鸡ACE2基因的克隆、蛋白的表达和纯化及其多克隆抗体的制备
文献发布时间:2023-06-19 11:52:33
技术领域
本发明属于分子克隆技术领域,具体涉及一种鸡的新基因ACE2 (Angiotensinconverting enzyme 2)的克隆,蛋白的表达和纯化以及多克隆抗 体的制备。
背景技术
血管紧张素转换酶2(ACE2)是2000年发现的ACE同系物中的一员, 是肾素-血管紧张素系统的一个重要调节因子。属于I型跨膜糖蛋白,一种含 锌离子的单羧基肽糖蛋白酶。广泛存在于动物的心脏、肝脏、肺脏、肾脏、 睾丸以及肠道等主要器官中。ACE2将血管紧张素II(AngII)降解为具有舒 张血管及抗增殖作用血管紧张素(1-7)[Ang(1-7)],也可将血管紧张素I (AngI)降解为Ang(1-9)。Ang(1-7)通过其受体Mas(MasR)发挥抗 氧化、抗纤维化、抗炎等作用,引起了科研工作者的广泛关注。
目前ACE2的研究集中在以下三个方面:ACE2作为RAS系统的负调 节分子,通过靶向降解AngII,发挥抗损伤、抗纤维化等机体保护功能;作 为急性严重呼吸系统综合征冠状病毒(SARS-CoV)主要功能性受体,具有 保护肺损伤的功能;ACE2协同氨基酸转运载体转运,与氨基酸吸收不良、 肠道微生态平衡和胃肠道炎症发挥重要作用。
人、小鼠、大鼠、猫、绵羊、猪以及斑马鱼等多种哺乳动物及水生动 物,哺乳动物的ACE2基因鉴定并证实。但在鸟类中,ACE2基因鲜有报道, 尤其是家禽鸡的ACE2基因还未曾被注释。
在本发明研究中,根据NCBI公布的预测的鸡ACE2序列,首次通过 PCR扩增得到了鸡的ACE2基因,利用原核表达系统表达了pET32a-ACE2
重组蛋白,通过纯化pET32a-ACE2重组蛋白制备鼠抗ACE2血清多抗。 本发明研究填补了GenBank上缺乏鸡ACE2全长序列的资料空白,为ACE2 在肉鸡及禽类方面的研究提供了基础资料。
有鉴于此,特提出本发明。
发明内容
本发明的目的是根据ACE2作为RAS系统的负调节分子,通过靶向降 解AngII,发挥抗损伤、抗纤维化等机体保护功能;作为急性严重呼吸系统 综合征冠状病毒(SARS-CoV)主要功能性受体,具有保护肺损伤的功能; 协同氨基酸转运载体转运,与氨基酸吸收不良、肠道微生态平衡和胃肠道 炎症所发挥的关键作用,以及到目前为止鸡的ACE2基因还尚未被注释, 克隆得到鸡ACE2基因全长,不仅填补了GeneBank上缺乏鸡ACE2全长序 列的资料空白,而且通过原核表达ACE2蛋白并制备相应的多克隆抗体, 还可以为ACE2在肉鸡及禽类方面的研究提供基础资料。
为实现上述目的,本发明采用以下的技术方案为:
一种鸡的ACE2蛋白,其氨基酸序列如SEQ ID NO.1所示。
一种编码上述鸡的ACE2蛋白的基因,所述基因编码如SEQ ID NO.1 所示的氨基酸序列,或所述基因的核苷酸序列如SEQ ID NO.2所示。
一种鸡的ACE2的连接载体或表达载体,所述连接载体或表达载体含 有如SEQ IDNO.2所示的核苷酸序列。
一种鸡的ACE2克隆载体或表达载体的制备方法,其包括以下步骤:
S1、21日龄白羽肉鸡,颈部放血处死后取心脏、肝脏、肺脏、肾脏、 回肠、盲肠、空肠等,经生理盐水漂洗后迅速放入液氮速冻。提取基因组 RNA,使用随机引物将RNA反转录为cDNA。设计ACE2的上下游引物用 于ACE2全长基因扩增,并在ACE2基因片段两侧分别加入BamHI和HindIII 两个酶切位点,以cDNA为模板,扩增获得ACE2基因全长,扩增条件为: 95℃5min;95℃20s,59℃20s,72℃1min15s,40个循环;72℃10min。
S2、切胶纯化目的片段,取0.5uL pMD19-T载体,4.5uL纯化产物, 5uLSolutionI于200uLEP管中,混匀后在PCR仪器上运行16℃,14h,进行 连接。经菌液PCR、酶切、测序验证获得质粒pMD19T-ACE2。
如上所述的制备方法,优选地,还包括分别用BamHI和HindIII酶切 pET32a载体和pMD19T-ACE2质粒,切胶回收目的片段,用T4连接酶将 pET32a载体回收产物和pMD-19T-ACE2重组质粒回收产物进行连接,连接 体系(10uL);1uL pET32a载体,7uL目的序列,1uLT4连接酶,1uL10×Buffer 于200uLEP管中,混匀后在PCR仪器上运行22℃,14h,进行连接,经菌 液PCR、酶切、测序验证获得质粒pET32a-ACE2。
一种鸡的ACE2蛋白的制备方法,将上述鸡的ACE2原核表达载体, 转入DH5α、BL21感受态细胞中,经IPTG诱导表达后,分别获得 pET32a-ACE2重组蛋白。
如上所述的制备方法,质粒pET-32a-ACE2转入DH5α、BL21感受态 细胞后,诱导表达pET-32a-ACE2蛋白以包涵体的形式存在,纯化步骤:配 制10%分离胶,5%浓缩胶,不插梳子,待胶凝固后加入500uL的经上样缓 冲液处理过的包涵体提取液,另按常规方法配胶,加入标准分子量蛋白做 参考,按常规方法进行SDS-PAGE。
电泳毕,将卸下的胶放入一干净大平皿中,在摇床上进行0.25mol/LKCl 染色10min,染色完后,用手术刀片切下染成银白色的目的条带,用蒸馏水 洗5次后,将胶移入干净的小袋中,加入500uLPBS震荡混匀,-20℃反复 冻融3次,12000r/min,离心3min取上清SDS-PAGE电泳、Western blot 检测纯化效果。
一种鸡的ACE2蛋白多克隆抗体的制备,其包括如下步骤:将获得的 鸡pET32a-ACE2重组蛋白免疫Wistar大鼠,接种方式为颈背部皮下多点注 射,首免使用弗氏完全佐剂,分别于第二周进行一免,二免、三免和四免 使用弗氏不完全佐剂,于加强免疫后三天取大鼠血清。
本发明的有益效果在于:
本发明是首次扩增到了鸡的ACE2基因,不仅填补了GenBank上缺乏 鸡ACE2全长序列的资料空白,而且为ACE2在肉鸡及禽类方面的研究提 供了基础资料。
本发明中采用了大肠杆菌原核表达系统,构建了pET32a-ACE2质粒载 体,并进行了蛋白的表达和纯化,不仅能够制备效价较高的多克隆抗体, 而且还为之后ACE2相关功能的研究奠定了基础。
本发明中扩增得到的ACE2基因,目前ACE2的功能研究只要集中在 以下三个方面:ACE2作为RAS系统的负调节分子,通过靶向降解AngII, 发挥抗损伤、抗纤维化等机体保护功能;作为急性严重呼吸系统综合征冠 状病毒(SARS-CoV)主要功能性受体,具有保护肺损伤的功能;ACE2协同 氨基酸转运载体转运,与氨基酸吸收不良、肠道微生态平衡和胃肠道炎症 发挥重要作用。ACE2可能为鸡的肠道炎症治疗提供新的思路,为养禽业的 可持续发展奠定基础。
附图说明
为了更清楚地说明本发明具体实施方式或现有技术中的技术方案,下 面将对具体实施方式或现有技术描述中所需要使用的附图作简单地介绍, 显而易见地,下面描述中的附图是本发明的一些实施方式,对于本领域普 通技术人员来讲,在不付出创造性劳动的前提下,还可以根据这些附图获 得其他的附图。
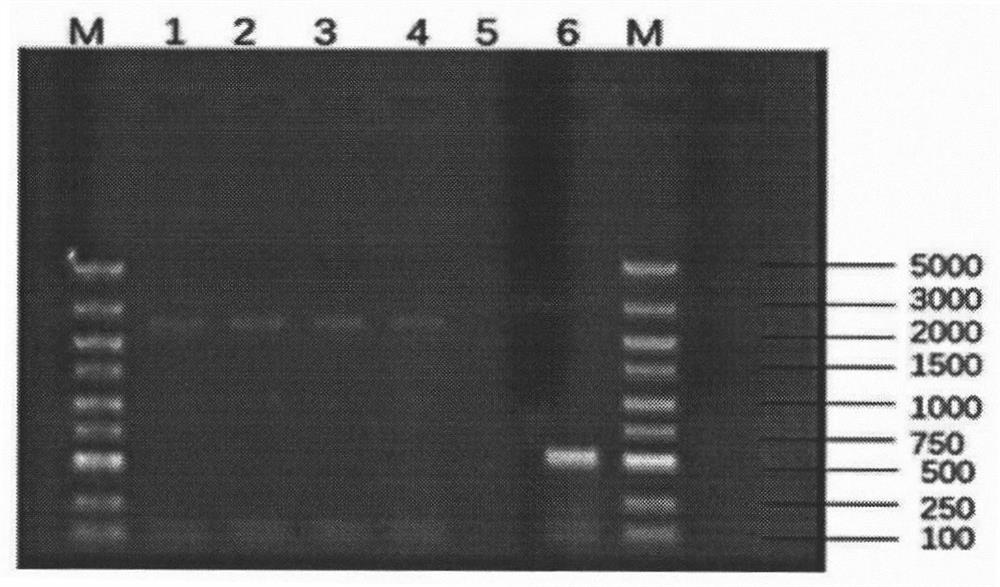
图1为本发明实例中利用RT-PCR方法扩增ACE2基因鉴定结果 (2444bp);其中,M:DNA Maker;泳道1-4:从白羽肉鸡空肠组织通过RT-PCR 扩增得到ACE2片段;泳道5:阴性对照;泳道6:18SrRNA基因;
图2A为菌液PCR扩增ACE2基因产物结果图;其中,M:DNA Maker 注:泳道1、2:pMD-19T-ACE2-阳性菌落泳道3:阴性对照;
图2B为pMD-19T-ACE2重组质粒单、双酶切鉴定结果图;其中,M: DNA Maker;泳道1、4:pMD-19T-ACE2重组质粒1;泳道2、5: pMD-19T-ACE2重组质粒1单酶切;泳道3、6:pMD-19T-ACE2重组质粒1 双酶切;
图3A为pET32a质粒双酶切结果图,其中,M:DL10000;泳道1-8: pET32a质粒;
图3B为pMD-19T-ACE2重组质粒双酶切结果图,其中,M:DL10000; 泳道1-7:pMD-19T-ACE2重组质粒;
图4A为DH5α感受态细胞的单克隆菌落结果图;
图4B为ACE2基因菌液PCR扩增产物结果图,其中,M:DL10000; 泳道1:pET32a-ACE2-阳性菌落1;泳道2:pET32a-ACE2-阳性菌落2;泳 道3:pET32a-ACE2-阳性菌落3;泳道4:pET32a-ACE2-阳性菌落4;泳道 5:pET32a-ACE2-阳性菌落5;泳道6:pET32a-ACE2-阳性菌落6;
图4C为pET32a-ACE2重组质粒单、双酶切鉴定结果图,其中,M: DL10000;1:pET32a-ACE2重组质粒1;2:pET32a-ACE2重组质粒1单酶切; 3:pET32a-ACE2重组质粒1双酶切;4:pET32a-ACE2重组质粒2;5: pET32a-ACE2重组质粒2单酶切;6:pET32a-ACE2重组质粒2双酶切;
图5A为不同浓度IPTG刺激pET32a-ACE2重组蛋白表达结果图,其 中,M:标准蛋白分子质量;泳道1-8:经IPTG终浓度为0、0.125、0.25、 0.5、1、2、4、8mmol/L诱导的pET32a-ACE2重组蛋白表达情况;
图5B为不同时间IPTG诱导的pET32a-ACE2重组蛋白表达结果图,其 中,M:标准蛋白分子质量;泳道1-9:经1mmol/L IPTG分别诱导0h、2h、 4h、6h、8h、10h、12h、过夜、过夜pET32a-ACE2重组蛋白表达情况;
图6为pET32a-ACE2重组蛋白表达形式的鉴定结果图,其中,M:标 准蛋白分子质量;泳道1、5:诱导后pET32a-ACE2重组菌表达产物;泳道 2、6:诱导后pET32a-ACE2重组菌超声破碎表达产物;泳道3、7:诱导后 pET32a-ACE2重组菌超声破碎上清;泳道4、8:诱导后pET32a-ACE2重 组菌超声破碎沉淀;
图7A为KCL切胶纯化pET32a-ACE2包涵体蛋白的SDS-PAGE结果 图,其中,M:标准蛋白分子质量;泳道1:诱导后pET32a-ACE2重组菌 表达产物;泳道2:诱导后pET32a-ACE2重组菌超声破碎表达产物;泳道 3:KCL切胶纯化pET32a-ACE2重组蛋白;泳道4:KCL切胶纯化pET32a-ACE2重组蛋白反复冻融3次;
图7B为KCL切胶纯化pET32a-ACE2包涵体蛋白的WB验证结果图, 其中,泳道1:未纯化pET32a-ACE2包涵体蛋白;泳道2:KCL切胶纯化 pET32a-ACE2包涵体蛋白;
图8为鼠抗鸡ACE2多克隆抗体效价检测的结果图;
图9为检测血清多抗的免疫印迹结果图,其中,A:阳性血清1-3: pET32a-ACE2重组蛋白;B:阴性血清1-3:pET32a-ACE2重组蛋白;
图10为本发明抗体检测鸡各组织ACE2蛋白表达结果图,其中,泳道 1:心脏;泳道2:肝脏;泳道3:肺脏;泳道4:肾脏;泳道5:空肠;泳 道6:回肠;泳道7:十二指肠;泳道8:盲肠;泳道9:直肠;
图11为本发明抗体检测不同物种组织ACE2蛋白表达结果图,其中, 泳道1:鸡回肠;泳道2:鸡十二指肠;泳道3:羊回肠;泳道4:羊十二 指肠;泳道5:大鼠回肠;泳道6:大鼠十二指肠;泳道7:猪回肠;泳道 8:猪十二指肠;
具体实施方式
下面将结合附图对本发明的技术方案进行清楚、完整地描述,显然, 所描述的实施例是本发明一部分实施例,而不是全部的实施例。基于本发 明中的实施例,本领域普通技术人员在没有做出创造性劳动前提下所获得 的所有其他实施例,都属于本发明保护的范围。
除非另有说明,本文中所用的专业与科学术语与本领域熟练人员所熟 悉的意义相同。此外,任何与所记载内容相似或均等的方法或材料也可应 用于本发明中。
下面结合具体实施例和对比例,对本发明作进一步说明。
实施例1 ACE2基因的扩增,以及连接载体的构建
引物设计根据GenBank中的预测的鸡ACE2基因序列 (XM-416822.5)设计,上下游引物分别插入BamHI和HindIII酶切位点由 南京擎科生物科技有限公司合成引物。将RNA反转录成cDNA,以cDNA 为模板进行PCR扩增,反应体系:cDNA模板2uL,反应条件:95℃ 5min;95℃ 20s,59℃ 20s,72℃ 1min15s,40个循环;72℃ 10min。PCR扩增结 果如图1,有单一条带出现(见泳道1-4),条带大小2000-3000bp之间,与 预期(2444bp)大小一致。
利用DNA胶回收试剂盒纯化回收PCR产物,具体步骤参照DNA胶 回收试剂盒试剂盒说明书进行。回收产物和pMD19T载体进行连接(16℃, 14h),将连接产物转化至感受态细胞DH5α中。将菌体重悬涂布在LB/Amp/X-Gal/IPTG培养基平板上,37℃培养16h。挑取培养皿上的单克隆 接种于含有氨苄的液体培养基中,培养过夜。利用质粒小型提取试剂盒提 取质粒,提取的质粒用HindIII单酶切及BamHI、HindIII双酶切(37℃, 14h)进行验证,将鉴定的阳性质粒送上海英俊生物技术有限公司测序。菌 落PCR验证结果如图2A,2000-3000bp之间有单一条带(图4,泳道1-2)。 PCR鉴定插入载体的片段大小与PCR扩增片段一致。提示阳性菌落中目的 片段插入成功。酶切验证结果如图2B所示:Hind III单酶切后发现有1条5136bp左右的条带(泳道2和泳道5),大小与pMD-19T载体(2692bp) 和目的基因(2444bp)总和一致。BamHI、Hind III双酶切可见2444bp和 2692bp左右两条带(泳道3和泳道6)。此结果证实已成功构建 pMD-19T-ACE2重组载体。测序获得的基因片段(SEQ ID NO.2):
编码氨基酸序列如下(SEQ ID NO.1):
引物序列如下:
ACE2-F(SEQ ID NO.3):CGGGATCCATGTTGCTTCACTTCTGGCTTCTCTGTG
ACE2-R(SEQ ID NO.4):GCGAAGCTTCTAAAAGGATGTTTGTGTCTCTTCAGATTGCT CA
实施例2 蛋白的表达实验
利用HindIII/BamHI双酶切经测序鉴定正确的pMD19T-ACE2重组质粒 和pET32a原核表达载体,pET32a质粒双酶切结果图如图3A,pET32a质 粒经BamHI、HindIII双酶切进行验证,酶切产物用1%琼脂糖凝胶电泳鉴 定,发现在5900bp处有单一目的条带,另一条条带约50bp,图上不显示。 证明酶切成功。pMD-19T-ACE2重组质粒双酶切结果如图3B, pMD-19T-ACE2重组质粒BamHI、HindIII双酶切可见2444bp和2692bp左 右两条带,证明酶切成功。
回收ACE2基因片段连接到pET32a原核表达载体,连接产物转化DH5α 感受态细胞,DH5α感受态细胞的单克隆菌落结果如图4A,白斑表明重组 质粒成功转入,形成了单克隆菌落。经菌液PCR及HindIII/BamHI双酶切 鉴定得到阳性重组表达载体pET32a-ACE2,ACE2基因菌液PCR扩增产物 结果如图4B,泳道1、2出现单一条带,条带大小与预期(2444bp)一致。pET32a-ACE2重组质粒单、双酶切鉴定结果如图4C,HindIII单酶切后发 现有1条约8344bp的条带,大小与pET32a载体(5900bp)和目的基因 (2444bp)总和一致(泳道2、5)。BamHI、HindIII双酶切可见5900bp和 2444bp左右两条带(泳道3、6)。此结果证实已成功构建pET32a-ACE2重 组载体。将质粒送上海英俊生物技术有限公司测序。
将测序鉴定正确的pET32a-ACE2重组质粒转化BL21(DE3)感受态细 胞,挑取单菌落,挑取白色菌落接种于20mLLB培养基,37℃震荡培养至 OD600为0.6-0.8,取1mL菌液作为诱导前对照,取1mL剩余菌液分别加 入20mg/mLIPTG至终浓度为8、4、2、1、0.5、0.25、0.125mmol/L。在28℃ 诱导6h后收集1mL菌液,8000rpm/min离心15min,收集菌液沉淀,100uLPBS重悬沉淀,经SDS-PAGE电泳确定IPTG最佳诱导浓度;挑取白 色菌落至加有20mLLB培养基,37℃震荡培养至OD600为0.6-0.8,取1mL 菌液作为诱导前对照,取1mL剩余菌液分别加入20mg/mLIPTG至终浓度 为1mmol/L。在28℃诱导2、4、6、8、10h后收集1mL菌液,8000rpm/min 离心15min,收集菌液沉淀,100uLPBS重悬沉淀,经SDS-PAGE电泳确定 IPTG最佳诱导时间。pET32a-ACE2重组蛋白诱导条件的优化结果如图5, IPTG确实可以诱导目的蛋白的表达,在IPTG终浓度为0.5、1、2、4mmol/L, 目的蛋白表达量较高,因此,筛选的IPTG诱导浓度为1mmol/L。随着诱导 时间的增加,重组蛋白表达量增加,但到诱导8h后,再增加诱导时间后, 重组蛋白表达量没有显著增加,因此确定诱导时间为10h。
选取表达量高的IPTG诱导浓度及时间,进行pET32a-ACE2重组蛋白 大量诱导表达,在28℃诱导10h后收集菌液,8000rpm/min离心15min,收 集菌液沉淀,PBS洗涤2次后,利用超声波裂解菌体,超声波裂解5s,间 隔10s,SDS-PAGE分析,鉴定目的蛋白的表达形式。pET32a-ACE2重组蛋 白表达形式的鉴定结果如图6可知,泳道4、8(沉淀)中目的蛋白含量显著高于泳道3、7(上清),即pET32a-ACE2重组蛋白主要以包涵体形式存 在。
实施例3 蛋白纯化实验
配制10%分离胶,5%浓缩胶,不插梳子,待胶凝固后加入500uL的经 上样缓冲液处理过的包涵体提取液,另按常规方法配胶,加入标准分子量 蛋白做参考,按常规方法进行SDS-PAGE。
电泳毕,将卸下的胶放入一干净大平皿中,在摇床上进行0.25mol/LKCl 染色10min,染色完后,用手术刀片切下染成银白色的目的条带,用蒸馏水 洗5次后,将胶移入干净的小袋中,加入500uLPBS震荡混匀,-20℃反复 冻融3次,12000r/min,离心3min取上清SDS-PAGE电泳、Westernblot检 验纯度。ACE2重组蛋白的Westernblot鉴定具体操作:将纯化的ACE2重 组蛋白和pET32a载体诱导表达菌经SDS-PAGE电泳后,采用湿转法将凝胶 上的相应条带转印至PVDF膜上,转印条件为恒流100V,90min。1∶3000 稀释的ACE2一抗,1∶5000稀释的辣根酶标记的山羊抗兔IgG。切胶纯化 重组蛋白鉴定结果如图7,相比于未纯化组经切胶纯化的组可见一条单一目 的条带(100kDa),且获得的纯化蛋白含量较高。
实施例4 ACE2多克隆抗体的制备
1、Wistar大鼠的免疫。
4只SPF级Wistar雌性大鼠(7周龄)购自扬州大学实验动物中心(实 验动物许可证:SCXK(苏)2017-0007),饲养于逸夫楼实验动物中心屏障 环境动物房(使用许可证:SYXK(苏)2017-0007)。大鼠,随机分为对照 组1只和实验组3只,分别称重后,断尾采血(作阴性对照),然后按如下程 序进行免疫。
首次免疫:将600uL的弗氏完全佐剂与准备好的600uL抗原(白羽肉 鸡pET32a-ACE2纯化重组蛋白,浓度为1402ug/mL)充分混匀使完全乳化。 取乳化好抗原液给老鼠皮下注射,实验组背腹部皮下多点注射,每个部位 300uL。对照组同样背腹部皮下多点注射1.2mL弗氏完全佐剂。二免、三免 和四免使用弗氏不完全佐剂,免疫周期为28天,共免疫4次。4次免疫后 的第7天,大鼠眼眶采血,收集血液置于37℃灭活2h后,4℃过夜使其凝 结释放血清,将凝结好的血液10000r/min,10min;收集上清,即为血清。
2、ELISA测定血清效价
使用间接ELISA方法测定血清效价。用纯化的pET32a-ACE2蛋白作为 包被抗原,抗原的包被浓度为403ug/mL,包被条件为4℃过夜,测定大鼠的 血清效价。结果如图8,与阴性对照相比,6只小鼠的血清效价均能达到 1∶6400。
实施例5 多克隆抗体的鉴定
1、纯化的重组蛋白Western Blot鉴定
将纯化的ACE2重组蛋白蛋白经SDS-PAGE凝胶电泳后电转至PVDF 膜上,转印条件为恒流100V,90min。用5%脱脂乳室温封闭2h,用PBST 洗涤3次;使用大鼠阳性(1∶5000),阴性(1∶5000)和ACE2单抗(1∶3000)作为 一抗,室温孵育2h,PBST洗涤三次,每次10min;辣根过氧化酶标记的山 羊抗鼠IgG作为二抗(1∶5000),常温孵育1h,PBST洗涤三次,每次10min,最后用ECL发光液曝光。
结果如图9所示,表明,免疫的大鼠血清可以特异性的与ACE2蛋白 进行结合(A),阴性的小鼠血清不能与ACE2蛋白进行结合(B)。
2、组织中ACE2蛋白的Western Blot鉴定
(1)白羽肉鸡各种组织中ACE2蛋白的Western Blot鉴定
采用两步法对白羽肉鸡心脏、肝脏、肺脏、肾脏、回肠、盲肠、空肠 组织蛋白进行提取,BCA试剂盒测定各组织蛋白浓度,将各组织蛋白稀释 至同一浓度后,-20℃保存。
将上步提取的组织总蛋白进行SDS-PAGE电泳后,浓缩胶5%,分离胶 10%。浓缩胶工作电压80V,分离胶工作电压120V,90min。采用湿转法将 凝胶上的相应条带转印至PVDF膜上,转印条件为恒流100V,90min。后 将PVDF膜置于50g/mL的脱脂奶粉中,室温封闭2h,然后转入用TBST 按1∶3000稀释的制备的ACE2多克隆抗,4℃温育过夜,TBST洗膜5次, 再转入用TBST液按1∶5000稀释的辣根酶标记的山羊抗兔IgG中,室温 孵育2h,TBST洗涤。显色后于全自动化学发光图像分析系统中观察拍照。
结果如图10所示,ACE2在心、肝、肾、空肠、回肠、十二指肠、盲 肠、直肠中表达,且心脏、回肠、直肠、空肠表达量较高。
(2)鸡、羊、猪及大鼠不同组织Western Blot鉴定抗体特异性
以制备的ACE2多克隆抗体为为一抗,羊抗鼠IgG-HRP作为二抗, 以鸡回肠、鸡十二指肠、羊回肠、羊十二指肠、大鼠回肠、大鼠十二指肠、 猪回肠、猪十二指肠蛋白进行Western Blot分析鉴定,结果如图11所示, 在鸡回肠、十二指肠在100kDa出现特异性的条带,而羊回肠、羊十二指肠、 大鼠回肠、大鼠十二指肠、猪回肠、猪十二指肠均未出现特异性条带。以 上结果表明,本发明制备的多克隆抗体能特异性识别鸡组织中的ACE2蛋 白,但与其他物种组织中的ACE2蛋白不能很好的识别,即制备的鸡抗鼠 的ACE2多克隆抗体具有较好的特异性。
最后应说明的是:以上各实施例仅用以说明本发明的技术方案,而非 对其限制;尽管参照前述各实施例对本发明进行了详细的说明,本领域的 普通技术人员应当理解:其依然可以对前述各实施例所记载的技术方案进 行修改,或者对其中部分或者全部技术特征进行等同替换;而这些修改或 者替换,并不使相应技术方案的本质脱离本发明各实施例技术方案的范围。
- 鸡ACE2基因的克隆、蛋白的表达和纯化及其多克隆抗体的制备
- 鸡CR2基因的克隆、蛋白的表达和纯化及其多克隆抗体的制备