PeLAP1基因在促进‘南林895杨’分枝中的应用
文献发布时间:2024-04-18 19:58:21
技术领域
本发明属于植物基因工程技术领域,更具体地说,涉及PeLAP1基因在促进‘南林895杨’分枝中的应用。
背景技术
‘南林895杨’是南京林业大学通过杂交育种培育出来的国家级良种(国S-SC-PE-002-2002),具有速生、优质、高产、高抗等优点。南京林业大学杨树课题组长期从事黑杨派树种遗传改良与栽培推广工作,在黑杨选、引、育等方面卓有成效。以南林895杨、南林3804杨、南林3412杨、南林862杨等为代表的一系列黑杨派良种,在我国黄淮、江淮及长江中下游平原地区表现出良好适应性和较高生产力,构成了江苏省森林资源的主体。第九次全国森林资源清查结果显示(《中国森林资源报告(2014-2018)》),江苏省森林面积155.99公顷,活立木蓄积量9609.62万立方米,森林蓄积(乔木林蓄积)7044.48万立方木;其中,全省杨树林面积达53.90万公顷、占乔木林面积(126.77万公顷)的42.52%,杨树林蓄积4484.98万立方米、占乔木林蓄积的63.67%。
随着现代遗传学理论与生物技术工具的不断发展,甄别、创制及应用种质的方法与途径也不断涌现。作为林木基础生物学研究的模式树种,近年来杨树功能基因组学研究突飞猛进,在杂种优势利用基础上,结合现代分子生物技术,全方位解析杨树生长发育及逆境适应性状的遗传调控机制,以黑杨良种为受体材料创建目标性状基因工程育种体系,将极大地推进杨树遗传改良进程、提高育种效率。目前基因工程技术已经广泛应用于杨树功能基因组学研究领域,但其受体材料多局限于银腺杨84K、欧美山杨T89、银灰杨717、毛白杨741等少数基因型。毫不疑问,这些受体材料是林木功能基因组学研究的较好模式系统,但其大田栽培表现差、适应性窄,不宜大面积造林推广。基于派内种间及派间远缘杂交策略选育出了的一系列速生优质黑杨新品种在我国杨树产业中占有重要地位,通过基因工程技术高效改良这些良种的株型性状,既可加速创制株型理想的杨树新优种质,又能引领林木良种改良进程。
发明内容
针对现有技术存在的上述问题,本发明所要解决的技术问题在于PeLAP1基因在促进‘南林895杨’分枝中的应用,用于遗传改良育种。
为了解决上述技术问题,本发明所采用的技术方案如下:
PeLAP1基因在促进‘南林895杨’分枝中的应用,所述PeLAP1基因的核苷酸序列如SEQ ID NO.1所示,包括以下步骤:
(1)构建PeLAP1基因的载体;
(2)将所构建的PeLAP1基因的载体转化到‘南林895杨’叶片中;
(3)培育筛选得到分枝增多的转基因‘南林895杨’植株。
所述的载体是植物表达载体。
所述的植物表达载体是35S::PeLAP1。
PeLAP1基因在促进‘南林895杨’生根时间提前中的应用,所述PeLAP1基因的核苷酸序列如SEQ ID NO.1所示。
PeLAP1基因在提高‘南林895杨’生根数量中的应用,所述PeLAP1基因的核苷酸序列如SEQ ID NO.1所示。
PeLAP1基因在降低‘南林895杨’株高中的应用,所述PeLAP1基因的核苷酸序列如SEQ ID NO.1所示。
PeLAP1基因在降低‘南林895杨’节间数量中的应用,所述PeLAP1基因的核苷酸序列如SEQ ID NO.1所示。
PeLAP1基因在提高‘南林895杨’体内分枝相关基因表达量中的应用,所述PeLAP1基因的核苷酸序列如SEQ ID NO.1所示。
所述的分枝相关基因为RAX2、RAX3。
相比于现有技术,本发明的有益效果为:
本发明以‘南林895杨’组培苗为材料,通过克隆得到PeLAP1基因,在此基础上构建植物表达载体35S::PeLAP1,转入‘南林895杨’叶片中,培育筛选得到分枝数量增多的‘南林895杨’PeLAP1突变体植株。转基因株系比CK生根时间提前了2-3天;生长30天的转基因株系生根数目显著多于CK,转基因植株高度显著低于CK;生长70天的转基因株系分枝数量显著多于CK,节间数量少于CK;转基因株系体内分枝相关基因RAX2的表达量是CK的1.8、4.7、4.6倍,RAX3基因的表达量为CK的3.2、4.0、3.8倍。
附图说明
图1为PeLAP1基因在各组织器官中的表达水平图;
图2为ABA处理后PeLAP1的表达量图;
图3为低温胁迫后PeLAP1的表达量图;
图4为35S::PeLAP1过表达载体构建的菌液检测图(a为大肠杆菌菌液检测结果,b为农杆菌菌液检测结果);
图5为转PeLAP1基因杨树分子检测图;
图6为PeLAP1在转基因株系中的表达水平图;
图7为转PeLAP1基因株系生根情况图;
图8为30天的转PeLAP1基因株系不定根数量统计图;
图9为微扦插10天的转PeLAP1基因株系表型图;
图10为30天的转PeLAP1基因株系苗高统计图;
图11为移栽后7天的转PeLAP1基因株系表型图;
图12为转PeLAP1基因株系分枝及节间数量统计图(a为过表达株系的分枝数量统计,b为过表达株系的节间数量统计);
图13为分枝相关基因在转PeLAP1基因株系中的表达量图。
具体实施方式
为使本发明的目的、技术方案和优点更加清楚,下面结合具体实施例对本发明进一步进行描述。以下实施例中如无特殊说明,所用的技术手段均为本领域技术人员所熟知的常规手段。
本发明所用植物材料为‘南林895杨’组培苗,在WPM生根培养基上生长,培养条件为16h光照,8h黑暗,湿度设定为70%。
Trans1-T1大肠杆菌感受态细胞、
EHA105农杆菌菌株购于擎科(北京)生物有限公司。
过量表达载体pH35GS保存于南京林业大学实验室。
RNAprepPure多糖多酚植物总RNA提取试剂盒、多糖多酚植物基因组DNA提取试剂盒购于天根生化科技(北京)有限公司;AxyPrep
实施例1
1、总RNA的提取
使用多糖多酚植物总RNA提取试剂盒提取30天‘南林895杨’组培苗的根、茎、叶的RNA,进行等量混合。实验步骤如下:
在1.5mLRNase-Free离心管中加入475μL裂解液SL和25μLβ-巯基乙醇;将植物组织加入高温消毒过的研钵中,并立即迅速研磨,将研磨成粉末状的植物组织加入含有裂解液的1.5mL离心管中,之后剧烈震荡1min;12000rpm室温离心2min;上清液转移至过滤柱CS,12000rpm室温离心2min后将上清液小心吸取至新的1.5mLRNase-Free离心管中;往离心管中加入300μL无水乙醇,用移液枪吸打混匀后转移至吸附柱CR3,12000rpm离心15s,弃废液,CR3重新放置在收集管中;加入350μL去蛋白液RW1,12000rpm离心15s,弃废液;从-20℃冰箱取出10μLDNaseI,加入70μLRDD,轻柔混匀;将DNaseI配置液加入CR3吸附柱的吸附膜中央,室温静置15min;加入350μL去蛋白液RW1,12000rpm离心15s,弃废液;加入500μL漂洗液PW,12000rpm离心15s,弃废液,CR3重新置于收集管中,洗涤两次;12000rpm离心2min,将CR3放入新的离心管中;向CR3吸附柱中的吸附膜中央滴加35μL的RNase-Free ddH
RNA质量检测:利用分光光度计测定总RNA的质量,其中OD260/280应该在1.8-2.1范围内;利用1%的琼脂糖凝胶电泳检测RNA的完整性。
2、cDNA合成
利用PrimeScriptRTMasterMix试剂盒将植物总RNA反转录为cDNA。
反应体系为:Prime Script RT Master Mix(5×)4μL、Total RNA1000ng XμL、RNase-FreeddH
反应程序为:37℃,15min;85℃,5s。反应结束后,根据具体需求对cDNA产物进行稀释,并放入4℃保存。
3、DNA提取
使用多糖多酚DNA提取试剂盒提取杨树基因组DNA,具体步骤见说明书。
4、引物设计
在JGI(https://phytozome-next.jgi.doe.gov/)网站下载毛果杨v4版本基因组数据,根据数据库中所提供的LAP1序列设计特异性扩增引物。使用Oligo6设计引物,长度控制在23-28nt,GC含量40%-70%,退火温度Tm≥60℃,引物序列如下所示:
PeLAP1-F:5’-ATGGGAAGAGGTAGGGTTCAGTT-3’,
PeLAP1-R:5’-TCATGTTCCAAAGCATCCAAGGT-3’。
5、目的片段扩增
以‘南林895杨’cDNA和DNA作为模板,使用高保真酶PrimeSTARMaxDNAPolymerase按照50μL体系对目的片段进行扩增。
反应体系为:
反应程序为:
使用2%琼脂糖凝胶电泳检测PCR扩增产物,回收单一且清晰的凝胶条带,用于回收纯化。
6、目的片段纯化与连接
使用凝胶回收试剂盒AxyPrep
将分离的琼脂糖胶块放入2mL离心管中并去皮称重,按照胶块重量:DE-A=1:3的比例向离心管中加入DE-A溶液,放置于75℃水浴锅中融化;加入1/2体积DE-A的DE-B溶液,充分混匀呈黄色溶液,吸取至DNA制备管中,12000rpm离心1min,倒出废液;加入500μLW1,12000rpm离心30s,倒废液;加入700μLW2,12000rpm离心30s,倒废液;加入700μLW2,12000rpm离心1min;将DNA制备管放入新的1.5mL离心管中,往吸附膜正中央垂直滴加20μLddH
回收后的PCR产物经过分光光度计检测浓度和质量后,连接到
反应程序为:25℃,15min。连接产物放4℃冰箱保存备用。
将5μL连接产物加入化冻的Trans1-T1大肠杆菌感受态菌株中,轻拨混匀,冰浴30min;置于42℃水浴锅中,热激40s后立刻放回冰盒中,冰浴4min;加入250μLLB液体培养基,在摇床上设置200rpm,37℃摇菌1h;将菌液从摇床取出,4000rpm离心5min;在超净工作台内吸除600μL上清液,剩余菌液用移液枪吹打混匀后滴在加带氨苄西林(Amp)抗性的LB固体培养基中,涂布棒均匀涂开;倒置于37℃恒温培养箱中,培养12h。
从平板中挑取大小合适的单菌落,每个菌落分别接种于携带Amp抗性的800μLLB液体培养基中,在温控摇床上设置250rpm,37℃摇菌4-6h。之后以菌液为模板进行PCR克隆检测。
反应体系如下;
反应程序为:
利用1.5%琼脂糖凝胶电泳对PCR产物的片段大小进行初步检测。选取条带位置与目的片段一致的对应阳性菌液送至生工公司进行测序。根据测序结果,确定目的基因的序列如SEQ ID NO.1所示,长度为684bp,命名为PeLAP1。其表达的蛋白序列如SEQ ID NO.2所示。
实施例2
1、按照实施例1中的方法使用多糖多酚植物总RNA提取试剂盒提取30天‘南林895杨’组培苗的根、茎、叶、茎尖、侧枝的RNA,并反转录为cDNA,具体方法如实施例1所示。采用实时荧光定量PCR(RT-qPCR)技术检测PeLAP1基因在不同器官中的特异性表达情况。引物序列如下所示:
qRTLAP1-F:5’-CGCCACGAGAGGTATTCTTATGC-3’,
qRTLAP1-R:5’-ACCTTTGCCTTGAGCCTGTTATAC-3’。
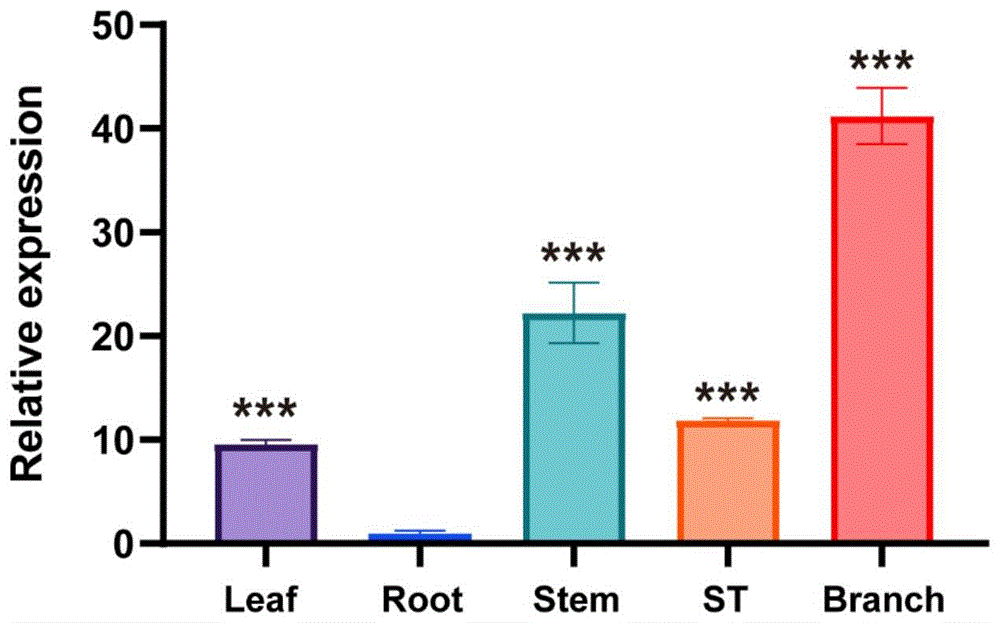
结果如图1所示,PeLAP1基因在根、茎、叶、茎尖、侧枝中均有表达,以表达量最低的根作为基准,并将它的表达水平设为1。其中,PeLAP1在侧枝中表达量最高,其相对表达量是根中的41.2倍;其次是在茎中,表达量是根中的22.2倍。PeLAP1基因在茎尖和叶中的水平次之,分别是根中11.8和9.5倍。根据上述结果得出结论,杨树侧枝中发挥作用。
2、使用浓度为100μmol/mL的脱落酸喷洒处理生长70天的‘南林895杨’盆栽苗,分别在0h、3h、6h、12h、24h取样,样品用液氮速冻并保存至-80℃超低温冰箱。按照实施例1中的方法分别提取总RNA后,以提取的总RNA为模板,通过反转录得到cDNA链。采用实时荧光定量PCR(RT-qPCR)技术检测脱落酸处理条件下PeLAP1基因的特异性表达情况,每个反应进行三次技术重复,以EF1a为内参基因,引物序列如下所示:
qRTLAP1-F:5’-CGCCACGAGAGGTATTCTTATGC-3’,
qRTLAP1-R:5’-ACCTTTGCCTTGAGCCTGTTATAC-3’;
EF1α-F:5’-GGCAAGGAGAAGGTACACAT-3’,
EF1α-R:5’-CAATCACACGCTTGTCAATA-3’。
qRT-PCR反应体系如下:
反应程序如下:
结果如图2所示,在经ABA处理的0-6h内,PeLAP1基因在杨树中的表达量并未显示变化,但在12h时显著升高;在处理12-24h内,PeLAP1的表达量又下降到与未处理时相同的水平,说明PeLAP1基因在一定程度内受脱落酸诱导。
3、在4℃低温下处理长势基本一致的‘南林895杨’组培苗,分别在0h、3h、6h、12h、24h取样,样品用液氮速冻并保存至-80℃超低温冰箱。
利用Beacon Designer 8软件设计实时定量引物,使用PowerUPTM SYBR GreenMaster mix试剂,在ABI ViiA
qRTLAP1-F:5’-CGCCACGAGAGGTATTCTTATGC-3’,
qRTLAP1-R:5’-ACCTTTGCCTTGAGCCTGTTATAC-3’;
EF1α-F:5’-GGCAAGGAGAAGGTACACAT-3’,
EF1α-R:5’-CAATCACACGCTTGTCAATA-3’。
qRT-PCR反应体系如下:
反应程序如下:
结果如图3所示,在4℃低温处理下,PeLAP1基因的表达量在0-6h内持续升高,6-12h时降低,12h之后又迅速升高,在24h达到最大值,说明PeLAP1基因在一定程度内参与植物的低温胁迫响应。
实施例3
1、过表达载体构建
利用One Step Cloning Kit试剂盒将PeLAP1的ORF序列构建到过表达载体pH35GS中,得到植物过表达载体35S::PeLAP1。具体的步骤如下:用灭菌过的接种环蘸取过表达载体pH35GS菌液,并在携带Spc抗性的LB固体培养基平板上划线;37℃培养箱中倒置约12h,挑取单克隆菌落并摇菌后进行菌液检测;将阳性菌液进行扩繁培养并提取质粒,对质粒纯度及浓度进行检验以备用;选用XhoI和KpnI-HF两种内切酶对pH35GS载体质粒进行线性化处理,反应体系如下:
反应程序为:37℃,60min,65℃,25min。
酶切产物用1%的琼脂糖凝胶进行电泳检测,之后使用AxyPrepTM DNA GelExtraction Kit回收纯化。
在目的片段的正反向扩增引物的5'端引入线性化载体两末端的同源序列,从而使扩增后获得带有和线性化克隆载体两末端对应一致的同源序列。添加完接头的引物分别为35S::PeLAP1-F和35S::PeLAP1-R,引物序列如下所示:
35S::PeLAP1-F:
5’-GGGGACTCTAGAATAGGTACCATGGGAAGAGGTAGGGTTCAGTT-3’,
35S::PeLAP1-R:
5’-CCTCAGCTACTTAAGCTCGAGTCATGTTCCAAAGCATCCAAGGT-3’。
以PeLAP1质粒为模板,利用35S::PeLAP1-F和35S::PeLAP1-R引物与高保真酶PrimeSTAR Max DNA Polymerase扩增目的片段,并用AxyPrepTM DNA Gel Extraction Kit试剂盒回收纯化扩增产物。
反应体系为:
反应程序为:
根据
反应程序为:37℃反应30min,结束后立即置于冰上;将重组质粒转入大肠杆菌感受态菌株Trans1-T1,具体方法参考实施例1,挑选单克隆进行菌液检测(图4a),最后经测序验证后获得插入方向及序列都正确的过表达重组载体阳性菌液,提取质粒以备用。剩余菌液保存至-80℃超低温备用。
2、转化农杆菌
将EHA105感受态细胞放置于冰盒中,待其融化后向其中加入200ng回收纯化的过量表达载体(质粒)混匀,冰浴30min;液氮速冻1min;37℃水浴3min,冰浴2min;加入800μLLB液体培养基,置于温控摇床上摇菌3h,设置为28℃,100rpm;3000rpm离心3min后吸除上清液;用移液枪吸打混匀菌液,用无菌涂布棒均匀涂抹于Kan+rif抗性的固体LB培养基上,并在28℃培养箱中倒置培养30-48h;在1.5mL离心管中加入800μL含Kan+rif抗性的LB液体培养基,挑取单菌落接种于离心管中;28℃,250rpm于温控摇床上摇菌24h;PCR检测菌液阳性,根据农杆菌菌液检测结果选择条带长度为950bp左右的阳性克隆菌液作为后续的遗传转化材料(图4b),得到的阳性菌液即作为遗传转化实验的目的种子液。
3、叶盘法转化杨树
取1mL目的种子液加入到50mL含Kan+rif抗性的液体LB培养基中,28℃,250rpm摇菌5-7h,直至OD600达到0.5-0.8;将菌液置于离心机中5000rpm,4℃离心10min,弃上清液;加50mL的液体无糖MS培养基重悬菌斑,混匀;将侵染液置于温控摇床上,28℃,90rpm激活1h;取生长状态优良的38-40d‘南林895杨’组培苗叶片,将叶片均匀剪出伤口,其中一部分作为空白对照,直接放入不加任何激素的分化培养基;剩余叶片放入侵染液中,28℃,90rpm温控摇床上侵染15-20min;取出叶盘,用灭菌滤纸吸干多余菌液,放进不含抗生素的分化培养基中,和正对照组一起暗培48h;用无糖MS液体培养基清洗三次叶盘,之后转入携带抑制农杆菌活性的Tim抗性分化培养基中光培7d;将叶盘更换至携带Kan+Tim抗性的分化培养基进行阳性植株筛选培养;待不定芽长出后,将叶盘移入携带Kan+Tim抗性的壮苗培养基中进行生长;待抗性不定芽生长至1cm左右时,将其剪下并转到携带Kan+Tim抗性的生根培养基中进行生长筛选;以上所有操作均在超净工作台中进行。
4、阳性植株筛选
提取转基因株系(T3、T4、T5、T6、T7、T8、T9、T16、T22、T23、T26、T29、T30、T31、T35)及对照组CK的DNA,以DNA为模板,利用P35S-F(5’-AGGAAGGTGGCTCCTACAAATGCCATC-3’)和35S::LAP1-R(5’-CCTCAGCTACTTAAGCTCGAGTCATGTTCCAAAGCATCCAAGGT-3’)进行PCR扩增,P35S-F在过表达载体上的位置距离目的片段260bp。
结果如图5所示,对照组CK泳道未显示扩增条带,其余泳道有单一明亮且与目的片段大小(950bp)相符的条带,初步筛选到11株阳性植株(T3、T5、T6、T8、T16、T22、T23、T26、T29、T31、T35)。
利用多糖多酚RNA提取试剂盒提取上述筛选出的11株阳性转基因杨树及对照组CK的RNA,凝胶电泳及分光光度计检测质量和浓度后将其反转录成cDNA。利用BeaconDesigner 8设计相关定量引物,以EF1a作为内参基因,CK和转基因株系的cDNA为模板进行qRT-PCR检测,测定PeLAP1以及分枝相关基因在转基因株系中的相对表达量,引物序列如下所示:
qRTPeLAP1-F:5’-CGCCACGAGAGGTATTCTTATGC-3’,
qRTPeLAP1-R:5’-ACCTTTGCCTTGAGCCTGTTATAC-3’;
EF1α-F:5’-GGCAAGGAGAAGGTACACAT-3’,
EF1α-R:5’-CAATCACACGCTTGTCAATA-3’。
结果如图6所示,选取表达量最高的4个PeLAP1-OE株系(OE#5对应T5、OE#16对应T16、OE#22对应T22、OE#31对应T31)进行后续表型观测实验。以对照组CK为基准,PeLAP1的相对表达量在PeLAP1-OE株系中增加了7-19倍左右,其中在OE#16中表达量最高,达到了18.6倍;在OE#5、OE#22、OE#31三个株系中分别为13.1、7.2、13.5倍。结果进一步证明35S::PeLAP1已经成功转入到杨树基因组中。
5、转基因株系的表型观测
1)选择6株生长状态基本一致的CK和4个转基因株系OE#5、OE#16#、OE#22、OE#31(每个株系5个重复,共计20株)在不含外源激素的WPM培养基中培养40天进行微扦插实验。观察统计生根时间、生根数量、植株高度等各方面的发育情况。
结果如图7所示,在扦插后的第7天,PeLAP1-OE转基因株系开始生根,比CK生根时间提前了2-3天;第12天时,PeLAP1-OE生根率已经达到100%,而第15天时CK的生根率才达到100%。
结果如表1、图8所示,对生长30天的CK及转基因株系的生根数量进行统计,结果表明,PeLAP1-OE转基因株系生根数目显著多于CK。
表1 PeLAP1-OE株系不定根数量统计
注:统计数据为三个独立重复的平均值±标准差(SD);*表示0.01 结果如图9所示,微扦插10天时,PeLAP1-OE株系与CK在表型上并未出现明显不同;10-20天时,PeLAP1-OE株系和CK生长较为缓慢;20-30天时,PeLAP1-OE株系和CK均迅速生长,但转基因植株高度显著低于CK。 结果如表2、图10所示,30天的4个PeLAP1-OE株系相对于CK在株高上表现出了不同程度的劣势,其中OE#16的变化最明显。 表2 PeLAP1-OE转基因株系的株高统计 注:统计数据为三个独立重复的平均值±标准差(SD);*表示0.01 2)将组培苗移栽至灭菌过的一定配比的营养土(营养土:珍珠岩:蛭石=7:2:1)中,每隔10天对植株的节间、株高和分枝情况进行统计。 结果如图11所示,在移栽后的第7天,即从微扦插算起第47天,过表达转基因株系已出现了分枝现象,CK未发现分枝。OE#5在第一个节点出现了1个分枝,OE#16在第二、三节点分别出现一个分枝,OE#31在第二个节点出现了1个分枝,而CK未发现分枝。 结果如表3、图12所示,对70天的CK和PeLAP1-OE株系进行分枝数量统计,发现PeLAP1-OE株系分枝数量显著多于CK,节间数量少于CK。过量表达PeLAP1基因在促进了分枝生长,抑制了节间数目以及株高生长。 表3 PeLAP1-OE转基因株系的分枝及节间数量统计 6、分枝相关基因在PeLAP1-OE株系中的表达水平 利用qRT-PCR检测调控杨树分枝发育的关键基因RAX2、RAX3、BRC1在PeLAP1-OE株系中的表达水平,引物序列如下所示: qRTRAX2-F:5’-GGCTCCTTGCTGTGACAA-3’, qRTRAX2-R:5’-TTCGCCATGTACTCCTTCAG-3’; qRTRAX3-F:5’-CAGCATGGCACTGGTGGTAACT-3’, qRTRAX3-R:5’-CCATCTAAGGCGGCAGCTCTTG-3’; qRTBRC1-F:5’-GGCAAGTGGAGTCCCTCTCTTCCC-3’, qRTBRC1-R:5’-TCCCATGTTTTGTACAAAGGATAC-3’; EF1α-F:5’-GGCAAGGAGAAGGTACACAT-3’, EF1α-R:5’-CAATCACACGCTTGTCAATA-3’。 结果如图13所示,RAX2基因在三个株系中的表达水平整体呈上升趋势,与对照组CK相比,表达量是CK的1.8、4.7、4.6倍;RAX3基因的表达情况与RAX2基因类似,在三个株系中的表达量为CK的3.2、4.0、3.8倍;而BRC1基因在3个株系中的表达量是CK的0.75、0.36、0.53倍。
- 南林895杨PdMYB2R032基因在提高植物抗旱性中的应用
- 一种“南林895杨”外源基因高效瞬时转化体系的建立方法