BZR1基因在调控植物对虫害胁迫抗性中的应用
文献发布时间:2023-06-19 13:45:04
技术领域
本发明涉及生物技术领域,具体涉及一种通过缺失BZR1基因功能来提高植物对虫害尤其是棉铃虫抗性的方法。
背景技术
番茄(Solanum lycopersicum L.)是世界上消费和种植最广泛的蔬菜之一。然而,在整个生长周期中,番茄面临各种病虫的侵袭,导致了巨大的经济损失。若大量施用杀虫剂防治,则会带来农药残留、环境污染和耐药性等问题。鉴于番茄具备生命周期相对较短、基因组较小、遗传多样性高、转基因和栽培容易等优点,加之全基因组测序完成和分子遗传学研究的发展,番茄已经成为研究植物与病虫害互作的经典模式植物。因此,以番茄为对象,探究植物抵御虫害的分子机制对于虫害控制和农产品安全生产具有积极的生态、经济意义。
棉铃虫(Helicoverpa armigera)是在中国棉区和蔬菜种植区均有发生的重要作物虫害。棉铃虫取食范围广,寄主植物多达30多科200余种,其中包括重要的园艺作物——番茄。棉铃虫在番茄种植的整个周期均可发生,其最大的危害是专蛀果实,引起果实腐变,产量损失达30%以上,已经成为发展晚茬番茄生产的一个主要障碍因素。因此,探究番茄内在防御棉铃虫的分子机制对培育抗虫新品种具有一定的指导意义。
作为固着生物,植物已经进化出大量的防御策略来对抗草食动物,以成功生存和繁殖。植物对草食动物攻击的防御反应是由植物激素(Pieterse,C.M.J.等,“SignallingNetworks Involved in Induced Resistance.”In Induced Resistance for PlantDefense,2014,58–80)介导的,其中,茉莉酸(JA)信号通路已被公认为是调控植物对草食动物防御反应的核心通路(Howe GA等,“Modularity in jasmonate signaling formultistress resilience.”Annu.Rev.Plant Biol.,2018,69,387–415)。当草食动物攻击时,JA信号激活,修饰植物的物理和化学表型,从而增加植物的抵御能力。过去20年的深入研究揭示了JA通路的重要分子组成(Howe GA等,“Modularity in jasmonate signalingfor multistress resilience.”Annu.Rev.Plant Biol.,2018,69,387–415)。在植物受到损伤时,脂酰水解酶从质体膜中的半乳糖脂中释放α-亚麻酸(18:3),由13-脂肪加氧酶氧化,然后环氧化和环化反应生成12-氧代-植二烯酸(OPDA)。OPDA被运输到过氧化物酶体中,经还原和乙酰辅酶A氧化酶(Acx1)等介导的三个β氧化循环形成(+)-7-iso-JA,通过差向异构转变为较为稳定的反式构型,即JA。进一步,JA与异亮氨酸结合形成具有活性的衍生物茉莉酸-异亮氨酸(JA-Ile)。JA-Ile结合由COI1、JAZ及肌醇聚磷酸盐辅因子组成的受体,致使抑制因子JAZ被26S蛋白酶体降解,从而释放MYC转录因子,激活下游防御基因的表达。虽然JA信号转导途径已经得到了广泛的研究,但对于JA合成途径的调控机制亟待进一步的探究。
转录调控在植物抗性形成调控网络中起到关键的作用,是研究植物产生防御反应的关键所在。转录调控是由转录因子通过结合启动子区域的顺式作用元件,调控一系列基因的表达,以抵御病虫害的侵袭。BZR1(BRASSINAZOLE RESISTANT 1)为油菜素内酯信号通路中的关键转录因子,参与植物细胞伸长、细胞分裂、开花及种子发育等多个生长发育过程,还发现其在抵御高低温、盐胁迫等非生物胁迫中发挥重要作用(Qianfeng Li等,“Thebrassinosteroid-regulated transcription factors BZR1/BES1 function as acoordinator in multisignal-regulated plant growth.”BBA-Gene RegulatoryMechanisms,2018,861,561–571;Hui Li等,“BZR1 Positively Regulates FreezingTolerance viaCBF-Dependent and CBF-Independent Pathwaysin Arabidopsis.”Molecular Plant,2017,10,545-559)。但是对于BZR1在虫害胁迫中的作用及机制仍不清楚。因此,研究番茄SlBZR1基因应对棉铃虫的抗性机制对于抗棉铃虫番茄品种的选育具有一定的指导意义。
发明内容
本发明的目的在于提供一种可调控植物对虫害胁迫尤其是棉铃虫抗性的基因,通过基因改造达到提高植物对棉铃虫抗性的目的,为培育抗虫害的植物品种提供依据。
为实现上述目的,本发明采用如下技术方案:
本发明提供了BZR1基因在调控植物对虫害胁迫抗性中的应用,所述应用为:利用生物技术手段使得所述BZR1基因功能缺失,进而提高植物对虫害胁迫的抗性;
所述BZR1基因的编码序列如SEQ ID NO.1所示或与SEQ ID NO.1所示序列具有至少70%同源性且编码的蛋白在功能上等价。
进一步的,所述BZR1基因编码的蛋白质负调控茉莉酸的合成。
进一步的,所述BZR1基因编码的蛋白质结合到茉莉酸合成相关基因Acx1的启动子上,负调控Acx1基因的转录,降低茉莉酸的含量。
本发明研究表明,BZR1基因编码的蛋白负调控茉莉酸(JA)合成相关基因Acx1(Acyl-CoA oxidase 1)的转录,Acx1基因编码的乙酰辅酶A氧化酶为JA合成的关键酶,茉莉酸在植物抵御生物胁迫中具有重要作用。通过缺失BZR1基因功能,可显著提高植物对虫害胁迫的抗性。
进一步的,所述应用包括:利用基因突变、基因敲除、基因干扰或基因沉默技术导致所述BZR1基因的表达降低或缺失,从而获得对虫害胁迫抗性增强的突变体植株。
进一步的,所述植物为番茄。
进一步的,所述虫害胁迫为棉铃虫。
本发明对番茄SlBZR1(基因编号:Solyc04g079980)进行序列分析,查找PAM序列,将NGG前的20bp的序列定义为sgRNA,选定位于基因蛋白编码区上且具有高度特异性的sgRNA序列。通过基因编辑技术和组培技术,构建了SlBZR1基因的CRISPR/Cas9载体,并筛选获得了遗传稳定并且不含外源Cas9蛋白的纯合突变体株系。通过接种棉铃虫发现突变体植株能够显著增强番茄对棉铃虫的抗性。
所述SlBZR1基因的全基因DNA序列如SEQ ID NO.4所示,蛋白编码区的核苷酸序列如SEQ ID NO.1所示,其蛋白编码区长度为987bp;该SlBZR1基因编码的蛋白为油菜素内酯信号通路中的关键转录因子,由328个氨基酸组成,其序列如SEQ ID NO.2所示。
在棉铃虫胁迫条件下,与野生型番茄(WT)相比,bzr1缺失突变体中茉莉酸含量显著上升。因此,缺失番茄Slbzr1基因有利于提高番茄对棉铃虫的抗性。
SlBZR1基因编码蛋白负调控JA合成相关基因Acx1(基因编号:Solyc08g078390)的转录。进一步的,所述Acx1基因启动子的核苷酸序列如SEQ ID NO.3所示。
本发明还提供了一种抗棉铃虫番茄的培育方法,包括以下步骤:
(1)在SlBZR1基因的蛋白编码区选取含有PAM结构的靶标片段,以靶标片段PAM结构前20个碱基为依据,进行引物设计,构建CRISPR/Cas9载体;所述SlBZR1基因的核苷酸序列如SEQ ID NO.4所示;
(2)构建含步骤(1)所述CRISPR/Cas9载体的农杆菌基因工程菌;
(3)将步骤(2)所述基因工程菌转化番茄子叶,获得不含外源Cas9蛋白、且靶标序列发生变异的稳定遗传的纯合突变体株系。
进一步的,所述靶标片段PAM结构前20bp的核苷酸序列如SEQ ID NO.5所示。
本发明具备的有益效果:
本发明首次公开BZR1基因在调控植物对虫害胁迫尤其是棉铃虫抗性方面的用途,通过基因编辑技术缺失BZR1基因功能,可显著增强植物对对棉铃虫的抗性,可将其应用到抗虫害胁迫植株品种的培育中。
附图说明
图1为CRISPR/Cas9介导的敲除SlBZR1的纯合突变体植株的基因编辑位点及编码氨基酸的变化;
其中,WT为未转基因的野生型番茄,bzr1为SlBZR1基因敲除的突变体番茄;与WT相比,突变体在sgRNA的位置发生10bp的碱基缺失,导致蛋白翻译提前终止。
图2为番茄SlBZR1基因敲除的突变体植株接种棉铃虫3天后的棉铃虫表型(A)及重量(B);
其中,棉铃虫体型、重量越大,表示棉铃虫取食越多,番茄植株受虫害越严重;小写字母a、b代表不同植株之间在T检验中P<0.05水平上的差异显著。
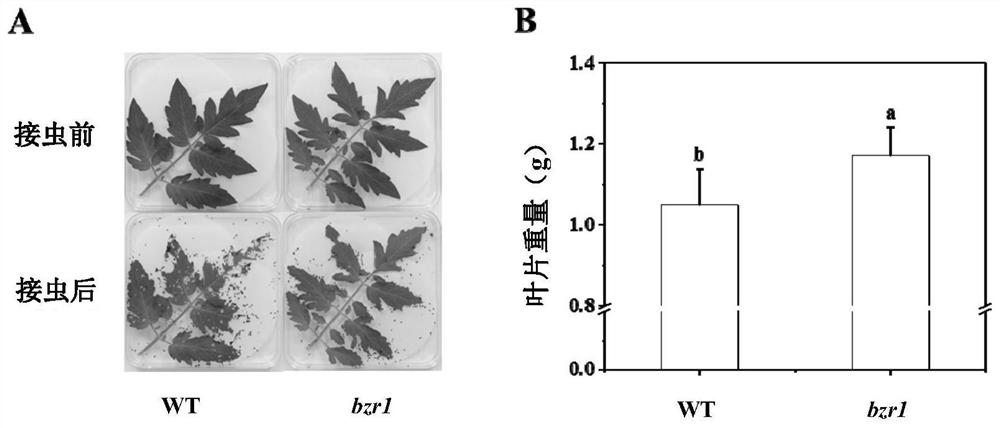
图3为番茄SlBZR1基因敲除的突变体植株接种棉铃虫3天后的叶片表型(A)及重量(B);
其中,番茄叶片受损越多,重量越小,表示棉铃虫取食越多,番茄植株受虫害越严重;小写字母a、b代表不同植株之间在T检验中P<0.05水平上的差异显著。
图4为番茄SlBZR1基因敲除的突变体植株进行机械伤加棉铃虫口器分泌物处理(W+OS)模拟棉铃虫取食后的JA含量;
其中,JA含量越多,代表植株抗虫性越强;小写字母a、b代表不同植株之间在单因素ANOVA检验中P<0.05水平上的差异显著。
图5为SlBZR1蛋白调控Acx1基因转录的荧光素酶报告基因检测比值;
其中,小写字母a、b代表不同处理之间在T检验中P<0.05水平上的差异显著。
具体实施方式
下面结合具体实施例对本发明作进一步描述,以下列举的仅是本发明的具体实施例,但本发明的保护范围不仅限于此。
下述实施例中所用实验方法如无特殊说明,均为常规方法。
下述实施例中所用实验材料、试剂等,如无特殊说明,均可从商业途径得到。
下述实施例中采用的番茄品种为番茄常规品种Condine Red。
实施例1番茄SlBZR1缺失突变体植株的制备与鉴定
1、含特异sgRNA的CRISPR/Cas9载体的构建
首先,在SGN网站https://solgenomics.net/上找到番茄SlBZR1基因的DNA全长序列如SEQ ID NO.4所示,将其输入http://crispr.hzau.edu.cn/CRISPR2/网站,找出onscore得分高,且GC含量>40%,位于蛋白编码区的一段PAM结构前的20bp碱基序列作为特异性靶向基因蛋白编码区的sgRNA,如SEQ ID NO.5所示。
合成单链sgRNA序列后退火形成双链sgRNA,且其两端具有BbsⅠ限制性内切酶酶切位点,上述合成片段插入到AtU6-sgRNA-AtUBQ-Cas9载体的BbsⅠ位点,得到中间载体。
利用HindⅢ和KpnⅠ限制性内切酶对上述中间载体及pCAMBIA1301载体进行双酶切并连接转化大肠杆菌DH5α感受态,挑单克隆进行测序,测序正确的质粒电击转化GV3101农杆菌感受态,获得可用于构建CRISPR/Cas9基因编辑材料的农杆菌菌株。
2、SlBZR1基因突变体材料的制备与鉴定
番茄种子经消毒后播种于1/2MS培养基中,黑暗培养至种子发芽后,移至光下培养。待子叶长出后,将子叶切下置于看护培养基中,黑暗培养12-24h。将准备好的用于构建CRISPR/Cas9基因编辑材料的农杆菌培养至对数期,离心去上清,加入MS0.2重悬浮,用于侵染子叶外植体,暗下侵染2.5min后,吸干子叶外植体表面剩余菌液后置于看护培养基上,黑暗下共培养两天。共培养后,子叶外植体转移至2Z培养基中诱导愈伤组织,2-3周后转入0.2Z培养基中诱导发芽,每2周转入新的0.2Z培养基,直至长出小苗后,转入R培养基诱导生根,约2-3周后待根系发育良好,将其转移至基质中并进行保湿,成活后对T
取少量T
验证引物如下:
验证前引物:5’-AGTAAGCAAAACCCACCTGC-3’(SEQ ID NO.6);
验证后引物:5’-TTCATAATCAATTCTAGATC-3’(SEQ ID NO.7)。
将T
对上述株系进一步自交繁种,种子播种后获得不含外源基因Cas9,且sgRNA发生变异的稳定遗传的T
以下实施例均以T
实施例2番茄SlBZR1基因抗棉铃虫表型的鉴定
棉铃虫采用河南省济源白云实业有限公司购买的3龄棉铃虫,挑选大小一致的棉铃虫至24孔板中,进行饥饿一晚的处理。
待番茄幼苗长至六叶一心时,选取从上往下第2-4节位叶片,称取相同质量叶片(1±0.05g)置于装有1%琼脂的培养皿中,以防止离体叶片失水,并对接虫前番茄叶片进行拍照记录。然后将两头经饥饿处理的棉铃虫接种到一个培养皿的番茄叶片上,盖上培养皿盖并用皮筋固定,防止棉铃虫离开培养皿。弱光下接虫处理3天后,检测棉铃虫重量,然后用70%甲醇浸泡棉铃虫至其不再活动,按体型大小排列后进行拍照记录。对接虫后番茄叶片也相应进行拍照记录及称重。
结果如图2和图3所示,饲喂bzr1突变体叶片的棉铃虫相比饲喂野生型叶片的棉铃虫体型较小、重量较轻,且bzr1突变体叶片经接虫处理后受到的损失相比野生型叶片更轻,受损后叶片重量较重,说明通过敲除SlBZR1基因可以提高番茄的棉铃虫抗性。
实施例3番茄SlBZR1基因对抗性激素JA的影响
待番茄幼苗长至六叶一心时,选取从上往下第3-4节位叶片进行W+OS模拟棉铃虫取食处理,并于0、1h采集叶片样品分析JA激素水平,每个时间点设置4个重复。
W+OS模拟棉铃虫取食处理方式如下:用止血钳在小叶主叶脉两侧叶面各夹一次制造机械伤,然后在创伤处涂抹稀释五倍的棉铃虫口器分泌物。
棉铃虫口器分泌物的获取方式为:棉铃虫饥饿12h后,饲喂番茄叶片,12h后吸取棉铃虫口器分泌物为原液。
JA激素测定方式为:称取0.1g番茄叶片,经液氮研磨后,加入1ml色谱级乙酸乙酯,避光加入25μl激素内标,涡旋混匀后,包上一层锡箔纸以避光,置于4℃摇床过夜,摇床频率为150rpm。4℃、12000rpm离心10min后,吸取850μl上清于新的10ml管中,4℃避光保存。剩下的沉淀中再加入1ml色谱级乙酸乙酯,重悬后置于4℃摇床震荡1h,再次4℃、12000rpm离心10min后,吸取900μl上清。合并两次上清,利用氮气吹干。加入500μl 70%色谱级甲醇,涡旋30s,转移液体至1.5ml管中,瞬离后吸取200μl上清至进样瓶内衬中,利用高效液相色谱串联质谱法测定(Varian320-MSLC/MS,Agilent Technologies,Amstelveen,Netherlands)。
结果如图4所示,W+OS处理后1h,bzr1突变体植株的JA含量显著高于野生型植株,说明敲除SlBZR1基因可以提高番茄植株在棉铃虫取食后的JA含量,从而提高番茄的棉铃虫抗性。
实施例4番茄SlBZR1蛋白负调控Acx1基因的转录验证
1、构建含SlBZR1基因的pGreenII 0029 62-SK载体:以番茄cDNA为模板,进行PCR扩增,得到SlBZR1 CDS序列。引物如下:
SlBZR1-SK-F:
5’-cgctctagaactagtggatccATGTGGGAAGGTGGAGGGTT-3’(SEQ ID NO.8);SlBZR1-SK-R:
5’-gataagcttgatatcgaattcTCACATCCGAGCAGTCCCAC-3’(SEQ ID NO.9);
利用BamHⅠ和EcoRⅠ限制性内切酶对上述载体及PCR产物进行双酶切,并连接转化大肠杆菌DH5α感受态。
2、构建含Acx1基因启动子序列的pGreen II 0800-LUC载体:方法同上,以番茄基因组DNA为模板,酶切位点为NotⅠ和NcoⅠ,所用引物如下:
pAcx1-LUC-F:
5’-tccactagttctagagcggccgcTTTTTTTCATGTTGGACGGTCTG-3’(SEQ ID NO.10);
pAcx1-LUC-R:
5’-tgtttttggcgtcttccatggAATAAAATGTTCGTCCACTTTAATATATTC-3’(SEQ IDNO.11)。
3、将测序正确的质粒电击转化农杆菌GV3101感受态,获得可用于双荧光素酶报告基因检测实验的农杆菌菌株。
4、将上述两种农杆菌菌液在含有150μmol/L乙酰丁香酮的侵染液中重悬并调至最佳浓度(OD
结果如图5所示,相比空载体,BZR1的表达使得Acx1启动子受到抑制,LUC/REN相对值较低,说明BZR1抑制了Acx1的转录。
序列表
<110> 浙江大学
浙江大学山东(临沂)现代农业研究院
<120> BZR1基因在调控植物对虫害胁迫抗性中的应用
<160> 11
<170> SIPOSequenceListing 1.0
<210> 1
<211> 987
<212> DNA
<213> 番茄(Solanum lycopersicum L.)
<400> 1
atgatgtggg aaggtggagg gttgccggtg gagggtggtg gtggtgttgg tgaaggtggt 60
ggtgttggtg gtggtggagg tggtggtagt gggaggagga agccatcatg gagggaaagg 120
gagaataata ggaggaggga aaggaggaga agggcaatag cagctaagat ttatagtgga 180
ttaagagcac aggggaatta taatcttcct aaacattgtg ataacaatga ggttttgaag 240
gctctttgtg ttgaagctgg atggattgtt gagcctgatg gaactactta tagaaaggga 300
tgcaggccaa ctccaatgga gattggaggc acttcagcca acattacgcc aagttcttca 360
cgaaatccaa gtcctccctc ttcatacttt gctagcccga ttccatctta ccaagttagt 420
ccaacatcct cgtctttccc aagtccatct cgtggtgatg ctaacatgtc gtcacatcca 480
tttgcatttc tccatagttc cattcccttg tcgctaccac cattacgaat atcaaacagt 540
gcccctgtaa caccacctct ttcatcacca actagagtcc ctaagcagat atttaatctt 600
gagactttgg ctagagagtc tatgtctgct ctaaatatcc ctttctttgc tgcttcagcc 660
ccaactagcc caactcgagg tcagcgattc actcctgcta caataccaga gtgtgacgaa 720
tctgattcat ccaccattga ttctggccag tggatgagct ttcaaaagta cgcagccaat 780
gggatcccta cttctccgac ttttaatctt attaagcctg tagctcagag aattccttct 840
aatgatatga tcatcgacaa gggtaagagc attgaatttg actttgagaa tgtatcagtt 900
aaggcagcat gggaaggtga aaagattcat gaggttggtt tagatgatct ggagctcact 960
ctcggaagtg ggactgctcg gatgtga 987
<210> 2
<211> 328
<212> PRT
<213> 番茄(Solanum lycopersicum L.)
<400> 2
Met Met Trp Glu Gly Gly Gly Leu Pro Val Glu Gly Gly Gly Gly Val
1 5 10 15
Gly Glu Gly Gly Gly Val Gly Gly Gly Gly Gly Gly Gly Ser Gly Arg
20 25 30
Arg Lys Pro Ser Trp Arg Glu Arg Glu Asn Asn Arg Arg Arg Glu Arg
35 40 45
Arg Arg Arg Ala Ile Ala Ala Lys Ile Tyr Ser Gly Leu Arg Ala Gln
50 55 60
Gly Asn Tyr Asn Leu Pro Lys His Cys Asp Asn Asn Glu Val Leu Lys
65 70 75 80
Ala Leu Cys Val Glu Ala Gly Trp Ile Val Glu Pro Asp Gly Thr Thr
85 90 95
Tyr Arg Lys Gly Cys Arg Pro Thr Pro Met Glu Ile Gly Gly Thr Ser
100 105 110
Ala Asn Ile Thr Pro Ser Ser Ser Arg Asn Pro Ser Pro Pro Ser Ser
115 120 125
Tyr Phe Ala Ser Pro Ile Pro Ser Tyr Gln Val Ser Pro Thr Ser Ser
130 135 140
Ser Phe Pro Ser Pro Ser Arg Gly Asp Ala Asn Met Ser Ser His Pro
145 150 155 160
Phe Ala Phe Leu His Ser Ser Ile Pro Leu Ser Leu Pro Pro Leu Arg
165 170 175
Ile Ser Asn Ser Ala Pro Val Thr Pro Pro Leu Ser Ser Pro Thr Arg
180 185 190
Val Pro Lys Gln Ile Phe Asn Leu Glu Thr Leu Ala Arg Glu Ser Met
195 200 205
Ser Ala Leu Asn Ile Pro Phe Phe Ala Ala Ser Ala Pro Thr Ser Pro
210 215 220
Thr Arg Gly Gln Arg Phe Thr Pro Ala Thr Ile Pro Glu Cys Asp Glu
225 230 235 240
Ser Asp Ser Ser Thr Ile Asp Ser Gly Gln Trp Met Ser Phe Gln Lys
245 250 255
Tyr Ala Ala Asn Gly Ile Pro Thr Ser Pro Thr Phe Asn Leu Ile Lys
260 265 270
Pro Val Ala Gln Arg Ile Pro Ser Asn Asp Met Ile Ile Asp Lys Gly
275 280 285
Lys Ser Ile Glu Phe Asp Phe Glu Asn Val Ser Val Lys Ala Ala Trp
290 295 300
Glu Gly Glu Lys Ile His Glu Val Gly Leu Asp Asp Leu Glu Leu Thr
305 310 315 320
Leu Gly Ser Gly Thr Ala Arg Met
325
<210> 3
<211> 480
<212> DNA
<213> 番茄(Solanum lycopersicum L.)
<400> 3
tttttttcat gttggacggt ctgcctactt tatttttttt tgttcatttt ccgttagtga 60
aatctgatta aatttaaata atagagcatg aaatttattt ggagtgatat atttaacaca 120
attttttcta tatcatgtgt ttgaatttga aattttttat taaagatgaa ataatttctc 180
cgttgcacga taactcattt tagaagtacc attattgatc aaaatactat taaatattca 240
aataaaattt attgacaaat tgatgtatcg ctctttttaa aaaaaacata ttaggagtac 300
tgaaaataga aaaatatggg ctaaatatcc tcttatcgtc actataccga ctgttcacac 360
tgcggaccac acgaagcaaa aaaaaaaagg ctataattat actcttaata tactatgagc 420
attttttttt aaaaaaaata tatatactat gaatatatta aagtggacga acattttatt 480
<210> 4
<211> 3493
<212> DNA
<213> 番茄(Solanum lycopersicum L.)
<400> 4
taaaataaac ttattcattt attatttata aacaagttat attataattt ttgataatca 60
aactttctga ttcaacctca ttataaaata tccttgttat tgccttatcg ctttttctat 120
ctataaaatt tatcacaatt atacaaaaaa aaaattactt taattttatc tatgtgtcta 180
tcataactaa cctcgattta agaccagaaa acttataata acatagctat taccccatta 240
ataaaatttt aaaaagcagt caaacattaa aaactttata tttaaattcc aaaatataaa 300
ctagtaaata acaattatta tcttgcttca tttgataagt aaaaaattat ttcaactctt 360
aaataacagt caaaaaaatt aatttagtag tctaatcatg taatttgtaa aatctaaaaa 420
agtagtcaaa gttttttttt ctttttgcca tttttaaaaa atagaatgaa aaacgcttac 480
acatgataat tctgaagctg taacaaactt tatctcttca attattataa ttttgctaaa 540
aaacaaaaca cacaaaacac catttcatca tcattttttt tttatatata tatatatata 600
acacaaacac acagagagaa aagtaaagaa gaaaattctt cagattgttt gtagtagtat 660
aaactttaga gcgtatttct cgtgagtagt cccaaaaaaa tgctttatag agtggcaccc 720
ctagtaccta tacaacacag taagcaaaac ccacctgctg tctgaacaca cccacctggg 780
accagccccc ctttttcttc ttctttcttc tttggattca tcatataaaa aaaacccatt 840
ttcttaattt tgagaaatct tgaaaaaaat cgattctttt tggggagaaa atgatgtggg 900
aaggtggagg gttgccggtg gagggtggtg gtggtgttgg tgaaggtggt ggtgttggtg 960
gtggtggagg tggtggtagt gggaggagga agccatcatg gagggaaagg gagaataata 1020
ggaggaggga aaggaggaga agggcaatag cagctaagat ttatagtgga ttaagagcac 1080
aggggaatta taatcttcct aaacattgtg ataacaatga ggttttgaag gctctttgtg 1140
ttgaagctgg atggattgtt gagcctgatg gaactactta tagaaaggtt agtaaaattc 1200
atcttctttt atgaattgat gtatggttaa agtagtgttt gtgtcttagt ttttgtttgt 1260
ttggattgaa ggggtgatgt gtttttggtt ggagttttgc ttagatctat agatttggaa 1320
aatggggttg ttgaatttgg atcaaagatt gtgtttttat ctgttttcag ttggtttttg 1380
actgttttat ggttttctga tctagaattg attatgaaca gcttctgagt ttgtgttttc 1440
tttagttcaa ttgttctttt atgtacttat aatctgttca tgttctcatt tctggataat 1500
tggtatgaat gtatgtagtt taatttgtct gagaatggga gttgtttctg gaaaaaacaa 1560
gaaggaagaa ttgacttgtt cttaatatag aaatggataa gcagggtgtt ttggtgtcta 1620
aggtggggtt tgatgcattt ggttgtggtg tttttagttt atcaactact tttcgagatt 1680
caaagttgca atctgttgaa gttgttatct cagattacag agccttttga tcatttatgt 1740
tcactagttt gatttgaaaa tgtcctcttt tactatgttt tcagggatgc aggccaactc 1800
caatggagat tggaggcact tcagccaaca ttacgccaag ttcttcacga aatccaagtc 1860
ctccctcttc atactttgct agcccgattc catcttacca agttagtcca acatcctcgt 1920
ctttcccaag tccatctcgt ggtgatgcta acatgtcgtc acatccattt gcatttctcc 1980
atagttccat tcccttgtcg ctaccaccat tacgaatatc aaacagtgcc cctgtaacac 2040
cacctctttc atcaccaact agagtcccta agcagatatt taatcttgag actttggcta 2100
gagagtctat gtctgctcta aatatccctt tctttgctgc ttcagcccca actagcccaa 2160
ctcgaggtca gcgattcact cctgctacaa taccagagtg tgacgaatct gattcatcca 2220
ccattgattc tggccagtgg atgagctttc aaaagtacgc agccaatggg atccctactt 2280
ctccgacttt taatcttatt aagcctgtag ctcagagaat tccttctaat gatatgatca 2340
tcgacaaggg taagagcatt gaatttgact ttgagaatgt atcagttaag gcagcatggg 2400
aaggtgaaaa gattcatgag gttggtttag atgatctgga gctcactctc ggaagtggga 2460
ctgctcggat gtgattttca tgattaaaag aactgctcag tgtggattta cagattgaga 2520
ttatccgagc ttgctagctg atgattcact caagcagtcc agtggatgtt cgttacagag 2580
atagatcttg catgttgctg tttaccttat atacagacat tttttttttc cttttcgcct 2640
tatctgatga ggcctttaag tttagtttgg gtcttgtcat aagcatttag gtatagatag 2700
ttgatggttt ctatctcctt gtggttgaga attgtttcaa gtcattgtag actagcagga 2760
gttctcctta aacctgaact atccacagtt atattttcat gattatagta gatccctttg 2820
ttgttatgca actggatttc ttgttttatg atctagcgtc aattcaaaat caatggatgt 2880
gctattcttt aagtctcttt gatgctttat gtccagatca aaacattagt actagttctc 2940
tttgtttaca cacactttca ctataccatt ttaactcata tgtccttaaa ggacaagcaa 3000
tggtgggcca gaaaagactt tgacgactca agggttcata agtgagtgaa agtaagatag 3060
ccctctaaag gaagatctat tcagctcgat catctaattc actccaacag acatgcatga 3120
ttccgtaatc gaaataactt ccatagggct ggccaatggc aagatggatg tcaagaagtt 3180
ggaatttttt gacaatcaaa tcaaagttgt tgaattttct ttgtcactca tttggctcca 3240
ttagtaaact accagtagtc atattaataa taatagcatg ttcatgcaca cgtttgttat 3300
caacacaagc acagacagag aagaagaaag ataaaacaaa tggtgtttaa ctgatttaat 3360
gtacaagacc acggcctaat gtctggatgt ggggttctgt actactgtat tacacatgtt 3420
cctctacagt tactttttca ttattttttt ctcataacgc ggttattgtc acatgtcttg 3480
cttgctgcta tat 3493
<210> 5
<211> 20
<212> DNA
<213> 番茄(Solanum lycopersicum L.)
<400> 5
ggaagccatc atggagggaa 20
<210> 6
<211> 20
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 6
agtaagcaaa acccacctgc 20
<210> 7
<211> 20
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 7
ttcataatca attctagatc 20
<210> 8
<211> 41
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 8
cgctctagaa ctagtggatc catgtgggaa ggtggagggt t 41
<210> 9
<211> 41
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 9
gataagcttg atatcgaatt ctcacatccg agcagtccca c 41
<210> 10
<211> 46
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 10
tccactagtt ctagagcggc cgcttttttt catgttggac ggtctg 46
<210> 11
<211> 51
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 11
tgtttttggc gtcttccatg gaataaaatg ttcgtccact ttaatatatt c 51