基于植物功能群构建的杉木林土壤生态服务功能提升方法
文献发布时间:2023-06-19 12:25:57
技术领域
本发明属于林业生态修复领域,涉及土壤修复技术,具体涉及基于植物功能群构建的杉木林土壤生态服务功能提升方法。
背景技术
杉木(Cunninghamia lanceolata(Lamb.)Hook.)是我国南方最主要的用材树种之一,其具有速生、材质好、产量高、用途广等诸多优点。但近几十年来,杉木人工林种植面积的不断扩张、看重短暂经济效益的错误理念以及不当的营林方式,出现了杉木人工林物种多样性减少、地力退化、碳汇功能降低等一系列生态问题。因此,为了扭转杉木人工林现状,提升其林分生物多样性、生产力以及固碳减排能力等生态系统服务功能,使之既可以呈现健康稳定的人工林生态系统状态又可以满足人类对木材生产上的需求,是林学界和生态学界共同面临的问题。
林下植被恢复对于退化杉木人工林生态系统服务功能的提升具有重要作用,但现行做法通常是针对整个林区采用施肥、保留枯落物、混交、改变炼山或整地方式、密度控制等单一措施改善杉木林下土壤单一方面的问题,也未针对林龄不同进行区别处理,对土壤生态服务功能提升有限。
发明内容
本发明提供一种基于植物功能群构建的杉木林土壤生态服务功能提升方,旨在通过在杉木林不同林龄区域构建不同的特定植物功能群以综合性解决杉木人工林物种多样性减少、地力退化、碳汇功能降低等一系列生态问题。
本发明解决上述技术问题的技术方案如下:基于植物功能群构建的杉木林土壤生态服务功能提升方法,其包括如下内容:
1)调查林地植被状况,对需要林下植被及土壤修复的杉木林进行识别并按照林龄不同进行划片分区,确定各待修复区域为杉木幼龄林、中龄林、近熟林、成熟林和/或过熟林及各自边界位置;
2)在春季向各待修复区域引入植物功能群,根据林龄不同分别引入由南方红豆杉、南岭黄檀、鸡血藤和山柰中的两种及以上植物组合形成的不同植物功能群。
在上述技术方案的基础上,本发明还可以有如下进一步的具体选择。
具体的,对于幼龄林和近熟林引入由南方红豆杉、鸡血藤和山柰组成的植物功能群A,对于中龄林和过熟林引入由南岭黄檀和山柰组成的植物功能群B,对于成熟林引入由南岭黄檀和南方红豆杉组成的植物功能群C。
具体的,植物功能群A中南方红豆杉、鸡血藤的株数和山柰簇数比为1:1:1;植物功能群B中南岭黄檀的株数与山柰簇数比为1:1;植物功能群C中南岭黄檀与南方红豆杉的株数比为1:1。
具体的,植物功能群A中南方红豆杉、鸡血藤和山柰采用条带状混交种植,南方红豆杉和鸡血藤在同一行内交错种植,南方红豆杉和鸡血藤的株行距均为2m×2m,同一行内南方红豆杉与相邻的鸡血藤株间距为1m,山柰采用行间种植,两行间相邻两株鸡血藤之间种植有一簇山柰,每簇山柰内有四个山柰块茎且分布于边长0.5m正方形的四个顶点。
具体的,植物功能群B中南岭黄檀与山柰采用条带状混交种植,南岭黄檀的株行距为2m×2m,两行间相邻两株南岭黄檀之间种植有一簇山柰,每簇山柰内有四个山柰块茎且分布于边长0.5m正方形的四个顶点。
具体的,植物功能群C中的南岭黄檀和南方红豆杉采用块状交错种植,南岭黄檀和南方红豆杉的株行距均为2m×2m,行内和行间南岭黄檀和南方红豆杉均交错种植。
具体的,春季引入时,南方红豆杉、南岭黄檀和鸡血藤均采用直接种植苗高1米以下的相应树苗,山柰采用块茎直接埋入。
与现有技术相比,本发明的有益效果是:
本发明首次通过植物功能群引入来提高杉木人工林土壤生态服务功能,并且根据林龄不同引入不同的特定植物组合的植物功能群,可综合解决杉木人工林经营管理过程中存在的肥力下降(氮、磷营养缺乏等)、生物多样性降低、碳汇减少等多种土壤生态服务功能下降问题,本发明首次提出向不同林龄引入不同林下植物功能群的概念并付诸实践,取得了较好的效果。
附图说明
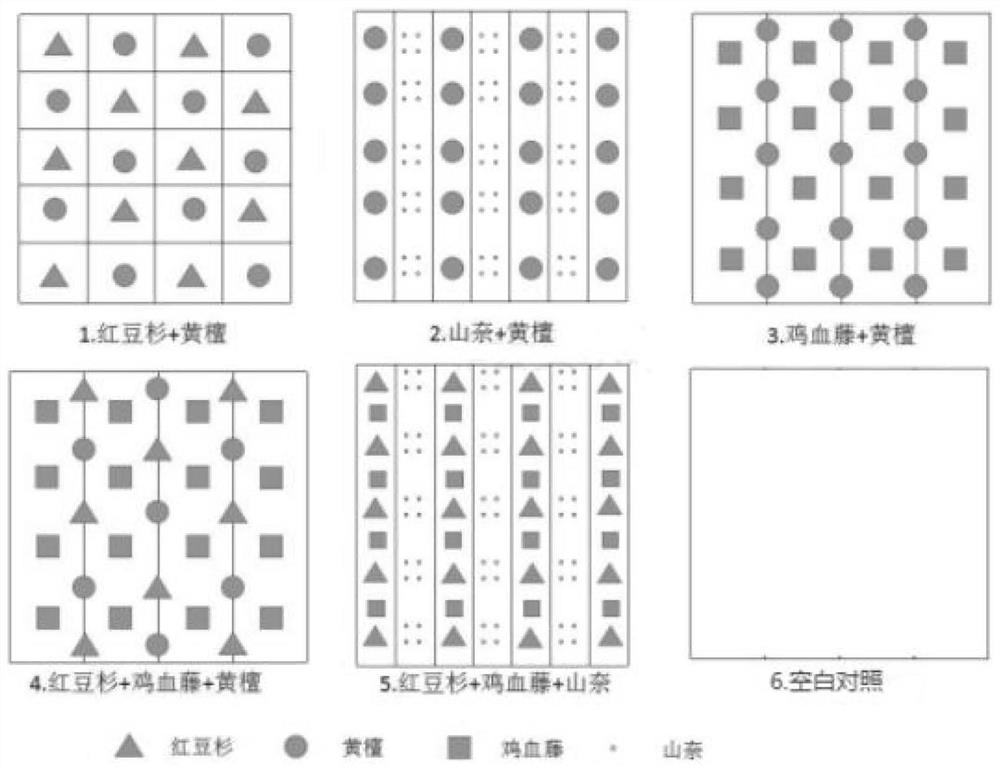
图1为本发明提供的五种植物功能群的配置示意图;
图2为幼龄林引入各植物功能群后林下土壤的真菌群落主成分分析图;
图3为中龄林引入各植物功能群后林下土壤的真菌群落主成分分析图;
图4为近熟林引入各植物功能群后林下土壤的真菌群落主成分分析图;
图5为成熟林引入各植物功能群后林下土壤的真菌群落主成分分析图;
图6为过熟林引入各植物功能群后林下土壤的真菌群落主成分分析图;
图7为幼龄林引入各植物功能群后林下土壤的细菌群落主成分分析图;
图8为中龄林引入各植物功能群后林下土壤的细菌群落主成分分析图;
图9为近熟林引入各植物功能群后林下土壤的细菌群落主成分分析图;
图10为成熟林引入各植物功能群后林下土壤的细菌群落主成分分析图;
图11为过熟林引入各植物功能群后林下土壤的细菌群落主成分分析图;
图12为幼龄林引入各植物功能群后土壤真菌的相对丰度图;
图13为中龄林引入各植物功能群后土壤真菌的相对丰度图;
图14为近熟林引入各植物功能群后土壤真菌的相对丰度图;
图15为成熟林引入各植物功能群后土壤真菌的相对丰度图;
图16为过熟林引入各植物功能群后土壤真菌的相对丰度图;
图17为幼龄林引入各植物功能群后土壤细菌的相对丰度图;
图18为中龄林引入各植物功能群后土壤细菌的相对丰度图;
图19为近熟林引入各植物功能群后土壤细菌的相对丰度图;
图20为成熟林引入各植物功能群后土壤细菌的相对丰度图;
图21为过熟林引入各植物功能群后土壤细菌的相对丰度图。
具体实施方式
以下结合附图及具体实施例对本发明提供的技术方案作进一步的详描述,所举实例只用于解释本发明,并非用于限定本发明的范围。
本发明提供一种基于植物功能群构建的杉木林土壤生态服务功能提升方法,其包括如下内容:
1)调查林地植被状况,对需要林下植被及土壤修复的杉木林进行识别并按照林龄不同进行划片分区,确定各待修复区域为杉木幼龄林、中龄林、近熟林、成熟林和/或过熟林及各自边界位置;
2)在春季向各待修复区域引入植物功能群,根据林龄不同分别引入由南方红豆杉、南岭黄檀、鸡血藤和山柰中的两种及以上植物组合形成的不同植物功能群。
如图1所示,本发明供提供了五种特定比例组成及特定种植分布的植物功能群:
标号1代表的植物功能群(植物功能群C)为南方红豆杉和南岭黄檀按照1:1的株数比块状交错种植,南方红豆杉和南岭黄檀的株行距均为2m×2m,同一行内及相邻行间南方红豆杉和南岭黄檀均交错且南方红豆杉与相邻的南岭黄檀株之间间距为1m;
标号2代表的植物功能群(植物功能群B)为南岭黄檀的株数和山柰簇数按照1:1比条带状混交种植,南岭黄檀的株行距为2m×2m,行间种植山柰,山柰的畦宽1m、高0.2m,株行距0.5m,穴深6cm,山柰块茎每穴1块;
标号3代表的植物功能群为南岭黄檀和鸡血藤按照1:1的株数比条带状错位混交种植,南岭黄檀和鸡血藤的株行距均为2m×2m;
标号4代表的植物功能群为南方红豆杉、南岭黄檀和鸡血藤按照1:1:2的株数比条带状错位混交种植,南方红豆杉和南岭黄檀的株行距分别为4m×4m,鸡血藤的株行距为2m×2m,鸡血藤单独成行,鸡血藤的行间由南方红豆杉和南岭黄檀交错种植成行并均与鸡血藤错开。
标号5代表的植物功能群(植物功能群A)为南方红豆杉、南岭黄檀和山柰按照1:1:1的株数比条带状混交种植,南方红豆杉和南岭黄檀的株行距均为2m×2m,同一行内南方红豆杉和南岭黄檀交错且两行株间距为1m,行间种植山柰,山柰的畦宽1m、高0.2m,株行距0.5m,穴深6cm,山柰根茎每穴1块。
标号6代表空白对照,即在林下不引入植物功能群。
春季引入时,南方红豆杉、南岭黄檀和鸡血藤均采用直接种植苗高1米以下的相应树苗,山柰采用块茎直接埋入且每穴通常埋入3颗块茎。
以上述的植物功能群在春季时对林下土壤待生态修复的杉木人工林的幼龄林(Y)、中龄林(Z)、近熟林(J)、成熟林(C)和过熟林(G)进行分别引入,引入两年后测试林下土壤中理化性质以及微生物数量、种类和丰度等,为便于对土壤生态恢复的结果进行描述,以下内容中以Y1代表向幼龄林引入标号为1的植物功能群,即向幼龄林引入由南方红豆杉和南岭黄檀1:1组成的植物功能群,字母代表相应林龄的杉木人工林,数字代表图1中相应标号的植物功能群(标号1-5)或空白对照(标号6)。恢复效果如下:
1、不同植物功能群对土壤理化性质的影响
在杉木幼龄林,林下土壤有机质含量、全氮含量、全磷含量、碱解氮含量、有效磷含量以及速效钾含量均在Y5(红豆杉+鸡血藤+山柰)模式样地得到提升,与对照组相比,有机质含量提升了30.81%,全氮含量提升了45.77%,全磷含量提升了27.59%,碱解氮含量提高了41.66%,速效钾含量提升了20.75%。幼龄林的pH值在Y4(黄檀+鸡血藤+红豆杉)时最高,但与其他模式组差异基本都不显著。幼龄林全钾含量在Y1(黄檀+红豆杉)最高,提升了188.59%,显著高于其他种植模式组。
在杉木中龄林,土壤有机质、全氮、碱解氮、有效磷含量在Z2(黄檀+山柰)组显著高于其他组,与对照组相比有机质含量提高了31.87%,全氮含量提高了28.95%,碱解氮含量提高了43.94%,有效磷含量提高了174.35%。土壤pH和全钾含量在Z4(黄檀+红豆杉+鸡血藤)组显著高于其他各个组,土壤全钾含量提升了19.57%。全磷含量在Z1(黄檀+红豆杉)和Z4(黄檀+红豆杉+鸡血藤)组显著高于其他种植模式组,Z1组的全磷含量相比对照组提升最高,为46.43%。各个种植模式对于中龄林速效钾的影响不显著。
在杉木近熟林,土壤pH值和全磷含量在J3(黄檀+鸡血藤)模式显著高于其他种植模式,全磷含量相比对照组提升了40%。J5(红豆杉+鸡血藤+山柰)组对于土壤有机质含量和碱解氮含量的提升最显著,有机质含量提升了55.46%,碱解氮含量提升了93.72%。J2(黄檀+山柰)对土壤全氮和有效磷含量的提升效果最显著,全氮含量提升了62.73%,有效磷含量提高了44.08%。不同的种植模式对于速效钾的含量没有显著影响。
在杉木成熟林,C1(黄檀+红豆杉)组对于土壤全氮、碱解氮以及有效磷含量提升最显著,使全氮含量提升了13.97%,碱解氮含量提高了62.56%,有效磷含量提高了326.86%。对土壤有机质提升最高,但与其他组差异不显著。土壤全磷和全钾在C5(红豆杉+鸡血藤+山柰)组含量显著高于其他各组,全磷含量提升了15.15%,全钾含量提升了7.18%。C3(黄檀+鸡血藤)模式和C1(黄檀+红豆杉)模式对土壤速效钾的提升效果最显著,速效钾在C3模式相比对照组的提升最高,为61.04%。土壤pH值在C2(黄檀+山柰)模式显著高于其他模式组。
在杉木过熟林,G1(黄檀+红豆杉)组使得土壤有机质、全磷、碱解氮、速效钾含量均显著高于其他模式组,相比对照组,有机质含量提升了44.20%,全磷含量提升了16.28%,碱解氮含量提升了248.74%,速效钾含量提升了51.28%。G5(红豆杉+鸡血藤+山柰)对于土壤全氮和全钾的含量提升最高,但与其他种植模式组之间差异不显著。G2(黄檀+山柰)对于土壤有效磷的含量效果最显著。G3(黄檀+鸡血藤)对于土壤pH值的提升最高,但与其他模式组之间的差异不显著。
在Y5(红豆杉+鸡血藤+山柰)模式下,杉木幼龄林中的土壤脲酶活性相比对照组提高了35.74%,脲酶活性显著高于其他种植模式;在Z4(黄檀+红豆杉+鸡血藤)模式下,中龄林脲酶活性相比对照组提升了50.42%,且显著高于其他种植模式;在J1/C1(黄檀+鸡血藤)模式下,近熟林和成熟林的脲酶活性相比对照组提升最大,分别提高了21.18%和32.76%,且脲酶活性显著高于其他模式组;在G3(黄檀+鸡血藤)模式下,过熟林脲酶活性相比对照组提高了26.97%,且脲酶活性显著高于其他模式组。在Y4/Y2/Y5模式下,杉木幼龄林中的土壤酸性磷酸酶活性均显著高于其他各组,其中Y4(黄檀+红豆杉+鸡血藤)模式的酸性磷酸酶活性较对照组的提升最多(38.97%);在G1(黄檀+鸡血藤)模式下,杉木过熟林中的酸性磷酸酶活性相比对照组来说提升最大(163.76%),且酸性磷酸酶活性显著高于其他各组;不同的种植模式对于杉木中龄林、近熟林以及过熟林的土壤酸性磷酸酶没有影响。在Y2模式(黄檀+山柰),杉木幼龄林土壤纤维素酶活性显著高于其他各组,且相比对照组来说提高了101.04%;在Z5/J5(红豆杉+鸡血藤+山柰)模式,杉木中龄林和近熟林土壤纤维素酶活性显著高于同一林龄其他各组,且相比对照组分别上升了82.44%和73.95%;在C3/G3(黄檀+鸡血藤)模式,杉木成熟林和过熟林土壤纤维素酶活性显著高于相同林龄其他各组,且相比对照组分别上升了86.49%和40.74%。。
2、不同植物功能群对土壤微生物的影响
2.1不同林龄土壤真菌群落主成分分析:
由主成分分析可知,各组间距离越大,组间差异越大。幼龄林中Y1、Y2、Y3、Y4模式与Y5、Y6模式在PC1轴完全分开,说明Y1、Y2、Y3、Y4模式与Y6模式土壤真菌群落结构差异明显,植入Y5模式后土壤真菌群落结构变化不显著。中龄林中Z1、Z5、Z6模式与Z2、Z3、Z4模式在PC1轴完全分开,Z1、Z6模式和Z5模式围绕PC2轴完全分开,说明Z2、Z3、Z4模式与Z6模式土壤真菌群落结构差异明显,Z5模式与Z6模式土壤真菌群落结构差异明显,植入Z1模式后土壤真菌相对Z6对照组的变化不显著。近熟林中J1、J2、J3、J4、J5模式组和J6对照组在PC1轴完全分开,说明在近熟林各个种植模式与对照组相比土壤真菌群落结构均发生了显著变化。在成熟林,C2模式与C1、C3、C4、C5、C6模式组在PC1轴完全分开,C3、C5模式和C1、C4、C6模式组在PC2轴完全分开,说明C2模式组与C6对照组的真菌群落结构差异显著,C3、C5模式与C6对照组的真菌群落结构差异显著,C1、C4种植模式与C6对照组相比土壤真菌群落无明显变化。过熟林中,G1模式与G2、G3、G4、G5、G6模式之间围绕PC1轴完全分开,G5模式与G2、G3、G4、G6模式之间围绕PC2轴完全分开,说明G1模式与G6对照组的土壤真菌群落结构差异显著,G5模式与G6对照组的土壤真菌群落结构差异显著,G2、G3、G4种植模式的土壤真菌群落结构与对照组相比变化不明显。
2.2不同林龄土壤细菌群落主成分分析:
幼龄林中Y4、Y6模式和Y1、Y2、Y3、Y5模式在PC1轴完全分开,说明Y1、Y2、Y3、Y5模式与Y6对照组的土壤细菌群落结构差异显著,与Y6对照组相比,种植Y4模式后细菌群落结构变化不明显。中龄林中Z1、Z2模式与Z3、Z4、Z5、Z6模式围绕PC1轴完全分开,Z3、Z5模式和Z4、Z6模式围绕PC2轴完全分开,说明Z1、Z2模式与对照组的土壤细菌群落结构差异显著,Z3、Z5模式和Z6对照组的细菌群落结构差异明显,而Z4种植模式下细菌群落结构与对照组相比差异不明显。近熟林中,J1、J2模式与J3、J4、J5、J6模式围绕PC1轴完全分开,J4、J5和J3、J6组围绕PC2轴完全分开,说明J1、J2模式与J6对照组的土壤细菌群落结构差异显著,J4、J5模式与对照组的土壤细菌群落结构差异显著,而J3模式与对照组相比土壤细菌群落结构的变化不显著。成熟林中,C3模式与C1、C2、C4、C5、C6模式围绕PC1轴完全分开,C1、C4模式与C2、C5、C6模式围绕PC2轴完全分开,说明C3模式与对照组的细菌群落结构差异明显,C1、C4模式与对照组的细菌群落结构差异也显著,而C2、C5模式与对照组之间的群落结构差异不明显。在过熟林,G1、G2、G5、G6模式与G3、G4模式围绕PC1轴分开,G1、G2模式与G5、G6模式围绕PC2轴分开,说明G3、G4模式与G6对照组的土壤细菌群落结构差异显著,G1、G2模式与对照组细菌群落结构差异显著,种植G5模式后的土壤细菌群落结构与对照相比变化不明显。
2.3不同林龄土壤微生物Alpha多样性分析
2.3.1不同林龄土壤真菌群落丰富度和多样性分析:
在杉木幼龄林中,从表1中的Chao1指数和ACE指数可以看出,不同种植模式间土壤真菌群落丰富度大小依次为Y3>Y1>Y2>Y5>Y4>Y6,土壤真菌Shannon指数大小为Y5>Y2>Y1>Y3>Y4>Y6,5种模式组的土壤真菌的丰富度和多样性水平均高于对照组,可见不同的种植模式对幼龄林土壤真菌丰富度和多样性都有改善。在杉木中龄林中,不同模式下土壤真菌群落丰富度大小依次为Z1>Z5>Z2>Z3>Z4>Z6,土壤真菌Shannon指数大小为Z3>Z2>Z1>Z5>Z4>Z6,5个种植模式内土壤真菌的Chao1指数、ACE指数、Shannon指数均高于对照组,说明施加不同种植模式对于中龄林土壤真菌丰富度和多样性也有积极影响。在杉木近熟林,不同模式土壤真菌群落丰富度大小顺序为J5>J2>J4>J1>J3>J6,土壤真菌Shannon指数大小为J1>J3>J2>J5>J4>J6,5个种植模式的土壤真菌的丰富度和多样性水平均高于对照组,说明5种模式对近熟林土壤真菌的丰富度和多样性均有所提升。在杉木成熟林,各组土壤真菌丰富度指数大小为Y4>Y1>Y2>Y6>Y5>Y3,其中Y4、Y1、Y2这三种种植模式的土壤真菌丰富度高于对照组。各个模式的土壤真菌Simpson指数大小为C3>C2>C5>C1>C4>C6,Shannon指数大小为C6>C1>C4>C5>C2>C3,结果表明对照组的土壤真菌群落多样性水平高于各个种植组,说明这5种模式对于成熟林内土壤真菌多样性没有影响。在杉木过熟林内,不同模式下土壤真菌群落丰富度Chao1指数和ACE指数大小顺序为G3>G2>G1>G5>G4>G6,5种种植模式内土壤真菌丰富度Chao1指数和ACE指数均高于对照组,过熟林土壤真菌Simpson指数大小顺序为G1>G5>G3>G6>G2>G4,过熟林土壤真菌Shannon指数大小顺序为G4>G2>G3>G6>G5>G1,G4、G2两种模式的土壤真菌多样性高于对照组。
2.3.2不同林龄土壤细菌群落丰富度和多样性分析:
在杉木幼龄林中,从Chao1指数和ACE指数可以看出,不同种植模式间土壤细菌群落丰富度大小依次为Y2>Y3>Y1>Y5>Y4>Y6,5个种植模式下土壤细菌丰富度均高于对照组。不同种植模式土壤细菌Simpson指数大小为Y6>Y3>Y4>Y2>Y5>Y1,Shannon指数大小为Y5>Y2>Y1>Y3>Y4>Y6,由此可知5个种植模式对幼龄林的土壤细菌多样性均有提升效果。在杉木中龄林中,不同模式下土壤细菌群落丰富度大小依次为Z5>Z6>Z2>Z4>Z3>Z1,Z5模式下的土壤细菌丰富度高于对照组。土壤细菌Simpson指数大小依次为Z2>Z3>Z4>Z6>Z1>Z5,Shannon指数大小为Z5>Z1>Z6>Z4>Z3>Z2,Z5和Z1两种模式下的土壤细菌多样性水平高于对照组土壤。在杉木近熟林,不同模式土壤细菌群落丰富度大小顺序为J3>J4>J6>J2>J5>J1,其中J3和J4两种模式下土壤细菌丰富度高于对照组。不同种植模式下土壤细菌Simpson指数大小为J2>J1>J3>J5>J6>J4,Shannon指数大小为J6>J4>J5>J3>J1>J2,对照组的细菌多样性指数较高,说明不同模式对于土壤细菌多样性影响不大。在杉木成熟林,各个模式下土壤细菌群落丰富度大小依次为C2>C5>C4>C6>C1>C3,其中C2、C5、C4三个种植模式的土壤细菌群落丰富度相比对照组有所提升。各个模式的土壤细菌Simpson指数大小为C3>C1>C2>C5>C4>C6,Shannon指数大小为C4>C6>C2>C5>C1>C3,对照组的土壤细菌多样性较高,施加其他种植模式对于成熟林土壤细菌群落丰富度和多样性水平基本没有提高。在杉木过熟林,不同模式下土壤细菌群落丰富度Chao1指数和ACE指数大小顺序为G2>G3>G4>G6>G5>G1,其中G2、G3、G4这三种模式的土壤细菌丰富度超过了对照组。过熟林土壤细菌Simpson指数大小顺序为G3>G4>G1>G2>G6>G5,过熟林土壤细菌Shannon指数大小顺序为G5>G2>G6>G4>G1>G3,G5的土壤细菌Simpson指数和Shannon指数与对照组相比有所增加。
表1.不同植物功能群种植模式土壤微生物群落丰富度和多样性指数
2.4不同林龄土壤微生物群落组成及相对丰度影响分析
2.4.1不同植物功能群模式对土壤真菌群落组成及相对相度影响分析
对不同林龄杉木人工林土壤真菌的分类学分析显示丰度大于1%的真菌菌门共4种,包括子囊菌门(Ascomycota)、担子菌门(Basidiomycota)、接合菌门(Zygomycota)、以及罗兹菌门(Rozellomycota)。
幼龄林土壤真菌在门水平的群落结构组成按其所占比例大小依次为子囊菌门(Ascomycota)、担子菌门(Basidiomycota)、接合菌门(Zygomycota)、以及罗兹菌门(Rozellomycota)。在5种种植模式中占比最高的均为子囊菌门,所占比例基本都在40%以上,而对照组的担子菌门比例最高(45.85%),5个模式组的担子菌门比例相比对照组均有所下降。在Y3模式下土壤真菌在Rozellomycota占比为23.82%,而在其他模式以及对照组中的Rozellomycota在各自模式下所占比例均小于3%。
中龄林土壤真菌在门水平按所占比例大小依次为子囊菌门、担子菌门、接合菌门。在Z2、Z3、Z4、Z5模式下,土壤真菌中子囊菌门的占比最高,均超过了50%,接合菌门占比相对最低。在Z1模式和对照组中,占比最高的是接合菌门(37.15%,45.31%),担子菌门相对占比最低。
近熟林土壤真菌在门水平按所占比例大小依次为子囊菌门、担子菌门、接合菌门。JI和J2模式下土壤真菌在担子菌门占比最高,占比大小分别为42.61%和44.65%。J3、J4、J5模式以及对照组土壤真菌均在子囊菌门占比最高,所占比例大小均超过40%。各个模式下以及对照组的土壤真菌在Rozellomycota占比最低。
成熟林土壤真菌在门水平按所占比例大小依次为子囊菌门、担子菌门、接合菌门。C2模式土壤真菌在担子菌门占比最高(53.99%),C5模式土壤真菌在接合菌门占比最高(62.81%),其余各模式组以及对照组的土壤真菌在子囊菌门占比最高,尤其是C4模式和对照组,子囊菌门所占比例均超过70%。
过熟林土壤真菌在门水平按所占比例大小依次为担子菌门、子囊菌门、接合菌门。G1、G4、G5中担子菌门所占比例最高(52.78,45.47%,66.18%),G2、G3以及对照组中子囊菌门所占比例最高(59.72%,45.29%,41.64%)。不同种植模式以及对照组中的Rozellomycota占比最低。
5种不同模式的植入对于杉木人工林土壤中真菌的丰富度和多样性水平提高效果都较为明显。尤其是在幼龄林和近熟林中,不同种植模式下的土壤真菌丰富度和多样+性各个指数相比对照组均上。在中龄林Z4(黄檀+鸡血藤+红豆杉)、Z5(红豆杉+鸡血藤+山柰)模式的真菌Simpson指数低于对照组,除此之外各个种植模式的其他丰富度和多样性指数与对照组相比均有增加。在过熟林中不同种植模式对土壤真菌丰富度水平均有所提升,但只有G4(黄檀+鸡血藤+红豆杉)和G2(黄檀+山柰)模式的真菌多样性高于对照组。在成熟林经过不同模式植物的种植后未见土壤真菌群落丰富度和多样性水平有所提高。土壤真菌群落组成中,子囊菌门在过熟林中G5(红豆杉+鸡血藤+山柰)模式下的占比最高,在其他4个林龄中均在模式4(黄檀+鸡血藤+红豆杉)下占比最高。担子菌门在幼龄林中对照组下的占比最高,在其他4个林龄中均在模式2(黄檀+山柰)下占比最高。接合菌门在各个林龄中分别在Y2(黄檀+山柰)、Z6(对照)、J4(黄檀+鸡血藤+红豆杉)、C5(红豆杉+鸡血藤+山柰)、G3模式下占比最高。Rozellomycota菌门在幼龄林和近熟林的Y3(黄檀+鸡血藤)模式下占比最高,在成熟林和过熟林的C2(黄檀+山柰)和G4(黄檀+鸡血藤+红豆杉)模式下占比最高,中龄林中无Rozellomycota菌门。
2.4.2不同林龄土壤细菌群落组成及相对相度影响分析:
对不同林龄杉木人工林土壤细菌进行分类学分析,共得到丰度大于1%的细菌菌门共11种,分别是变形菌门(Proteobacteria)、酸杆菌门(Acidobacteria)、绿弯菌门(Chloroflexi)、放线菌门(Actinobacteria)、浮霉菌门(Planctomycetes)、厚壁菌门(Firmicutes)、疣微菌门(Verrucomicrobia)、芽孢杆菌门(Gemmatimonadetes)、拟杆菌门(Bacteroidetes)、螺旋体菌门(Saccharibacteria)以及蓝藻细菌门(Cyanobacteria)。
幼龄林各个种植模式下的变形菌门和酸杆菌门占据一定的优势,所占比例均大于20%,对照组土壤细菌中绿弯菌门占比最高(42.18%),变形菌门和酸杆菌门也占据一定的优势(17.30%,19.14%)。除Y3模式下的浮霉菌门(1.04%)外,绿弯菌门、放线菌门、浮霉菌门在各个模式下的占比都大于3%,其余门水平细菌占比基本上都低于3%。
中龄林中不同模式下的土壤细菌酸杆菌门、变形菌门和绿弯菌门都占有一定优势,Z1、Z3、Z5模式下的酸杆菌门占比最高,分别达32.57%、28.02%、28.88%。Z2模式下的变形菌门占比最高(36.13%),Z4和对照组在绿弯菌门占比最高,分别达32.00%和28.07%。各个模式下的放线菌门占比均大于7%,Z4和Z5模式的浮霉菌门、对照组的浮霉菌门和疣微菌门在各自模式下的占比均大于3%,其余门水平细菌占比均低于3%。
近熟林中不同模式下的土壤细菌酸杆菌门、变形菌门和绿弯菌门都占有一定优势,J2、J4、J5模式下的酸杆菌门占比最高,分别达32.24%、33.66%、34.88%。J3和对照组的变形菌门占比最高,分别达31.58%和29.05%。J1模式下绿弯菌门占比最高(29.08%)。各个模式下的放线菌门、J1、J2、J3、J5、J6模式下的厚壁菌门以及J2、J4、J5模式下的浮霉菌门在各自模式中的占比大于3%,其余门水平细菌占比均小于3%。
成熟林中不同模式下的土壤细菌酸杆菌门、变形菌门和绿弯菌门都占有一定优势,C3模式下的绿弯菌门占比最高(41.08%),除C3外其他模式下的变形菌门占比最高。所有模式中的放线菌门,C1、C3、C4以及对照组的浮霉菌门,C4和C6模式下的疣微菌门以及C4模式下的厚壁菌门,在各自模式下的占比均大于3%。
过熟林中不同模式下的土壤细菌变形菌门、酸杆菌门、绿弯菌门都占有一定优势,G1、G3、G4、G6模式下的变形菌门占比最高,所占比例分别为32.00%、53.54%、33.88%、27.57%。G2和G5模式下的酸杆菌门占比最高,分别为31.69%和28.49%。所有模式下的放线菌门,G1、G4、G5、G6模式下的浮霉菌门,G1、G4、G5、G6模式下的疣微菌门,G4和G5模式下的厚壁菌门,再各自模式下的占比均大于3%,其余门水平细菌占比小于3%。
5种模式对土壤细菌的影响不一致,在幼龄林各个模式的土壤细菌丰富度和多样性水平较幼龄林均有所提高。在中龄林Z5(红豆杉+鸡血藤+山柰)模式下的土壤细菌丰富度和多样性相比对照组均有所提高。在近熟林J3(黄檀+鸡血藤)模式最能够提高土壤细菌丰富度水平。在成熟林C2(黄檀+山柰)模式最能够提高土壤细菌丰富度水平。在过熟林G2(黄檀+山柰)模式最能够提高土壤细菌丰富度水平,G5(红豆杉+鸡血藤+山柰)模式最能够提高土壤细菌多样性水平。土壤细菌群落组成中,在各个林龄占比最高的4个门为变形菌门、酸杆菌门、绿弯菌门、放线菌门。其中变形菌门在不同林龄中的Y3、Z2、J3、C5、G3模式占比最高,酸杆菌门在不同林龄中的Y1、Z1、J5、C1、G2模式占比最高,绿弯菌门在不同林龄中的Y6、Z4、J2、C3、G6模式占比最高,放线菌门在不同林龄中的Y4、Z2、J1、C5、G3模式占比最高。
根据上述的不同植物功能群对土壤理化性质及微生物影响的相关结果,针对杉木林土壤氮磷亏缺严重、土壤微生物多样性降低、以及林下植被本身生物学特性(如黄檀、鸡血藤豆科植物固氮特性、红豆杉C4植物特性、山柰快速养分归还及其经济效益)等,综合分析可优选出以下较佳的针对不同林龄林下土壤生态功能提升的植物功能群配置:对于幼龄林和近熟林引入由南方红豆杉、鸡血藤和山柰组成的植物功能群A(图1中标号5对应的植物功能群配置),对于中龄林和过熟林引入由南岭黄檀和山柰组成的植物功能群B(图1中标号2对应的植物功能群配置),对于成熟林引入由南岭黄檀和南方红豆杉组成的植物功能群C(图1中标号1对应的植物功能群配置)。
以上所述仅为本发明的较佳实施例,并不用以限制本发明,凡在本发明的精神和原则之内,所作的任何修改、等同替换、改进等,均应包含在本发明的保护范围之内。
- 基于植物功能群构建的杉木林土壤生态服务功能提升方法
- 一种基于土壤速效水的农业干旱指数构建方法