一种双环吉玛烯合成酶突变体和基因序列以及应用
文献发布时间:2023-06-19 19:27:02
技术领域
本发明属于酶基因工程及酶工程领域,具体涉及一种来源于马鞭草(Lippiadulcis)的双环吉玛烯合成酶LdTPS5突变体及其编码基因以及应用。
背景技术
双环吉玛烯是一种重要的双环倍半萜,是雄性激素(androgen)、雌性激素(estrogen)、孕酮(progesterone)和多巴胺(dopamine)等人体激素的潜在受体,有治疗内分泌疾病的潜力。双环吉玛烯可能是具有抑菌活性的香橙烯、白千层醇、蓝桉醇、喇叭醇和喇叭茶醇(palustrol)等香橙烯型倍半萜合成的重要中间体,也可能是一些具有抑菌活性的杂环萜如psiguadials A和macrocarpal A的重要合成骨架。目前,双环吉玛烯合成酶的催化效率以及特异性催化限制了双环吉玛烯的利用,因此通过酶工程手段对双环吉玛烯合成酶进行改造以提高双环吉玛烯产量具有重要的生产意义和应用价值。
发明内容
本发明的目的在于,提供一种双环吉玛烯合成酶LdTPS5突变体及其基因序列,以解决双环吉玛烯合成酶的催化效率低以及产物特异性差导致的双环吉玛烯产量低的问题。
为了解决上述技术问题,本发明采用以下技术方案:
根据本发明的第一方面,提供一种双环吉玛烯合成酶突变体,所述双环吉玛烯合成酶突变体包括以SEQ ID NO.1所示野生型的双环吉玛烯合成酶核苷酸序列为模板,分别进行四个位点突变,获得四个突变体,所述四个突变体为:第290位异亮氨酸突变为缬氨酸形成的突变体I290V,其氨基酸序列如SEQ ID NO.2所示;第316位异亮氨酸突变为亮氨酸形成的突变体I316L,其氨基酸序列如SEQ IDNO.3所示;第454位亮氨酸突变为半胱氨酸形成的突变体L454C,其氨基酸序列如SEQ ID NO.4所示;第290位异亮氨酸突变为缬氨酸,第316位异亮氨酸突变为亮氨酸以及第454位亮氨酸突变为半胱氨酸形成的组合突变体I290V/I316L/L454C,其氨基酸序列如SEQ ID NO.5所示。
根据本发明的第二方面,提供一种双环吉玛烯合成酶突变体的编码基因,其特征在于,所述编码基因编码如上所述的任意一种双环吉玛烯合成酶突变体的氨基酸序列。
根据本发明的第三方面,提供一种包含所述双环吉玛烯合成酶突变体的编码基因的重组表达载体。
根据本发明的一个优选实施方案,所述重组表达载体中的载体质粒为pESC-URA,启动子为Gal1诱导性启动子。但是应当理解的是,本发明并不仅限于该载体质粒和启动子,还可以是其他任何适合的载体质粒和启动子。
根据本发明的第四方面,提供一种如上所述的双环吉玛烯合成酶突变体的编码基因的重组基因工程菌。
根据本发明的第五方面,提供一种所述双环吉玛烯合酶突变体在酵母中合成双环吉玛烯中的应用。
所述应用包括:将一种包含所述双环吉玛烯合酶突变体的编码基因的重组表达载体转入酵母宿主菌中,将所述重组基因工程菌接种到尿嘧啶缺陷型选择性培养基中发酵120h,从而获得包含双环吉玛烯的产物。
在所述应用中,优选地,在24h时添加终浓度为10g/L的半乳糖进行蛋白的诱导表达,同时添加10%正十二烷进行双相发酵培养以减少双环吉玛烯的损失。
有益效果:本发明通过理性设计改造初步获得了4种双环吉玛烯合酶突变体,I290V,I316L,L454C以及组合突变体I290V/I316L/L454C,并通过基因工程技术进一步构建出基因重组酵母菌株,最终通过实验证明,相比野生型菌株,突变体菌株生产双环吉玛烯的产量分别提高了17.6,11.2,12.3和52.7倍。
综上所述,本发明利用基因工程和酶工程的手段,提供了一种双环吉玛烯合酶突变体以及在酵母中合成双环吉玛烯的应用,通过提供催化活性提高的双环吉玛烯合酶突变体,提高双环吉玛烯的产量,为双环吉玛烯的工业化生产创造了条件。
附图说明
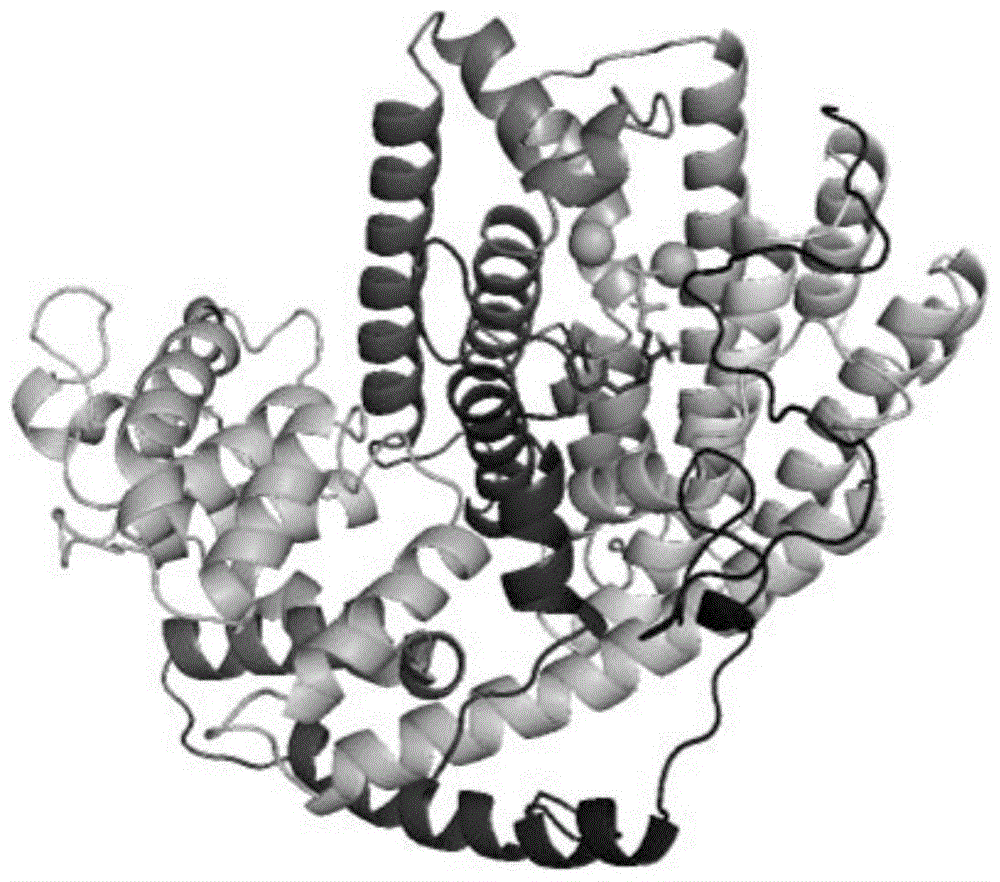
图1为双环吉玛烯合成酶LdTPS5的三维结构模型图;
图2示出了双环吉玛烯合成酶LdTPS5的基因表达载体的质粒示意图;
图3示出了基因重组酵母菌株的双环吉玛烯产量图。
具体实施方式
以下结合具体实施例,对本发明做进一步说明。应理解,以下实施例仅用于说明本发明而非用于限制本发明的范围。
下列实施例中未注明具体条件的实验方法,通常按照常规条件,如《分子克隆:实验室手册》(New York:Cold Spring Harbor Laboratory Press,1989)中所述的条件进行。引物和序列为金唯智生物科技有限公司合成。
实施例1双环吉玛烯合成酶三维结构模型的建立
利用同源建模工具,以PDB ID:3G4F为参考模型对双环吉玛烯合成酶进行同源建模。经模型评估后,获得可靠的三维结构模型,三维结构模型图如图1所示。
实施例2构建双环吉玛烯合成酶突变体
首先,委托金唯智公司合成经过密码子优化过的LdTPS5(该密码子优化后的核苷酸序列如序列SEQ ID No.1所示)。利用PCR技术,以合成的LdTPS5为模板,对LdTPS5的290,316和454位分别进行定点突变,对290/316/454位进行组合突变。获得突变体I290V,I316L和L454C及组合突变体I290V/I316L/L454C的编码基因。
表1引物序列表
I290V-F、I290V-R、LdTPS5-F和LdTPS5-R用于构建I290V突变体
I316L-F、I316L-R、LdTPS5-F和LdTPS5-R用于构建I316L突变体
L454C-F、L454C-R、LdTPS5-F和LdTPS5-R用于构建L454C突变体
I290V-F、I290V-R、I316L-F、I316L-R、L454C-F、L454C-R、LdTPS5-F和LdTPS5-R用于构建组合突变I290V/I316L/L454C突变体
实施例3:构建双环吉玛烯合酶突变体表达载体
选择表达载体pESC-URA空载质粒,使用限制性核酸内切酶BamHI和Hind III对空载质粒进行双酶切后经琼脂糖凝胶电泳验证后进行回收纯化。
然后使用诺唯赞Vazyme的同源重组酶CloneExpressⅡ对实施例2中构建的LdTPS5野生型及突变体和线性化pESC-URA空载质粒分别进行连接。混合好的连接混合物于37℃反应30min并通过大肠杆菌转化导入DH5α感受态。使用LdTPS5-F/LdTPS5-R引物PCR验证,将阳性条带对应的菌株送去DNA测序,得到阳性LdTPS5突变体重组质粒。
实施例4:表达突变体的重组酿酒酵母的构建
分别以实施例2和实施例3构建的野生型和突变体质粒作为双环吉马烯合成酶表达载体利用醋酸锂转化法导入酿酒酵母菌株。将转入质粒的重组酿酒酵母细胞涂布于SD-URA营养缺陷型筛选固体培养基平板,30℃培养箱培养2~3天,分别得到酿酒酵母双环吉马烯合成酶突变体表达菌株I290V,I316L,L454C,组合突变I290V/I316L/L454C以及双环吉马烯合成酶野生型表达菌株。
实施例5:重组酿酒酵母摇瓶发酵生产双环吉马烯
挑取实施例4中构建的重组酿酒酵母菌株进行摇瓶发酵,双环吉马烯合成酶野生型表达菌株作为对照。接单菌落于10mL SD-URA培养基中,于30℃摇床中220rpm培养16h。按初始OD
如图3所示,I290V,I316L,L454C以及组合突变体I290V/I316L/L454C突变体菌株的催化活性有所提高,双环吉马烯产量相比野生型表达菌株有明显提升,双环吉玛烯的产量分别提高了17.6,11.2,12.3和52.7倍。
以上所述的,仅为本发明的较佳实施例,并非用以限定本发明的范围,本发明的上述实施例还可以做出各种变化。凡是依据本发明申请的权利要求书及说明书内容所作的简单、等效变化与修饰,皆落入本发明专利的权利要求保护范围。本发明未详尽描述的均为常规技术内容。
- 一种7β-羟基类固醇脱氢酶突变体、编码序列、重组表达载体、基因工程菌及应用
- 一种用于制备桥式四氢双环戊二烯的催化剂及制备方法与应用
- 一种紫花苜蓿过表达鲨烯环氧酶基因的方法与应用
- 一种γ-谷酰胺甲胺合成酶的突变体,其编码基因、氨基酸序列及其应用
- 一种具有热稳定性的γ-谷酰胺甲胺合成酶的突变体,其编码基因、氨基酸序列及其应用