Leu Phe Ala Ser Pro Glu Thr Lys Thr Asp Leu Gly Ile Leu Arg
290 295 300
Val Leu Glu Val Val Ala His Glu Phe Phe His Tyr Trp Ser Gly Asp
305 310 315 320
Arg Val Thr Ile Arg Asp Trp Phe Asn Leu Pro Leu Lys Glu Gly Leu
325 330 335
Thr Thr Phe Arg Ala Ala Met Phe Arg Glu Glu Leu Phe Gly Thr Asp
340 345 350
Leu Ile Arg Leu Leu Asp Gly Lys Asn Leu Asp Glu Arg Ala Pro Arg
355 360 365
Gln Ser Ala Tyr Thr Ala Val Arg Ser Leu Tyr Thr Ala Ala Ala Tyr
370 375 380
Glu Lys Ser Ala Asp Ile Phe Arg Met Met Met Leu Phe Ile Gly Lys
385 390 395 400
Glu Pro Phe Ile Glu Ala Val Ala Lys Phe Phe Lys Asp Asn Asp Gly
405 410 415
Gly Ala Val Thr Leu Glu Asp Phe Ile Glu Ser Ile Ser Asn Ser Ser
420 425 430
Gly Lys Asp Leu Arg Ser Phe Leu Ser Trp Phe Thr Glu Ser Gly Ile
435 440 445
Pro Glu Leu Ile Val Thr Asp Glu Leu Asn Pro Asp Thr Lys Gln Tyr
450 455 460
Phe Leu Lys Ile Lys Thr Val Asn Gly Arg Asn Arg Pro Ile Pro Ile
465 470 475 480
Leu Met Gly Leu Leu Asp Ser Ser Gly Ala Glu Ile Val Ala Asp Lys
485 490 495
Leu Leu Ile Val Asp Gln Glu Glu Ile Glu Phe Gln Phe Glu Asn Ile
500 505 510
Gln Thr Arg Pro Ile Pro Ser Leu Leu Arg Ser Phe Ser Ala Pro Val
515 520 525
His Met Lys Tyr Glu Tyr Ser Tyr Gln Asp Leu Leu Leu Leu Met Gln
530 535 540
Phe Asp Thr Asn Leu Tyr Asn Arg Cys Glu Ala Ala Lys Gln Leu Ile
545 550 555 560
Ser Ala Leu Ile Asn Asp Phe Cys Ile Gly Lys Lys Ile Glu Leu Ser
565 570 575
Pro Gln Phe Phe Ala Val Tyr Lys Ala Leu Leu Ser Asp Asn Ser Leu
580 585 590
Asn Glu Trp Met Leu Ala Glu Leu Ile Thr Leu Pro Ser Leu Glu Glu
595 600 605
Leu Ile Glu Asn Gln Asp Lys Pro Asp Phe Glu Lys Leu Asn Glu Gly
610 615 620
Arg Gln Leu Ile Gln Asn Ala Leu Ala Asn Glu Leu Lys Thr Asp Phe
625 630 635 640
Tyr Asn Leu Leu Phe Arg Ile Gln Ile Ser Gly Asp Asp Asp Lys Gln
645 650 655
Lys Leu Lys Gly Phe Asp Leu Lys Gln Ala Gly Leu Arg Arg Leu Lys
660 665 670
Ser Val Cys Phe Ser Tyr Leu Leu Asn Val Asp Phe Glu Lys Thr Lys
675 680 685
Glu Lys Leu Ile Leu Gln Phe Glu Asp Ala Leu Gly Lys Asn Met Thr
690 695 700
Glu Thr Ala Leu Ala Leu Ser Met Leu Cys Glu Ile Asn Cys Glu Glu
705 710 715 720
Ala Asp Val Ala Leu Glu Asp Tyr Tyr His Tyr Trp Lys Asn Asp Pro
725 730 735
Gly Ala Val Asn Asn Trp Phe Ser Ile Gln Ala Leu Ala His Ser Pro
740 745 750
Asp Val Ile Glu Arg Val Lys Lys Leu Met Arg His Gly Asp Phe Asp
755 760 765
Leu Ser Asn Pro Asn Lys Val Tyr Ala Leu Leu Gly Ser Phe Ile Lys
770 775 780
Asn Pro Phe Gly Phe His Ser Val Thr Gly Glu Gly Tyr Gln Leu Val
785 790 795 800
Ala Asp Ala Ile Phe Asp Leu Asp Lys Ile Asn Pro Thr Leu Ala Ala
805 810 815
Asn Leu Thr Glu Lys Phe Thr Tyr Trp Asp Lys Tyr Asp Val Asn Arg
820 825 830
Gln Ala Met Met Ile Ser Thr Leu Lys Ile Ile Tyr Ser Asn Ala Thr
835 840 845
Ser Ser Asp Val Arg Thr Met Ala Lys Lys Gly Leu Asp Lys Val Lys
850 855 860
Glu Asp Leu Pro Leu Pro Ile His Leu Thr Phe His Gly Gly Ser Thr
865 870 875 880
Met Gln Asp Arg Thr Ala Gln Leu Ile Ala Asp Gly Asn Lys Glu Asn
885 890 895
Ala Tyr Gln Leu His
900
<210> 46
<211> 265
<212> PRT
<213> 大肠杆菌
<400> 46
Met Gly Thr Ala Ile Ser Ile Lys Thr Pro Glu Asp Ile Glu Lys Met
1 5 10 15
Arg Val Ala Gly Arg Leu Ala Ala Glu Val Leu Glu Met Ile Glu Pro
20 25 30
Tyr Val Lys Pro Gly Val Ser Thr Gly Glu Leu Asp Arg Ile Cys Asn
35 40 45
Asp Tyr Ile Val Asn Glu Gln His Ala Val Ser Ala Cys Leu Gly Tyr
50 55 60
His Gly Tyr Pro Lys Ser Val Cys Ile Ser Ile Asn Glu Val Val Cys
65 70 75 80
His Gly Ile Pro Asp Asp Ala Lys Leu Leu Lys Asp Gly Asp Ile Val
85 90 95
Asn Ile Asp Val Thr Val Ile Lys Asp Gly Phe His Gly Asp Thr Ser
100 105 110
Lys Met Phe Ile Val Gly Lys Pro Thr Ile Met Gly Glu Arg Leu Cys
115 120 125
Arg Ile Thr Gln Glu Ser Leu Tyr Leu Ala Leu Arg Met Val Lys Pro
130 135 140
Gly Ile Asn Leu Arg Glu Ile Gly Ala Ala Ile Gln Lys Phe Val Glu
145 150 155 160
Ala Glu Gly Phe Ser Val Val Arg Glu Tyr Cys Gly His Gly Ile Gly
165 170 175
Arg Gly Phe His Glu Glu Pro Gln Val Leu His Tyr Asp Ser Arg Glu
180 185 190
Thr Asn Val Val Leu Lys Pro Gly Met Thr Phe Thr Ile Glu Pro Met
195 200 205
Val Asn Ala Gly Lys Lys Glu Ile Arg Thr Met Lys Asp Gly Trp Thr
210 215 220
Val Lys Thr Lys Asp Arg Ser Leu Ser Ala Gln Tyr Glu His Thr Ile
225 230 235 240
Val Val Thr Asp Asn Gly Cys Glu Ile Leu Thr Leu Arg Lys Asp Asp
245 250 255
Thr Ile Pro Ala Ile Ile Ser His Asp
260 265
<210> 47
<211> 322
<212> PRT
<213> 耻垢分枝杆菌
<400> 47
Met Gly Thr Leu Glu Ala Asn Thr Asn Gly Pro Gly Ser Met Leu Ser
1 5 10 15
Arg Met Pro Val Ser Ser Arg Thr Val Pro Phe Gly Asp His Glu Thr
20 25 30
Trp Val Gln Val Thr Thr Pro Glu Asn Ala Gln Pro His Ala Leu Pro
35 40 45
Leu Ile Val Leu His Gly Gly Pro Gly Met Ala His Asn Tyr Val Ala
50 55 60
Asn Ile Ala Ala Leu Ala Asp Glu Thr Gly Arg Thr Val Ile His Tyr
65 70 75 80
Asp Gln Val Gly Cys Gly Asn Ser Thr His Leu Pro Asp Ala Pro Ala
85 90 95
Asp Phe Trp Thr Pro Gln Leu Phe Val Asp Glu Phe His Ala Val Cys
100 105 110
Thr Ala Leu Gly Ile Glu Arg Tyr His Val Leu Gly Gln Ser Trp Gly
115 120 125
Gly Met Leu Gly Ala Glu Ile Ala Val Arg Gln Pro Ser Gly Leu Val
130 135 140
Ser Leu Ala Ile Cys Asn Ser Pro Ala Ser Met Arg Leu Trp Ser Glu
145 150 155 160
Ala Ala Gly Asp Leu Arg Ala Gln Leu Pro Ala Glu Thr Arg Ala Ala
165 170 175
Leu Asp Arg His Glu Ala Ala Gly Thr Ile Thr His Pro Asp Tyr Leu
180 185 190
Gln Ala Ala Ala Glu Phe Tyr Arg Arg His Val Cys Arg Val Val Pro
195 200 205
Thr Pro Gln Asp Phe Ala Asp Ser Val Ala Gln Met Glu Ala Glu Pro
210 215 220
Thr Val Tyr His Thr Met Asn Gly Pro Asn Glu Phe His Val Val Gly
225 230 235 240
Thr Leu Gly Asp Trp Ser Val Ile Asp Arg Leu Pro Asp Val Thr Ala
245 250 255
Pro Val Leu Val Ile Ala Gly Glu His Asp Glu Ala Thr Pro Lys Thr
260 265 270
Trp Gln Pro Phe Val Asp His Ile Pro Asp Val Arg Ser His Val Phe
275 280 285
Pro Gly Thr Ser His Cys Thr His Leu Glu Lys Pro Glu Glu Phe Arg
290 295 300
Ala Val Val Ala Gln Phe Leu His Gln His Asp Leu Ala Ala Asp Ala
305 310 315 320
Arg Val
<210> 48
<211> 446
<212> PRT
<213> 鼠疫耶氏杆菌
<400> 48
Met Thr Gln Gln Glu Tyr Gln Asn Arg Arg Gln Ala Leu Leu Ala Lys
1 5 10 15
Met Ala Pro Gly Ser Ala Ala Ile Ile Phe Ala Ala Pro Glu Ala Thr
20 25 30
Arg Ser Ala Asp Ser Glu Tyr Pro Tyr Arg Gln Asn Ser Asp Phe Ser
35 40 45
Tyr Leu Thr Gly Phe Asn Glu Pro Glu Ala Val Leu Ile Leu Val Lys
50 55 60
Ser Asp Glu Thr His Asn His Ser Val Leu Phe Asn Arg Ile Arg Asp
65 70 75 80
Leu Thr Ala Glu Ile Trp Phe Gly Arg Arg Leu Gly Gln Glu Ala Ala
85 90 95
Pro Thr Lys Leu Ala Val Asp Arg Ala Leu Pro Phe Asp Glu Ile Asn
100 105 110
Glu Gln Leu Tyr Leu Leu Leu Asn Arg Leu Asp Val Ile Tyr His Ala
115 120 125
Gln Gly Gln Tyr Ala Tyr Ala Asp Asn Ile Val Phe Ala Ala Leu Glu
130 135 140
Lys Leu Arg His Gly Phe Arg Lys Asn Leu Arg Ala Pro Ala Thr Leu
145 150 155 160
Thr Asp Trp Arg Pro Trp Leu His Glu Met Arg Leu Phe Lys Ser Ala
165 170 175
Glu Glu Ile Ala Val Leu Arg Arg Ala Gly Glu Ile Ser Ala Leu Ala
180 185 190
His Thr Arg Ala Met Glu Lys Cys Arg Pro Gly Met Phe Glu Tyr Gln
195 200 205
Leu Glu Gly Glu Ile Leu His Glu Phe Thr Arg His Gly Ala Arg Tyr
210 215 220
Pro Ala Tyr Asn Thr Ile Val Gly Gly Gly Glu Asn Gly Cys Ile Leu
225 230 235 240
His Tyr Thr Glu Asn Glu Cys Glu Leu Arg Asp Gly Asp Leu Val Leu
245 250 255
Ile Asp Ala Gly Cys Glu Tyr Arg Gly Tyr Ala Gly Asp Ile Thr Arg
260 265 270
Thr Phe Pro Val Asn Gly Lys Phe Thr Pro Ala Gln Arg Ala Val Tyr
275 280 285
Asp Ile Val Leu Ala Ala Ile Asn Lys Ser Leu Thr Leu Phe Arg Pro
290 295 300
Gly Thr Ser Ile Arg Glu Val Thr Glu Glu Val Val Arg Ile Met Val
305 310 315 320
Val Gly Leu Val Glu Leu Gly Ile Leu Lys Gly Asp Ile Glu Gln Leu
325 330 335
Ile Ala Glu Gln Ala His Arg Pro Phe Phe Met His Gly Leu Ser His
340 345 350
Trp Leu Gly Met Asp Val His Asp Val Gly Asp Tyr Gly Ser Ser Asp
355 360 365
Arg Gly Arg Ile Leu Glu Pro Gly Met Val Leu Thr Val Glu Pro Gly
370 375 380
Leu Tyr Ile Ala Pro Asp Ala Asp Val Pro Pro Gln Tyr Arg Gly Ile
385 390 395 400
Gly Ile Arg Ile Glu Asp Asp Ile Val Ile Thr Ala Thr Gly Asn Glu
405 410 415
Asn Leu Thr Ala Ser Val Val Lys Asp Pro Asp Asp Ile Glu Ala Leu
420 425 430
Met Ala Leu Asn His Ala Gly Glu Asn Leu Tyr Phe Gln Glu
435 440 445
<210> 49
<211> 303
<212> PRT
<213> 激烈火球菌
<400> 49
Met Asp Thr Glu Lys Leu Met Lys Ala Gly Glu Ile Ala Lys Lys Val
1 5 10 15
Arg Glu Lys Ala Ile Lys Leu Ala Arg Pro Gly Met Leu Leu Leu Glu
20 25 30
Leu Ala Glu Ser Ile Glu Lys Met Ile Met Glu Leu Gly Gly Lys Pro
35 40 45
Ala Phe Pro Val Asn Leu Ser Ile Asn Glu Ile Ala Ala His Tyr Thr
50 55 60
Pro Tyr Lys Gly Asp Thr Thr Val Leu Lys Glu Gly Asp Tyr Leu Lys
65 70 75 80
Ile Asp Val Gly Val His Ile Asp Gly Phe Ile Ala Asp Thr Ala Val
85 90 95
Thr Val Arg Val Gly Met Glu Glu Asp Glu Leu Met Glu Ala Ala Lys
100 105 110
Glu Ala Leu Asn Ala Ala Ile Ser Val Ala Arg Ala Gly Val Glu Ile
115 120 125
Lys Glu Leu Gly Lys Ala Ile Glu Asn Glu Ile Arg Lys Arg Gly Phe
130 135 140
Lys Pro Ile Val Asn Leu Ser Gly His Lys Ile Glu Arg Tyr Lys Leu
145 150 155 160
His Ala Gly Ile Ser Ile Pro Asn Ile Tyr Arg Pro His Asp Asn Tyr
165 170 175
Val Leu Lys Glu Gly Asp Val Phe Ala Ile Glu Pro Phe Ala Thr Ile
180 185 190
Gly Ala Gly Gln Val Ile Glu Val Pro Pro Thr Leu Ile Tyr Met Tyr
195 200 205
Val Arg Asp Val Pro Val Arg Val Ala Gln Ala Arg Phe Leu Leu Ala
210 215 220
Lys Ile Lys Arg Glu Tyr Gly Thr Leu Pro Phe Ala Tyr Arg Trp Leu
225 230 235 240
Gln Asn Asp Met Pro Glu Gly Gln Leu Lys Leu Ala Leu Lys Thr Leu
245 250 255
Glu Lys Ala Gly Ala Ile Tyr Gly Tyr Pro Val Leu Lys Glu Ile Arg
260 265 270
Asn Gly Ile Val Ala Gln Phe Glu His Thr Ile Ile Val Glu Lys Asp
275 280 285
Ser Val Ile Val Thr Gln Asp Met Ile Asn Lys Ser Thr Leu Glu
290 295 300
<210> 50
<211> 428
<212> PRT
<213> 温和气单胞菌
<400> 50
His Met Ser Ser Pro Leu His Tyr Val Leu Asp Gly Ile His Cys Glu
1 5 10 15
Pro His Phe Phe Thr Val Pro Leu Asp His Gln Gln Pro Asp Asp Glu
20 25 30
Glu Thr Ile Thr Leu Phe Gly Arg Thr Leu Cys Arg Lys Asp Arg Leu
35 40 45
Asp Asp Glu Leu Pro Trp Leu Leu Tyr Leu Gln Gly Gly Pro Gly Phe
50 55 60
Gly Ala Pro Arg Pro Ser Ala Asn Gly Gly Trp Ile Lys Arg Ala Leu
65 70 75 80
Gln Glu Phe Arg Val Leu Leu Leu Asp Gln Arg Gly Thr Gly His Ser
85 90 95
Thr Pro Ile His Ala Glu Leu Leu Ala His Leu Asn Pro Arg Gln Gln
100 105 110
Ala Asp Tyr Leu Ser His Phe Arg Ala Asp Ser Ile Val Arg Asp Ala
115 120 125
Glu Leu Ile Arg Glu Gln Leu Ser Pro Asp His Pro Trp Ser Leu Leu
130 135 140
Gly Gln Ser Phe Gly Gly Phe Cys Ser Leu Thr Tyr Leu Ser Leu Phe
145 150 155 160
Pro Asp Ser Leu His Glu Val Tyr Leu Thr Gly Gly Val Ala Pro Ile
165 170 175
Gly Arg Ser Ala Asp Glu Val Tyr Arg Ala Thr Tyr Gln Arg Val Ala
180 185 190
Asp Lys Asn Arg Ala Phe Phe Ala Arg Phe Pro His Ala Gln Ala Ile
195 200 205
Ala Asn Arg Leu Ala Thr His Leu Gln Arg His Asp Val Arg Leu Pro
210 215 220
Asn Gly Gln Arg Leu Thr Val Glu Gln Leu Gln Gln Gln Gly Leu Asp
225 230 235 240
Leu Gly Ala Ser Gly Ala Phe Glu Glu Leu Tyr Tyr Leu Leu Glu Asp
245 250 255
Ala Phe Ile Gly Glu Lys Leu Asn Pro Ala Phe Leu Tyr Gln Val Gln
260 265 270
Ala Met Gln Pro Phe Asn Thr Asn Pro Val Phe Ala Ile Leu His Glu
275 280 285
Leu Ile Tyr Cys Glu Gly Ala Ala Ser His Trp Ala Ala Glu Arg Val
290 295 300
Arg Gly Glu Phe Pro Ala Leu Ala Trp Ala Gln Gly Lys Asp Phe Ala
305 310 315 320
Phe Thr Gly Glu Met Ile Phe Pro Trp Met Phe Glu Gln Phe Arg Glu
325 330 335
Leu Ile Pro Leu Lys Glu Ala Ala His Leu Leu Ala Glu Lys Ala Asp
340 345 350
Trp Gly Pro Leu Tyr Asp Pro Val Gln Leu Ala Arg Asn Lys Val Pro
355 360 365
Val Ala Cys Ala Val Tyr Ala Glu Asp Met Tyr Val Glu Phe Asp Tyr
370 375 380
Ser Arg Glu Thr Leu Lys Gly Leu Ser Asn Ser Arg Ala Trp Ile Thr
385 390 395 400
Asn Glu Tyr Glu His Asn Gly Leu Arg Val Asp Gly Glu Gln Ile Leu
405 410 415
Asp Arg Leu Ile Arg Leu Asn Arg Asp Cys Leu Glu
420 425
<210> 51
<211> 348
<212> PRT
<213> 激烈火球菌
<400> 51
Met Lys Glu Arg Leu Glu Lys Leu Val Lys Phe Met Asp Glu Asn Ser
1 5 10 15
Ile Asp Arg Val Phe Ile Ala Lys Pro Val Asn Val Tyr Tyr Phe Ser
20 25 30
Gly Thr Ser Pro Leu Gly Gly Gly Tyr Ile Ile Val Asp Gly Asp Glu
35 40 45
Ala Thr Leu Tyr Val Pro Glu Leu Glu Tyr Glu Met Ala Lys Glu Glu
50 55 60
Ser Lys Leu Pro Val Val Lys Phe Lys Lys Phe Asp Glu Ile Tyr Glu
65 70 75 80
Ile Leu Lys Asn Thr Glu Thr Leu Gly Ile Glu Gly Thr Leu Ser Tyr
85 90 95
Ser Met Val Glu Asn Phe Lys Glu Lys Ser Asn Val Lys Glu Phe Lys
100 105 110
Lys Ile Asp Asp Val Ile Lys Asp Leu Arg Ile Ile Lys Thr Lys Glu
115 120 125
Glu Ile Glu Ile Ile Glu Lys Ala Cys Glu Ile Ala Asp Lys Ala Val
130 135 140
Met Ala Ala Ile Glu Glu Ile Thr Glu Gly Lys Arg Glu Arg Glu Val
145 150 155 160
Ala Ala Lys Val Glu Tyr Leu Met Lys Met Asn Gly Ala Glu Lys Pro
165 170 175
Ala Phe Asp Thr Ile Ile Ala Ser Gly His Arg Ser Ala Leu Pro His
180 185 190
Gly Val Ala Ser Asp Lys Arg Ile Glu Arg Gly Asp Leu Val Val Ile
195 200 205
Asp Leu Gly Ala Leu Tyr Asn His Tyr Asn Ser Asp Ile Thr Arg Thr
210 215 220
Ile Val Val Gly Ser Pro Asn Glu Lys Gln Arg Glu Ile Tyr Glu Ile
225 230 235 240
Val Leu Glu Ala Gln Lys Arg Ala Val Glu Ala Ala Lys Pro Gly Met
245 250 255
Thr Ala Lys Glu Leu Asp Ser Ile Ala Arg Glu Ile Ile Lys Glu Tyr
260 265 270
Gly Tyr Gly Asp Tyr Phe Ile His Ser Leu Gly His Gly Val Gly Leu
275 280 285
Glu Ile His Glu Trp Pro Arg Ile Ser Gln Tyr Asp Glu Thr Val Leu
290 295 300
Lys Glu Gly Met Val Ile Thr Ile Glu Pro Gly Ile Tyr Ile Pro Lys
305 310 315 320
Leu Gly Gly Val Arg Ile Glu Asp Thr Val Leu Ile Thr Glu Asn Gly
325 330 335
Ala Lys Arg Leu Thr Lys Thr Glu Arg Glu Leu Leu
340 345
<210> 52
<211> 298
<212> PRT
<213> 脑膜败血伊莉莎白菌
<400> 52
Met Ile Pro Ile Thr Thr Pro Val Gly Asn Phe Lys Val Trp Thr Lys
1 5 10 15
Arg Phe Gly Thr Asn Pro Lys Ile Lys Val Leu Leu Leu His Gly Gly
20 25 30
Pro Ala Met Thr His Glu Tyr Met Glu Cys Phe Glu Thr Phe Phe Gln
35 40 45
Arg Glu Gly Phe Glu Phe Tyr Glu Tyr Asp Gln Leu Gly Ser Tyr Tyr
50 55 60
Ser Asp Gln Pro Thr Asp Glu Lys Leu Trp Asn Ile Asp Arg Phe Val
65 70 75 80
Asp Glu Val Glu Gln Val Arg Lys Ala Ile His Ala Asp Lys Glu Asn
85 90 95
Phe Tyr Val Leu Gly Asn Ser Trp Gly Gly Ile Leu Ala Met Glu Tyr
100 105 110
Ala Leu Lys Tyr Gln Gln Asn Leu Lys Gly Leu Ile Val Ala Asn Met
115 120 125
Met Ala Ser Ala Pro Glu Tyr Val Lys Tyr Ala Glu Val Leu Ser Lys
130 135 140
Gln Met Lys Pro Glu Val Leu Ala Glu Val Arg Ala Ile Glu Ala Lys
145 150 155 160
Lys Asp Tyr Ala Asn Pro Arg Tyr Thr Glu Leu Leu Phe Pro Asn Tyr
165 170 175
Tyr Ala Gln His Ile Cys Arg Leu Lys Glu Trp Pro Asp Ala Leu Asn
180 185 190
Arg Ser Leu Lys His Val Asn Ser Thr Val Tyr Thr Leu Met Gln Gly
195 200 205
Pro Ser Glu Leu Gly Met Ser Ser Asp Ala Arg Leu Ala Lys Trp Asp
210 215 220
Ile Lys Asn Arg Leu His Glu Ile Ala Thr Pro Thr Leu Met Ile Gly
225 230 235 240
Ala Arg Tyr Asp Thr Met Asp Pro Lys Ala Met Glu Glu Gln Ser Lys
245 250 255
Leu Val Gln Lys Gly Arg Tyr Leu Tyr Cys Pro Asn Gly Ser His Leu
260 265 270
Ala Met Trp Asp Asp Gln Lys Val Phe Met Asp Gly Val Ile Lys Phe
275 280 285
Ile Lys Asp Val Asp Thr Lys Ser Phe Asn
290 295
<210> 53
<211> 310
<212> PRT
<213> 淋病奈瑟氏菌
<400> 53
Met Tyr Glu Ile Lys Gln Pro Phe His Ser Gly Tyr Leu Gln Val Ser
1 5 10 15
Glu Ile His Gln Ile Tyr Trp Glu Glu Ser Gly Asn Pro Asp Gly Val
20 25 30
Pro Val Ile Phe Leu His Gly Gly Pro Gly Ala Gly Ala Ser Pro Glu
35 40 45
Cys Arg Gly Phe Phe Asn Pro Asp Val Phe Arg Ile Val Ile Ile Asp
50 55 60
Gln Arg Gly Cys Gly Arg Ser His Pro Tyr Ala Cys Ala Glu Asp Asn
65 70 75 80
Thr Thr Trp Asp Leu Val Ala Asp Ile Glu Lys Val Arg Glu Met Leu
85 90 95
Gly Ile Gly Lys Trp Leu Val Phe Gly Gly Ser Trp Gly Ser Thr Leu
100 105 110
Ser Leu Ala Tyr Ala Gln Thr His Pro Glu Arg Val Lys Gly Leu Val
115 120 125
Leu Arg Gly Ile Phe Leu Cys Arg Pro Ser Glu Thr Ala Trp Leu Asn
130 135 140
Glu Ala Gly Gly Val Ser Arg Ile Tyr Pro Glu Gln Trp Gln Lys Phe
145 150 155 160
Val Ala Pro Ile Ala Glu Asn Arg Arg Asn Arg Leu Ile Glu Ala Tyr
165 170 175
His Gly Leu Leu Phe His Gln Asp Glu Glu Val Cys Leu Ser Ala Ala
180 185 190
Lys Ala Trp Ala Asp Trp Glu Ser Tyr Leu Ile Arg Phe Glu Pro Glu
195 200 205
Gly Val Asp Glu Asp Ala Tyr Ala Ser Leu Ala Ile Ala Arg Leu Glu
210 215 220
Asn His Tyr Phe Val Asn Gly Gly Trp Leu Gln Gly Asp Lys Ala Ile
225 230 235 240
Leu Asn Asn Ile Gly Lys Ile Arg His Ile Pro Thr Val Ile Val Gln
245 250 255
Gly Arg Tyr Asp Leu Cys Thr Pro Met Gln Ser Ala Trp Glu Leu Ser
260 265 270
Lys Ala Phe Pro Glu Ala Glu Leu Arg Val Val Gln Ala Gly His Cys
275 280 285
Ala Phe Asp Pro Pro Leu Ala Asp Ala Leu Val Gln Ala Val Glu Asp
290 295 300
Ile Leu Pro Arg Leu Leu
305 310
<210> 54
<211> 870
<212> PRT
<213> 大肠杆菌
<400> 54
Met Thr Gln Gln Pro Gln Ala Lys Tyr Arg His Asp Tyr Arg Ala Pro
1 5 10 15
Asp Tyr Gln Ile Thr Asp Ile Asp Leu Thr Phe Asp Leu Asp Ala Gln
20 25 30
Lys Thr Val Val Thr Ala Val Ser Gln Ala Val Arg His Gly Ala Ser
35 40 45
Asp Ala Pro Leu Arg Leu Asn Gly Glu Asp Leu Lys Leu Val Ser Val
50 55 60
His Ile Asn Asp Glu Pro Trp Thr Ala Trp Lys Glu Glu Glu Gly Ala
65 70 75 80
Leu Val Ile Ser Asn Leu Pro Glu Arg Phe Thr Leu Lys Ile Ile Asn
85 90 95
Glu Ile Ser Pro Ala Ala Asn Thr Ala Leu Glu Gly Leu Tyr Gln Ser
100 105 110
Gly Asp Ala Leu Cys Thr Gln Cys Glu Ala Glu Gly Phe Arg His Ile
115 120 125
Thr Tyr Tyr Leu Asp Arg Pro Asp Val Leu Ala Arg Phe Thr Thr Lys
130 135 140
Ile Ile Ala Asp Lys Ile Lys Tyr Pro Phe Leu Leu Ser Asn Gly Asn
145 150 155 160
Arg Val Ala Gln Gly Glu Leu Glu Asn Gly Arg His Trp Val Gln Trp
165 170 175
Gln Asp Pro Phe Pro Lys Pro Cys Tyr Leu Phe Ala Leu Val Ala Gly
180 185 190
Asp Phe Asp Val Leu Arg Asp Thr Phe Thr Thr Arg Ser Gly Arg Glu
195 200 205
Val Ala Leu Glu Leu Tyr Val Asp Arg Gly Asn Leu Asp Arg Ala Pro
210 215 220
Trp Ala Met Thr Ser Leu Lys Asn Ser Met Lys Trp Asp Glu Glu Arg
225 230 235 240
Phe Gly Leu Glu Tyr Asp Leu Asp Ile Tyr Met Ile Val Ala Val Asp
245 250 255
Phe Phe Asn Met Gly Ala Met Glu Asn Lys Gly Leu Asn Ile Phe Asn
260 265 270
Ser Lys Tyr Val Leu Ala Arg Thr Asp Thr Ala Thr Asp Lys Asp Tyr
275 280 285
Leu Asp Ile Glu Arg Val Ile Gly His Glu Tyr Phe His Asn Trp Thr
290 295 300
Gly Asn Arg Val Thr Cys Arg Asp Trp Phe Gln Leu Ser Leu Lys Glu
305 310 315 320
Gly Leu Thr Val Phe Arg Asp Gln Glu Phe Ser Ser Asp Leu Gly Ser
325 330 335
Arg Ala Val Asn Arg Ile Asn Asn Val Arg Thr Met Arg Gly Leu Gln
340 345 350
Phe Ala Glu Asp Ala Ser Pro Met Ala His Pro Ile Arg Pro Asp Met
355 360 365
Val Ile Glu Met Asn Asn Phe Tyr Thr Leu Thr Val Tyr Glu Lys Gly
370 375 380
Ala Glu Val Ile Arg Met Ile His Thr Leu Leu Gly Glu Glu Asn Phe
385 390 395 400
Gln Lys Gly Met Gln Leu Tyr Phe Glu Arg His Asp Gly Ser Ala Ala
405 410 415
Thr Cys Asp Asp Phe Val Gln Ala Met Glu Asp Ala Ser Asn Val Asp
420 425 430
Leu Ser His Phe Arg Arg Trp Tyr Ser Gln Ser Gly Thr Pro Ile Val
435 440 445
Thr Val Lys Asp Asp Tyr Asn Pro Glu Thr Glu Gln Tyr Thr Leu Thr
450 455 460
Ile Ser Gln Arg Thr Pro Ala Thr Pro Asp Gln Ala Glu Lys Gln Pro
465 470 475 480
Leu His Ile Pro Phe Ala Ile Glu Leu Tyr Asp Asn Glu Gly Lys Val
485 490 495
Ile Pro Leu Gln Lys Gly Gly His Pro Val Asn Ser Val Leu Asn Val
500 505 510
Thr Gln Ala Glu Gln Thr Phe Val Phe Asp Asn Val Tyr Phe Gln Pro
515 520 525
Val Pro Ala Leu Leu Cys Glu Phe Ser Ala Pro Val Lys Leu Glu Tyr
530 535 540
Lys Trp Ser Asp Gln Gln Leu Thr Phe Leu Met Arg His Ala Arg Asn
545 550 555 560
Asp Phe Ser Arg Trp Asp Ala Ala Gln Ser Leu Leu Ala Thr Tyr Ile
565 570 575
Lys Leu Asn Val Ala Arg His Gln Gln Gly Gln Pro Leu Ser Leu Pro
580 585 590
Val His Val Ala Asp Ala Phe Arg Ala Val Leu Leu Asp Glu Lys Ile
595 600 605
Asp Pro Ala Leu Ala Ala Glu Ile Leu Thr Leu Pro Ser Val Asn Glu
610 615 620
Met Ala Glu Leu Phe Asp Ile Ile Asp Pro Ile Ala Ile Ala Glu Val
625 630 635 640
Arg Glu Ala Leu Thr Arg Thr Leu Ala Thr Glu Leu Ala Asp Glu Leu
645 650 655
Leu Ala Ile Tyr Asn Ala Asn Tyr Gln Ser Glu Tyr Arg Val Glu His
660 665 670
Glu Asp Ile Ala Lys Arg Thr Leu Arg Asn Ala Cys Leu Arg Phe Leu
675 680 685
Ala Phe Gly Glu Thr His Leu Ala Asp Val Leu Val Ser Lys Gln Phe
690 695 700
His Glu Ala Asn Asn Met Thr Asp Ala Leu Ala Ala Leu Ser Ala Ala
705 710 715 720
Val Ala Ala Gln Leu Pro Cys Arg Asp Ala Leu Met Gln Glu Tyr Asp
725 730 735
Asp Lys Trp His Gln Asn Gly Leu Val Met Asp Lys Trp Phe Ile Leu
740 745 750
Gln Ala Thr Ser Pro Ala Ala Asn Val Leu Glu Thr Val Arg Gly Leu
755 760 765
Leu Gln His Arg Ser Phe Thr Met Ser Asn Pro Asn Arg Ile Arg Ser
770 775 780
Leu Ile Gly Ala Phe Ala Gly Ser Asn Pro Ala Ala Phe His Ala Glu
785 790 795 800
Asp Gly Ser Gly Tyr Leu Phe Leu Val Glu Met Leu Thr Asp Leu Asn
805 810 815
Ser Arg Asn Pro Gln Val Ala Ser Arg Leu Ile Glu Pro Leu Ile Arg
820 825 830
Leu Lys Arg Tyr Asp Ala Lys Arg Gln Glu Lys Met Arg Ala Ala Leu
835 840 845
Glu Gln Leu Lys Gly Leu Glu Asn Leu Ser Gly Asp Leu Tyr Glu Lys
850 855 860
Ile Thr Lys Ala Leu Ala
865 870
<210> 55
<211> 889
<212> PRT
<213> 恶性疟原虫
<400> 55
Pro Lys Ile His Tyr Arg Lys Asp Tyr Lys Pro Ser Gly Phe Ile Ile
1 5 10 15
Asn Gln Val Thr Leu Asn Ile Asn Ile His Asp Gln Glu Thr Ile Val
20 25 30
Arg Ser Val Leu Asp Met Asp Ile Ser Lys His Asn Val Gly Glu Asp
35 40 45
Leu Val Phe Asp Gly Val Gly Leu Lys Ile Asn Glu Ile Ser Ile Asn
50 55 60
Asn Lys Lys Leu Val Glu Gly Glu Glu Tyr Thr Tyr Asp Asn Glu Phe
65 70 75 80
Leu Thr Ile Phe Ser Lys Phe Val Pro Lys Ser Lys Phe Ala Phe Ser
85 90 95
Ser Glu Val Ile Ile His Pro Glu Thr Asn Tyr Ala Leu Thr Gly Leu
100 105 110
Tyr Lys Ser Lys Asn Ile Ile Val Ser Gln Cys Glu Ala Thr Gly Phe
115 120 125
Arg Arg Ile Thr Phe Phe Ile Asp Arg Pro Asp Met Met Ala Lys Tyr
130 135 140
Asp Val Thr Val Thr Ala Asp Lys Glu Lys Tyr Pro Val Leu Leu Ser
145 150 155 160
Asn Gly Asp Lys Val Asn Glu Phe Glu Ile Pro Gly Gly Arg His Gly
165 170 175
Ala Arg Phe Asn Asp Pro Pro Leu Lys Pro Cys Tyr Leu Phe Ala Val
180 185 190
Val Ala Gly Asp Leu Lys His Leu Ser Ala Thr Tyr Ile Thr Lys Tyr
195 200 205
Thr Lys Lys Lys Val Glu Leu Tyr Val Phe Ser Glu Glu Lys Tyr Val
210 215 220
Ser Lys Leu Gln Trp Ala Leu Glu Cys Leu Lys Lys Ser Met Ala Phe
225 230 235 240
Asp Glu Asp Tyr Phe Gly Leu Glu Tyr Asp Leu Ser Arg Leu Asn Leu
245 250 255
Val Ala Val Ser Asp Phe Asn Val Gly Ala Met Glu Asn Lys Gly Leu
260 265 270
Asn Ile Phe Asn Ala Asn Ser Leu Leu Ala Ser Lys Lys Asn Ser Ile
275 280 285
Asp Phe Ser Tyr Ala Arg Ile Leu Thr Val Val Gly His Glu Tyr Phe
290 295 300
His Gln Tyr Thr Gly Asn Arg Val Thr Leu Arg Asp Trp Phe Gln Leu
305 310 315 320
Thr Leu Lys Glu Gly Leu Thr Val His Arg Glu Asn Leu Phe Ser Glu
325 330 335
Glu Met Thr Lys Thr Val Thr Thr Arg Leu Ser His Val Asp Leu Leu
340 345 350
Arg Ser Val Gln Phe Leu Glu Asp Ser Ser Pro Leu Ser His Pro Ile
355 360 365
Arg Pro Glu Ser Tyr Val Ser Met Glu Asn Phe Tyr Thr Thr Thr Val
370 375 380
Tyr Asp Lys Gly Ser Glu Val Met Arg Met Tyr Leu Thr Ile Leu Gly
385 390 395 400
Glu Glu Tyr Tyr Lys Lys Gly Phe Asp Ile Tyr Ile Lys Lys Asn Asp
405 410 415
Gly Asn Thr Ala Thr Cys Glu Asp Phe Asn Tyr Ala Met Glu Gln Ala
420 425 430
Tyr Lys Met Lys Lys Ala Asp Asn Ser Ala Asn Leu Asn Gln Tyr Leu
435 440 445
Leu Trp Phe Ser Gln Ser Gly Thr Pro His Val Ser Phe Lys Tyr Asn
450 455 460
Tyr Asp Ala Glu Lys Lys Gln Tyr Ser Ile His Val Asn Gln Tyr Thr
465 470 475 480
Lys Pro Asp Glu Asn Gln Lys Glu Lys Lys Pro Leu Phe Ile Pro Ile
485 490 495
Ser Val Gly Leu Ile Asn Pro Glu Asn Gly Lys Glu Met Ile Ser Gln
500 505 510
Thr Thr Leu Glu Leu Thr Lys Glu Ser Asp Thr Phe Val Phe Asn Asn
515 520 525
Ile Ala Val Lys Pro Ile Pro Ser Leu Phe Arg Gly Phe Ser Ala Pro
530 535 540
Val Tyr Ile Glu Asp Gln Leu Thr Asp Glu Glu Arg Ile Leu Leu Leu
545 550 555 560
Lys Tyr Asp Ser Asp Ala Phe Val Arg Tyr Asn Ser Cys Thr Asn Ile
565 570 575
Tyr Met Lys Gln Ile Leu Met Asn Tyr Asn Glu Phe Leu Lys Ala Lys
580 585 590
Asn Glu Lys Leu Glu Ser Phe Gln Leu Thr Pro Val Asn Ala Gln Phe
595 600 605
Ile Asp Ala Ile Lys Tyr Leu Leu Glu Asp Pro His Ala Asp Ala Gly
610 615 620
Phe Lys Ser Tyr Ile Val Ser Leu Pro Gln Asp Arg Tyr Ile Ile Asn
625 630 635 640
Phe Val Ser Asn Leu Asp Thr Asp Val Leu Ala Asp Thr Lys Glu Tyr
645 650 655
Ile Tyr Lys Gln Ile Gly Asp Lys Leu Asn Asp Val Tyr Tyr Lys Met
660 665 670
Phe Lys Ser Leu Glu Ala Lys Ala Asp Asp Leu Thr Tyr Phe Asn Asp
675 680 685
Glu Ser His Val Asp Phe Asp Gln Met Asn Met Arg Thr Leu Arg Asn
690 695 700
Thr Leu Leu Ser Leu Leu Ser Lys Ala Gln Tyr Pro Asn Ile Leu Asn
705 710 715 720
Glu Ile Ile Glu His Ser Lys Ser Pro Tyr Pro Ser Asn Trp Leu Thr
725 730 735
Ser Leu Ser Val Ser Ala Tyr Phe Asp Lys Tyr Phe Glu Leu Tyr Asp
740 745 750
Lys Thr Tyr Lys Leu Ser Lys Asp Asp Glu Leu Leu Leu Gln Glu Trp
755 760 765
Leu Lys Thr Val Ser Arg Ser Asp Arg Lys Asp Ile Tyr Glu Ile Leu
770 775 780
Lys Lys Leu Glu Asn Glu Val Leu Lys Asp Ser Lys Asn Pro Asn Asp
785 790 795 800
Ile Arg Ala Val Tyr Leu Pro Phe Thr Asn Asn Leu Arg Arg Phe His
805 810 815
Asp Ile Ser Gly Lys Gly Tyr Lys Leu Ile Ala Glu Val Ile Thr Lys
820 825 830
Thr Asp Lys Phe Asn Pro Met Val Ala Thr Gln Leu Cys Glu Pro Phe
835 840 845
Lys Leu Trp Asn Lys Leu Asp Thr Lys Arg Gln Glu Leu Met Leu Asn
850 855 860
Glu Met Asn Thr Met Leu Gln Glu Pro Gln Ile Ser Asn Asn Leu Lys
865 870 875 880
Glu Tyr Leu Leu Arg Leu Thr Asn Lys
885
<210> 56
<211> 919
<212> PRT
<213> 未知
<220>
<223> 嘌呤霉素敏感型氨肽酶
<400> 56
Met Trp Leu Ala Ala Ala Ala Pro Ser Leu Ala Arg Arg Leu Leu Phe
1 5 10 15
Leu Gly Pro Pro Pro Pro Pro Leu Leu Leu Leu Val Phe Ser Arg Ser
20 25 30
Ser Arg Arg Arg Leu His Ser Leu Gly Leu Ala Ala Met Pro Glu Lys
35 40 45
Arg Pro Phe Glu Arg Leu Pro Ala Asp Val Ser Pro Ile Asn Tyr Ser
50 55 60
Leu Cys Leu Lys Pro Asp Leu Leu Asp Phe Thr Phe Glu Gly Lys Leu
65 70 75 80
Glu Ala Ala Ala Gln Val Arg Gln Ala Thr Asn Gln Ile Val Met Asn
85 90 95
Cys Ala Asp Ile Asp Ile Ile Thr Ala Ser Tyr Ala Pro Glu Gly Asp
100 105 110
Glu Glu Ile His Ala Thr Gly Phe Asn Tyr Gln Asn Glu Asp Glu Lys
115 120 125
Val Thr Leu Ser Phe Pro Ser Thr Leu Gln Thr Gly Thr Gly Thr Leu
130 135 140
Lys Ile Asp Phe Val Gly Glu Leu Asn Asp Lys Met Lys Gly Phe Tyr
145 150 155 160
Arg Ser Lys Tyr Thr Thr Pro Ser Gly Glu Val Arg Tyr Ala Ala Val
165 170 175
Thr Gln Phe Glu Ala Thr Asp Ala Arg Arg Ala Phe Pro Cys Trp Asp
180 185 190
Glu Pro Ala Ile Lys Ala Thr Phe Asp Ile Ser Leu Val Val Pro Lys
195 200 205
Asp Arg Val Ala Leu Ser Asn Met Asn Val Ile Asp Arg Lys Pro Tyr
210 215 220
Pro Asp Asp Glu Asn Leu Val Glu Val Lys Phe Ala Arg Thr Pro Val
225 230 235 240
Met Ser Thr Tyr Leu Val Ala Phe Val Val Gly Glu Tyr Asp Phe Val
245 250 255
Glu Thr Arg Ser Lys Asp Gly Val Cys Val Arg Val Tyr Thr Pro Val
260 265 270
Gly Lys Ala Glu Gln Gly Lys Phe Ala Leu Glu Val Ala Ala Lys Thr
275 280 285
Leu Pro Phe Tyr Lys Asp Tyr Phe Asn Val Pro Tyr Pro Leu Pro Lys
290 295 300
Ile Asp Leu Ile Ala Ile Ala Asp Phe Ala Ala Gly Ala Met Glu Asn
305 310 315 320
Trp Gly Leu Val Thr Tyr Arg Glu Thr Ala Leu Leu Ile Asp Pro Lys
325 330 335
Asn Ser Cys Ser Ser Ser Arg Gln Trp Val Ala Leu Val Val Gly His
340 345 350
Glu Leu Ala His Gln Trp Phe Gly Asn Leu Val Thr Met Glu Trp Trp
355 360 365
Thr His Leu Trp Leu Asn Glu Gly Phe Ala Ser Trp Ile Glu Tyr Leu
370 375 380
Cys Val Asp His Cys Phe Pro Glu Tyr Asp Ile Trp Thr Gln Phe Val
385 390 395 400
Ser Ala Asp Tyr Thr Arg Ala Gln Glu Leu Asp Ala Leu Asp Asn Ser
405 410 415
His Pro Ile Glu Val Ser Val Gly His Pro Ser Glu Val Asp Glu Ile
420 425 430
Phe Asp Ala Ile Ser Tyr Ser Lys Gly Ala Ser Val Ile Arg Met Leu
435 440 445
His Asp Tyr Ile Gly Asp Lys Asp Phe Lys Lys Gly Met Asn Met Tyr
450 455 460
Leu Thr Lys Phe Gln Gln Lys Asn Ala Ala Thr Glu Asp Leu Trp Glu
465 470 475 480
Ser Leu Glu Asn Ala Ser Gly Lys Pro Ile Ala Ala Val Met Asn Thr
485 490 495
Trp Thr Lys Gln Met Gly Phe Pro Leu Ile Tyr Val Glu Ala Glu Gln
500 505 510
Val Glu Asp Asp Arg Leu Leu Arg Leu Ser Gln Lys Lys Phe Cys Ala
515 520 525
Gly Gly Ser Tyr Val Gly Glu Asp Cys Pro Gln Trp Met Val Pro Ile
530 535 540
Thr Ile Ser Thr Ser Glu Asp Pro Asn Gln Ala Lys Leu Lys Ile Leu
545 550 555 560
Met Asp Lys Pro Glu Met Asn Val Val Leu Lys Asn Val Lys Pro Asp
565 570 575
Gln Trp Val Lys Leu Asn Leu Gly Thr Val Gly Phe Tyr Arg Thr Gln
580 585 590
Tyr Ser Ser Ala Met Leu Glu Ser Leu Leu Pro Gly Ile Arg Asp Leu
595 600 605
Ser Leu Pro Pro Val Asp Arg Leu Gly Leu Gln Asn Asp Leu Phe Ser
610 615 620
Leu Ala Arg Ala Gly Ile Ile Ser Thr Val Glu Val Leu Lys Val Met
625 630 635 640
Glu Ala Phe Val Asn Glu Pro Asn Tyr Thr Val Trp Ser Asp Leu Ser
645 650 655
Cys Asn Leu Gly Ile Leu Ser Thr Leu Leu Ser His Thr Asp Phe Tyr
660 665 670
Glu Glu Ile Gln Glu Phe Val Lys Asp Val Phe Ser Pro Ile Gly Glu
675 680 685
Arg Leu Gly Trp Asp Pro Lys Pro Gly Glu Gly His Leu Asp Ala Leu
690 695 700
Leu Arg Gly Leu Val Leu Gly Lys Leu Gly Lys Ala Gly His Lys Ala
705 710 715 720
Thr Leu Glu Glu Ala Arg Arg Arg Phe Lys Asp His Val Glu Gly Lys
725 730 735
Gln Ile Leu Ser Ala Asp Leu Arg Ser Pro Val Tyr Leu Thr Val Leu
740 745 750
Lys His Gly Asp Gly Thr Thr Leu Asp Ile Met Leu Lys Leu His Lys
755 760 765
Gln Ala Asp Met Gln Glu Glu Lys Asn Arg Ile Glu Arg Val Leu Gly
770 775 780
Ala Thr Leu Leu Pro Asp Leu Ile Gln Lys Val Leu Thr Phe Ala Leu
785 790 795 800
Ser Glu Glu Val Arg Pro Gln Asp Thr Val Ser Val Ile Gly Gly Val
805 810 815
Ala Gly Gly Ser Lys His Gly Arg Lys Ala Ala Trp Lys Phe Ile Lys
820 825 830
Asp Asn Trp Glu Glu Leu Tyr Asn Arg Tyr Gln Gly Gly Phe Leu Ile
835 840 845
Ser Arg Leu Ile Lys Leu Ser Val Glu Gly Phe Ala Val Asp Lys Met
850 855 860
Ala Gly Glu Val Lys Ala Phe Phe Glu Ser His Pro Ala Pro Ser Ala
865 870 875 880
Glu Arg Thr Ile Gln Gln Cys Cys Glu Asn Ile Leu Leu Asn Ala Ala
885 890 895
Trp Leu Lys Arg Asp Ala Glu Ser Ile His Gln Tyr Leu Leu Gln Arg
900 905 910
Lys Ala Ser Pro Pro Thr Val
915
<210> 57
<211> 919
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 57
Met Trp Leu Ala Ala Ala Ala Pro Ser Leu Ala Arg Arg Leu Leu Phe
1 5 10 15
Leu Gly Pro Pro Pro Pro Pro Leu Leu Leu Leu Val Phe Ser Arg Ser
20 25 30
Ser Arg Arg Arg Leu His Ser Leu Gly Leu Ala Ala Met Pro Glu Lys
35 40 45
Arg Pro Phe Glu Arg Leu Pro Ala Asp Val Ser Pro Ile Asn Tyr Ser
50 55 60
Leu Cys Leu Lys Pro Asp Leu Leu Asp Phe Thr Phe Glu Gly Lys Leu
65 70 75 80
Glu Ala Ala Ala Gln Val Arg Gln Ala Thr Asn Gln Ile Val Met Asn
85 90 95
Cys Ala Asp Ile Asp Ile Ile Thr Ala Ser Tyr Ala Pro Glu Gly Asp
100 105 110
Glu Glu Ile His Ala Thr Gly Phe Asn Tyr Gln Asn Glu Asp Glu Lys
115 120 125
Val Thr Leu Ser Phe Pro Ser Thr Leu Gln Thr Gly Thr Gly Thr Leu
130 135 140
Lys Ile Asp Phe Val Gly Glu Leu Asn Asp Lys Met Lys Gly Phe Tyr
145 150 155 160
Arg Ser Lys Tyr Thr Thr Pro Ser Gly Glu Val Arg Tyr Ala Ala Val
165 170 175
Thr Gln Phe Glu Ala Thr Asp Ala Arg Arg Ala Phe Pro Cys Trp Asp
180 185 190
Glu Pro Ala Ile Lys Ala Thr Phe Asp Ile Ser Leu Val Val Pro Lys
195 200 205
Asp Arg Val Ala Leu Ser Asn Met Asn Val Ile Asp Arg Lys Pro Tyr
210 215 220
Pro Asp Asp Glu Asn Leu Val Glu Val Lys Phe Ala Arg Thr Pro Val
225 230 235 240
Met Ser Thr Tyr Leu Val Ala Phe Val Val Gly Glu Tyr Asp Phe Val
245 250 255
Glu Thr Arg Ser Lys Asp Gly Val Cys Val Arg Val Tyr Thr Pro Val
260 265 270
Gly Lys Ala Glu Gln Gly Lys Phe Ala Leu Glu Val Ala Ala Lys Thr
275 280 285
Leu Pro Phe Tyr Lys Asp Tyr Phe Asn Val Pro Tyr Pro Leu Pro Lys
290 295 300
Ile Asp Leu Ile Ala Ile Ala Asp Phe Ala Ala Gly Ala Met Glu Asn
305 310 315 320
Trp Gly Leu Val Thr Tyr Arg Glu Thr Ala Leu Leu Ile Asp Pro Lys
325 330 335
Asn Ser Cys Ser Ser Ser Arg Gln Trp Val Ala Leu Val Val Gly His
340 345 350
Val Leu Ala His Gln Trp Phe Gly Asn Leu Val Thr Met Glu Trp Trp
355 360 365
Thr His Leu Trp Leu Asn Glu Gly Phe Ala Ser Trp Ile Glu Tyr Leu
370 375 380
Cys Val Asp His Cys Phe Pro Glu Tyr Asp Ile Trp Thr Gln Phe Val
385 390 395 400
Ser Ala Asp Tyr Thr Arg Ala Gln Glu Leu Asp Ala Leu Asp Asn Ser
405 410 415
His Pro Ile Glu Val Ser Val Gly His Pro Ser Glu Val Asp Glu Ile
420 425 430
Phe Asp Ala Ile Ser Tyr Ser Lys Gly Ala Ser Val Ile Arg Met Leu
435 440 445
His Asp Tyr Ile Gly Asp Lys Asp Phe Lys Lys Gly Met Asn Met Tyr
450 455 460
Leu Thr Lys Phe Gln Gln Lys Asn Ala Ala Thr Glu Asp Leu Trp Glu
465 470 475 480
Ser Leu Glu Asn Ala Ser Gly Lys Pro Ile Ala Ala Val Met Asn Thr
485 490 495
Trp Thr Lys Gln Met Gly Phe Pro Leu Ile Tyr Val Glu Ala Glu Gln
500 505 510
Val Glu Asp Asp Arg Leu Leu Arg Leu Ser Gln Lys Lys Phe Cys Ala
515 520 525
Gly Gly Ser Tyr Val Gly Glu Asp Cys Pro Gln Trp Met Val Pro Ile
530 535 540
Thr Ile Ser Thr Ser Glu Asp Pro Asn Gln Ala Lys Leu Lys Ile Leu
545 550 555 560
Met Asp Lys Pro Glu Met Asn Val Val Leu Lys Asn Val Lys Pro Asp
565 570 575
Gln Trp Val Lys Leu Asn Leu Gly Thr Val Gly Phe Tyr Arg Thr Gln
580 585 590
Tyr Ser Ser Ala Met Leu Glu Ser Leu Leu Pro Gly Ile Arg Asp Leu
595 600 605
Ser Leu Pro Pro Val Asp Arg Leu Gly Leu Gln Asn Asp Leu Phe Ser
610 615 620
Leu Ala Arg Ala Gly Ile Ile Ser Thr Val Glu Val Leu Lys Val Met
625 630 635 640
Glu Ala Phe Val Asn Glu Pro Asn Tyr Thr Val Trp Ser Asp Leu Ser
645 650 655
Cys Asn Leu Gly Ile Leu Ser Thr Leu Leu Ser His Thr Asp Phe Tyr
660 665 670
Glu Glu Ile Gln Glu Phe Val Lys Asp Val Phe Ser Pro Ile Gly Glu
675 680 685
Arg Leu Gly Trp Asp Pro Lys Pro Gly Glu Gly His Leu Asp Ala Leu
690 695 700
Leu Arg Gly Leu Val Leu Gly Lys Leu Gly Lys Ala Gly His Lys Ala
705 710 715 720
Thr Leu Glu Glu Ala Arg Arg Arg Phe Lys Asp His Val Glu Gly Lys
725 730 735
Gln Ile Leu Ser Ala Asp Leu Arg Ser Pro Val Tyr Leu Thr Val Leu
740 745 750
Lys His Gly Asp Gly Thr Thr Leu Asp Ile Met Leu Lys Leu His Lys
755 760 765
Gln Ala Asp Met Gln Glu Glu Lys Asn Arg Ile Glu Arg Val Leu Gly
770 775 780
Ala Thr Leu Leu Pro Asp Leu Ile Gln Lys Val Leu Thr Phe Ala Leu
785 790 795 800
Ser Glu Glu Val Arg Pro Gln Asp Thr Val Ser Val Ile Gly Gly Val
805 810 815
Ala Gly Gly Ser Lys His Gly Arg Lys Ala Ala Trp Lys Phe Ile Lys
820 825 830
Asp Asn Trp Glu Glu Leu Tyr Asn Arg Tyr Gln Gly Gly Phe Leu Ile
835 840 845
Ser Arg Leu Ile Lys Leu Ser Val Glu Gly Phe Ala Val Asp Lys Met
850 855 860
Ala Gly Glu Val Lys Ala Phe Phe Glu Ser His Pro Ala Pro Ser Ala
865 870 875 880
Glu Arg Thr Ile Gln Gln Cys Cys Glu Asn Ile Leu Leu Asn Ala Ala
885 890 895
Trp Leu Lys Arg Asp Ala Glu Ser Ile His Gln Tyr Leu Leu Gln Arg
900 905 910
Lys Ala Ser Pro Pro Thr Val
915
<210> 58
<211> 864
<212> PRT
<213> 土伦病法兰西斯氏菌
<400> 58
Met Ile Tyr Glu Phe Val Met Thr Asp Pro Lys Ile Lys Tyr Leu Lys
1 5 10 15
Asp Tyr Lys Pro Ser Asn Tyr Leu Ile Asp Glu Thr His Leu Ile Phe
20 25 30
Glu Leu Asp Glu Ser Lys Thr Arg Val Thr Ala Asn Leu Tyr Ile Val
35 40 45
Ala Asn Arg Glu Asn Arg Glu Asn Asn Thr Leu Val Leu Asp Gly Val
50 55 60
Glu Leu Lys Leu Leu Ser Ile Lys Leu Asn Asn Lys His Leu Ser Pro
65 70 75 80
Ala Glu Phe Ala Val Asn Glu Asn Gln Leu Ile Ile Asn Asn Val Pro
85 90 95
Glu Lys Phe Val Leu Gln Thr Val Val Glu Ile Asn Pro Ser Ala Asn
100 105 110
Thr Ser Leu Glu Gly Leu Tyr Lys Ser Gly Asp Val Phe Ser Thr Gln
115 120 125
Cys Glu Ala Thr Gly Phe Arg Lys Ile Thr Tyr Tyr Leu Asp Arg Pro
130 135 140
Asp Val Met Ala Ala Phe Thr Val Lys Ile Ile Ala Asp Lys Lys Lys
145 150 155 160
Tyr Pro Ile Ile Leu Ser Asn Gly Asp Lys Ile Asp Ser Gly Asp Ile
165 170 175
Ser Asp Asn Gln His Phe Ala Val Trp Lys Asp Pro Phe Lys Lys Pro
180 185 190
Cys Tyr Leu Phe Ala Leu Val Ala Gly Asp Leu Ala Ser Ile Lys Asp
195 200 205
Thr Tyr Ile Thr Lys Ser Gln Arg Lys Val Ser Leu Glu Ile Tyr Ala
210 215 220
Phe Lys Gln Asp Ile Asp Lys Cys His Tyr Ala Met Gln Ala Val Lys
225 230 235 240
Asp Ser Met Lys Trp Asp Glu Asp Arg Phe Gly Leu Glu Tyr Asp Leu
245 250 255
Asp Thr Phe Met Ile Val Ala Val Pro Asp Phe Asn Ala Gly Ala Met
260 265 270
Glu Asn Lys Gly Leu Asn Ile Phe Asn Thr Lys Tyr Ile Met Ala Ser
275 280 285
Asn Lys Thr Ala Thr Asp Lys Asp Phe Glu Leu Val Gln Ser Val Val
290 295 300
Gly His Glu Tyr Phe His Asn Trp Thr Gly Asp Arg Val Thr Cys Arg
305 310 315 320
Asp Trp Phe Gln Leu Ser Leu Lys Glu Gly Leu Thr Val Phe Arg Asp
325 330 335
Gln Glu Phe Thr Ser Asp Leu Asn Ser Arg Asp Val Lys Arg Ile Asp
340 345 350
Asp Val Arg Ile Ile Arg Ser Ala Gln Phe Ala Glu Asp Ala Ser Pro
355 360 365
Met Ser His Pro Ile Arg Pro Glu Ser Tyr Ile Glu Met Asn Asn Phe
370 375 380
Tyr Thr Val Thr Val Tyr Asn Lys Gly Ala Glu Ile Ile Arg Met Ile
385 390 395 400
His Thr Leu Leu Gly Glu Glu Gly Phe Gln Lys Gly Met Lys Leu Tyr
405 410 415
Phe Glu Arg His Asp Gly Gln Ala Val Thr Cys Asp Asp Phe Val Asn
420 425 430
Ala Met Ala Asp Ala Asn Asn Arg Asp Phe Ser Leu Phe Lys Arg Trp
435 440 445
Tyr Ala Gln Ser Gly Thr Pro Asn Ile Lys Val Ser Glu Asn Tyr Asp
450 455 460
Ala Ser Ser Gln Thr Tyr Ser Leu Thr Leu Glu Gln Thr Thr Leu Pro
465 470 475 480
Thr Ala Asp Gln Lys Glu Lys Gln Ala Leu His Ile Pro Val Lys Met
485 490 495
Gly Leu Ile Asn Pro Glu Gly Lys Asn Ile Ala Glu Gln Val Ile Glu
500 505 510
Leu Lys Glu Gln Lys Gln Thr Tyr Thr Phe Glu Asn Ile Ala Ala Lys
515 520 525
Pro Val Ala Ser Leu Phe Arg Asp Phe Ser Ala Pro Val Lys Val Glu
530 535 540
His Lys Arg Ser Glu Lys Asp Leu Leu His Ile Val Lys Tyr Asp Asn
545 550 555 560
Asn Ala Phe Asn Arg Trp Asp Ser Leu Gln Gln Ile Ala Thr Asn Ile
565 570 575
Ile Leu Asn Asn Ala Asp Leu Asn Asp Glu Phe Leu Asn Ala Phe Lys
580 585 590
Ser Ile Leu His Asp Lys Asp Leu Asp Lys Ala Leu Ile Ser Asn Ala
595 600 605
Leu Leu Ile Pro Ile Glu Ser Thr Ile Ala Glu Ala Met Arg Val Ile
610 615 620
Met Val Asp Asp Ile Val Leu Ser Arg Lys Asn Val Val Asn Gln Leu
625 630 635 640
Ala Asp Lys Leu Lys Asp Asp Trp Leu Ala Val Tyr Gln Gln Cys Asn
645 650 655
Asp Asn Lys Pro Tyr Ser Leu Ser Ala Glu Gln Ile Ala Lys Arg Lys
660 665 670
Leu Lys Gly Val Cys Leu Ser Tyr Leu Met Asn Ala Ser Asp Gln Lys
675 680 685
Val Gly Thr Asp Leu Ala Gln Gln Leu Phe Asp Asn Ala Asp Asn Met
690 695 700
Thr Asp Gln Gln Thr Ala Phe Thr Glu Leu Leu Lys Ser Asn Asp Lys
705 710 715 720
Gln Val Arg Asp Asn Ala Ile Asn Glu Phe Tyr Asn Arg Trp Arg His
725 730 735
Glu Asp Leu Val Val Asn Lys Trp Leu Leu Ser Gln Ala Gln Ile Ser
740 745 750
His Glu Ser Ala Leu Asp Ile Val Lys Gly Leu Val Asn His Pro Ala
755 760 765
Tyr Asn Pro Lys Asn Pro Asn Lys Val Tyr Ser Leu Ile Gly Gly Phe
770 775 780
Gly Ala Asn Phe Leu Gln Tyr His Cys Lys Asp Gly Leu Gly Tyr Ala
785 790 795 800
Phe Met Ala Asp Thr Val Leu Ala Leu Asp Lys Phe Asn His Gln Val
805 810 815
Ala Ala Arg Met Ala Arg Asn Leu Met Ser Trp Lys Arg Tyr Asp Ser
820 825 830
Asp Arg Gln Ala Met Met Lys Asn Ala Leu Glu Lys Ile Lys Ala Ser
835 840 845
Asn Pro Ser Lys Asn Val Phe Glu Ile Val Ser Lys Ser Leu Glu Ser
850 855 860
<210> 59
<211> 353
<212> PRT
<213> 掘越氏火球菌
<400> 59
Met Glu Val Arg Asn Met Val Asp Tyr Glu Leu Leu Lys Lys Val Val
1 5 10 15
Glu Ala Pro Gly Val Ser Gly Tyr Glu Phe Leu Gly Ile Arg Asp Val
20 25 30
Val Ile Glu Glu Ile Lys Asp Tyr Val Asp Glu Val Lys Val Asp Lys
35 40 45
Leu Gly Asn Val Ile Ala His Lys Lys Gly Glu Gly Pro Lys Val Met
50 55 60
Ile Ala Ala His Met Asp Gln Ile Gly Leu Met Val Thr His Ile Glu
65 70 75 80
Lys Asn Gly Phe Leu Arg Val Ala Pro Ile Gly Gly Val Asp Pro Lys
85 90 95
Thr Leu Ile Ala Gln Arg Phe Lys Val Trp Ile Asp Lys Gly Lys Phe
100 105 110
Ile Tyr Gly Val Gly Ala Ser Val Pro Pro His Ile Gln Lys Pro Glu
115 120 125
Asp Arg Lys Lys Ala Pro Asp Trp Asp Gln Ile Phe Ile Asp Ile Gly
130 135 140
Ala Glu Ser Lys Glu Glu Ala Glu Asp Met Gly Val Lys Ile Gly Thr
145 150 155 160
Val Ile Thr Trp Asp Gly Arg Leu Glu Arg Leu Gly Lys His Arg Phe
165 170 175
Val Ser Ile Ala Phe Asp Asp Arg Ile Ala Val Tyr Thr Ile Leu Glu
180 185 190
Val Ala Lys Gln Leu Lys Asp Ala Lys Ala Asp Val Tyr Phe Val Ala
195 200 205
Thr Val Gln Glu Glu Val Gly Leu Arg Gly Ala Arg Thr Ser Ala Phe
210 215 220
Gly Ile Glu Pro Asp Tyr Gly Phe Ala Ile Asp Val Thr Ile Ala Ala
225 230 235 240
Asp Ile Pro Gly Thr Pro Glu His Lys Gln Val Thr His Leu Gly Lys
245 250 255
Gly Thr Ala Ile Lys Ile Met Asp Arg Ser Val Ile Cys His Pro Thr
260 265 270
Ile Val Arg Trp Leu Glu Glu Leu Ala Lys Lys His Glu Ile Pro Tyr
275 280 285
Gln Leu Glu Ile Leu Leu Gly Gly Gly Thr Asp Ala Gly Ala Ile His
290 295 300
Leu Thr Lys Ala Gly Val Pro Thr Gly Ala Leu Ser Val Pro Ala Arg
305 310 315 320
Tyr Ile His Ser Asn Thr Glu Val Val Asp Glu Arg Asp Val Asp Ala
325 330 335
Thr Val Glu Leu Met Thr Lys Ala Leu Glu Asn Ile His Glu Leu Lys
340 345 350
Ile
<210> 60
<211> 408
<212> PRT
<213> 水生栖热菌
<400> 60
Met Asp Ala Phe Thr Glu Asn Leu Asn Lys Leu Ala Glu Leu Ala Ile
1 5 10 15
Arg Val Gly Leu Asn Leu Glu Glu Gly Gln Glu Ile Val Ala Thr Ala
20 25 30
Pro Ile Glu Ala Val Asp Phe Val Arg Leu Leu Ala Glu Lys Ala Tyr
35 40 45
Glu Asn Gly Ala Ser Leu Phe Thr Val Leu Tyr Gly Asp Asn Leu Ile
50 55 60
Ala Arg Lys Arg Leu Ala Leu Val Pro Glu Ala His Leu Asp Arg Ala
65 70 75 80
Pro Ala Trp Leu Tyr Glu Gly Met Ala Lys Ala Phe His Glu Gly Ala
85 90 95
Ala Arg Leu Ala Val Ser Gly Asn Asp Pro Lys Ala Leu Glu Gly Leu
100 105 110
Pro Pro Glu Arg Val Gly Arg Ala Gln Gln Ala Gln Ser Arg Ala Tyr
115 120 125
Arg Pro Thr Leu Ser Ala Ile Thr Glu Phe Val Thr Asn Trp Thr Ile
130 135 140
Val Pro Phe Ala His Pro Gly Trp Ala Lys Ala Val Phe Pro Gly Leu
145 150 155 160
Pro Glu Glu Glu Ala Val Gln Arg Leu Trp Gln Ala Ile Phe Gln Ala
165 170 175
Thr Arg Val Asp Gln Glu Asp Pro Val Ala Ala Trp Glu Ala His Asn
180 185 190
Arg Val Leu His Ala Lys Val Ala Phe Leu Asn Glu Lys Arg Phe His
195 200 205
Ala Leu His Phe Gln Gly Pro Gly Thr Asp Leu Thr Val Gly Leu Ala
210 215 220
Glu Gly His Leu Trp Gln Gly Gly Ala Thr Pro Thr Lys Lys Gly Arg
225 230 235 240
Leu Cys Asn Pro Asn Leu Pro Thr Glu Glu Val Phe Thr Ala Pro His
245 250 255
Arg Glu Arg Val Glu Gly Val Val Arg Ala Ser Arg Pro Leu Ala Leu
260 265 270
Ser Gly Gln Leu Val Glu Gly Leu Trp Ala Arg Phe Glu Gly Gly Val
275 280 285
Ala Val Glu Val Gly Ala Glu Lys Gly Glu Glu Val Leu Lys Lys Leu
290 295 300
Leu Asp Thr Asp Glu Gly Ala Arg Arg Leu Gly Glu Val Ala Leu Val
305 310 315 320
Pro Ala Asp Asn Pro Ile Ala Lys Thr Gly Leu Val Phe Phe Asp Thr
325 330 335
Leu Phe Asp Glu Asn Ala Ala Ser His Ile Ala Phe Gly Gln Ala Tyr
340 345 350
Ala Glu Asn Leu Glu Gly Arg Pro Ser Gly Glu Glu Phe Arg Arg Arg
355 360 365
Gly Gly Asn Glu Ser Met Val His Val Asp Trp Met Ile Gly Ser Glu
370 375 380
Glu Val Asp Val Asp Gly Leu Leu Glu Asp Gly Thr Arg Val Pro Leu
385 390 395 400
Met Arg Arg Gly Arg Trp Val Ile
405
<210> 61
<211> 362
<212> PRT
<213> 脂肪嗜热芽孢杆菌
<400> 61
Met Ala Lys Leu Asp Glu Thr Leu Thr Met Leu Lys Ala Leu Thr Asp
1 5 10 15
Ala Lys Gly Val Pro Gly Asn Glu Arg Glu Ala Arg Asp Val Met Lys
20 25 30
Thr Tyr Ile Ala Pro Tyr Ala Asp Glu Val Thr Thr Asp Gly Leu Gly
35 40 45
Ser Leu Ile Ala Lys Lys Glu Gly Lys Ser Gly Gly Pro Lys Val Met
50 55 60
Ile Ala Gly His Leu Asp Glu Val Gly Phe Met Val Thr Gln Ile Asp
65 70 75 80
Asp Lys Gly Phe Ile Arg Phe Gln Thr Leu Gly Gly Trp Trp Ser Gln
85 90 95
Val Met Leu Ala Gln Arg Val Thr Ile Val Thr Lys Lys Gly Asp Ile
100 105 110
Thr Gly Val Ile Gly Ser Lys Pro Pro His Ile Leu Pro Ser Glu Ala
115 120 125
Arg Lys Lys Pro Val Glu Ile Lys Asp Met Phe Ile Asp Ile Gly Ala
130 135 140
Thr Ser Arg Glu Glu Ala Met Glu Trp Gly Val Arg Pro Gly Asp Met
145 150 155 160
Ile Val Pro Tyr Phe Glu Phe Thr Val Leu Asn Asn Glu Lys Met Leu
165 170 175
Leu Ala Lys Ala Trp Asp Asn Arg Ile Gly Cys Ala Val Ala Ile Asp
180 185 190
Val Leu Lys Gln Leu Lys Gly Val Asp His Pro Asn Thr Val Tyr Gly
195 200 205
Val Gly Thr Val Gln Glu Glu Val Gly Leu Arg Gly Ala Arg Thr Ala
210 215 220
Ala Gln Phe Ile Gln Pro Asp Ile Ala Phe Ala Val Asp Val Gly Ile
225 230 235 240
Ala Gly Asp Thr Pro Gly Val Ser Glu Lys Glu Ala Met Gly Lys Leu
245 250 255
Gly Ala Gly Pro His Ile Val Leu Tyr Asp Ala Thr Met Val Ser His
260 265 270
Arg Gly Leu Arg Glu Phe Val Ile Glu Val Ala Glu Glu Leu Asn Ile
275 280 285
Pro His His Phe Asp Ala Met Pro Gly Val Gly Thr Asp Ala Gly Ala
290 295 300
Ile His Leu Thr Gly Ile Gly Val Pro Ser Leu Thr Ile Ala Ile Pro
305 310 315 320
Thr Arg Tyr Ile His Ser His Ala Ala Ile Leu His Arg Asp Asp Tyr
325 330 335
Glu Asn Thr Val Lys Leu Leu Val Glu Val Ile Lys Arg Leu Asp Ala
340 345 350
Asp Lys Val Lys Gln Leu Thr Phe Asp Glu
355 360
<210> 62
<211> 484
<212> PRT
<213> 霍乱弧菌
<400> 62
Met Glu Asp Lys Val Trp Ile Ser Met Gly Ala Asp Ala Val Gly Ser
1 5 10 15
Leu Asn Pro Ala Leu Ser Glu Ser Leu Leu Pro His Ser Phe Ala Ser
20 25 30
Gly Ser Gln Val Trp Ile Gly Glu Val Ala Ile Asp Glu Leu Ala Glu
35 40 45
Leu Ser His Thr Met His Glu Gln His Asn Arg Cys Gly Gly Tyr Met
50 55 60
Val His Thr Ser Ala Gln Gly Ala Met Ala Ala Leu Met Met Pro Glu
65 70 75 80
Ser Ile Ala Asn Phe Thr Ile Pro Ala Pro Ser Gln Gln Asp Leu Val
85 90 95
Asn Ala Trp Leu Pro Gln Val Ser Ala Asp Gln Ile Thr Asn Thr Ile
100 105 110
Arg Ala Leu Ser Ser Phe Asn Asn Arg Phe Tyr Thr Thr Thr Ser Gly
115 120 125
Ala Gln Ala Ser Asp Trp Leu Ala Asn Glu Trp Arg Ser Leu Ile Ser
130 135 140
Ser Leu Pro Gly Ser Arg Ile Glu Gln Ile Lys His Ser Gly Tyr Asn
145 150 155 160
Gln Lys Ser Val Val Leu Thr Ile Gln Gly Ser Glu Lys Pro Asp Glu
165 170 175
Trp Val Ile Val Gly Gly His Leu Asp Ser Thr Leu Gly Ser His Thr
180 185 190
Asn Glu Gln Ser Ile Ala Pro Gly Ala Asp Asp Asp Ala Ser Gly Ile
195 200 205
Ala Ser Leu Ser Glu Ile Ile Arg Val Leu Arg Asp Asn Asn Phe Arg
210 215 220
Pro Lys Arg Ser Val Ala Leu Met Ala Tyr Ala Ala Glu Glu Val Gly
225 230 235 240
Leu Arg Gly Ser Gln Asp Leu Ala Asn Gln Tyr Lys Ala Gln Gly Lys
245 250 255
Lys Val Val Ser Val Leu Gln Leu Asp Met Thr Asn Tyr Arg Gly Ser
260 265 270
Ala Glu Asp Ile Val Phe Ile Thr Asp Tyr Thr Asp Ser Asn Leu Thr
275 280 285
Gln Phe Leu Thr Thr Leu Ile Asp Glu Tyr Leu Pro Glu Leu Thr Tyr
290 295 300
Gly Tyr Asp Arg Cys Gly Tyr Ala Cys Ser Asp His Ala Ser Trp His
305 310 315 320
Lys Ala Gly Phe Ser Ala Ala Met Pro Phe Glu Ser Lys Phe Lys Asp
325 330 335
Tyr Asn Pro Lys Ile His Thr Ser Gln Asp Thr Leu Ala Asn Ser Asp
340 345 350
Pro Thr Gly Asn His Ala Val Lys Phe Thr Lys Leu Gly Leu Ala Tyr
355 360 365
Val Ile Glu Met Ala Asn Ala Gly Ser Ser Gln Val Pro Asp Asp Ser
370 375 380
Val Leu Gln Asp Gly Thr Ala Lys Ile Asn Leu Ser Gly Ala Arg Gly
385 390 395 400
Thr Gln Lys Arg Phe Thr Phe Glu Leu Ser Gln Ser Lys Pro Leu Thr
405 410 415
Ile Gln Thr Tyr Gly Gly Ser Gly Asp Val Asp Leu Tyr Val Lys Tyr
420 425 430
Gly Ser Ala Pro Ser Lys Ser Asn Trp Asp Cys Arg Pro Tyr Gln Asn
435 440 445
Gly Asn Arg Glu Thr Cys Ser Phe Asn Asn Ala Gln Pro Gly Ile Tyr
450 455 460
His Val Met Leu Asp Gly Tyr Thr Asn Tyr Asn Asp Val Ala Leu Lys
465 470 475 480
Ala Ser Thr Gln
<210> 63
<211> 488
<212> PRT
<213> 耐盐发光杆菌
<400> 63
Met Glu Asp Lys Val Trp Ile Ser Ile Gly Ser Asp Ala Ser Gln Thr
1 5 10 15
Val Lys Ser Val Met Gln Ser Asn Ala Arg Ser Leu Leu Pro Glu Ser
20 25 30
Leu Ala Ser Asn Gly Pro Val Trp Val Gly Gln Val Asp Tyr Ser Gln
35 40 45
Leu Ala Glu Leu Ser His His Met His Glu Asp His Gln Arg Cys Gly
50 55 60
Gly Tyr Met Val His Ser Ser Pro Glu Ser Ala Ile Ala Ala Ser Asn
65 70 75 80
Met Pro Gln Ser Leu Val Ala Phe Ser Ile Pro Glu Ile Ser Gln Gln
85 90 95
Asp Thr Val Asn Ala Trp Leu Pro Gln Val Asn Ser Gln Ala Ile Thr
100 105 110
Gly Thr Ile Thr Ser Leu Thr Ser Phe Ile Asn Arg Phe Tyr Thr Thr
115 120 125
Thr Ser Gly Ala Gln Ala Ser Asp Trp Leu Ala Asn Glu Trp Arg Ser
130 135 140
Leu Ser Ala Ser Leu Pro Asn Ala Ser Val Arg Gln Val Ser His Phe
145 150 155 160
Gly Tyr Asn Gln Lys Ser Val Val Leu Thr Ile Thr Gly Ser Glu Lys
165 170 175
Pro Asp Glu Trp Ile Val Leu Gly Gly His Leu Asp Ser Thr Ile Gly
180 185 190
Ser His Thr Asn Glu Gln Ser Val Ala Pro Gly Ala Asp Asp Asp Ala
195 200 205
Ser Gly Ile Ala Ser Val Thr Glu Ile Ile Arg Val Leu Ser Glu Asn
210 215 220
Asn Phe Gln Pro Lys Arg Ser Ile Ala Phe Met Ala Tyr Ala Ala Glu
225 230 235 240
Glu Val Gly Leu Arg Gly Ser Gln Asp Leu Ala Asn Gln Tyr Lys Ala
245 250 255
Glu Gly Lys Gln Val Ile Ser Ala Leu Gln Leu Asp Met Thr Asn Tyr
260 265 270
Lys Gly Ser Val Glu Asp Ile Val Phe Ile Thr Asp Tyr Thr Asp Ser
275 280 285
Asn Leu Thr Thr Phe Leu Ser Gln Leu Val Asp Glu Tyr Leu Pro Ser
290 295 300
Leu Thr Tyr Gly Phe Asp Thr Cys Gly Tyr Ala Cys Ser Asp His Ala
305 310 315 320
Ser Trp His Lys Ala Gly Phe Ser Ala Ala Met Pro Phe Glu Ala Lys
325 330 335
Phe Asn Asp Tyr Asn Pro Met Ile His Thr Pro Asn Asp Thr Leu Gln
340 345 350
Asn Ser Asp Pro Thr Ala Ser His Ala Val Lys Phe Thr Lys Leu Gly
355 360 365
Leu Ala Tyr Ala Ile Glu Met Ala Ser Thr Thr Gly Gly Thr Pro Pro
370 375 380
Pro Thr Gly Asn Val Leu Lys Asp Gly Val Pro Val Asn Gly Leu Ser
385 390 395 400
Gly Ala Thr Gly Ser Gln Val His Tyr Ser Phe Glu Leu Pro Ala Gln
405 410 415
Lys Asn Leu Gln Ile Ser Thr Ala Gly Gly Ser Gly Asp Val Asp Leu
420 425 430
Tyr Val Ser Phe Gly Ser Glu Ala Thr Lys Gln Asn Trp Asp Cys Arg
435 440 445
Pro Tyr Arg Asn Gly Asn Asn Glu Val Cys Thr Phe Ala Gly Ala Thr
450 455 460
Pro Gly Thr Tyr Ser Ile Met Leu Asp Gly Tyr Arg Gln Phe Ser Gly
465 470 475 480
Val Thr Leu Lys Ala Ser Thr Gln
485
<210> 64
<211> 871
<212> PRT
<213> 鼠疫耶氏杆菌
<400> 64
Met Thr Gln Gln Pro Gln Ala Lys Tyr Arg His Asp Tyr Arg Ala Pro
1 5 10 15
Asp Tyr Thr Ile Thr Asp Ile Asp Leu Asp Phe Ala Leu Asp Ala Gln
20 25 30
Lys Thr Thr Val Thr Ala Val Ser Lys Val Lys Arg Gln Gly Thr Asp
35 40 45
Val Thr Pro Leu Ile Leu Asn Gly Glu Asp Leu Thr Leu Ile Ser Val
50 55 60
Ser Val Asp Gly Gln Ala Trp Pro His Tyr Arg Gln Gln Asp Asn Thr
65 70 75 80
Leu Val Ile Glu Gln Leu Pro Ala Asp Phe Thr Leu Thr Ile Val Asn
85 90 95
Asp Ile His Pro Ala Thr Asn Ser Ala Leu Glu Gly Leu Tyr Leu Ser
100 105 110
Gly Glu Ala Leu Cys Thr Gln Cys Glu Ala Glu Gly Phe Arg His Ile
115 120 125
Thr Tyr Tyr Leu Asp Arg Pro Asp Val Leu Ala Arg Phe Thr Thr Arg
130 135 140
Ile Val Ala Asp Lys Ser Arg Tyr Pro Tyr Leu Leu Ser Asn Gly Asn
145 150 155 160
Arg Val Gly Gln Gly Glu Leu Asp Asp Gly Arg His Trp Val Lys Trp
165 170 175
Glu Asp Pro Phe Pro Lys Pro Ser Tyr Leu Phe Ala Leu Val Ala Gly
180 185 190
Asp Phe Asp Val Leu Gln Asp Lys Phe Ile Thr Arg Ser Gly Arg Glu
195 200 205
Val Ala Leu Glu Ile Phe Val Asp Arg Gly Asn Leu Asp Arg Ala Asp
210 215 220
Trp Ala Met Thr Ser Leu Lys Asn Ser Met Lys Trp Asp Glu Thr Arg
225 230 235 240
Phe Gly Leu Glu Tyr Asp Leu Asp Ile Tyr Met Ile Val Ala Val Asp
245 250 255
Phe Phe Asn Met Gly Ala Met Glu Asn Lys Gly Leu Asn Val Phe Asn
260 265 270
Ser Lys Tyr Val Leu Ala Lys Ala Glu Thr Ala Thr Asp Lys Asp Tyr
275 280 285
Leu Asn Ile Glu Ala Val Ile Gly His Glu Tyr Phe His Asn Trp Thr
290 295 300
Gly Asn Arg Val Thr Cys Arg Asp Trp Phe Gln Leu Ser Leu Lys Glu
305 310 315 320
Gly Leu Thr Val Phe Arg Asp Gln Glu Phe Ser Ser Asp Leu Gly Ser
325 330 335
Arg Ser Val Asn Arg Ile Glu Asn Val Arg Val Met Arg Ala Ala Gln
340 345 350
Phe Ala Glu Asp Ala Ser Pro Met Ala His Ala Ile Arg Pro Asp Lys
355 360 365
Val Ile Glu Met Asn Asn Phe Tyr Thr Leu Thr Val Tyr Glu Lys Gly
370 375 380
Ser Glu Val Ile Arg Met Met His Thr Leu Leu Gly Glu Gln Gln Phe
385 390 395 400
Gln Ala Gly Met Arg Leu Tyr Phe Glu Arg His Asp Gly Ser Ala Ala
405 410 415
Thr Cys Asp Asp Phe Val Gln Ala Met Glu Asp Val Ser Asn Val Asp
420 425 430
Leu Ser Leu Phe Arg Arg Trp Tyr Ser Gln Ser Gly Thr Pro Leu Leu
435 440 445
Thr Val His Asp Asp Tyr Asp Val Glu Lys Gln Gln Tyr His Leu Phe
450 455 460
Val Ser Gln Lys Thr Leu Pro Thr Ala Asp Gln Pro Glu Lys Leu Pro
465 470 475 480
Leu His Ile Pro Leu Asp Ile Glu Leu Tyr Asp Ser Lys Gly Asn Val
485 490 495
Ile Pro Leu Gln His Asn Gly Leu Pro Val His His Val Leu Asn Val
500 505 510
Thr Glu Ala Glu Gln Thr Phe Thr Phe Asp Asn Val Ala Gln Lys Pro
515 520 525
Ile Pro Ser Leu Leu Arg Glu Phe Ser Ala Pro Val Lys Leu Asp Tyr
530 535 540
Pro Tyr Ser Asp Gln Gln Leu Thr Phe Leu Met Gln His Ala Arg Asn
545 550 555 560
Glu Phe Ser Arg Trp Asp Ala Ala Gln Ser Leu Leu Ala Thr Tyr Ile
565 570 575
Lys Leu Asn Val Ala Lys Tyr Gln Gln Gln Gln Pro Leu Ser Leu Pro
580 585 590
Ala His Val Ala Asp Ala Phe Arg Ala Ile Leu Leu Asp Glu His Leu
595 600 605
Asp Pro Ala Leu Ala Ala Gln Ile Leu Thr Leu Pro Ser Glu Asn Glu
610 615 620
Met Ala Glu Leu Phe Thr Thr Ile Asp Pro Gln Ala Ile Ser Thr Val
625 630 635 640
His Glu Ala Ile Thr Arg Cys Leu Ala Gln Glu Leu Ser Asp Glu Leu
645 650 655
Leu Ala Val Tyr Val Ala Asn Met Thr Pro Val Tyr Arg Ile Glu His
660 665 670
Gly Asp Ile Ala Lys Arg Ala Leu Arg Asn Thr Cys Leu Asn Tyr Leu
675 680 685
Ala Phe Gly Asp Glu Glu Phe Ala Asn Lys Leu Val Ser Leu Gln Tyr
690 695 700
His Gln Ala Asp Asn Met Thr Asp Ser Leu Ala Ala Leu Ala Ala Ala
705 710 715 720
Val Ala Ala Gln Leu Pro Cys Arg Asp Glu Leu Leu Ala Ala Phe Asp
725 730 735
Val Arg Trp Asn His Asp Gly Leu Val Met Asp Lys Trp Phe Ala Leu
740 745 750
Gln Ala Thr Ser Pro Ala Ala Asn Val Leu Val Gln Val Arg Thr Leu
755 760 765
Leu Lys His Pro Ala Phe Ser Leu Ser Asn Pro Asn Arg Thr Arg Ser
770 775 780
Leu Ile Gly Ser Phe Ala Ser Gly Asn Pro Ala Ala Phe His Ala Ala
785 790 795 800
Asp Gly Ser Gly Tyr Gln Phe Leu Val Glu Ile Leu Ser Asp Leu Asn
805 810 815
Thr Arg Asn Pro Gln Val Ala Ala Arg Leu Ile Glu Pro Leu Ile Arg
820 825 830
Leu Lys Arg Tyr Asp Ala Gly Arg Gln Ala Leu Met Arg Lys Ala Leu
835 840 845
Glu Gln Leu Lys Thr Leu Asp Asn Leu Ser Gly Asp Leu Tyr Glu Lys
850 855 860
Ile Thr Lys Ala Leu Ala Ala
865 870
<210> 65
<211> 483
<212> PRT
<213> 鳗弧菌
<400> 65
Met Glu Glu Lys Val Trp Ile Ser Ile Gly Gly Asp Ala Thr Gln Thr
1 5 10 15
Ala Leu Arg Ser Gly Ala Gln Ser Leu Leu Pro Glu Asn Leu Ile Asn
20 25 30
Gln Thr Ser Val Trp Val Gly Gln Val Pro Val Ser Glu Leu Ala Thr
35 40 45
Leu Ser His Glu Met His Glu Asn His Gln Arg Cys Gly Gly Tyr Met
50 55 60
Val His Pro Ser Ala Gln Ser Ala Met Ser Val Ser Ala Met Pro Leu
65 70 75 80
Asn Leu Asn Ala Phe Ser Ala Pro Glu Ile Thr Gln Gln Thr Thr Val
85 90 95
Asn Ala Trp Leu Pro Ser Val Ser Ala Gln Gln Ile Thr Ser Thr Ile
100 105 110
Thr Thr Leu Thr Gln Phe Lys Asn Arg Phe Tyr Thr Thr Ser Thr Gly
115 120 125
Ala Gln Ala Ser Asn Trp Ile Ala Asp His Trp Arg Ser Leu Ser Ala
130 135 140
Ser Leu Pro Ala Ser Lys Val Glu Gln Ile Thr His Ser Gly Tyr Asn
145 150 155 160
Gln Lys Ser Val Met Leu Thr Ile Thr Gly Ser Glu Lys Pro Asp Glu
165 170 175
Trp Val Val Ile Gly Gly His Leu Asp Ser Thr Leu Gly Ser Arg Thr
180 185 190
Asn Glu Ser Ser Ile Ala Pro Gly Ala Asp Asp Asp Ala Ser Gly Ile
195 200 205
Ala Gly Val Thr Glu Ile Ile Arg Leu Leu Ser Glu Gln Asn Phe Arg
210 215 220
Pro Lys Arg Ser Ile Ala Phe Met Ala Tyr Ala Ala Glu Glu Val Gly
225 230 235 240
Leu Arg Gly Ser Gln Asp Leu Ala Asn Arg Phe Lys Ala Glu Gly Lys
245 250 255
Lys Val Met Ser Val Met Gln Leu Asp Met Thr Asn Tyr Gln Gly Ser
260 265 270
Arg Glu Asp Ile Val Phe Ile Thr Asp Tyr Thr Asp Ser Asn Phe Thr
275 280 285
Gln Tyr Leu Thr Gln Leu Leu Asp Glu Tyr Leu Pro Ser Leu Thr Tyr
290 295 300
Gly Phe Asp Thr Cys Gly Tyr Ala Cys Ser Asp His Ala Ser Trp His
305 310 315 320
Ala Val Gly Tyr Pro Ala Ala Met Pro Phe Glu Ser Lys Phe Asn Asp
325 330 335
Tyr Asn Pro Asn Ile His Ser Pro Gln Asp Thr Leu Gln Asn Ser Asp
340 345 350
Pro Thr Gly Phe His Ala Val Lys Phe Thr Lys Leu Gly Leu Ala Tyr
355 360 365
Val Val Glu Met Gly Asn Ala Ser Thr Pro Pro Thr Pro Ser Asn Gln
370 375 380
Leu Lys Asn Gly Val Pro Val Asn Gly Leu Ser Ala Ser Arg Asn Ser
385 390 395 400
Lys Thr Trp Tyr Gln Phe Glu Leu Gln Glu Ala Gly Asn Leu Ser Ile
405 410 415
Val Leu Ser Gly Gly Ser Gly Asp Ala Asp Leu Tyr Val Lys Tyr Gln
420 425 430
Thr Asp Ala Asp Leu Gln Gln Tyr Asp Cys Arg Pro Tyr Arg Ser Gly
435 440 445
Asn Asn Glu Thr Cys Gln Phe Ser Asn Ala Gln Pro Gly Arg Tyr Ser
450 455 460
Ile Leu Leu His Gly Tyr Asn Asn Tyr Ser Asn Ala Ser Leu Val Ala
465 470 475 480
Asn Ala Gln
<210> 66
<211> 482
<212> PRT
<213> 未知
<220>
<223> 盐弧菌spYCSC6
<400> 66
Met Glu Asp Lys Lys Val Trp Ile Ser Ile Gly Ala Asp Ala Gln Gln
1 5 10 15
Thr Ala Leu Ser Ser Gly Ala Gln Pro Leu Leu Ala Gln Ser Val Ala
20 25 30
His Asn Gly Gln Ala Trp Ile Gly Glu Val Ser Glu Ser Glu Leu Ala
35 40 45
Ala Leu Ser His Glu Met His Glu Asn His His Arg Cys Gly Gly Tyr
50 55 60
Ile Val His Ser Ser Ala Gln Ser Ala Met Ala Ala Ser Asn Met Pro
65 70 75 80
Leu Ser Arg Ala Ser Phe Ile Ala Pro Ala Ile Ser Gln Gln Ala Leu
85 90 95
Val Thr Pro Trp Ile Ser Gln Ile Asp Ser Ala Leu Ile Val Asn Thr
100 105 110
Ile Asp Arg Leu Thr Asp Phe Pro Asn Arg Phe Tyr Thr Thr Thr Ser
115 120 125
Gly Ala Gln Ala Ser Asp Trp Ile Lys Gln Arg Trp Gln Ser Leu Ser
130 135 140
Ala Gly Leu Ala Gly Ala Ser Val Thr Gln Ile Ser His Ser Gly Tyr
145 150 155 160
Asn Gln Ala Ser Val Met Leu Thr Ile Glu Gly Ser Glu Ser Pro Asp
165 170 175
Glu Trp Val Val Val Gly Gly His Leu Asp Ser Thr Ile Gly Ser Arg
180 185 190
Thr Asn Glu Gln Ser Ile Ala Pro Gly Ala Asp Asp Asp Ala Ser Gly
195 200 205
Ile Ala Ala Val Thr Glu Val Ile Arg Val Leu Ala Gln Asn Asn Phe
210 215 220
Gln Pro Lys Arg Ser Ile Ala Phe Val Ala Tyr Ala Ala Glu Glu Val
225 230 235 240
Gly Leu Arg Gly Ser Gln Asp Val Ala Asn Gln Phe Lys Gln Ala Gly
245 250 255
Lys Asp Val Arg Gly Val Leu Gln Leu Asp Met Thr Asn Tyr Gln Gly
260 265 270
Ser Ala Glu Asp Ile Val Phe Ile Thr Asp Tyr Thr Asp Asn Gln Leu
275 280 285
Thr Gln Tyr Leu Thr Gln Leu Leu Asp Glu Tyr Leu Pro Thr Leu Asn
290 295 300
Tyr Gly Phe Asp Thr Cys Gly Tyr Ala Cys Ser Asp His Ala Ser Trp
305 310 315 320
His Gln Val Gly Tyr Pro Ala Ala Met Pro Phe Glu Ala Lys Phe Asn
325 330 335
Asp Tyr Asn Pro Asn Ile His Thr Pro Gln Asp Thr Leu Ala Asn Ser
340 345 350
Asp Ser Glu Gly Ala His Ala Ala Lys Phe Thr Lys Leu Gly Leu Ala
355 360 365
Tyr Thr Val Glu Leu Ala Asn Ala Asp Ser Ser Pro Asn Pro Gly Asn
370 375 380
Glu Leu Lys Leu Gly Glu Pro Ile Asn Gly Leu Ser Gly Ala Arg Gly
385 390 395 400
Asn Glu Lys Tyr Phe Asn Tyr Arg Leu Asp Gln Ser Gly Glu Leu Val
405 410 415
Ile Arg Thr Tyr Gly Gly Ser Gly Asp Val Asp Leu Tyr Val Lys Ala
420 425 430
Asn Gly Asp Val Ser Thr Gly Asn Trp Asp Cys Arg Pro Tyr Arg Ser
435 440 445
Gly Asn Asp Glu Val Cys Arg Phe Asp Asn Ala Thr Pro Gly Asn Tyr
450 455 460
Ala Val Met Leu Arg Gly Tyr Arg Thr Tyr Asp Asn Val Ser Leu Ile
465 470 475 480
Val Glu
<210> 67
<211> 301
<212> PRT
<213> 解蛋白弧菌
<400> 67
Met Pro Pro Ile Thr Gln Gln Ala Thr Val Thr Ala Trp Leu Pro Gln
1 5 10 15
Val Asp Ala Ser Gln Ile Thr Gly Thr Ile Ser Ser Leu Glu Ser Phe
20 25 30
Thr Asn Arg Phe Tyr Thr Thr Thr Ser Gly Ala Gln Ala Ser Asp Trp
35 40 45
Ile Ala Ser Glu Trp Gln Ala Leu Ser Ala Ser Leu Pro Asn Ala Ser
50 55 60
Val Lys Gln Val Ser His Ser Gly Tyr Asn Gln Lys Ser Val Val Met
65 70 75 80
Thr Ile Thr Gly Ser Glu Ala Pro Asp Glu Trp Ile Val Ile Gly Gly
85 90 95
His Leu Asp Ser Thr Ile Gly Ser His Thr Asn Glu Gln Ser Val Ala
100 105 110
Pro Gly Ala Asp Asp Asp Ala Ser Gly Ile Ala Ala Val Thr Glu Val
115 120 125
Ile Arg Val Leu Ser Glu Asn Asn Phe Gln Pro Lys Arg Ser Ile Ala
130 135 140
Phe Met Ala Tyr Ala Ala Glu Glu Val Gly Leu Arg Gly Ser Gln Asp
145 150 155 160
Leu Ala Asn Gln Tyr Lys Ser Glu Gly Lys Asn Val Val Ser Ala Leu
165 170 175
Gln Leu Asp Met Thr Asn Tyr Lys Gly Ser Ala Gln Asp Val Val Phe
180 185 190
Ile Thr Asp Tyr Thr Asp Ser Asn Phe Thr Gln Tyr Leu Thr Gln Leu
195 200 205
Met Asp Glu Tyr Leu Pro Ser Leu Thr Tyr Gly Phe Asp Thr Cys Gly
210 215 220
Tyr Ala Cys Ser Asp His Ala Ser Trp His Asn Ala Gly Tyr Pro Ala
225 230 235 240
Ala Met Pro Phe Glu Ser Lys Phe Asn Asp Tyr Asn Pro Arg Ile His
245 250 255
Thr Thr Gln Asp Thr Leu Ala Asn Ser Asp Pro Thr Gly Ser His Ala
260 265 270
Lys Lys Phe Thr Gln Leu Gly Leu Ala Tyr Ala Ile Glu Met Gly Ser
275 280 285
Ala Thr Gly Asp Thr Pro Thr Pro Gly Asn Gln Leu Glu
290 295 300
<210> 68
<211> 301
<212> PRT
<213> 解蛋白弧菌
<400> 68
Met Pro Pro Ile Thr Gln Gln Ala Thr Val Thr Ala Trp Leu Pro Gln
1 5 10 15
Val Asp Ala Ser Gln Ile Thr Gly Thr Ile Ser Ser Leu Glu Ser Phe
20 25 30
Thr Asn Arg Phe Tyr Thr Thr Thr Ser Gly Ala Gln Ala Ser Asp Trp
35 40 45
Ile Ala Ser Glu Trp Gln Phe Leu Ser Ala Ser Leu Pro Asn Ala Ser
50 55 60
Val Lys Gln Val Ser His Ser Gly Tyr Asn Gln Lys Ser Val Val Met
65 70 75 80
Thr Ile Thr Gly Ser Glu Ala Pro Asp Glu Trp Ile Val Ile Gly Gly
85 90 95
His Leu Asp Ser Thr Ile Gly Ser His Thr Asn Glu Gln Ser Val Ala
100 105 110
Pro Gly Ala Asp Asp Asp Ala Ser Gly Ile Ala Ala Val Thr Glu Val
115 120 125
Ile Arg Val Leu Ser Glu Asn Asn Phe Gln Pro Lys Arg Ser Ile Ala
130 135 140
Phe Met Ala Tyr Ala Ala Glu Glu Val Gly Leu Arg Gly Ser Gln Asp
145 150 155 160
Leu Ala Asn Gln Tyr Lys Ser Glu Gly Lys Asn Val Val Ser Ala Leu
165 170 175
Gln Leu Asp Met Thr Asn Tyr Lys Gly Ser Ala Gln Asp Val Val Phe
180 185 190
Ile Thr Asp Tyr Thr Asp Ser Asn Phe Thr Gln Tyr Leu Thr Gln Leu
195 200 205
Met Asp Glu Tyr Leu Pro Ser Leu Thr Tyr Gly Phe Asp Thr Cys Gly
210 215 220
Tyr Ala Cys Ser Asp His Ala Ser Trp His Asn Ala Gly Tyr Pro Ala
225 230 235 240
Ala Met Pro Phe Glu Ser Lys Phe Asn Asp Tyr Asn Pro Arg Ile His
245 250 255
Thr Thr Gln Asp Thr Leu Ala Asn Ser Asp Pro Thr Gly Ser His Ala
260 265 270
Lys Lys Phe Thr Gln Leu Gly Leu Ala Tyr Ala Ile Glu Met Gly Ser
275 280 285
Ala Thr Gly Asp Thr Pro Thr Pro Gly Asn Gln Leu Glu
290 295 300
<210> 69
<211> 348
<212> PRT
<213> 激烈火球菌
<400> 69
Met Val Asp Trp Glu Leu Met Lys Lys Ile Ile Glu Ser Pro Gly Val
1 5 10 15
Ser Gly Tyr Glu His Leu Gly Ile Arg Asp Leu Val Val Asp Ile Leu
20 25 30
Lys Asp Val Ala Asp Glu Val Lys Ile Asp Lys Leu Gly Asn Val Ile
35 40 45
Ala His Phe Lys Gly Ser Ala Pro Lys Val Met Val Ala Ala His Met
50 55 60
Asp Lys Ile Gly Leu Met Val Asn His Ile Asp Lys Asp Gly Tyr Leu
65 70 75 80
Arg Val Val Pro Ile Gly Gly Val Leu Pro Glu Thr Leu Ile Ala Gln
85 90 95
Lys Ile Arg Phe Phe Thr Glu Lys Gly Glu Arg Tyr Gly Val Val Gly
100 105 110
Val Leu Pro Pro His Leu Arg Arg Glu Ala Lys Asp Gln Gly Gly Lys
115 120 125
Ile Asp Trp Asp Ser Ile Ile Val Asp Val Gly Ala Ser Ser Arg Glu
130 135 140
Glu Ala Glu Glu Met Gly Phe Arg Ile Gly Thr Ile Gly Glu Phe Ala
145 150 155 160
Pro Asn Phe Thr Arg Leu Ser Glu His Arg Phe Ala Thr Pro Tyr Leu
165 170 175
Asp Asp Arg Ile Cys Leu Tyr Ala Met Ile Glu Ala Ala Arg Gln Leu
180 185 190
Gly Glu His Glu Ala Asp Ile Tyr Ile Val Ala Ser Val Gln Glu Glu
195 200 205
Ile Gly Leu Arg Gly Ala Arg Val Ala Ser Phe Ala Ile Asp Pro Glu
210 215 220
Val Gly Ile Ala Met Asp Val Thr Phe Ala Lys Gln Pro Asn Asp Lys
225 230 235 240
Gly Lys Ile Val Pro Glu Leu Gly Lys Gly Pro Val Met Asp Val Gly
245 250 255
Pro Asn Ile Asn Pro Lys Leu Arg Gln Phe Ala Asp Glu Val Ala Lys
260 265 270
Lys Tyr Glu Ile Pro Leu Gln Val Glu Pro Ser Pro Arg Pro Thr Gly
275 280 285
Thr Asp Ala Asn Val Met Gln Ile Asn Arg Glu Gly Val Ala Thr Ala
290 295 300
Val Leu Ser Ile Pro Ile Arg Tyr Met His Ser Gln Val Glu Leu Ala
305 310 315 320
Asp Ala Arg Asp Val Asp Asn Thr Ile Lys Leu Ala Lys Ala Leu Leu
325 330 335
Glu Glu Leu Lys Pro Met Asp Phe Thr Pro Leu Glu
340 345
<210> 70
<211> 2
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 70
Lys Phe
1
<210> 71
<211> 7
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa是叠氮赖氨酸
<400> 71
Lys Lys Met Lys Lys Met Xaa
1 5
<210> 72
<211> 7
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa是叠氮赖氨酸
<400> 72
Asp Asp Met Asp Asp Met Xaa
1 5
<210> 73
<211> 7
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa是叠氮赖氨酸
<400> 73
Phe Phe Met Phe Phe Met Xaa
1 5
<210> 74
<211> 7
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa是叠氮赖氨酸
<400> 74
Ala Ala Met Ala Ala Met Xaa
1 5
<210> 75
<211> 7
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa是叠氮赖氨酸
<400> 75
Phe Pro Phe Pro Phe Pro Xaa
1 5
<210> 76
<211> 7
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa是叠氮赖氨酸
<400> 76
Asp Pro Asp Pro Asp Pro Xaa
1 5
<210> 77
<211> 7
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa是叠氮赖氨酸
<400> 77
Lys Pro Lys Pro Lys Pro Xaa
1 5
<210> 78
<211> 13
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 78
Tyr Ala Ala Trp Ala Ala Phe Ala Asp Asp Asp Trp Lys
1 5 10
<210> 79
<211> 11
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 79
Phe Tyr Pro Leu Pro Trp Pro Asp Asp Asp Tyr
1 5 10
<210> 80
<211> 12
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 80
Phe Tyr Pro Leu Pro Trp Pro Asp Asp Asp Tyr Lys
1 5 10
<210> 81
<211> 11
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 81
Tyr Pro Leu Pro Trp Pro Asp Asp Asp Tyr Lys
1 5 10
<210> 82
<211> 7
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 82
Tyr Pro Tyr Pro Tyr Pro Lys
1 5
<210> 83
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 83
Pro Tyr Pro Tyr Pro Lys
1 5
<210> 84
<211> 4
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 84
Gly Pro Arg Pro
1
<210> 85
<211> 7
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa是叠氮赖氨酸
<400> 85
Asp Asp Pro Asp Asp Pro Xaa
1 5
<210> 86
<211> 7
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa是叠氮赖氨酸
<400> 86
Tyr Pro Tyr Pro Tyr Pro Xaa
1 5
<210> 87
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (6)..(6)
<223> Xaa是叠氮赖氨酸
<400> 87
Pro Tyr Pro Tyr Pro Xaa
1 5
<210> 88
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (6)..(6)
<223> Xaa是叠氮赖氨酸
<400> 88
Pro Phe Pro Phe Pro Xaa
1 5
<210> 89
<211> 7
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa是Lys-三唑-PEG4
<400> 89
Lys Lys Met Lys Lys Met Xaa
1 5
<210> 90
<211> 7
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa是叠氮赖氨酸
<400> 90
Ala Ala Pro Ala Ala Pro Xaa
1 5
<210> 91
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (6)..(6)
<223> Xaa是叠氮赖氨酸
<400> 91
Ala Pro Ala Ala Pro Xaa
1 5
<210> 92
<211> 7
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa是叠氮赖氨酸
<400> 92
Tyr Tyr Pro Tyr Tyr Pro Xaa
1 5
<210> 93
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (6)..(6)
<223> Xaa是叠氮赖氨酸
<400> 93
Tyr Pro Tyr Tyr Pro Xaa
1 5
<210> 94
<211> 7
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa是叠氮赖氨酸
<400> 94
Phe Phe Pro Phe Phe Pro Xaa
1 5
<210> 95
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (6)..(6)
<223> Xaa是叠氮赖氨酸
<400> 95
Phe Pro Phe Phe Pro Xaa
1 5
<210> 96
<211> 7
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa是叠氮赖氨酸
<400> 96
Arg Arg Pro Arg Arg Pro Xaa
1 5
<210> 97
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (6)..(6)
<223> Xaa是叠氮赖氨酸
<400> 97
Arg Pro Arg Arg Pro Xaa
1 5
<210> 98
<211> 7
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa是叠氮赖氨酸
<400> 98
Ala Ala Pro Ala Ala Pro Xaa
1 5
<210> 99
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (6)..(6)
<223> Xaa是叠氮赖氨酸
<400> 99
Ala Pro Ala Ala Pro Xaa
1 5
<210> 100
<211> 7
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa是叠氮赖氨酸
<400> 100
Lys Lys Pro Lys Lys Pro Xaa
1 5
<210> 101
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (6)..(6)
<223> Xaa是叠氮赖氨酸
<400> 101
Lys Pro Lys Lys Pro Xaa
1 5
<210> 102
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (6)..(6)
<223> Xaa是叠氮赖氨酸
<400> 102
Lys Met Lys Lys Met Xaa
1 5
<210> 103
<211> 7
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (7)..(7)
<223> Xaa是叠氮赖氨酸
<400> 103
Tyr Tyr Met Tyr Tyr Met Xaa
1 5
<210> 104
<211> 8
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (8)..(8)
<223> Xaa是叠氮赖氨酸
<400> 104
Asp Pro Ala Ala Ala Phe Lys Xaa
1 5
<210> 105
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 105
Pro Ala Ala Ala Phe Lys
1 5
<210> 106
<211> 10
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (10)..(10)
<223> Xaa是叠氮赖氨酸
<400> 106
Lys Ala Ala Ala Ala Ala Ala Phe Lys Xaa
1 5 10
<210> 107
<211> 12
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (12)..(12)
<223> Xaa是叠氮赖氨酸
<400> 107
Phe Tyr Pro Leu Pro Trp Pro Asp Asp Asp Tyr Xaa
1 5 10
<210> 108
<211> 11
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (11)..(11)
<223> Xaa是叠氮赖氨酸
<400> 108
Tyr Pro Leu Pro Trp Pro Asp Asp Asp Tyr Xaa
1 5 10
<210> 109
<211> 10
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (10)..(10)
<223> Xaa是叠氮赖氨酸
<400> 109
Pro Leu Pro Trp Pro Asp Asp Asp Tyr Xaa
1 5 10
<210> 110
<211> 9
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<220>
<221> misc_feature
<222> (9)..(9)
<223> Xaa是叠氮赖氨酸
<400> 110
Leu Pro Trp Pro Asp Asp Asp Tyr Xaa
1 5
<210> 111
<211> 10
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 111
Phe Ala Ala Ala Trp Pro Asp Asp Asp Phe
1 5 10
<210> 112
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 112
Trp Pro Asp Asp Phe
1 5
<210> 113
<211> 10
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 113
Trp Ala Ala Ala Phe Pro Asp Asp Asp Phe
1 5 10
<210> 114
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 114
Phe Pro Asp Asp Phe
1 5
<210> 115
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 115
Tyr Pro Asp Asp Phe
1 5
<210> 116
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 116
Arg Arg Pro Phe Gln Gln
1 5
<210> 117
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 117
Arg Pro Phe Gln Gln
1 5
<210> 118
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 118
Ala Ala Pro Phe Gln Gln
1 5
<210> 119
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 119
Ala Pro Phe Gln Gln
1 5
<210> 120
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 120
Lys Lys Pro Phe Gln Gln
1 5
<210> 121
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 121
Lys Pro Phe Gln Gln
1 5
<210> 122
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 122
Tyr Tyr Pro Phe Gln Gln
1 5
<210> 123
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 123
Tyr Pro Phe Gln Gln
1 5
<210> 124
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 124
Phe Phe Pro Phe Gln Gln
1 5
<210> 125
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 125
Phe Pro Phe Gln Gln
1 5
<210> 126
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 126
Asp Asp Pro Phe Gln Gln
1 5
<210> 127
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 127
Asp Pro Phe Gln Gln
1 5
<210> 128
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 128
Glu Glu Pro Phe Gln Gln
1 5
<210> 129
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 129
Glu Pro Phe Gln Gln
1 5
<210> 130
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 130
Asn Asn Pro Phe Gln Gln
1 5
<210> 131
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 131
Asn Pro Phe Gln Gln
1 5
<210> 132
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 132
Gln Gln Pro Phe Gln Gln
1 5
<210> 133
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 133
Gln Pro Phe Gln Gln
1 5
<210> 134
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 134
Val Val Pro Phe Gln Gln
1 5
<210> 135
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 135
Val Pro Phe Gln Gln
1 5
<210> 136
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 136
Ile Ile Pro Phe Gln Gln
1 5
<210> 137
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 137
Ile Pro Phe Gln Gln
1 5
<210> 138
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 138
Leu Leu Pro Phe Gln Gln
1 5
<210> 139
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 139
Leu Pro Phe Gln Gln
1 5
<210> 140
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 140
Ser Ser Pro Phe Gln Gln
1 5
<210> 141
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 141
Ser Pro Phe Gln Gln
1 5
<210> 142
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 142
Thr Thr Pro Phe Gln Gln
1 5
<210> 143
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 143
Thr Pro Phe Gln Gln
1 5
<210> 144
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 144
Cys Cys Pro Phe Gln Gln
1 5
<210> 145
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 145
Cys Pro Phe Gln Gln
1 5
<210> 146
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 146
Trp Trp Pro Phe Gln Gln
1 5
<210> 147
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 147
Trp Pro Phe Gln Gln
1 5
<210> 148
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 148
Met Met Pro Phe Gln Gln
1 5
<210> 149
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 149
Met Pro Phe Gln Gln
1 5
<210> 150
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 150
Pro Pro Pro Phe Gln Gln
1 5
<210> 151
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 151
Pro Pro Phe Gln Gln
1 5
<210> 152
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 152
Gly Gly Pro Phe Gln Gln
1 5
<210> 153
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 153
Gly Pro Phe Gln Gln
1 5
<210> 154
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 154
His His Pro Phe Gln Gln
1 5
<210> 155
<211> 5
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 155
His Pro Phe Gln Gln
1 5
<210> 156
<211> 12
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 156
Tyr Ala Ala Phe Ala Ala Trp Ala Asp Asp Asp Trp
1 5 10
<210> 157
<211> 6
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 157
Ala Asp Asp Asp Trp Lys
1 5
<210> 158
<211> 7
<212> PRT
<213> 人工序列
<220>
<223> 合成多肽
<400> 158
Trp Ala Asp Asp Asp Trp Lys
1 5
- 用于蛋白质测序的方法及组合物
- 特别适用于有机食品业的去矿物质化乳蛋白质组合物的生产方法以及去矿物质化乳蛋白质组合物